Identification of Adult Fasciola spp. Using Matrix-Assisted Laser/Desorption Ionization Time-of-Flight (MALDI-TOF) Mass Spectrometry

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
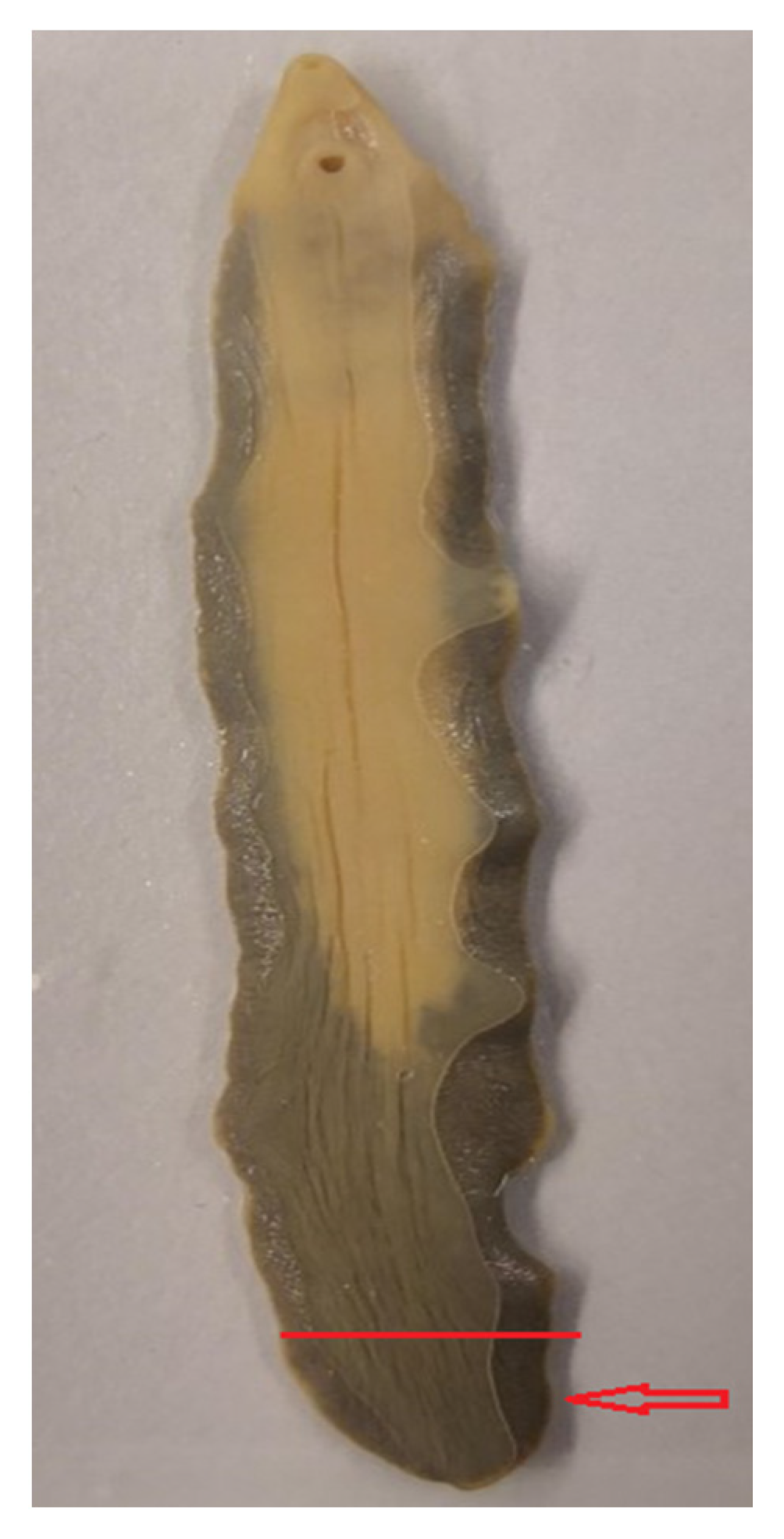
2.2. Sample Collection
2.3. Sample Preparation
2.4. Molecular Analysis
2.4.1. DNA Extraction, PCR and Sequencing
2.4.2. Sequence Analysis and Species Identification
2.5. MALDI-TOF Analysis
2.5.1. Protein Extraction
2.5.2. Target Plate Preparation and Measurements
2.5.3. MALDI-TOF MS Parameters
2.5.4. Spectral Analysis and Database Creation
2.5.5. Validation Test
3. Results
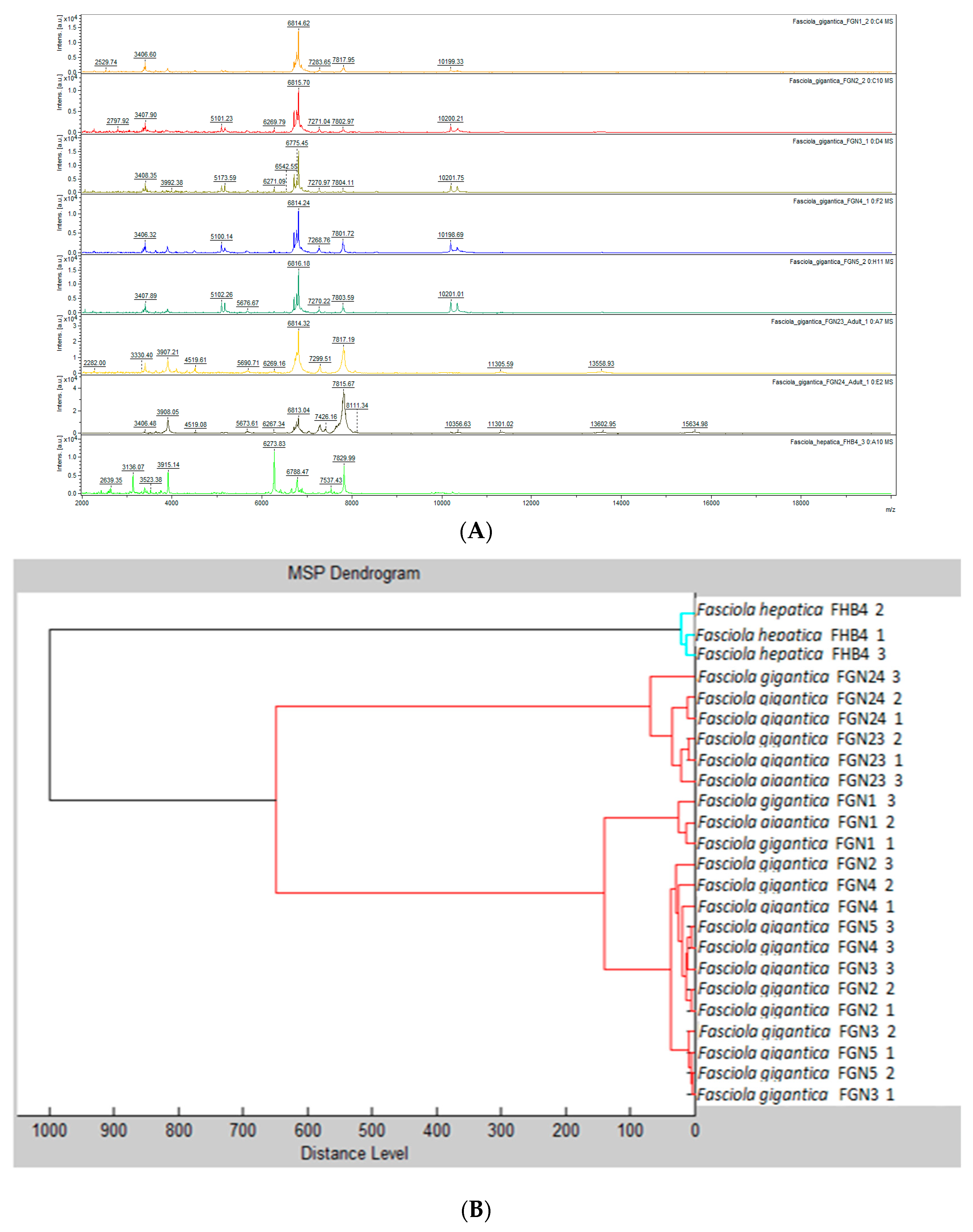
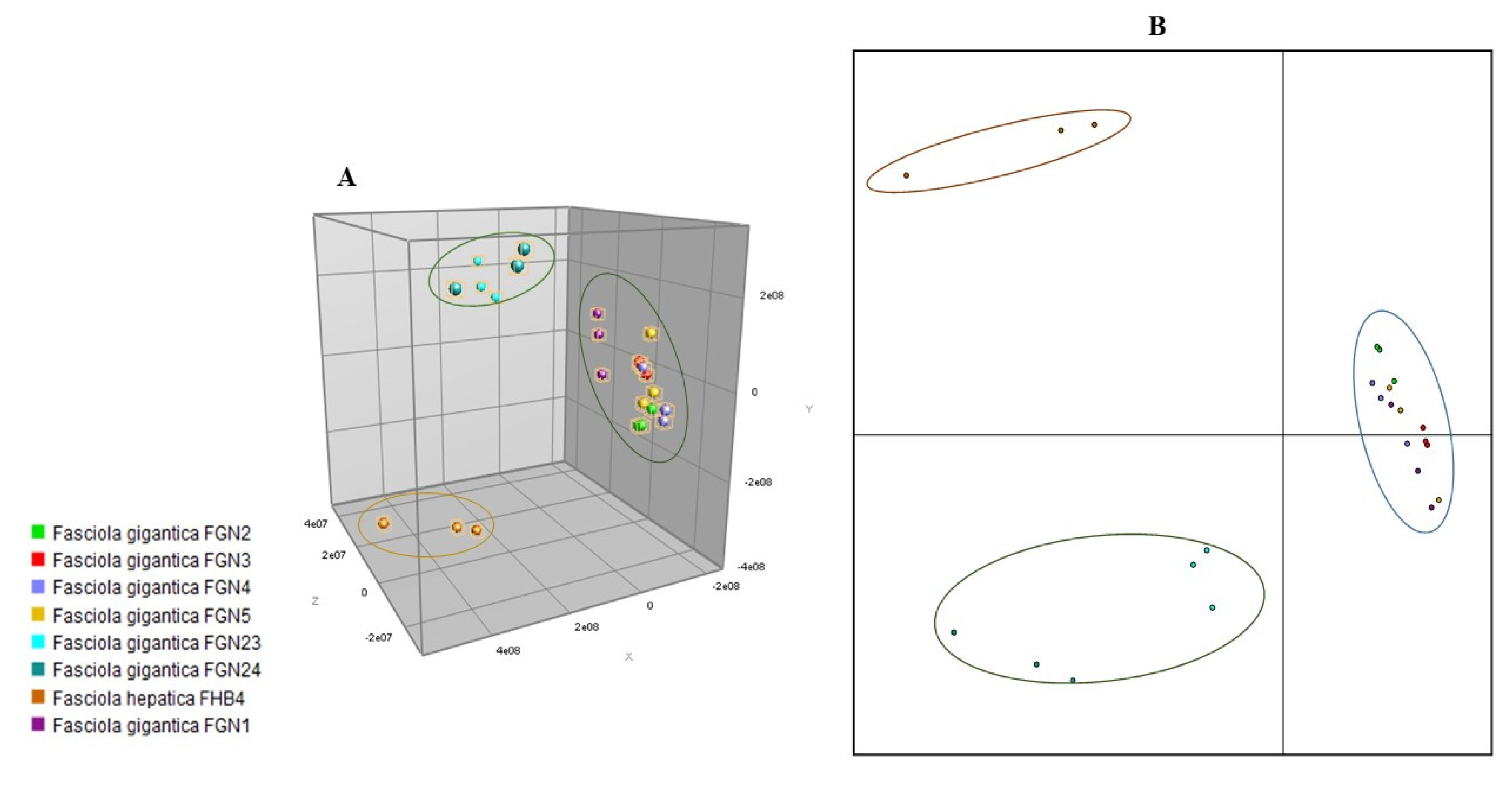
3.1. Comparative Analysis of Samples Used for MSP Database Creation
3.2. Internal Database Validation
3.3. Analysis of Samples for External Database Validation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Keiser, J.; Utzinger, J. Food-borne trematodiases. Clin. Microbiol. Rev. 2009, 22, 466–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amer, S.; ElKhatam, A.; Zidan, S.; Feng, Y.; Xiao, L. Identity of Fasciola spp. in sheep in Egypt. Parasit. Vectors 2016, 9, 623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvani, N.E.D.; Ichikawa-Seki, M.; Bush, R.D.; Khounsy, S.; Šlapeta, J. Which species is in the faeces at a time of global livestock movements: Single nucleotide polymorphism genotyping assays for the differentiation of Fasciola spp. Int. J. Parasitol. 2020, 50, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi-Storm, N.; Denwood, M.; Petersen, H.H.; Enemark, H.L.; Stensgaard, A.-S.; Sengupta, M.E.; Beesley, N.J.; Hodgkinson, J.; Williams, D.; Thamsborg, S.M. Patterns of Fasciola hepatica infection in Danish dairy cattle: Implications for on-farm control of the parasite based on different diagnostic methods. Parasit. Vectors 2018, 11, 674. [Google Scholar] [CrossRef]
- Perrodin, S.; Walti, L.; Gottstein, B.; Kim-Fuchs, C.; Candinas, D.; Banz, V. Fasciola hepatica in a country of low incidence: A tricky diagnosis. Hepatobiliary Surg. Nutr. 2019, 8, 597–603. [Google Scholar] [CrossRef]
- Mucheka, V.T.; Lamb, J.M.; Pfukenyi, D.M.; Mukaratirwa, S. DNA sequence analyses reveal co-occurrence of novel haplotypes of Fasciola gigantica with F. hepatica in South Africa and Zimbabwe. Vet. Parasitol. 2015, 214, 144–151. [Google Scholar] [CrossRef]
- Chougar, L.; Mas-Coma, S.; Artigas, P.; Harhoura, K.; Aissi, M.; Agramunt, V.H.; Bargues, M.D. Genetically “pure” Fasciola gigantica discovered in Algeria: DNA multimarker characterization, trans-Saharan introduction from a Sahel origin and spreading risk into north-western Maghreb countries. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Mas-Coma, S.; Valero, M.A.; Bargues, M.D. Fasciola, lymnaeids and human fascioliasis, with a global overview on disease transmission, epidemiology, evolutionary genetics, molecular epidemiology and control. Adv. Parasitol. 2009, 69, 41–146. [Google Scholar] [CrossRef]
- Jean-Richard, V.; Crump, L.; Abicho, A.A.; Naré, N.B.; Greter, H.; Hattendorf, J.; Schelling, E.; Zinsstag, J. Prevalence of Fasciola gigantica infection in slaughtered animals in south-eastern Lake Chad area in relation to husbandry practices and seasonal water levels. BMC Vet. Res. 2014, 10, 81. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.T.; Le, T.C.; Vo, M.D.C.; Van Cao, H.; Nguyen, L.T.; Ho, K.T.; Nguyen, Q.N.; Tran, V.Q.; Matsumoto, Y. High prevalence of cattle fascioliasis in coastal areas of Thua Thien Hue province, Vietnam. J. Vet. Med. Sci. 2017, 79, 1035–1042. [Google Scholar] [CrossRef] [Green Version]
- Chikowore, T.J.; Zishiri, O.T.; Mukaratirwa, S. Phylogenetic analysis of Fasciola spp. isolated from slaughtered cattle in KwaZulu-Natal and Mpumalanga provinces of South Africa based on the cytochrome c oxidase subunit I mitochondrial marker. Onderstepoort J. Vet. Res. 2019, 86, e1–e11. [Google Scholar] [CrossRef] [PubMed]
- Bargues, M.D.; Vigo, M.; Horak, P.; Dvorak, J.; Patzner, R.A.; Pointier, J.P.; Jackiewicz, M.; Meier-Brook, C.; Mas-Coma, S. European Lymnaeidae (Mollusca: Gastropoda), intermediate hosts of trematodiases, based on nuclear ribosomal DNA ITS-2 sequences. Infect. Genet. Evol. 2001, 1, 85–107. [Google Scholar] [CrossRef]
- Walker, S.M.; Makundi, A.E.; Namuba, F.V.; Kassuku, A.A.; Keyyu, J.; Hoey, E.M.; Prödohl, P.; Stothard, J.R.; Trudgett, A. The distribution of Fasciola hepatica and Fasciola gigantica within southern Tanzania--constraints associated with the intermediate host. Parasitology 2008, 135, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Pointier, J.-P.; Coustau, C.; Rondelaud, D.; Theron, A. Pseudosuccinea columella (Say 1817) (Gastropoda, Lymnaeidae), snail host of Fasciola hepatica: First record for France in the wild. Parasitol. Res. 2007, 101, 1389–1392. [Google Scholar] [CrossRef]
- Cwiklinski, K.; Dalton, J.P.; Dufresne, P.J.; La Course, J.; Williams, D.J.; Hodgkinson, J.; Paterson, S. The Fasciola hepatica genome: Gene duplication and polymorphism reveals adaptation to the host environment and the capacity for rapid evolution. Genome Biol. 2015, 16, 71. [Google Scholar] [CrossRef] [Green Version]
- Sumruayphol, S.; Siribat, P.; Dujardin, J.-P.; Dujardin, S.; Komalamisra, C.; Thaenkham, U. Fasciola gigantica, F. hepatica and Fasciola intermediate forms: Geometric morphometrics and an artificial neural network to help morphological identification. PeerJ 2020, 8, e8597. [Google Scholar] [CrossRef] [Green Version]
- Ai, L.; Chen, M.-X.; Alasaad, S.; Elsheikha, H.M.; Li, J.; Li, H.-L.; Lin, R.-Q.; Zou, F.-C.; Zhu, X.-Q.; Chen, J.-X. Genetic characterization, species differentiation and detection of Fasciola spp. by molecular approaches. Parasit. Vectors 2011, 4, 101. [Google Scholar] [CrossRef] [Green Version]
- Chabriere, E.; Bassène, H.; Drancourt, M.; Sokhna, C. MALDI-TOF MS and point of care are disruptive diagnostic tools in Africa. New Microbes New Infect 2018, 26, S83–S88. [Google Scholar] [CrossRef]
- Clark, A.E.; Kaleta, E.J.; Arora, A.; Wolk, D.M. Matrix-assisted laser desorption ionization-time of flight mass spectrometry: A fundamental shift in the routine practice of clinical microbiology. Clin. Microbiol. Rev. 2013, 26, 547–603. [Google Scholar] [CrossRef] [Green Version]
- Angeletti, S. Matrix assisted laser desorption time of flight mass spectrometry (MALDI-TOF MS) in clinical microbiology. J. Microbiol. Methods 2017, 138, 20–29. [Google Scholar] [CrossRef]
- Laroche, M.; Almeras, L.; Pecchi, E.; Bechah, Y.; Raoult, D.; Viola, A.; Parola, P. MALDI-TOF MS as an innovative tool for detection of Plasmodium parasites in Anopheles mosquitoes. Malar. J. 2017, 16, 5. [Google Scholar] [CrossRef] [PubMed]
- Nebbak, A.; Willcox, A.C.; Bitam, I.; Raoult, D.; Parola, P.; Almeras, L. Standardization of sample homogenization for mosquito identification using an innovative proteomic tool based on protein profiling. Proteomics 2016, 16, 3148–3160. [Google Scholar] [CrossRef] [PubMed]
- Diarra, A.Z.; Almeras, L.; Laroche, M.; Berenger, J.-M.; Koné, A.K.; Bocoum, Z.; Dabo, A.; Doumbo, O.; Raoult, D.; Parola, P. Molecular and MALDI-TOF identification of ticks and tick-associated bacteria in Mali. PLoS Negl. Trop. Dis. 2017, 11, e0005762. [Google Scholar] [CrossRef] [PubMed]
- Feucherolles, M.; Poppert, S.; Utzinger, J.; Becker, S.L. MALDI-TOF mass spectrometry as a diagnostic tool in human and veterinary helminthology: A systematic review. Parasit. Vectors 2019, 12, 245. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Proceedings of the Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; Volume 41, pp. 95–98. [Google Scholar]
- National Center for Biotechnology Information Nucleotide BLAST: Search Nucleotide Databases Using A Nucleotide Query. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastn&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome (accessed on 18 November 2020).
- Welker, M. Proteomics for routine identification of microorganisms. Proteomics 2011, 11, 3143–3153. [Google Scholar] [CrossRef]
- Shannon, S.; Kronemann, D.; Patel, R.; Schuetz, A.N. Routine use of MALDI-TOF MS for anaerobic bacterial identification in clinical microbiology. Anaerobe 2018, 54, 191–196. [Google Scholar] [CrossRef]
- Nebbak, A.; Almeras, L. Identification of Aedes mosquitoes by MALDI-TOF MS biotyping using protein signatures from larval and pupal exuviae. Parasit Vectors 2020, 13, 161. [Google Scholar] [CrossRef] [Green Version]
- Nebbak, A.; El Hamzaoui, B.; Berenger, J.-M.; Bitam, I.; Raoult, D.; Almeras, L.; Parola, P. Comparative analysis of storage conditions and homogenization methods for tick and flea species for identification by MALDI-TOF MS. Med. Vet. Entomol. 2017, 31, 438–448. [Google Scholar] [CrossRef]
- Ouarti, B.; Laroche, M.; Righi, S.; Meguini, M.N.; Benakhla, A.; Raoult, D.; Parola, P. Development of MALDI-TOF mass spectrometry for the identification of lice isolated from farm animals. Parasite 2020, 27, 28. [Google Scholar] [CrossRef]
- Karadjian, G.; Bilska-Zając, E.; Bahn, P.; Py, J.-S.; Johne, A.; Gassilloud, B.; Różycki, M.; Cencek, T.; Mayer-Scholl, A.; Vallée, I. Species identification of Trichinella originated from various host and different geographical location by MALDI-TOF. Exp. Parasitol. 2020, 213, 107890. [Google Scholar] [CrossRef]
- Marzano, V.; Pane, S.; Foglietta, G.; Levi Mortera, S.; Vernocchi, P.; Onetti Muda, A.; Putignani, L. Mass spectrometry-based proteomic analysis of Anisakis spp.: A preliminary study towards a new diagnostic tool. Genes 2020, 11, 693. [Google Scholar] [CrossRef] [PubMed]
- Sy, I.; Wendel, T.; Feucherolles, M.; Nimmesgern, A.; Stuermann, A.; Endriss, Y.; Utzinger, J.; Poppert, S.; Becker, S.L. Application of MALDI-TOF MS for identification of helminths in clinical samples. Abstract no. 6730. In Proceedings of the European Congress of Clinical Microbiology and Infectious Diseases, Paris, France, 18–21 April 2020; Cancelled Due to the COVID-19 Pandemic. p. 3232. [Google Scholar]
- Nguyen, T.B.N.; De, N.V.; Nguyen, T.K.L.; Quang, H.H.; Doan, H.T.T.; Agatsuma, T.; Le, T.H. Distribution status of hybrid types in large liver flukes, Fasciola species (Digenea: Fasciolidae), from ruminants and humans in Vietnam. Korean J. Parasitol. 2018, 56, 453–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.-H.; Gasser, R.B.; Young, N.D.; Song, H.-Q.; Ai, L.; Zhu, X.-Q. Complete mitochondrial genomes of the “intermediate form” of Fasciola and Fasciola gigantica, and their comparison with F. hepatica. Parasit. Vectors 2014, 7, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evack, J.G.; Schmidt, R.S.; Boltryk, S.D.; Voss, T.S.; Batil, A.A.; Ngandolo, B.N.; Greter, H.; Utzinger, J.; Zinsstag, J.; Balmer, O. Molecular confirmation of a Fasciola gigantica × Fasciola hepatica hybrid in a Chadian bovine. J. Parasitol. 2020, 106, 316–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Species | Number of Samples | Identification | LSV Range | |
|---|---|---|---|---|
| LSV ≥ 1.70 | LSV ≥ 2.00 | |||
| Fasciola gigantica | 75 | 74 (98.7%) | 31 (41.3%) | 1.73–2.13 |
| Fasciola hepatica | 3 | 3 (100%) | 3 (100%) | 2.15–2.23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sy, I.; Margardt, L.; Ngbede, E.O.; Adah, M.I.; Yusuf, S.T.; Keiser, J.; Rehner, J.; Utzinger, J.; Poppert, S.; Becker, S.L. Identification of Adult Fasciola spp. Using Matrix-Assisted Laser/Desorption Ionization Time-of-Flight (MALDI-TOF) Mass Spectrometry. Microorganisms 2021, 9, 82. https://doi.org/10.3390/microorganisms9010082
Sy I, Margardt L, Ngbede EO, Adah MI, Yusuf ST, Keiser J, Rehner J, Utzinger J, Poppert S, Becker SL. Identification of Adult Fasciola spp. Using Matrix-Assisted Laser/Desorption Ionization Time-of-Flight (MALDI-TOF) Mass Spectrometry. Microorganisms. 2021; 9(1):82. https://doi.org/10.3390/microorganisms9010082
Chicago/Turabian StyleSy, Issa, Lena Margardt, Emmanuel O. Ngbede, Mohammed I. Adah, Saheed T. Yusuf, Jennifer Keiser, Jacqueline Rehner, Jürg Utzinger, Sven Poppert, and Sören L. Becker. 2021. "Identification of Adult Fasciola spp. Using Matrix-Assisted Laser/Desorption Ionization Time-of-Flight (MALDI-TOF) Mass Spectrometry" Microorganisms 9, no. 1: 82. https://doi.org/10.3390/microorganisms9010082