Early Intracellular Trafficking and Subsequent Activity of Programmed Cell Death in Channel Catfish Macrophages Infected with Edwardsiella ictaluri


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Antibodies, Probes and Reagents
2.3. Infection Procedure
2.4. Immunofluorescent Microscopy
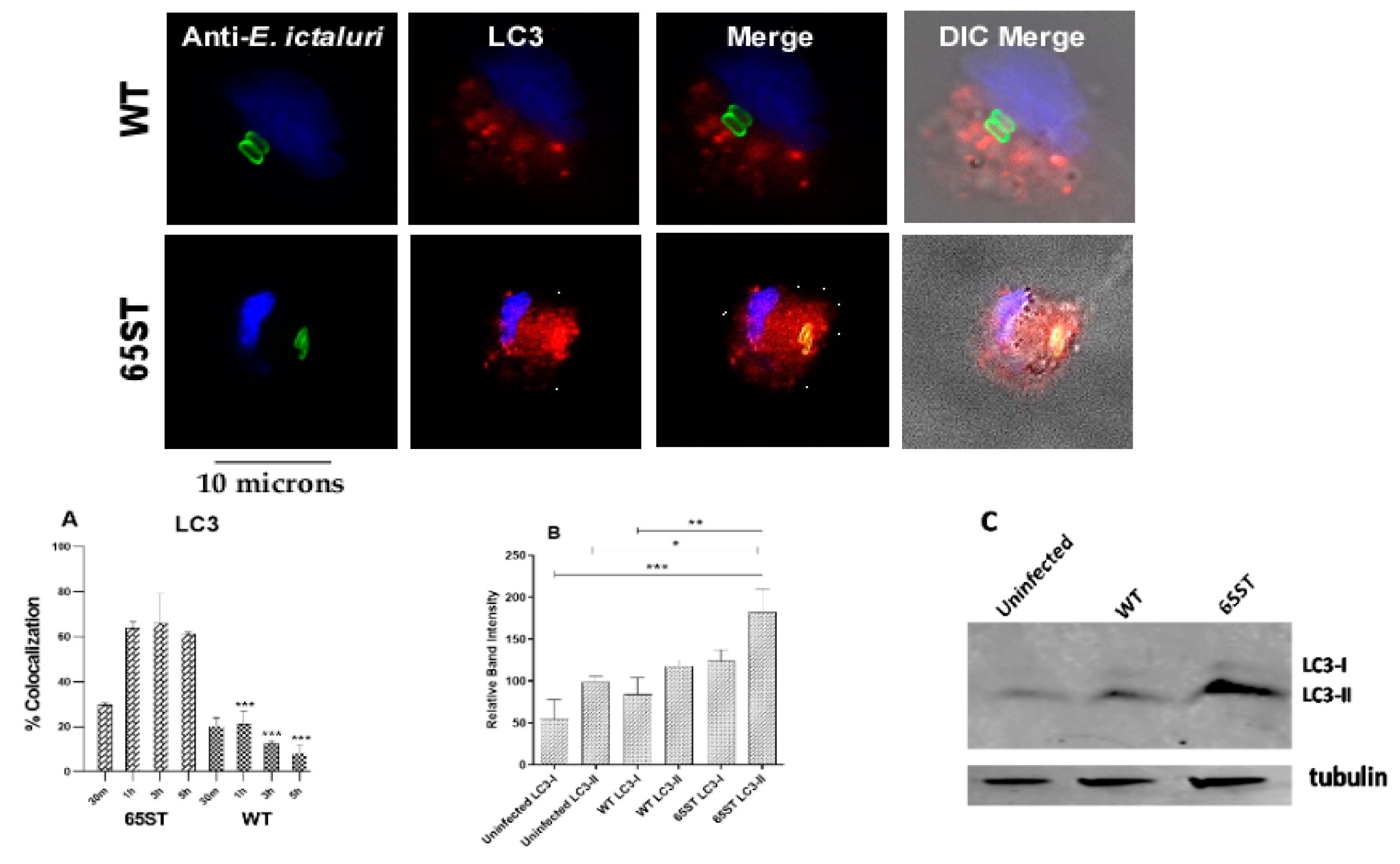
2.5. Immunoblotting of LC3/LC3-II
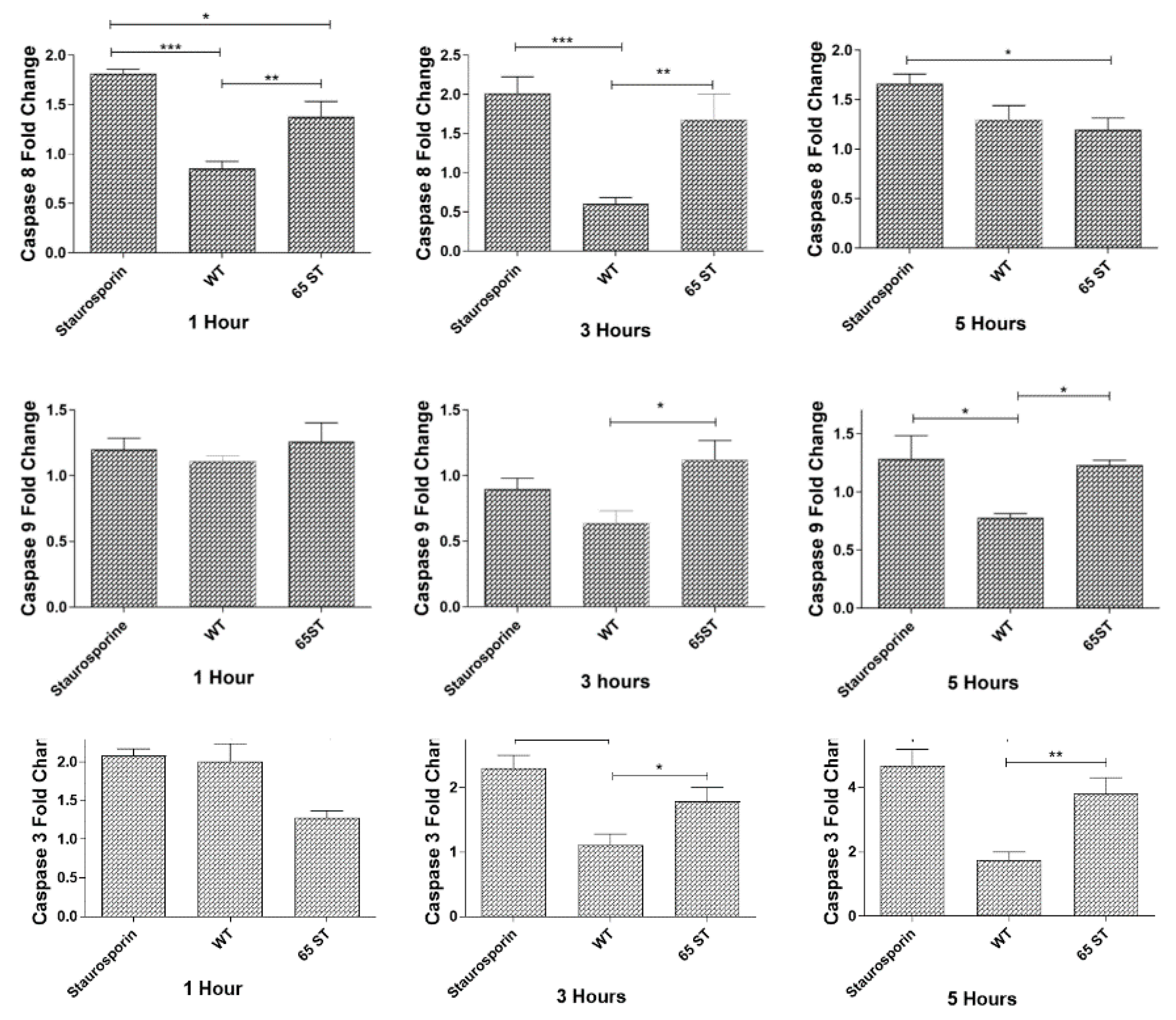
2.6. Caspase Activity
2.7. Assessment of Apoptosis by Flow Cytometry
2.8. Cytotoxicity Assay
2.9. Statistical Analysis
3. Results
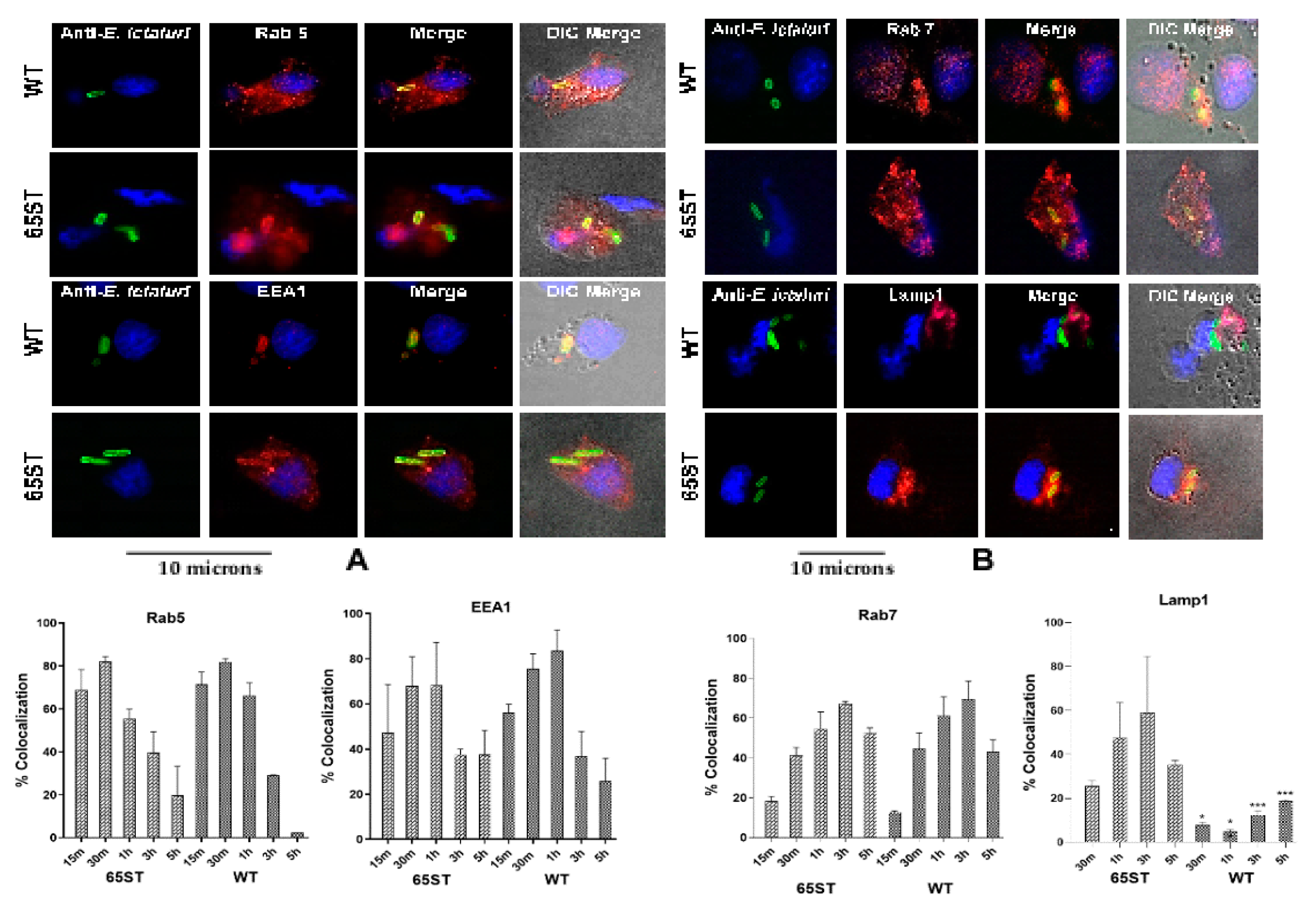
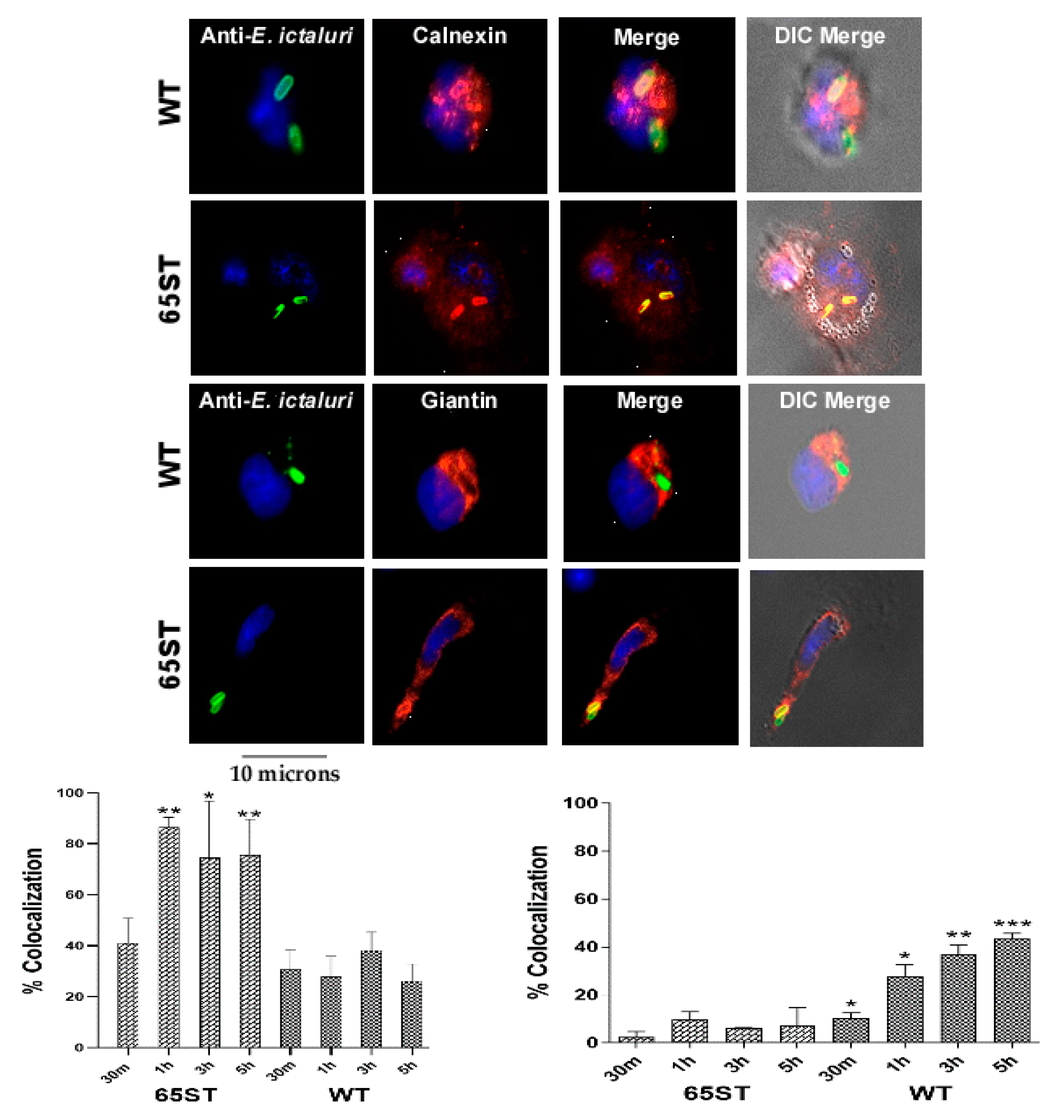
3.1. Immunofluorescence of Vacuolar Membrane Markers
3.2. Caspase Activation during E. ictaluri Infection of HKDM
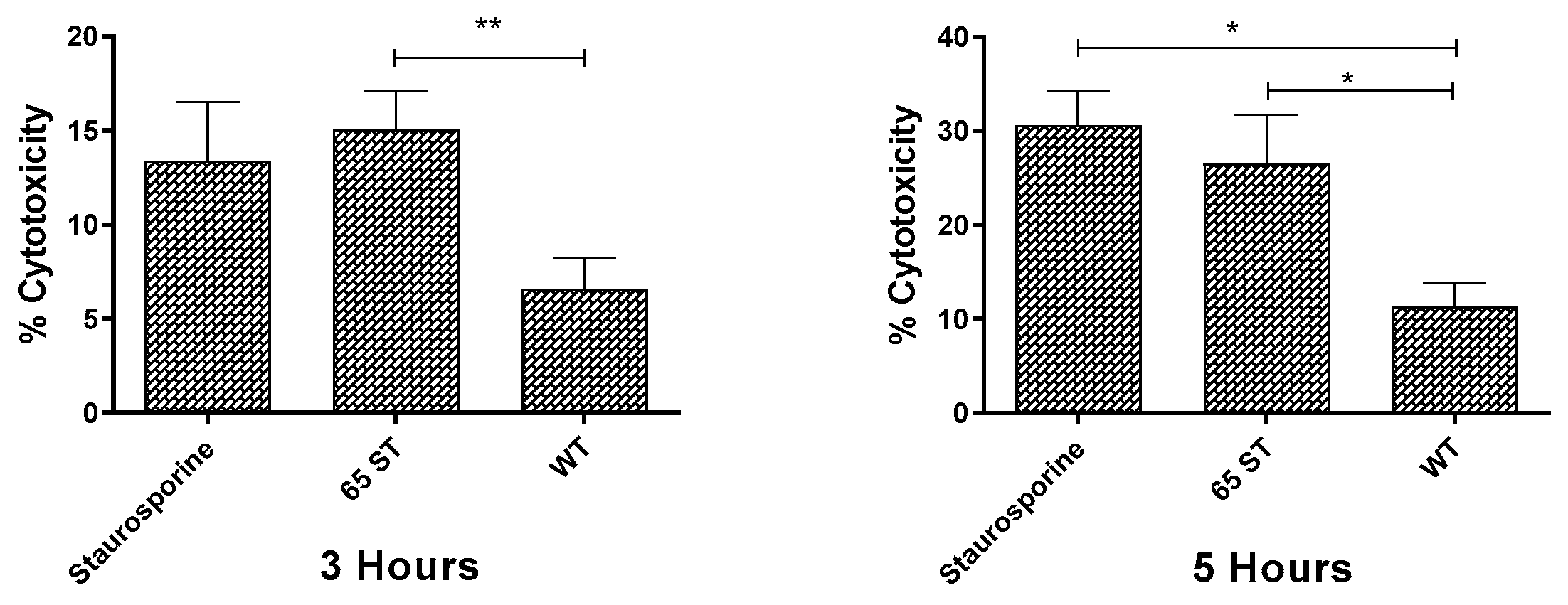
3.3. Cellular Damage in Macrophages Caused by Invasion of E. ictaluri
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Booth, N.J.; Beekman, J.B.; Thune, R.L. Edwardsiella ictaluri encodes an acid-activated urease that is required for intracellular replication in channel catfish (Ictalurus punctatus) macrophages. Appl. Environ. Microbiol. 2009, 75, 6712–6720. [Google Scholar] [CrossRef] [Green Version]
- Thune, R.L.; Fernandez, D.H.; Benoit, J.L.; Kelly-Smith, M.; Rogge, M.L.; Booth, N.J.; Landry, C.A.; Bologna, R.A. Signature-tagged mutagenesis of Edwardsiella ictaluri identifies virulence-related genes, including a Salmonella-pathogenicity-island-2 class of type III secretion system. Appl. Environ. Microbiol. 2007, 73, 7934–7946. [Google Scholar] [CrossRef] [Green Version]
- Dubytska, L.P.; Rogge, M.L.; Thune, R.L. Identification and characterization of putative translocated effector proteins of the Edwardsiella ictaluri type three secretion system. mSphere 2016, 1, e00039-00016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, D.H.; Pittman-Cooley, L.; Thune, R.L. Sequencing and analysis of the Edwardsiella ictaluri plasmids. Plasmid 2001, 45, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Rogge, M.; Thune, R. The Edwardsiella ictaluri type III secretion system is regulated by pH and phosphate concentration through EsrA, EsrB, and EsrC. Appl. Environ. Microbiol. 2011, 77, 4293–4302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumgartner, W.A.; Dubytska, L.; Rogge, M.L.; Mottram, P.J.; Thune, R.L. Modulation of vacuolar pH is required for replication of Edwardsiella ictaluri in channel catfish macrophages. Infect. Immun. 2014, 82, 2329–2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ainsworth, A.J.; Capley, G.; Waterstreet, P.; Munson, D. Use of monoclonal antibodies in the indirect fluorescent antibody technique for the diagnosis of Edwardsiella ictaluri. J. Fish Dis. 1986, 9, 439–444. [Google Scholar] [CrossRef]
- Booth, N.J.; ElKamel, A.; Thune, R.L. Intracellular replication of Edwardsiella ictaluri in channel catfish macrophages. J. Aquat. Anim. Health 2006, 18, 101–108. [Google Scholar] [CrossRef]
- Ainsworth, A.J.; Dexiang, C.; Greenway, T. Characterization of monoclonal antibodies to channel catfish, Ictalurus punctatus, leucocytes. Vet. Immunol. Immunopathol. 1990, 26, 81–92. [Google Scholar] [CrossRef]
- Zhao, D.; Lin, F.; Wu, X.; Zhao, Q.; Zhao, B.; Lin, P.; Zhang, Y.; Yu, X. Pseudolaric acid B induces apoptosis via proteasome-mediated Bcl-2 degradation in hormone-refractory prostate cancer DU145 cells. Toxicology 2012, 26, 595–602. [Google Scholar] [CrossRef]
- Cirillo, D.M.; Valdivia, R.H.; Monack, D.M.; Falkow, S. Macrophage-dependent induction of the Salmonella-pathogenicity-island-2 type III secretion system and its role in intracellular survival. Mol. Microbiol. 1998, 30, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Hensel, M. Whole genome scan for habitat-specific genes by signature-tagged mutagenesis. Electrophoresis 1998, 19, 608–612. [Google Scholar] [CrossRef] [PubMed]
- Campoy, E.; Colombo, M.I. Autophagy in intracellular bacterial infection. Biochim. Biophys. Acta 2009, 1793, 1465–1477. [Google Scholar] [CrossRef] [Green Version]
- DeLeo, F.R. Modulation of phagocyte apoptosis by bacterial pathogens. Apoptosis 2004, 9, 399–413. [Google Scholar] [CrossRef]
- Dabbs, E.R. Mapping of the genes for Bacillus subtilis ribosomal proteins s-6 and s-16 comparison of the chromosomal distribution of ribosomal protein genes in this bacterium with the distribution in Escherichia coli. Mol. Gen. Genet. 1983, 192, 386–390. [Google Scholar] [CrossRef]
- Diacovich, L.; Gorvel, J.P. Bacterial manipulation of innate immunity to promote infection. Nat. Rev. Microbiol. 2010, 8, 117–128. [Google Scholar] [CrossRef]
- Figueira, R.; Holden, D.W. Functions of the Salmonella pathogenicity island 2 (SPI-2) type III secretion system effectors. Microbiology 2012, 158, 1147–1161. [Google Scholar] [CrossRef] [Green Version]
- Pierini, R.; Cottam, E.; Roberts, R.; Wileman, T. Modulation of membrane traffic between endoplasmic reticulum, ERGIC and Golgi to generate compartments for the replication of bacteria and viruses. Semin. Cell Dev. Biol. 2009, 20, 828–833. [Google Scholar] [CrossRef]
- Gagnon, E.; Duclos, S.; Rondeau, C.; Chevet, E.; Cameron, P.H.; Steele-Mortimer, O.; Paiement, J.; Bergeron, J.J.; Desjardins, M. Endoplasmic reticulum-mediated phagocytosis is a mechanism of entry into macrophages. Cell 2002, 110, 119–131. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Q.; Yang, M.; Li, P.; Wu, C. Bacteria exploit autophagy for their own benefit. Infect. Drug Resist. 2019, 12, 3205–3215. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Chan, H.; Lu, L.; Wong, K.T.; Wong, S.H.; Li, M.X.; Xiao, Z.G.; Cho, C.H.; Gin, T.; Chan, M.T.V.; et al. Autophagy in intracellular bacterial infection. Semin. Cell Dev. Biol. 2020, 101, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Behar, S.M.; Briken, V. Apoptosis inhibition by intracellular bacteria and its consequence on host immunity. Curr. Opin. Immunol. 2019, 60, 103–110. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dubytska, L.P.; Thune, R.L. Early Intracellular Trafficking and Subsequent Activity of Programmed Cell Death in Channel Catfish Macrophages Infected with Edwardsiella ictaluri. Microorganisms 2020, 8, 1649. https://doi.org/10.3390/microorganisms8111649
Dubytska LP, Thune RL. Early Intracellular Trafficking and Subsequent Activity of Programmed Cell Death in Channel Catfish Macrophages Infected with Edwardsiella ictaluri. Microorganisms. 2020; 8(11):1649. https://doi.org/10.3390/microorganisms8111649
Chicago/Turabian StyleDubytska, Lidiya P., and Ronald L. Thune. 2020. "Early Intracellular Trafficking and Subsequent Activity of Programmed Cell Death in Channel Catfish Macrophages Infected with Edwardsiella ictaluri" Microorganisms 8, no. 11: 1649. https://doi.org/10.3390/microorganisms8111649