Differences in Microbial Community and Metabolites in Litter Layer of Plantation and Original Korean Pine Forests in North Temperate Zone


Abstract
:1. Introduction
2. Materials and Methods
2.1. Natural Survey of Study Area
2.2. Sample Collection
2.3. Methods
2.3.1. Extraction and Sequencing of Total DNA from Litter Layer
2.3.2. Determination of Untargeted Metabolites in Litter Layer
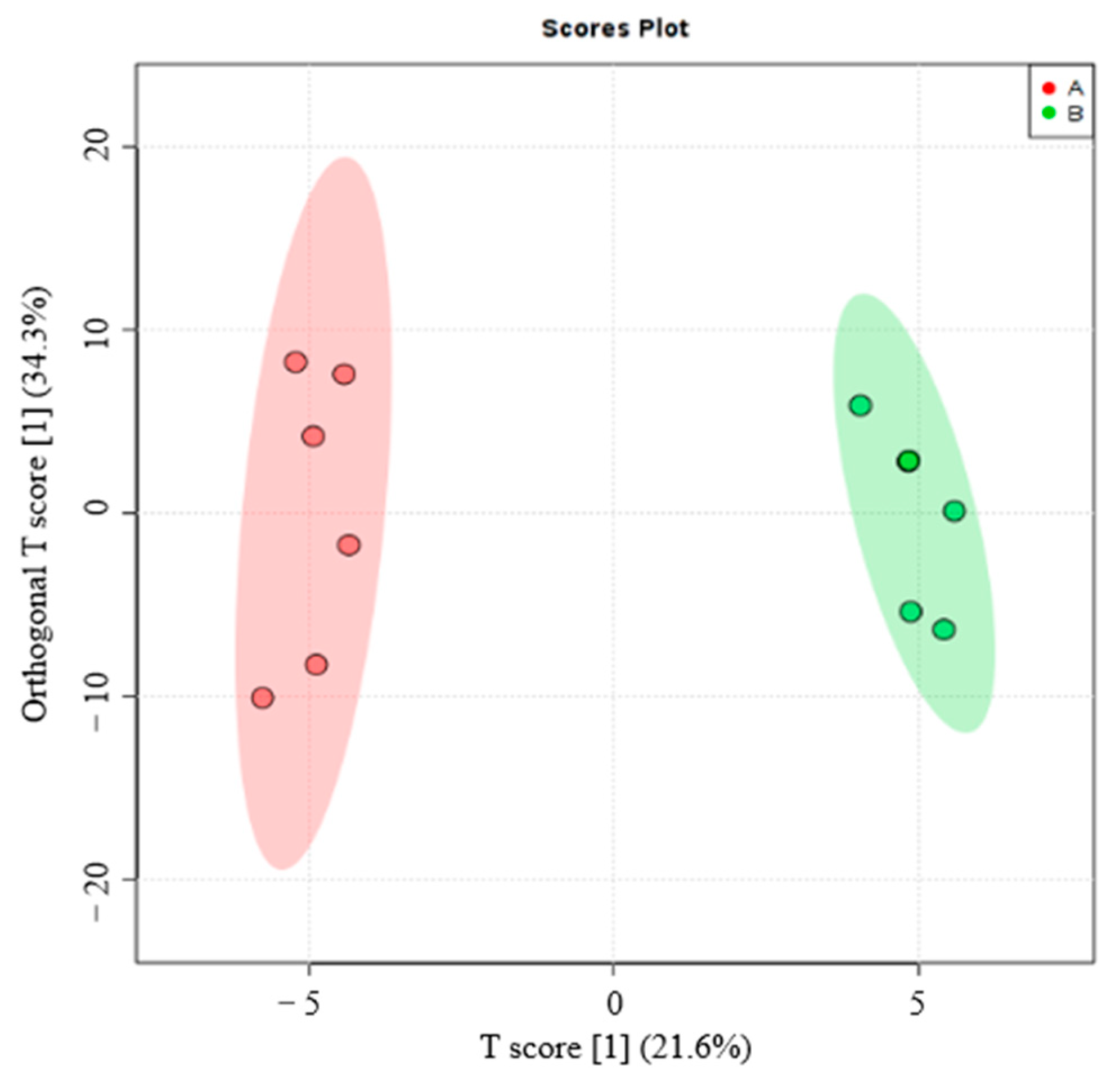
2.4. Data Statistics
3. Results and Analysis
3.1. Bacterial Community Structure of Litter Layer in Korean Pine Plantation and Original Korean Pine Forest
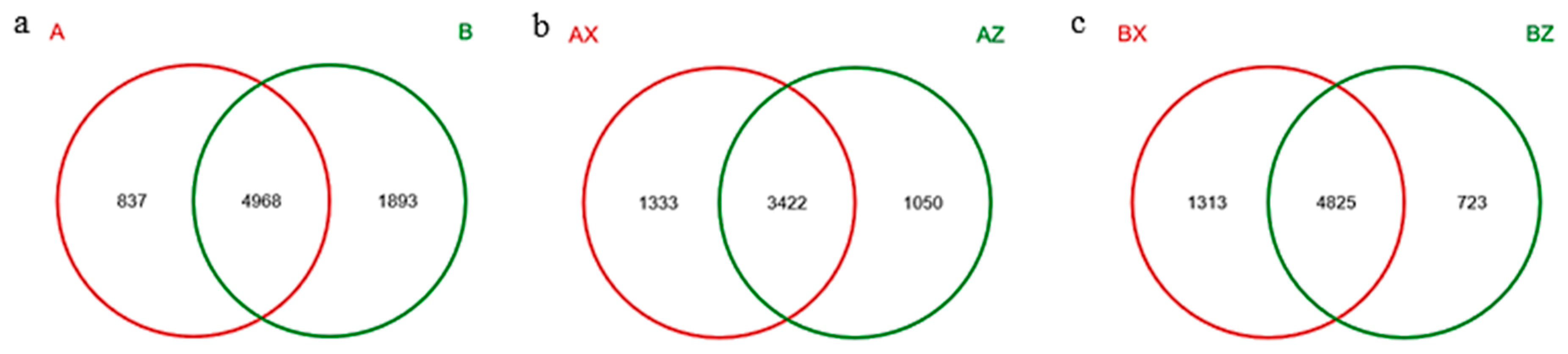
3.1.1. Results of Illumina-Miseq Sequencing of Litter-Layer Bacteria
3.1.2. α-Diversity of Bacterial Community in Litter Layer
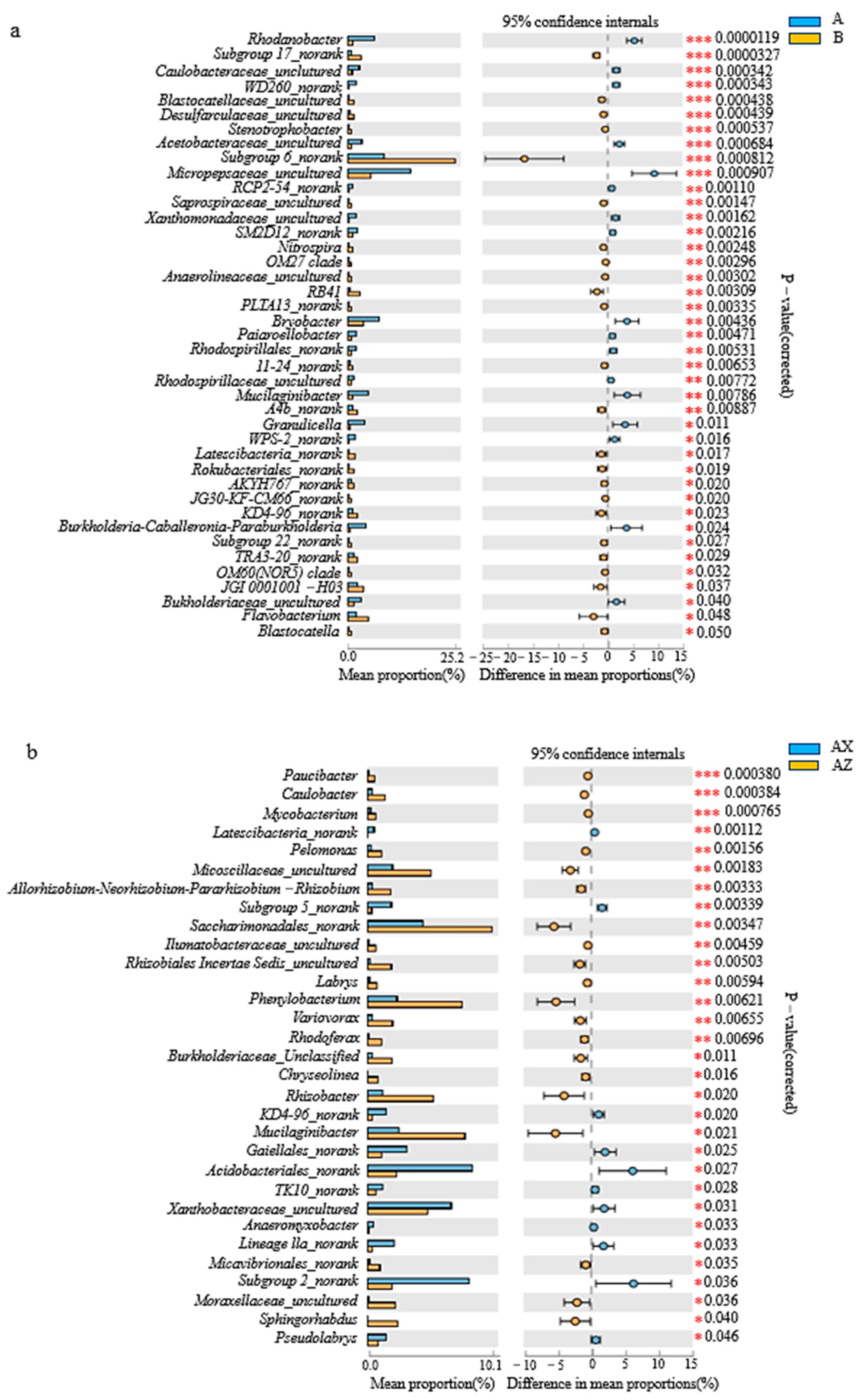
3.1.3. Bacterial Community Composition in Litter Layer of the Two Forests
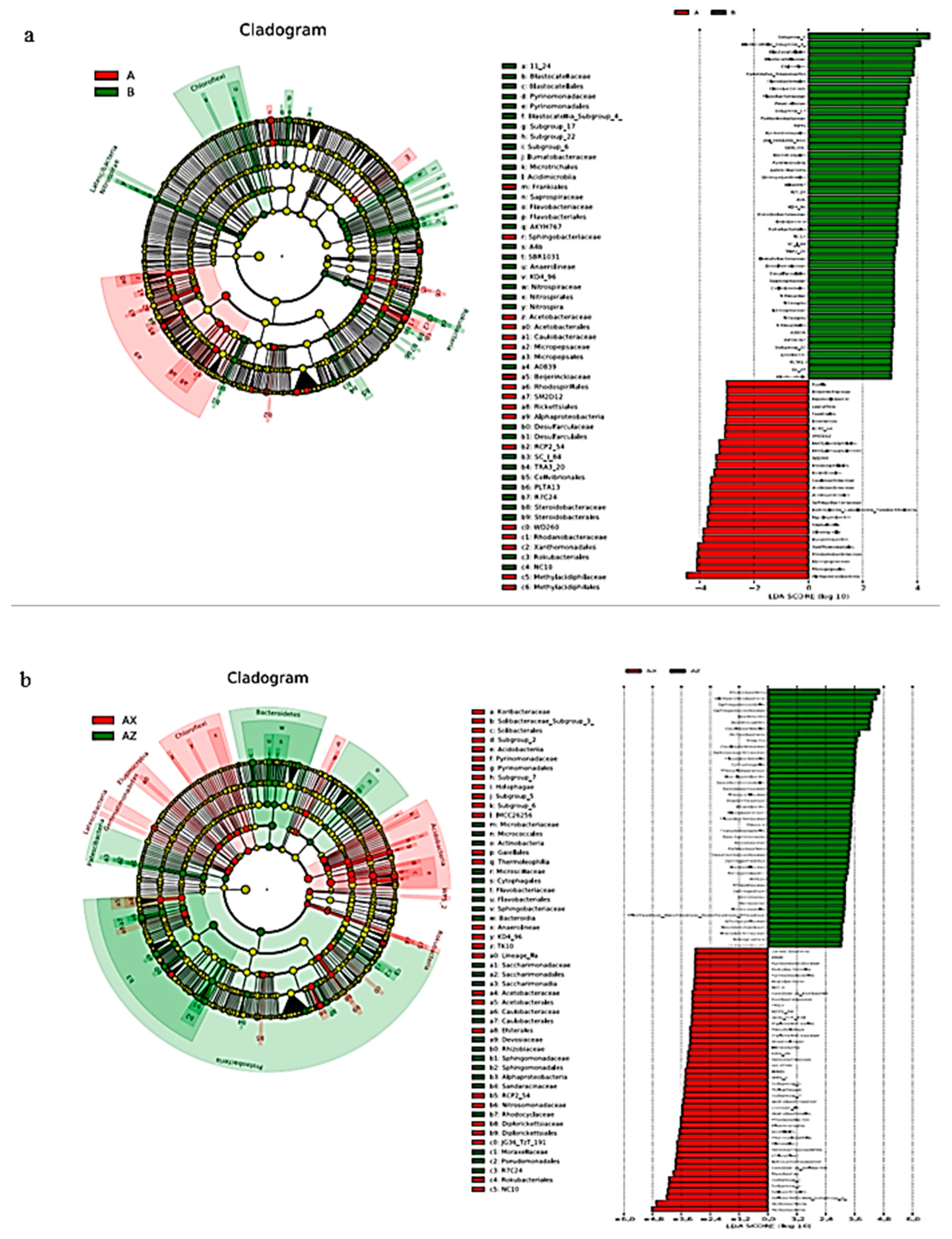
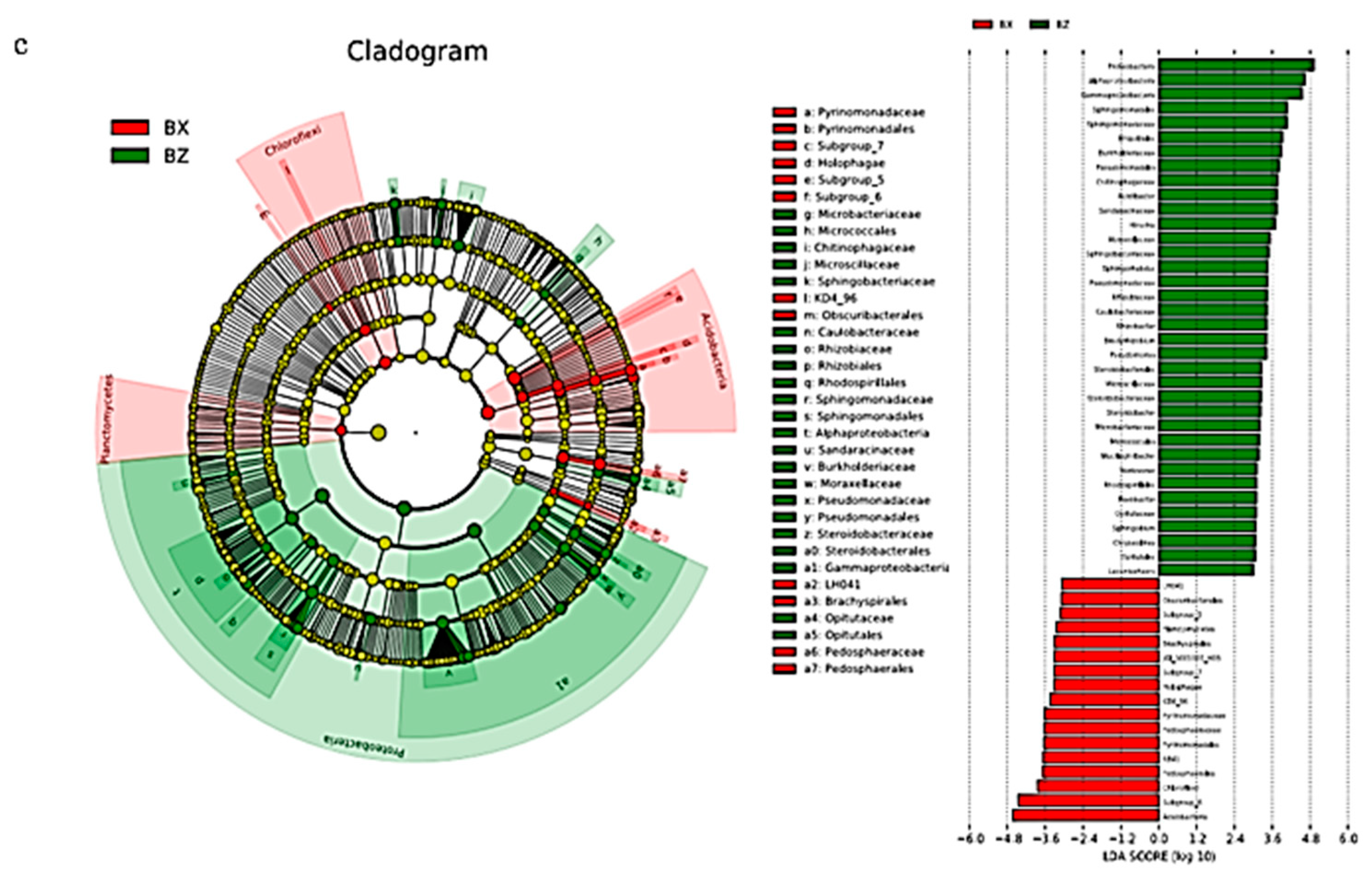
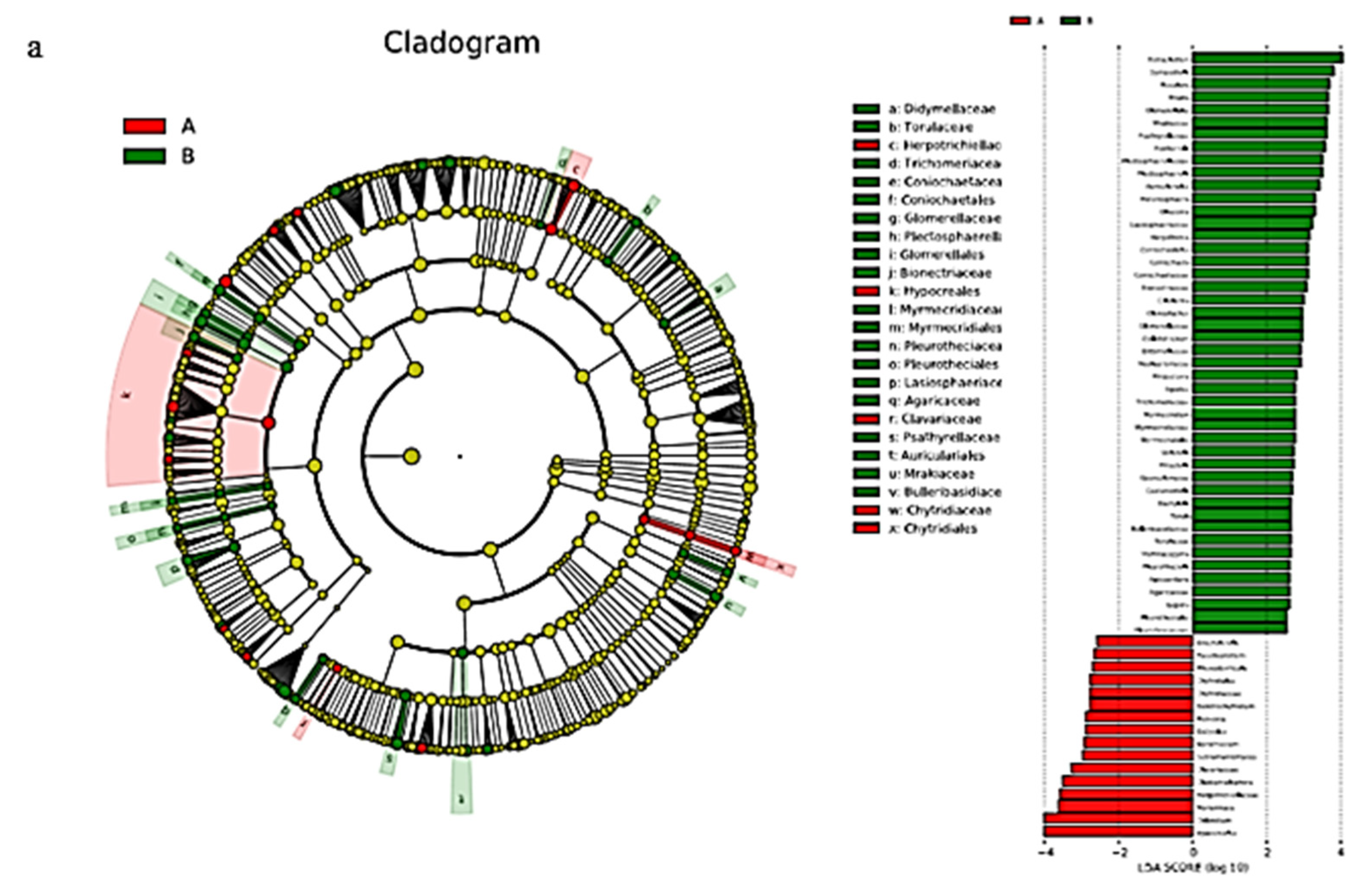
3.1.4. Bacterial Linear Discriminant Effect Size (LEfSe) Litter Analysis
3.2. Community Structure of Litter-Layer Fungi in Plantation and Original Korean Pine Forests
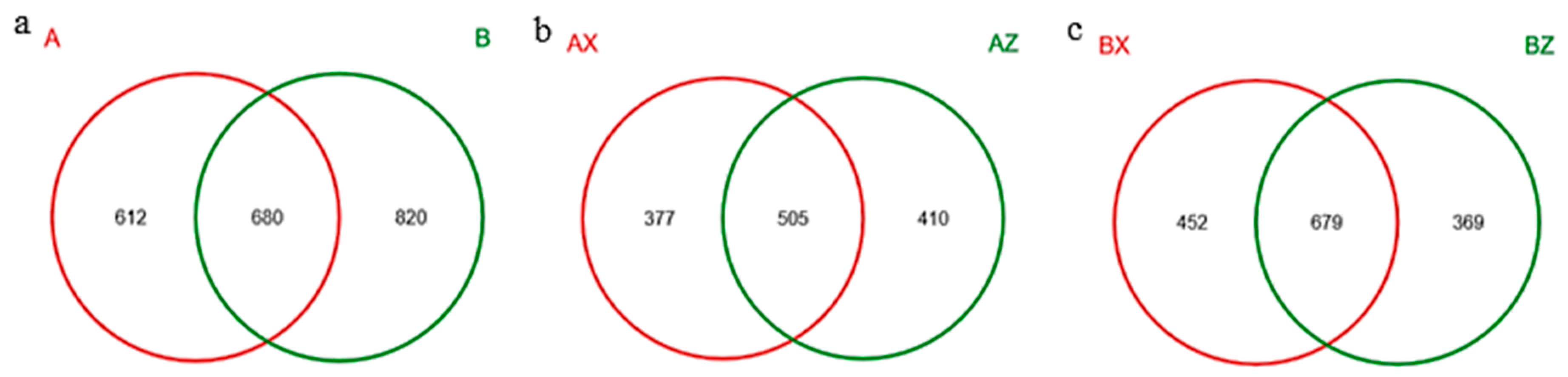
3.2.1. Results of Illumina-Miseq Sequencing of Litter-Layer Fungi
3.2.2. α-Diversity of Litter-Layer Fungus Communities
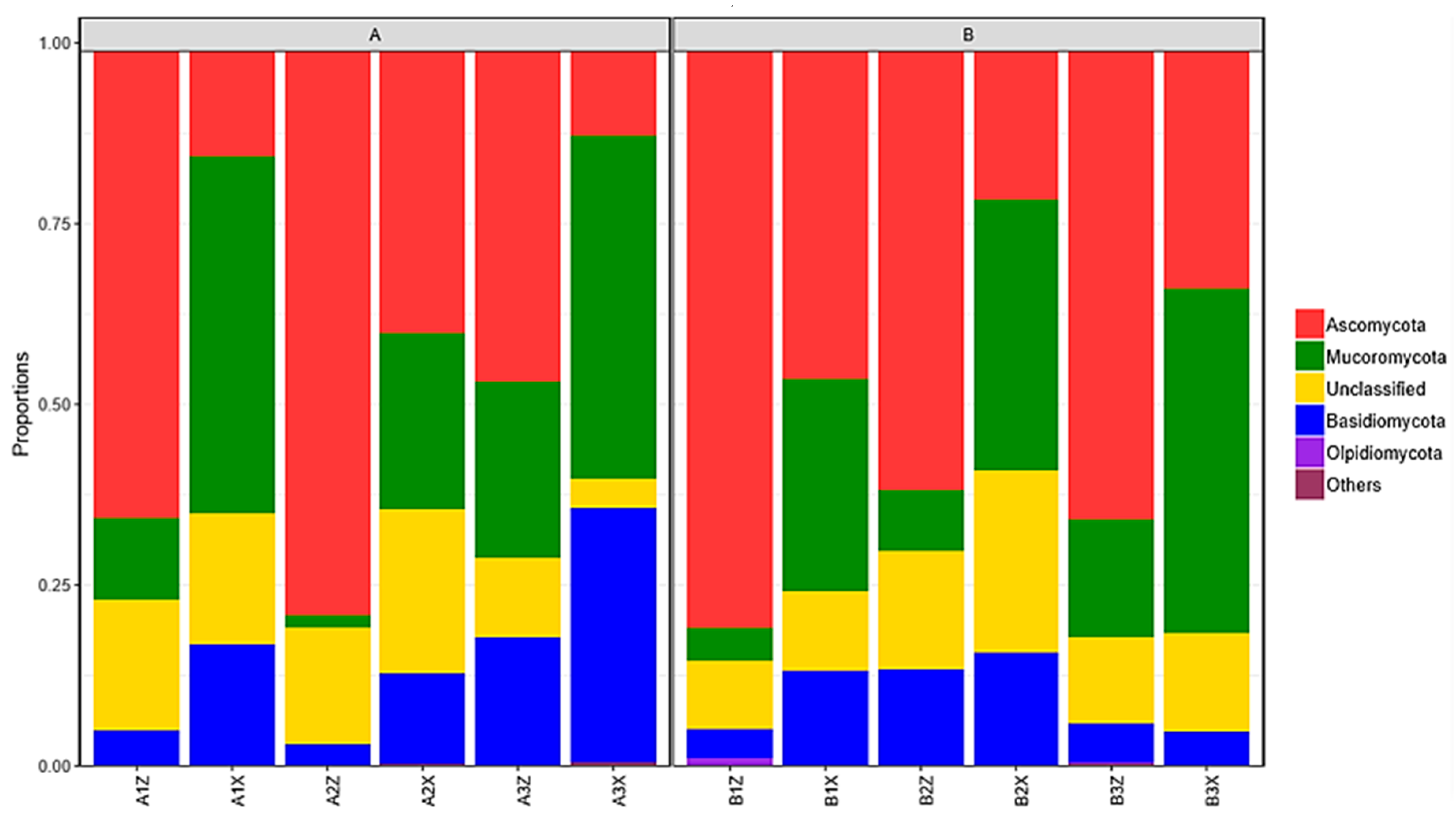
3.2.3. Composition of Litter-Layer Fungal Communities
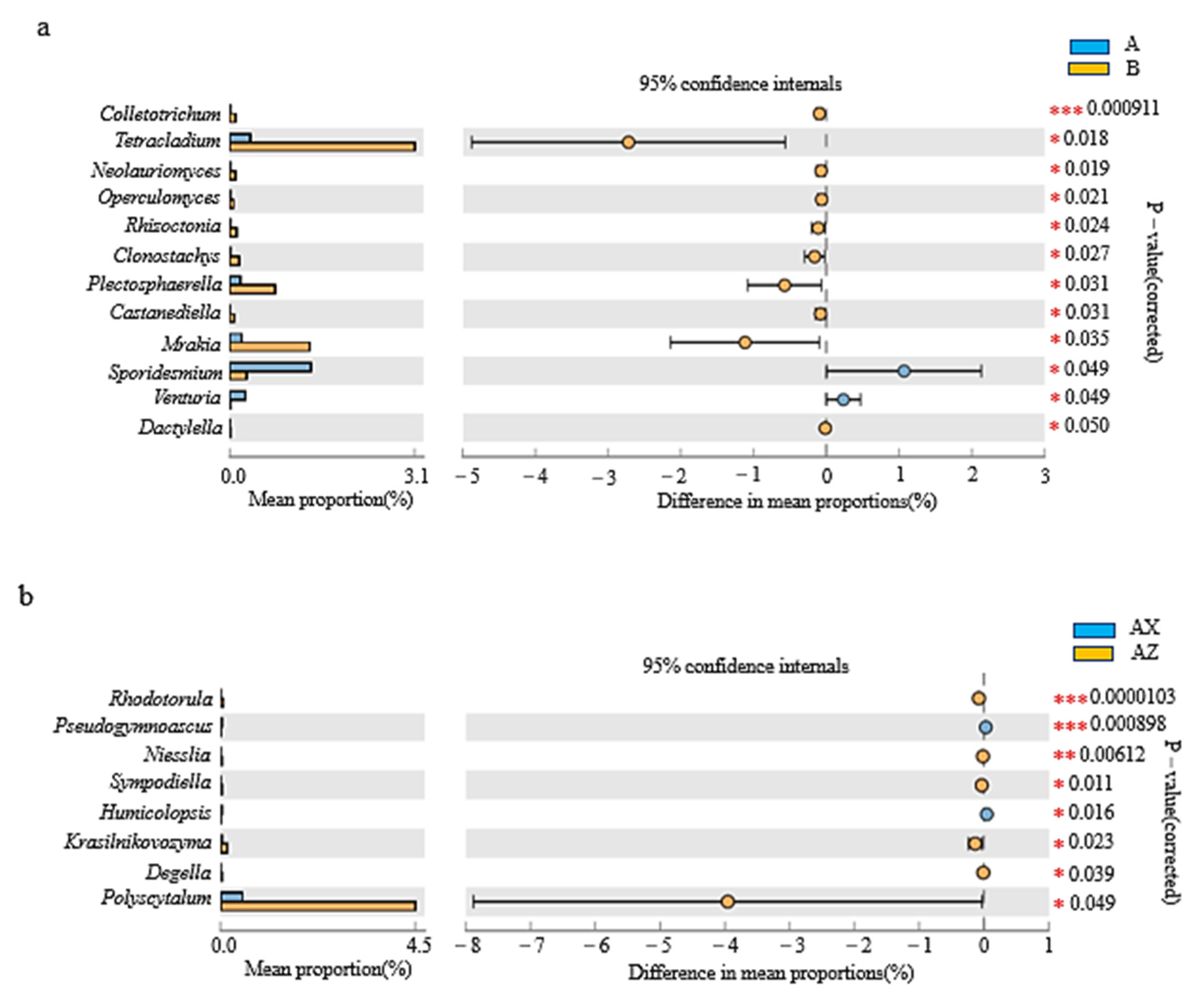
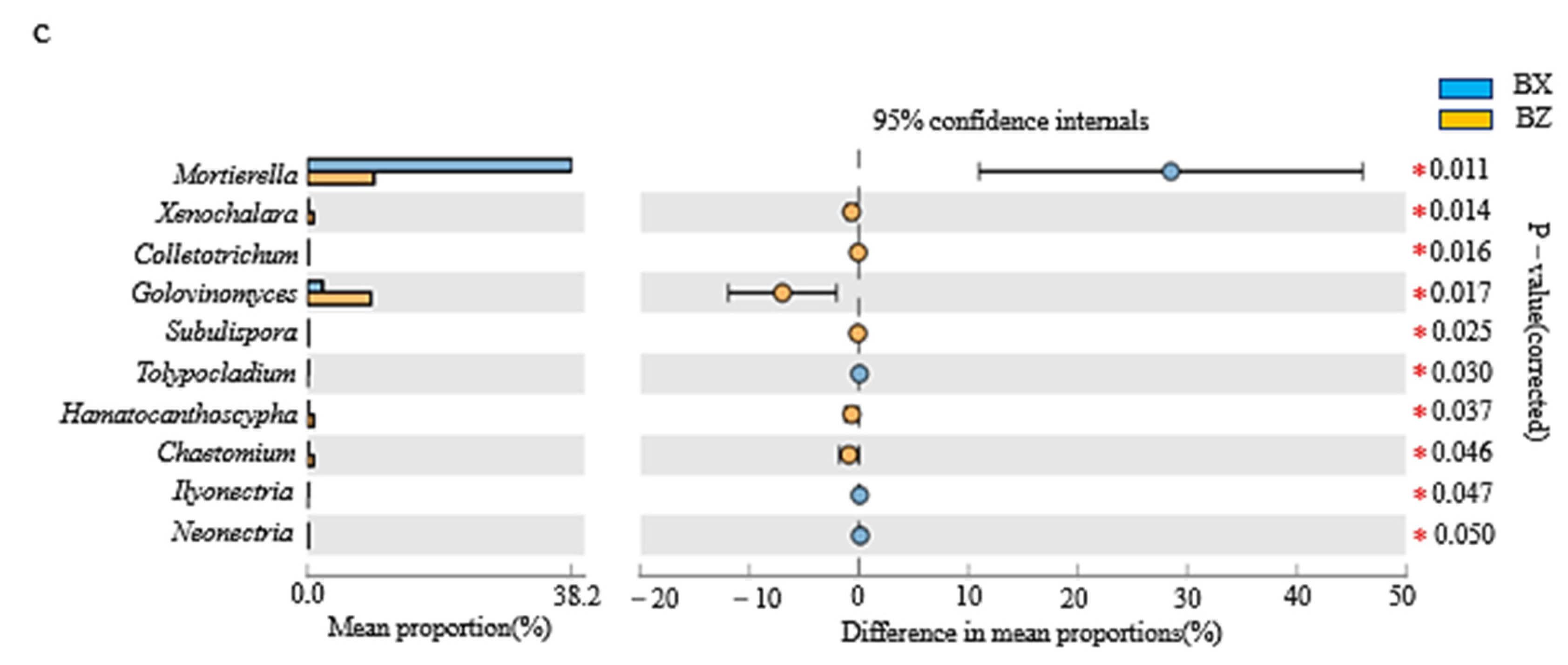
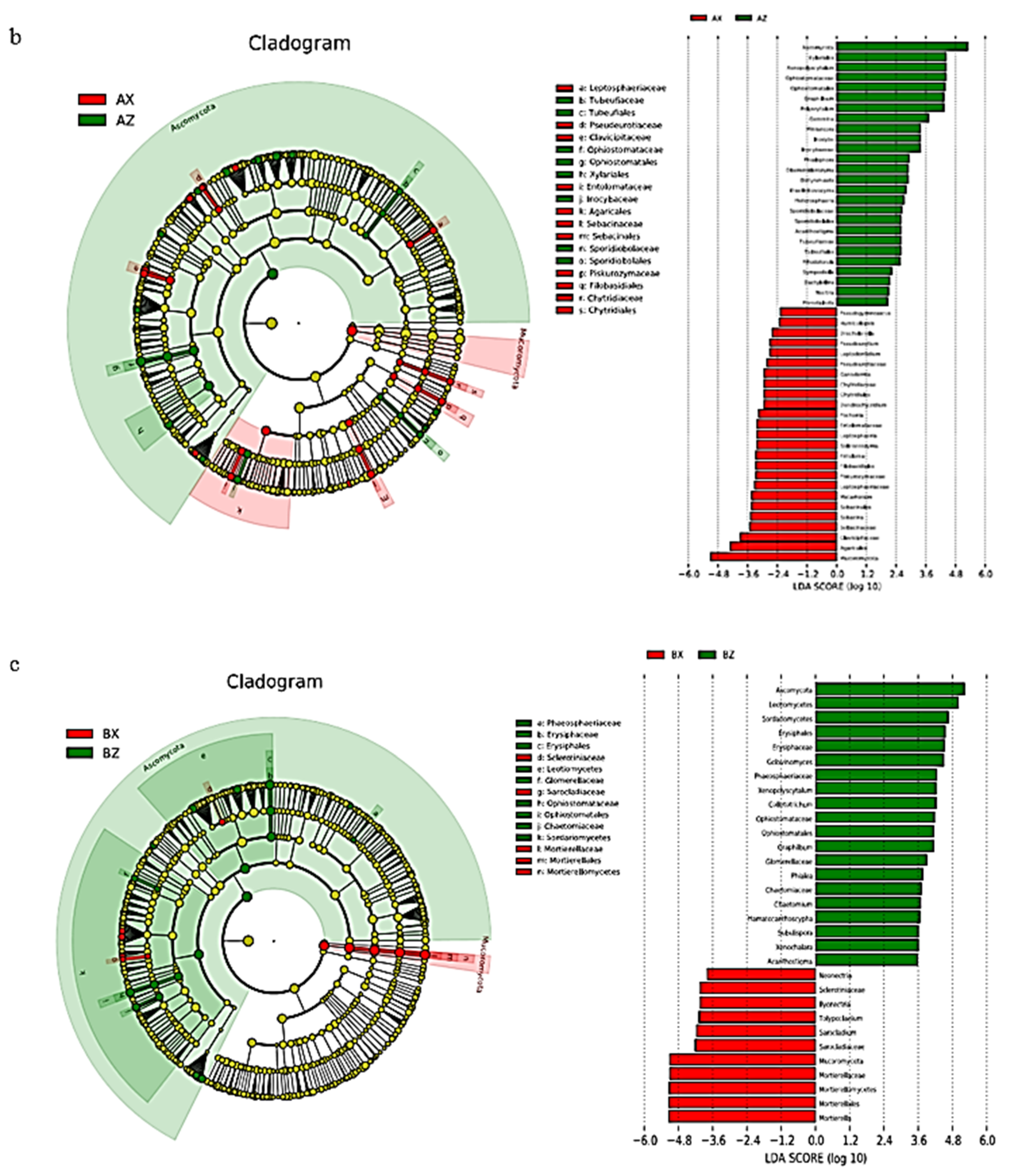
3.2.4. LEfSe Analysis of Litter Fungi
3.3. Litter-Layer Metabolomics of Korean Pine Plantation and Original Korean Pine Forest
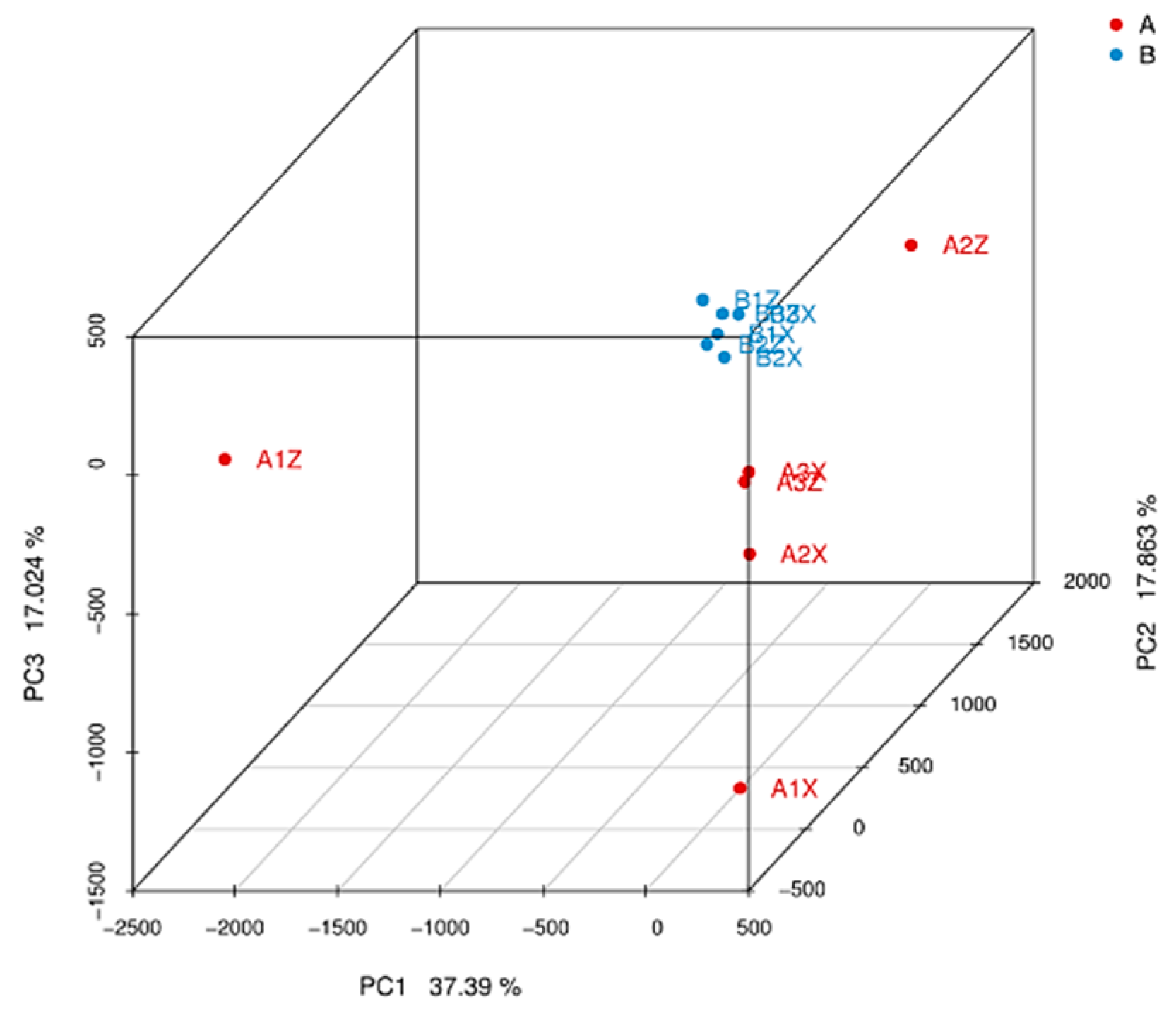
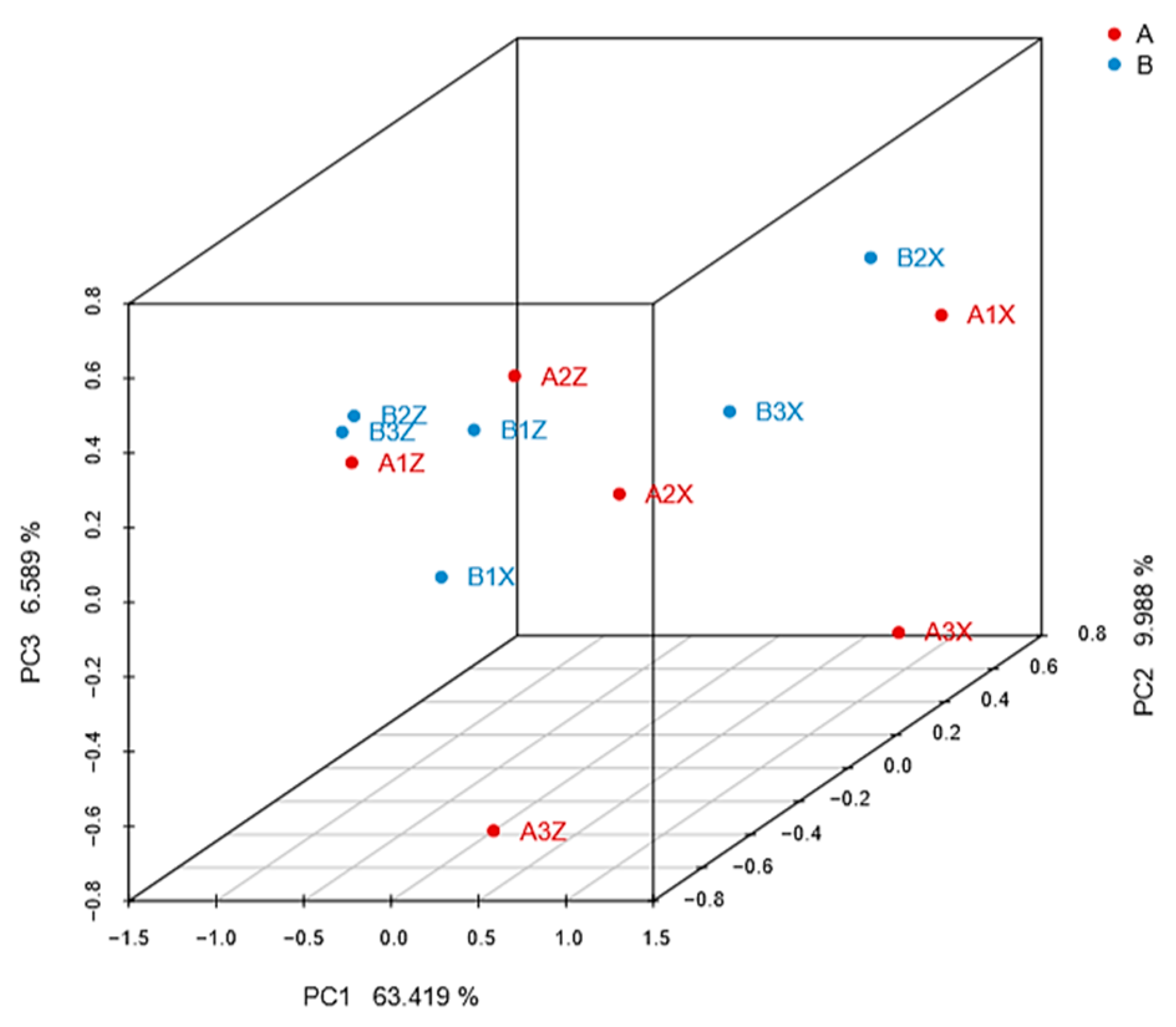
3.3.1. Screening of Different Litter Metabolites
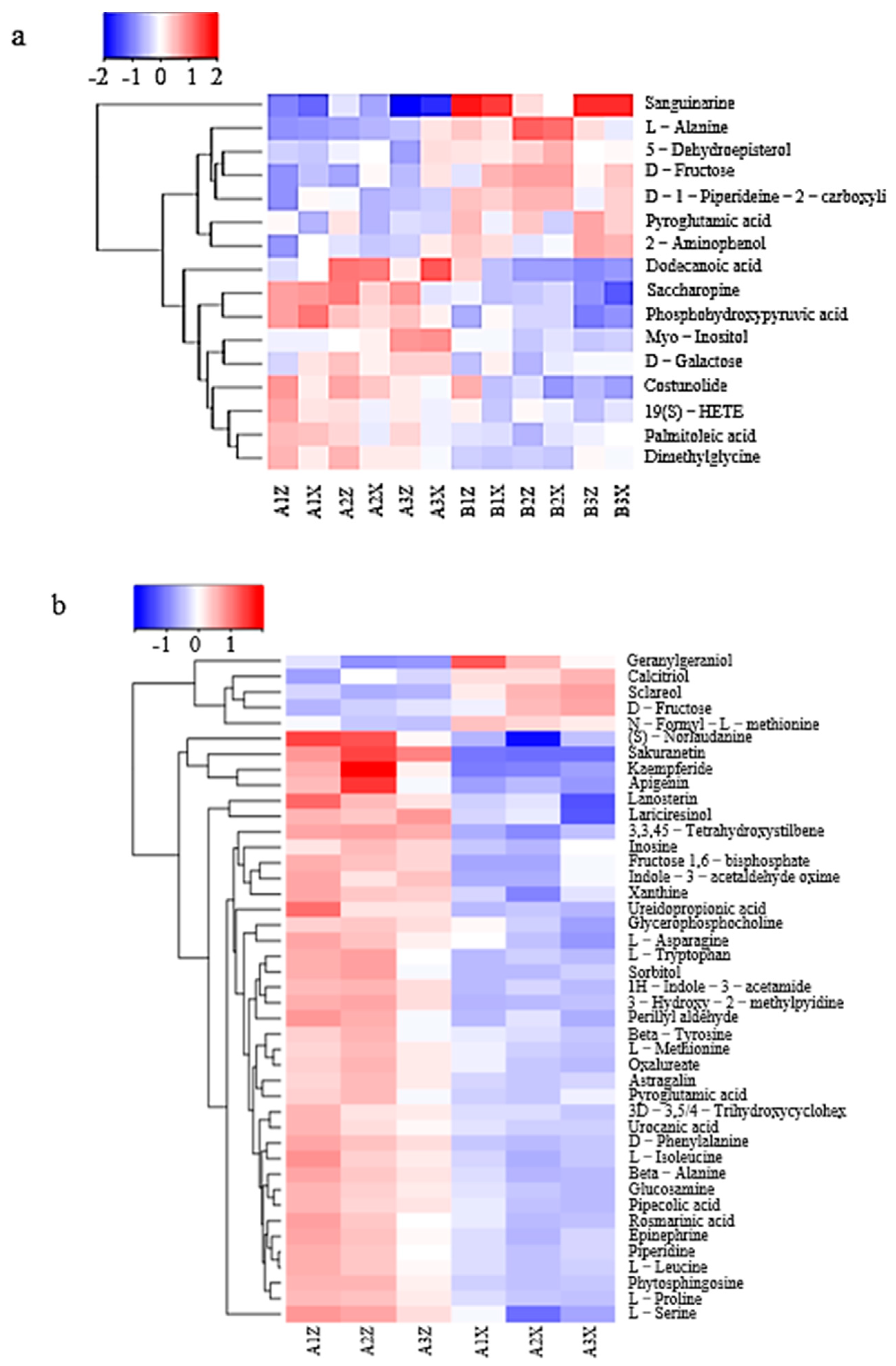
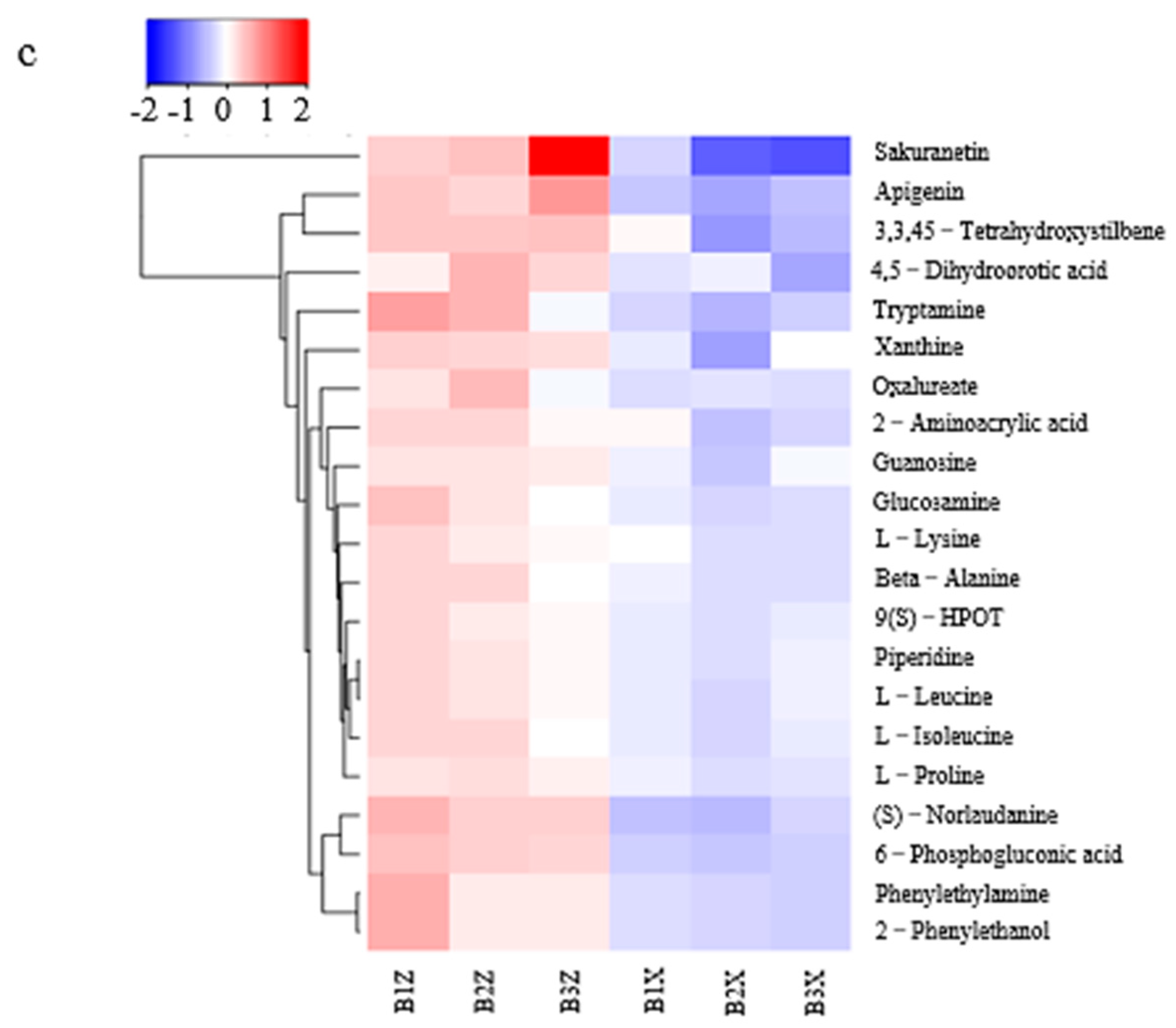
3.3.2. Differential-Metabolite-Cluster (Heatmap) Analysis of Litter
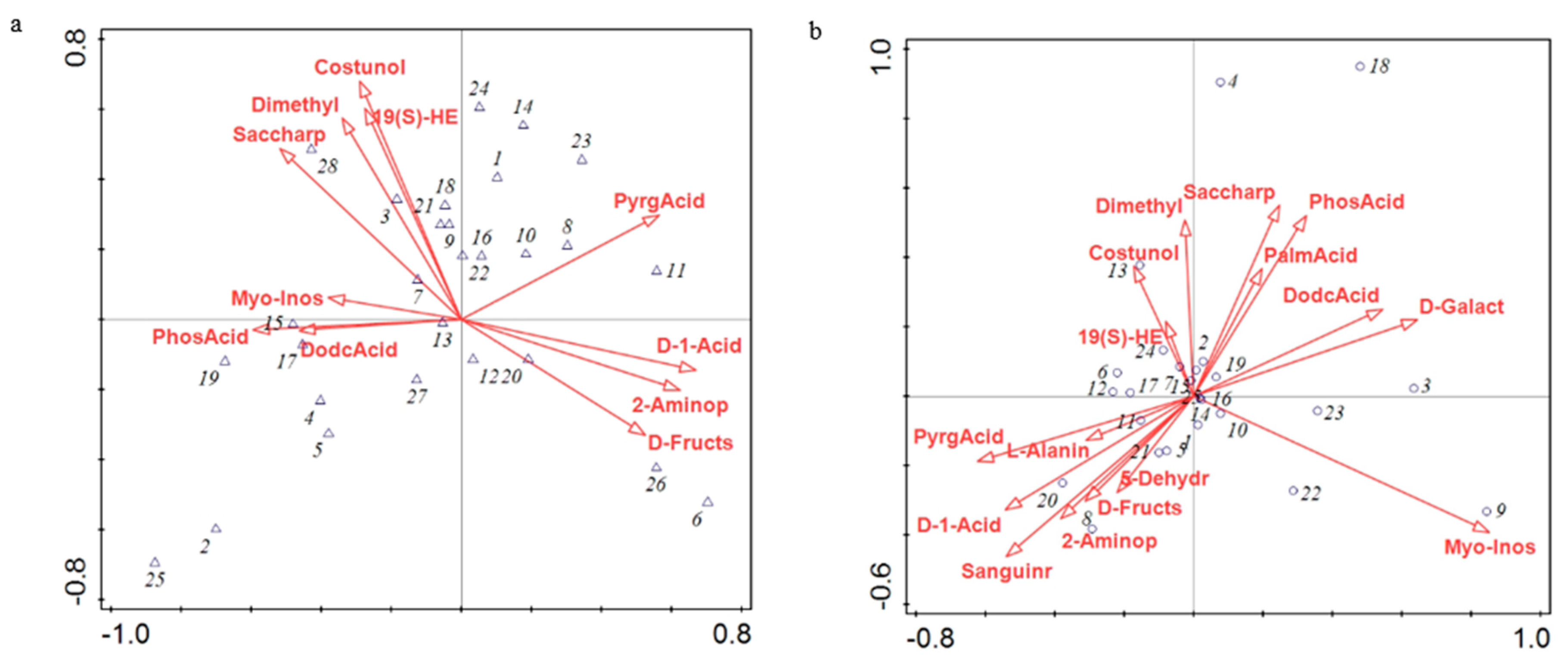
3.4. Canonical Correspondence Analysis (CCA) of Relative Microbial Abundance and Differential Metabolites
4. Discussion
4.1. Effects of Bacterial Community and Metabolites on Forest-Litter Decomposition
4.2. Effects of Fungal Communities and Metabolites on Forest-Litter Decomposition
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


References
- Froufe, L.C.M.; Schwiderke, D.K.; Castilhano, A.C.; Cezar, R.M.; Steenbock, W.; Seoane, C.E.S.; Bognola, I.A.; Vezzani, F.M. Nutrient cycling from leaf litter in multistrata successional agroforestry systems and natural regeneration at Brazilian Atlantic Rainforest Biome. Agrofor. Syst. 2020, 94, 159–171. [Google Scholar] [CrossRef]
- Birhane, E.; Desalegn, T.; Kebede, F.; Giday, K.; Hishe, H.; Hadgu, K.M. In situ leaf litter production, decomposition and nutrient release of dry Afromontane trees. East Afr. Agric. For. J. 2019, 83, 176–190. [Google Scholar] [CrossRef]
- Guo, J.; Wang, G.; Wu, Y.; Geng, Q.; Cao, F. Leaf litter and crop residue decomposition in ginkgo agroforestry systems in eastern China: Soil fauna diversity and abundance, microbial biomass and nutrient release. J. For. Res. 2019, 30, 1895–1902. [Google Scholar] [CrossRef]
- Azizan, K.A.; Ibrahim, S.; Ghani, N.H.A.; Nawawi, M.F. Metabolomics approach to investigate phytotoxic effects of Wedeliatrilobata leaves, litter and soil. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2019, 153, 691–699. [Google Scholar] [CrossRef]
- Du, J.; Cui, M.; Zhang, Y.; Guo, R.; Gao, Y.; Hu, D. Effects of nanometer zinc oxide on the microbial community structure and metabolic function of litters degradation. J. Ecotoxicol. 2012, 13, 149–158. [Google Scholar] [CrossRef]
- Deforest, J.L. Chronic phosphorus enrichment and elevated pH suppresses Quercus spp. leaf litter decomposition in a temperate forest. Soil Biol. Biochem. 2019, 135, 206–212. [Google Scholar] [CrossRef]
- Chen, M.; Lin, Y.; He, X.; Han, G. Effects of endophytic fungus colonization on litter decomposition and microbial activity of Chinese fir leaves. Bull. Microbiol. 2020, 47, 1404–1417. [Google Scholar] [CrossRef]
- Han, C.; Liu, T.; Lu, X.; Duan, L.; Singh, V.P.; Ma, L. Effect of litter on soil respiration in a man-made Populus, L. forest in a dune-meadow transitional region in China’s Horqin sandy land. Ecol. Eng. 2019, 127, 276–284. [Google Scholar] [CrossRef]
- Wang, Q.; He, T.; Liu, J. Litter input decreased the response of soil organic matter decomposition to warming in two subtropical forest soils. Sci. Rep. 2016, 6, 33814. [Google Scholar] [CrossRef]
- Krishna, M.P.; Mohan, M. Litter decomposition in forest ecosystems: A review. Energy Ecol. Environ. 2017, 2, 236–249. [Google Scholar] [CrossRef]
- Li, X.; Qu, C.; Bian, Y.; Gu, C.; Jiang, X.; Song, Y. New insights into the responses of soil microorganisms to polycyclic aromatic hydrocarbon stress by combining enzyme activity and sequencing analysis with metabolomics. Environ. Pollut. 2019, 255 (Pt 2), 113312. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.-J.; Li, Q.; Liu, C.; Jin, Z.-J.; Liang, Y.-M.; Huang, B.-H.; Lu, X.-X.; Peng, W.-J. Carbon Metabolism Characteristics of the Karst Soil Microbial Community for Pb-Zn Mine Tailings. Huanjing kexue 2018, 39, 2420–2430. [Google Scholar] [CrossRef] [PubMed]
- Mikryukov, V.; Dulya, O. Contamination-induced transformation of bacterial and fungal communities in spruce-fir and birch forest litter. Appl. Soil Ecol. 2017, 114, 111–122. [Google Scholar] [CrossRef]
- Joly, F.-X.; Fromin, N.; Kiikkilä, O.; Hättenschwiler, S. Diversity of leaf litter leachates from temperate forest trees and its consequences for soil microbial activity. Biogeochemistry 2016, 129, 373–388. [Google Scholar] [CrossRef]
- Treseder, K.K.; Bent, E.; Borneman, J.; McGuire, K.L. Shifts in fungal communities during decomposition of boreal forest litter. Fungal Ecol. 2014, 10, 58–69. [Google Scholar] [CrossRef]
- Zhang, Y.-Y.; Zhang, Q.; Zhang, Y.-M.; Wang, W.-W.; Zhang, L.; Yue-Ming, Z.; Bai, C.; Guo, J.-Z.; Fu, H.; She, Y.-B. A comprehensive automatic data analysis strategy for gas chromatography-mass spectrometry based untargeted metabolomics. J. Chromatogr. A 2019, 1616, 460787. [Google Scholar] [CrossRef]
- Xiao, W.; Guan, Q. Research status and Prospect of influence of Disturbance on decomposition of forest litters. J. Ecol. Environ. 2018, 27, 983–990. [Google Scholar] [CrossRef]
- Xi, X.; Zhang, H. Progress in microbial metabolomics research and application. Food Sci. 2016, 37, 283–289. [Google Scholar] [CrossRef]
- Withers, E.; Hill, P.W.; Chadwick, D.R.; Jones, D.L. Use of untargeted metabolomics for assessing soil quality and microbial function. Soil Biol. Biochem. 2020, 143, 107758. [Google Scholar] [CrossRef]
- Ristok, C.; Leppert, K.N.; Scherer-Lorenzen, M.; Niklaus, P.A.; Bruelheide, H. Soil macrofauna and leaf functional traits drive the decomposition of secondary metabolites in leaf litter. Soil Biol. Biochem. 2019, 135, 429–437. [Google Scholar] [CrossRef]
- Pollierer, M.M.; Ferlian, O.; Scheu, S. Temporal dynamics and variation with forest type of phospholipid fatty acids in litter and soil of temperate forests across regions. Soil Biol. Biochem. 2015, 91, 248–257. [Google Scholar] [CrossRef]
- Zhou, G.; Zhang, J.; Qiu, X.; Wei, F.; Xu, X. Decomposing litter and associated microbial activity responses to nitrogen deposition in two subtropical forests containing nitrogen-fixing or non-nitrogen-fixing tree species. Sci. Rep. 2018, 8, 12934. [Google Scholar] [CrossRef] [PubMed]
- Pei, G.; Liu, J.; Peng, B.; Gao, D.; Wang, C.; Dai, W.; Jiang, P.; Bai, E. Nitrogen, lignin, C/N as important regulators of gross nitrogen release and immobilization during litter decomposition in a temperate forest ecosystem. For. Ecol. Manag. 2019, 440, 61–69. [Google Scholar] [CrossRef]
- Wang, J.; Pisani, O.; Lin, L.H.; Lun, O.O.; Bowden, R.D.; Lajtha, K.; Simpson, A.J.; Simpson, M.J. Long-term litter manipulation alters soil organic matter turnover in a temperate deciduous forest. Sci. Total Environ. 2017, 607–608, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Radajewski, S.; Webster, G.; Reay, D.S.; Morris, S.A.; Ineson, P.; Nedwell, D.B.; Prosser, J.I.; Murrell, J.C. Identification of active methylotroph populations in an acidic forest soil by stable-isotope probing. Microbiology 2002, 148, 2331–2342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Zheng, Y.; Yang, Y. Anoxic energy metabolism of methane-oxidizing flora of proteobacteria in wetland. Acta Sodiae Sin. 2019, 7, 1–13. [Google Scholar] [CrossRef]
- Huttunen-Saarivirta, E.; Rajala, P.; Bomberg, M.; Carpén, L. Corrosion of copper in oxygen-deficient groundwater with and without deep bedrock micro-organisms: Characterisation of microbial communities and surface processes. Appl. Surf. Sci. 2017, 396, 1044–1057. [Google Scholar] [CrossRef]
- Lin, W.; Pan, Y. A putative greigite-type magnetosome gene cluster from the candidate phylum Latescibacteria. Environ. Microbiol. Rep. 2015, 7, 237–242. [Google Scholar] [CrossRef]
- Becraft, E.D.; Woyke, T.; Jarett, J.; Ivanova, N.; Godoy-Vitorino, F.; Poulton, N.; Brown, J.M.; Brown, J.; Lau, M.C.Y.; Onstott, T.; et al. Rokubacteria: Genomic Giants among the Uncultured Bacterial Phyla. Front. Microbiol. 2017, 8, 2264. [Google Scholar] [CrossRef]
- Fu, D.; Chen, Y.; Guo, Q.; Mao, L.; Fei, J.; Qiu, J. Progress in studies on the sexual type (Ascomycetes) metabolites of Epicuticella. Acta Mycol. 2008, 37, 541–554. [Google Scholar] [CrossRef]
- Lin, J.; Lin, X.R.; Ke, H.-A.; Wei, Y.; Ke, Y.; Wang, N.; Long, M. Oriental meat cut inside a bacterium EU7-22 cellulose enzyme Ⅰ heterologous expression. J. Biotechnol. 2019, 29, 1–6. [Google Scholar] [CrossRef]
- Luo, J.; Walsh, E.; Miller, S.; Blystone, D.; Dighton, J.; Zhang, N. Root endophytic fungal communities associated with pitch pine, switchgrass, and rosette grass in the pine barrens ecosystem. Fungal Biol. 2017, 121, 478–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dilegge, M.J.; Manter, D.K.; Vivanco, J.M. A novel approach to determine generalist nematophagous microbes reveals Mortierellaglobalpina as a new biocontrol agent against Meloidogyne spp. nematodes. Sci. Rep. 2019, 9, 7521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lay, C.-Y.; Hamel, C.; St-Arnaud, M. Taxonomy and pathogenicity of Olpidiumbrassicae and its allied species. Fungal Biol. 2018, 122, 837–846. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest | Component Characteristics | Age (Year) | Litter Thickness (cm) |
|---|---|---|---|
| Korean pine plantation | Artificial mixed forest of Pinus koraiensisSieb. et Zucc., Piceaasperata Mast., Larix gmelinii (Rupr.) Kuzen., Pinus sylvestris var. mongolicaLitv., Fraxinus mandshuricaRupr., and other tree species. | 65 (afforestation, 1954) | 2–13 |
| Original Korean pine forest | Pinus koraiensisSieb. et Zucc is the main species, accompanied by a variety of warm hardwood species (Tilia tuan Szyszyl., Betula., Quercus mongolica Fisch. ex Ledeb., Ulmus laciniata (Trautv.) Mayr., Acer mono Maxim., Populus ussuriensisKom.), and by some cold and temperate zones tree species in Eurasian coniferous forests, such as PiceakoraiensisNakai., Piceajezoensis var. microsperma., Abies nephrolepis (Trautv.) Maxim, coniferous, broad-leaved mixed forest. | >300 | 2–15 |
| Sample Number | Chao1 | Shannon | Simpson |
|---|---|---|---|
| AZ | 3733 ± 226 c | 6.77 ± 0.22 b | 0.0049 ± 0.0032 a |
| AX | 4030 ± 534 b | 7.06 ± 0.20 a,b | 0.0025 ± 0.0011 a,b |
| BZ | 4853 ± 292 a,b | 7.36 ± 0.11 a | 0.0015 ± 0.0003 b |
| BX | 5937 ± 795 a | 7.53 ± 0.11 a | 0.0012 ± 0.0003 b |
| Sample Number | Chao1 | Shannon | Simpson |
|---|---|---|---|
| AZ | 634 ± 125 a | 4.09 ± 0.42 b | 0.0436 ± 0.019 b |
| AX | 496 ± 96 b | 4.01 ± 0.67 b | 0.0538 ± 0.027 a |
| BZ | 589 ± 103 a,b | 4.34 ± 0.46 a | 0.0377 ± 0.024 b |
| BX | 582 ± 152 a,b | 4.08 ± 0.43 b | 0.0543 ± 0.015 a |
| Negative | Differential Metabolites | Positive | Differential Metabolites |
|---|---|---|---|
| A and B | 7 | A and B | 16 |
| AZ and AX | 22 | AZ and AX | 43 |
| BZ and BX | 9 | BZ and BX | 21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, T.; Li, C.; Song, F. Differences in Microbial Community and Metabolites in Litter Layer of Plantation and Original Korean Pine Forests in North Temperate Zone. Microorganisms 2020, 8, 2023. https://doi.org/10.3390/microorganisms8122023
Wang Y, Li T, Li C, Song F. Differences in Microbial Community and Metabolites in Litter Layer of Plantation and Original Korean Pine Forests in North Temperate Zone. Microorganisms. 2020; 8(12):2023. https://doi.org/10.3390/microorganisms8122023
Chicago/Turabian StyleWang, Yue, Ting Li, Chongwei Li, and Fuqiang Song. 2020. "Differences in Microbial Community and Metabolites in Litter Layer of Plantation and Original Korean Pine Forests in North Temperate Zone" Microorganisms 8, no. 12: 2023. https://doi.org/10.3390/microorganisms8122023