1. Introduction
The mammalian immune system consists of innate and adaptive immunity [
1]. Among them, innate immunity plays a role in recognizing a deviation from homeostasis due to infectious or non-infectious assaults; adaptive immunity, including humoral immunity and cell-mediated immunity; killing infected cells; and neutralizing antigens [
1,
2]. Innate immunity is mediated by phagocytes including macrophages and neutrophils and is an important component of host defense against invading microbial pathogens [
3,
4].
Lymphocytes and macrophages play an important role in innate and adaptive immunity [
5]. The activation of lymphocytes and macrophages causes the production of inflammatory cytokines and mediators, such as reactive oxygen species (ROS) and nitric oxide (NO), and the secretion of proinflammatory cytokines, such as tumor necrosis factor-α (TNF-α), interferon-(IFN)-γ, interleukin (IL)-6 and IL-12 [
6,
7,
8,
9]. These molecules are involved in the mitogen-activated protein kinase (MAPK) and the nuclear factor-κB (NF-κB) signaling pathways. NO is an important intracellular and intercellular regulatory molecule with many physiological functions. It is synthesized by inductive nitric oxide synthase (iNOS) and has many physiological roles, including immune response, vascular dilatation, neurotransmission and platelet aggregation inhibition [
7].
Loss of homeostasis in the immune system affects the immune response and leads to a variety of diseases [
10]. Cyclophosphamide (CTX) is a well-known alkylating cytotoxic drug that is effective in organ transplantation, the treatment of cancer and autoimmune diseases in part because of its ability to modulate immune responses [
11,
12]. CTX is one of the major immunomodulatory drugs used in clinical trials, which damages the DNA of normal cells and has immunosuppressive effects [
13]. However, immunomodulators, especially drugs containing single ingredients, can cause side effects such as fever, headache, hypertension and neurotoxicity. Immunosuppression can be fatal and is damaging to the immune system of the organism. Therefore, safe immunoregulatory agents that can prevent immunodeficiency in cancer patients receiving chemotherapy have been investigated, including dietary supplements and functional foods that promote health [
14].
The use of probiotics for improving immune health status in immunodeficient patients has gained a special interest in recent years [
15]. Probiotics are commonly used in various products because of the medically confirmed functions such as beneficial bacterial proliferation, harmful bacteria suppression, improvement of intestinal health, blood cholesterol reduction, immunity enhancement, endogenous infection suppression and anti-cancer effects [
16,
17]. Probiotics are used for the treatment and prevention of various diseases, but they have not been widely used due to a lack of understanding of the mechanism of action. In addition, although the immuno-promoting efficacy of lactic acid bacteria (LAB) has been actively studied, the efficacy of some specific strains have not been characterized; thus, further studies on various strains and the mechanisms of action are required [
18]. Among the 13 genera of LAB,
Lactococcus,
Lactobacillus,
Leuconostoc and
Enterococcus are known to play key roles in improving immunity, maintaining intestinal microbial balance and preventing gastrointestinal infections [
19,
20,
21,
22,
23]. Recent studies have shown that selenium exopolysaccharide produced by
Lactococcus lactis subsp.
lactis has antioxidant and immunomodulatory functions in immunosuppressive animal models [
24]. However, no studies have been reported on the mechanism of action related to the immunostimulating efficacy of
Lactococcus lactis in CTX-induced immunosuppressed mice.
In this study, we investigated the immunostimulating effects and associated mechanisms of Lactococcus lactis subsp. lactis GCWB1176 isolated from mozzarella cheese in mouse macrophages and CTX-induced immunosuppressed mice.
2. Materials and Methods
2.1. Materials
Roswell Park Memorial Institute (RPMI) 1640 medium, Dulbecco’s Modified Eagle’s Medium (DMEM), fetal bovine serum (FBS), streptomycin and penicillin were purchased from Life Technologies (Carlsbad, CA, USA). Enzyme-Linked immunosorbent assay (ELISA) kits for IL-1β, IL-2, IL-4, IL-10, IL-12p70, TNF-α and IFN-γ were from R&D Systems (Minneapolis, MN, USA). 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) was from USB Corp. (Cleveland, OH, USA) and the lactate dehydrogenase (LDH) release detection kit was from Roche Applied Science (Indianapolis, IN, USA). All kits were used according to the manufacturer’s protocols. CTX monohydrate, lipopolysaccharide (LPS), concanavalin (Con)A and dimethyl sulfoxide (DMSO) were from Sigma-Aldrich (St. Louis, MO, USA). YAC-1 cells, a mouse lymphoma cell line, and RAW264.7 cells, a mouse macrophage cell line, were obtained from ATCC (Manassas, VA, USA; no. ATCC®TIB-160TM and ATCC®TIB-71TM). Antibodies for β-actin, iNOS, NF-κB-p65 and secondary antibody (HRP-linked anti-rabbit IgG) were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). piNOS-Luc and pNF-κB-Luc were purchased from Addgene (Watertown, MA, USA).
2.2. Microbial Strain and Culture Conditions
Lactococcus lactis subsp. lactis GCWB1176 was isolated from mozzarella cheese and filed by GREEN CROSS Wellbeing (GCWB), Seongnam, Korea. This strain is deposited in the Korea Culture Center of Microorganisms under registration number KCCM12687P. Lactococcus lactis subsp. lactis GCWB1176 was cultured in optimized medium under anaerobic conditions. The strain was cultured in De Man, Rogosa and Sharpe (MRS) broth (Sigma-Aldrich, St. Louis, MO, USA) under anaerobic conditions at 37 °C for 24 h. The bacterial cells of stationary phase were centrifuged at 8000× g for 20 min at 4 °C. The cell pellet was washed twice with sterile phosphate-buffered saline (PBS) buffer (Life Technologies, Carlsbad, CA, USA) and adjusted to the appropriate density in DMEM or RPMI1640 medium containing 10% FBS 20 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) (Life Technologies, Carlsbad, CA, USA) for the in vitro experiments or in sterile PBS for the in vivo experiments.
2.3. Measurement of Live Cells
The numbers of live GCWB1176 (1 × 1011 CFU/g freeze-dried lactobacilli) were quantified by a 10-fold serial dilution and the plate-count method of MFDS (Ministry of Food and Drug Safety). Cell density during culturing in either the batch culture or MBR was analyzed by real-time measurement of optical density at 660 nm (G10S UV-Vis; Scinco Co., Gangnam, Korea).
2.4. Safety and Antimicrobial Susceptibility Testing (E-Test)
For the hemolysis test,
Lactococcus lactis subsp.
lactis GCWB1176 was plated on blood agar plates, containing 5% (
w/
v) sheep blood (Synergy Innovation, Seoul, Korea), and incubated for 18 h at 37 °C. Blood agar plates were examined for signs of β-hemolysis (clear zones around colonies), α-hemolysis (green-hued zones around colonies), or γ-hemolysis (no zones around colonies). For Lactococcus strains, the minimum inhibitory concentrations (MICs) (μg/mL) of nine antibiotics were determined using commercial E-test (Liofilchem MIC test Strip, Roseto degli Abruzzi, Italy): ampicillin, vancomycin, gentamicin, kanamycin, streptomycin, erythromycin, clindamycin, tetracycline and chloramphenicol. Bacterial suspensions with a turbidity equivalent to McFarland standard 1 were swabbed evenly onto MRS agar (BD DIFCO, 288210) plates with a sterile cotton swab. After drying the surfaces of the plates, the E-test strips (AB Biodisk, Solna, Sweden) of all antimicrobial agents tested (0.016–256 μg/mL) were applied. E-test strips were placed on the surface of the inoculated agar and incubated at 37 °C for 24 h. MICs were read directly from the test strip according to the instructions of the manufacturer. The concentration-dependent susceptibility was determined according to the European Food Safety Authority (EFSA) [
25].
2.5. Cell Culture and Sample Treatment
Mouse splenocytes, YAC-1 cells and RAW264.7 macrophages were cultured in RPMI 1640 and DMEM medium containing 10% FBS, streptomycin sulfate and penicillin at 37 °C and 5% CO2. Cells were treated with various concentrations (1.65 × 104–1.65 × 106 cfu/mL) of GCWB1176, LPS (10 ng/mL) and ConA (1 μg/mL) for various times.
2.6. Isolation of Splenocytes
The spleen tissues were aseptically extracted from the ICR mouse (six-week-old male, 20 ± 2 g). The tissues were disaggregated via passage through a 70 μm nylon mesh (Becton-Dickinson, Bedford, MA, USA) in RPMI-1640 medium (Life Technologies, Carlsbad, CA, USA) and the cells were purified via centrifugation at 450× g for 5 min and red blood cells (RBC) were removed with ACK lysis buffer (Sigma-Aldrich, St. Louis, MO, USA). Splenocytes were then washed with PBS, centrifuged at 1000× g for 5 min and suspended in RPMI containing 10% FBS, penicillin and streptomycin sulfate.
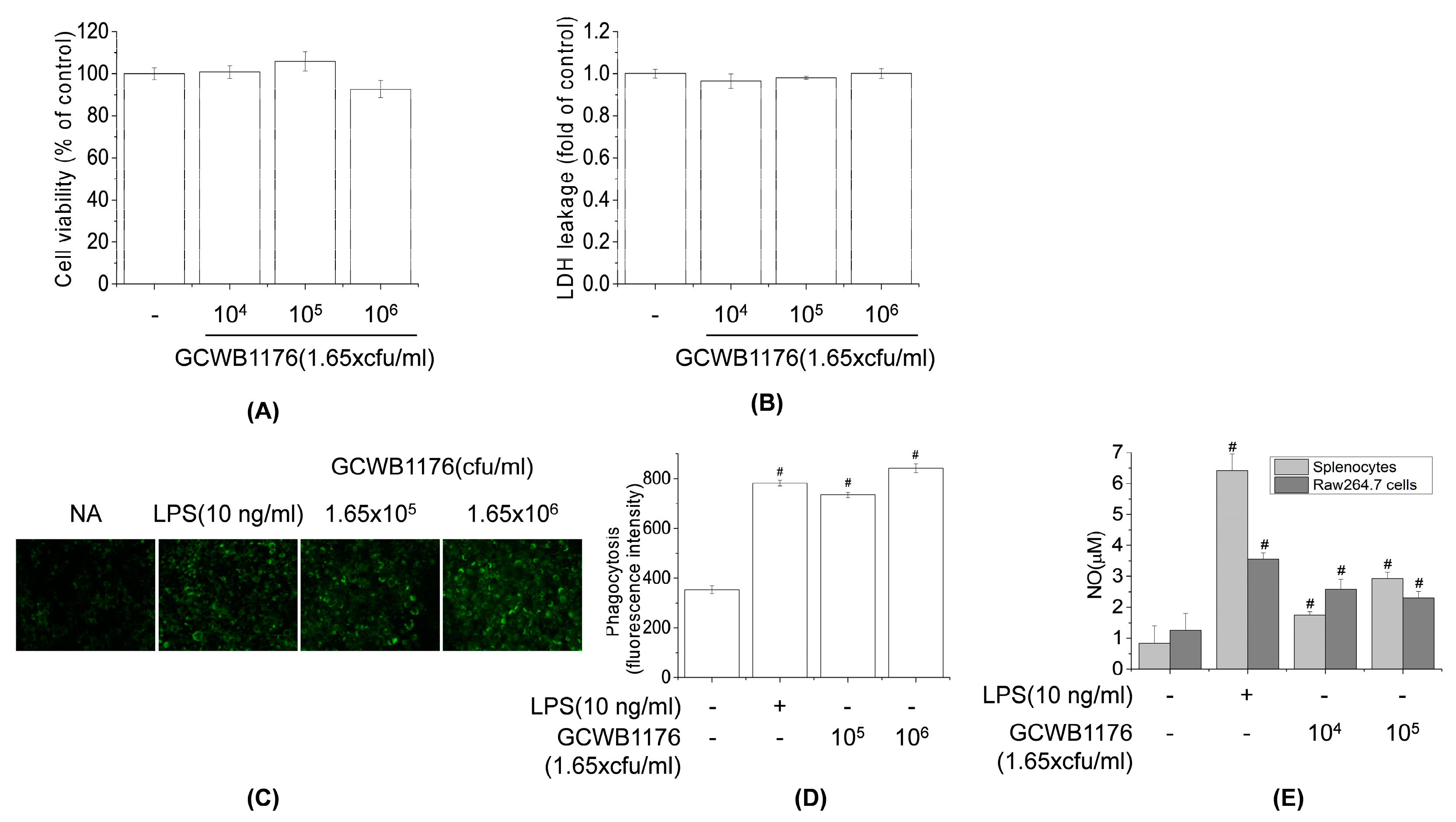
2.7. Measurement of Cell Cytotoxicity and Proliferation
Cell viability was assessed with the MTT assay or the LDH cytotoxicity assay. RAW264.7 mouse macrophage cells (1 × 106 cells/well) were seeded in 96-well plates containing 100 µL RPMI 1640 medium and 10% FBS and then incubated for 24 h. GCWB1176 (1.65 × 104–1.65 × 106 cfu/mL) was added, followed by incubation for 24 h. MTT solution (5 mg/mL in PBS) was added for 30 min at 37 °C and 5% CO2; the medium was then discarded and formazan crystals were dissolved with 200 µL DMSO, and the absorbance of each well at 550 nm was measured with a microplate reader (Varioskan; Thermo Fisher Scientific, Waltham, MA, USA). Cell proliferation was examined with a water-soluble tetrazolium (WST)-1 assay kit according to the manufacturer’s instructions. Briefly, mouse spleen cells (1 × 106 cells/well) were seeded in 96-well plates in DMEM with 10% FBS. After mixing with 10 µL RPMI 1640 medium (negative control) or 10 µL of 200 μg/mL ConA (positive control), the cells were incubated at 37 °C and 5% CO2 for 48 h. The supernatant was removed, and the cells were used in the WST-1 assay. Relative cytotoxicity was quantified by measuring the absorption at 550 nm with a microplate reader.
2.8. Preparation of the Cyclophosphamide-Induced Immunosuppression Model in Mice
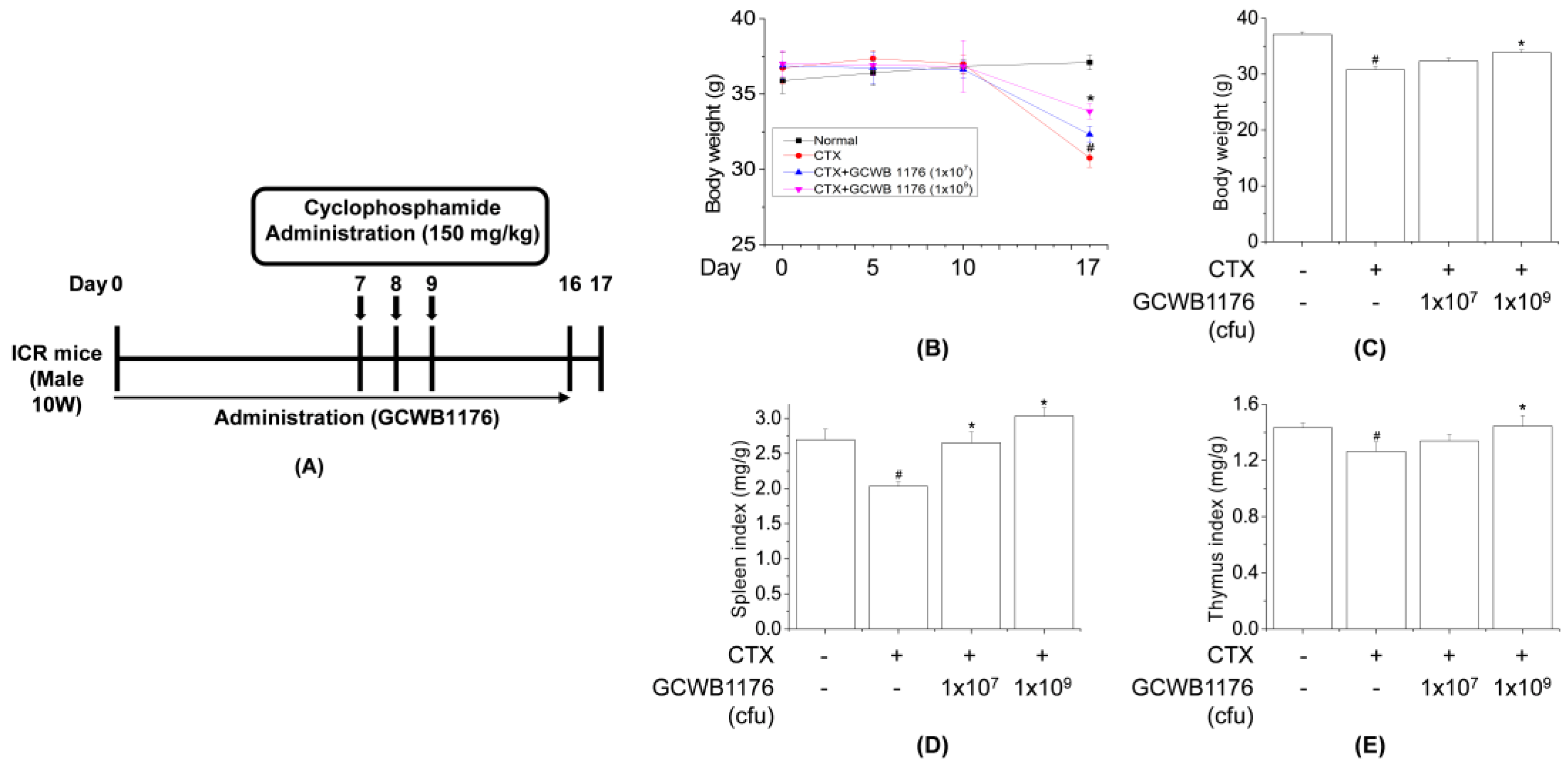
Ten-week-old male ICR mice (30 ± 2 g) were obtained from Samtako (Osan, Korea) and housed in a room with controlled temperature (22 °C ± 2 °C) and humidity (50% ± 5%) on a 12:12-h light/dark cycle with free access to food and water. The mice used in this study were handled in accordance with the Guidelines for the Care and Use of Laboratory Animals published by the U.S. National Institutes of Health (NIH Publication NO. 85-23, 1996), and all experimental procedures were approved by the Committee on Ethics of Animal Experiments of International University of Korea (Ethics NO, IUK-2019-10-1). ICR mice were divided into four groups (
n = 8), each consisting of normal, cyclophosphamide (CTX), CTX+GCWB low (1 × 10
7 cfu) and CTX+GCWB high (1 × 10
9 cfu). In total, 24 mice were injected intraperitoneally (i.p.) with CTX in sterile saline (150 mg/kg) for 3 consecutive days to establish immunosuppressed models (
Figure 1). GCWB1176 in saline was administered intragastrically at 1 × 10
7 cfu and 1 × 10
9 cfu once daily for 16 days. Mice were treated on Days 7–9 by intraperitoneal (i.p.) injection of cyclophosphamide (CTX, 150 mg/kg/day) in a total volume of 100 µL of saline. For the measurement of body and spleen weights, mice were weighed on Days 0 and 17. At the end of the experiment, the mice were sacrificed by injection of 200 mg/kg pentobarbital, and organs including the spleen were immediately removed and weighed. The immune organ index (%) was calculated according to the formula: Index = organ weight (mg)/body weight (g).
2.9. Assay for Macrophage Phagocytosis
The phagocytosis assay was performed as previously described [
26]. Briefly, 100 µL fluorescein-5-isothiocyanate (FITC)-labeled
Escherichia coli (Molecular Probes, Eugene, OR, USA) were added to the wells of a 96-well plate containing RAW264.7 macrophages. The plate was incubated for various times at 37 °C in a humidified atmosphere of 5% CO
2. Extracellular fluorescence was quenched by adding 100 mL Trypan blue. After 1 min, FITC-labeled bacteria that had not been phagocytosed by macrophages were washed away, and the macrophages were rinsed twice with PBS and then lysed with lysis buffer (10 mM Tris–HCl [pH 7.5], 130 mM NaCl, 1% Triton X-100, 10 mM Na
2HPO
4 and 10 mM Na
4P
2O
7). The relative fluorescence intensity of bacteria inside the macrophages was determined at excitation and emission wavelengths of 480 and 520 nm, respectively, using a microplate reader. The relative phagocytic activity was calculated as the percent fluorescence intensity of sample-supplemented vs. unsupplemented (control) FITC-labeled bacteria.
2.10. Assay of NK Cell Activity
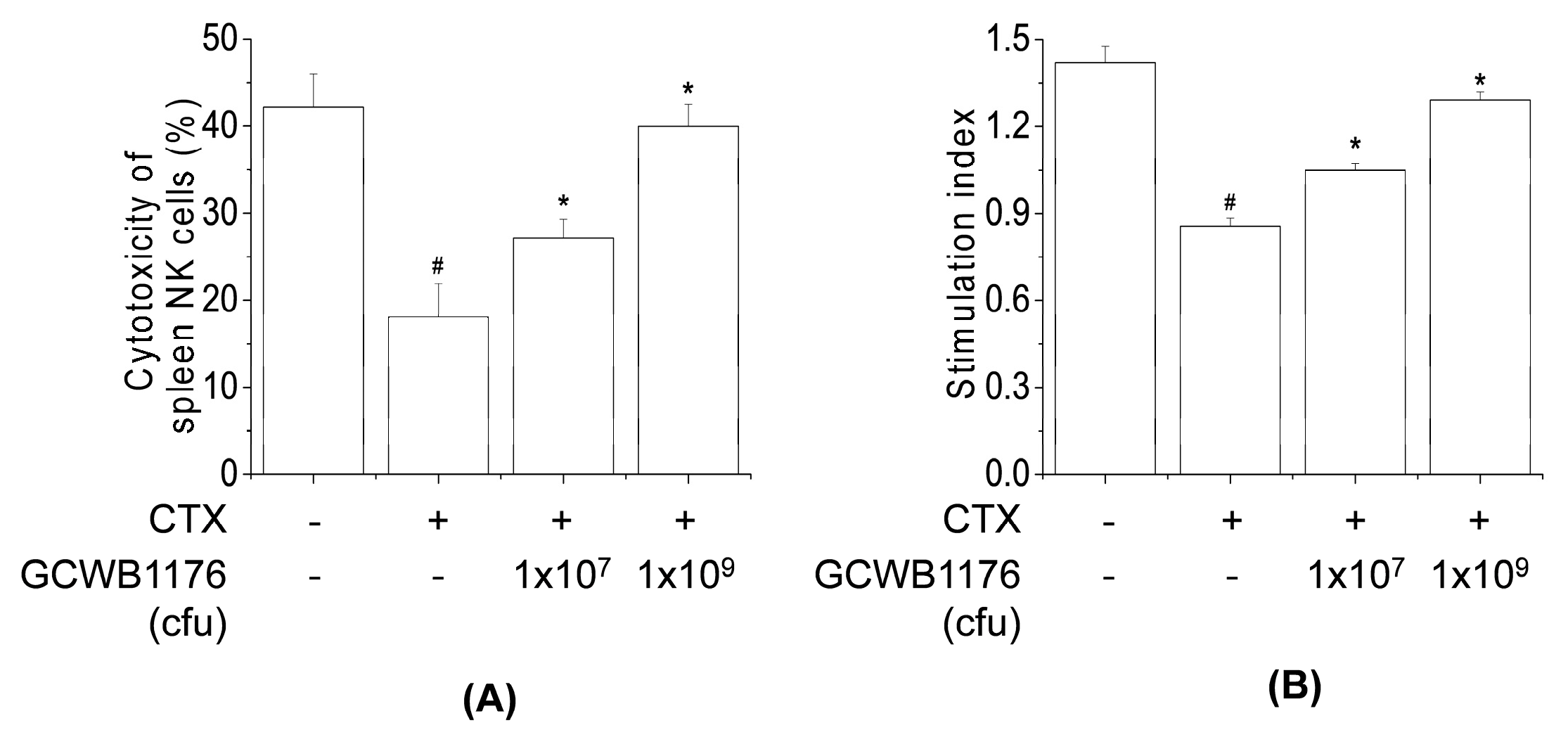
The spleen tissues were aseptically extracted from each mouse and ground into a single-cell suspension using sterile gauze and washed three times with RPMI 1640 medium. Cells were centrifuged at 1000× g for 5 min at room temperature and 100 µL of spleen cell suspension (1 × 106 cells/well) were seeded in a 96-well cell culture plate with 1 × 105 NK-sensitive YAC-1 cells at an effector cell:target cell ratio of 10:1, while 100 µL RPMI 1640 medium were used as a control. After 4-h incubation at 37 °C (5% CO2), the plate was centrifuged at 800× g for 5 min and the culture supernatant (100 µL/well) was mixed with LDH solution (Promega, Madison, WI, USA); the absorbance of each well was measured at 490 nm. NK cell cytotoxicity was calculated using the following formula: Cytotoxicity (%) = [(experimental release−spontaneous release)/(maximum release − spontaneous release)] × 100.
2.11. Measurement of Cytokine Levels
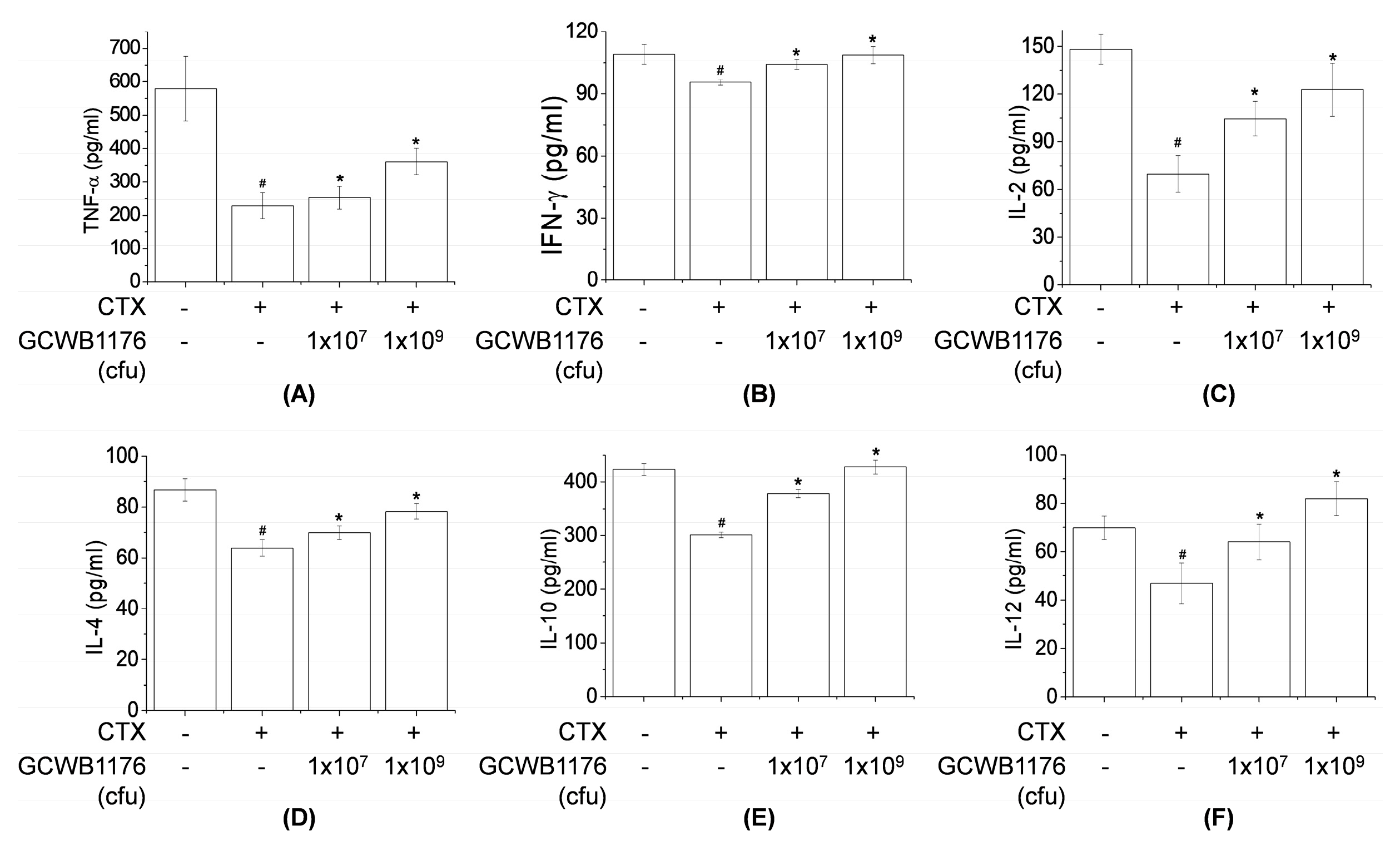
The cytokine levels in blood samples or cell culture media were quantified using ELISA kits, according to the manufacturer’s instructions. Briefly, RAW264.7 cells and splenocytes were cultured for 3 or 30 h at a density of 5 × 105 cells/well in 96-well plates. Supernatants were removed at the indicated times and cytokine (IL-1β, IL-2, IL-4, IL-10, IL-12p70, TNF-α and IFN-γ) production was analyzed based on sandwich immunoassays, as per the manufacturer’s protocol (R&D Systems).
2.12. Nitrite Assay
RAW264.7 cells (5 × 105 cells/well) were seeded in 96-well plates containing 200 µL RPMI1640 medium and 10% FBS and then incubated overnight. Cells were treated with various concentrations of GCWB1002 (1.65 × 104 and 1.65 × 105 cfu/mL) for 48 h. The cell culture media were collected, and nitrite was measured using Griess reagent. Equal volumes of Griess reagent (1:1 of 1% sulfanilamide in 5% phosphoric acid and 0.1% N-1 naphthylethylenediamine in 5% phosphoric acid) and sample were incubated together at room temperature for 5 min. Absorbance at 550 nm was measured using a microplate reader (Varioskan; Thermo Fisher Scientific, Waltham, MA, USA).
2.13. Western Blotting
RAW264.7 macrophages were treated with LPS (10 ng/mL) or GCWB1176 for 24 h or 1 h, and the protein levels of iNOS and NF-κB-p65 were determined immunochemically using specific antibody. The total protein or nuclear protein concentration of supernatants was estimated using the Bradford method and 50 μg protein was separated with 10% SDS-PAGE and then transferred onto polyvinylidene difluoride membranes. RAW264.7 macrophages were cultured with GCWB1176. Total cellular protein (50 μg) was then resolved by 10% SDS–PAGE and transferred onto polyvinylidene difluoride membranes. After blocking, the membranes were incubated with target antibody. Horseradish peroxidase-conjugated secondary antibody to IgG was used. The blots were probed using the ECL Western blot detection system, as instructed by the manufacturer.
2.14. Statistical Analysis
Results are expressed as mean values ± standard deviation (SD) of triplicate experiments. Data from the animal study are expressed as mean ± SD (n = 8). Mean differences were evaluated by analysis of variance followed by Dunnett’s post hoc test, and p values < 0.05 were considered statistically significant.
4. Discussion
The immunostimulatory potential of probiotics may be useful for the prevention and treatment of complex disorders ranging from diarrhea to allergy. Several LABs, including
Lactococcus,
Enterococcus,
Lactobacillus and
Leuconostoc, have been reported to have beneficial health effects by stimulating the immune system [
19,
20,
21,
22,
23]. The immunostimulatory mechanism of LAB comprises regulation of the T cell effector subset, the improvement of humoral immunity and activation of macrophages and lymphocytes [
32]. In the present study, we investigated the immunomodulatory properties of
Lactococcus lactis GCWB1176 isolated from mozzarella cheese using mice splenocytes and a mouse model of CTX-induced immunosuppression.
Probiotics have recently become popular among natural products with immunostimulating effects. One important component for the development of probiotics as a health functional food is the presence of antibiotic resistance genes. Interestingly, many studies have shown that several species of LAB, including
S. thermophilus, contain antibiotic-resistant genes or exhibit resistance to antibiotics, such as tetracycline, streptomycin chloramphenicol, gentamycin and kanamycin [
33]. In this study,
Lactococcus lactis subsp.
lactis, isolated from mozzarella cheese, is a nonpathogenic Gram-positive bacterium, and it is the most commonly used cheese starter. GCWB1176 did not exert a hemolysis effect on cells compared to the positive control
E. coli ATCC 25922. Although the EFSA cutoffs calculated here indicated that GCWB1176 does not show resistance against antibiotics, further studies are required to determine whether resistance genes can confer acquired resistance at a genetic level using PCR or whole genome sequencing.
CTX is commonly used to study immunomodulatory activity. CTX suppresses the immune system through inactivation of macrophages and lymphocytes and reduces the levels of inflammatory cytokines [
11,
12,
13]. We investigated the immunostimulating effect of GCWB1176 in CTX-induced immunosuppressive mice. In our model, CTX reduced the spleen and thymus index in mice, which reflects non-specific immunity. Several studies have reported that immunomodulatory agents, including
Lactobacillus plantarum, can restore spleen and thymus weight in CTX-induced immunosuppressive mice [
34,
35]. Similarly, we found that GCWB1176 orally administered for 16 consecutive days to CTX-treated mice increased spleen and thymus indices as compared to mice treated with CTX alone. The results also show that CTX markedly reduced the body weight. In addition, our results show that the spleen and thymus indices in GCWB1176 treatment groups were significantly higher than those in the CTX-treated group at 16 days, and the spleen and thymus indices were close to normal level at 16 days. These results suggest that GCWB1176 could resist the immunosuppressive effect on immune organ development.
Several authors have reported that immunostimulating agents including LAB could stimulate the production of proinflammatory cytokines and NK cell activity in cyclophosphamide-induced immunosuppressed animals [
13,
22,
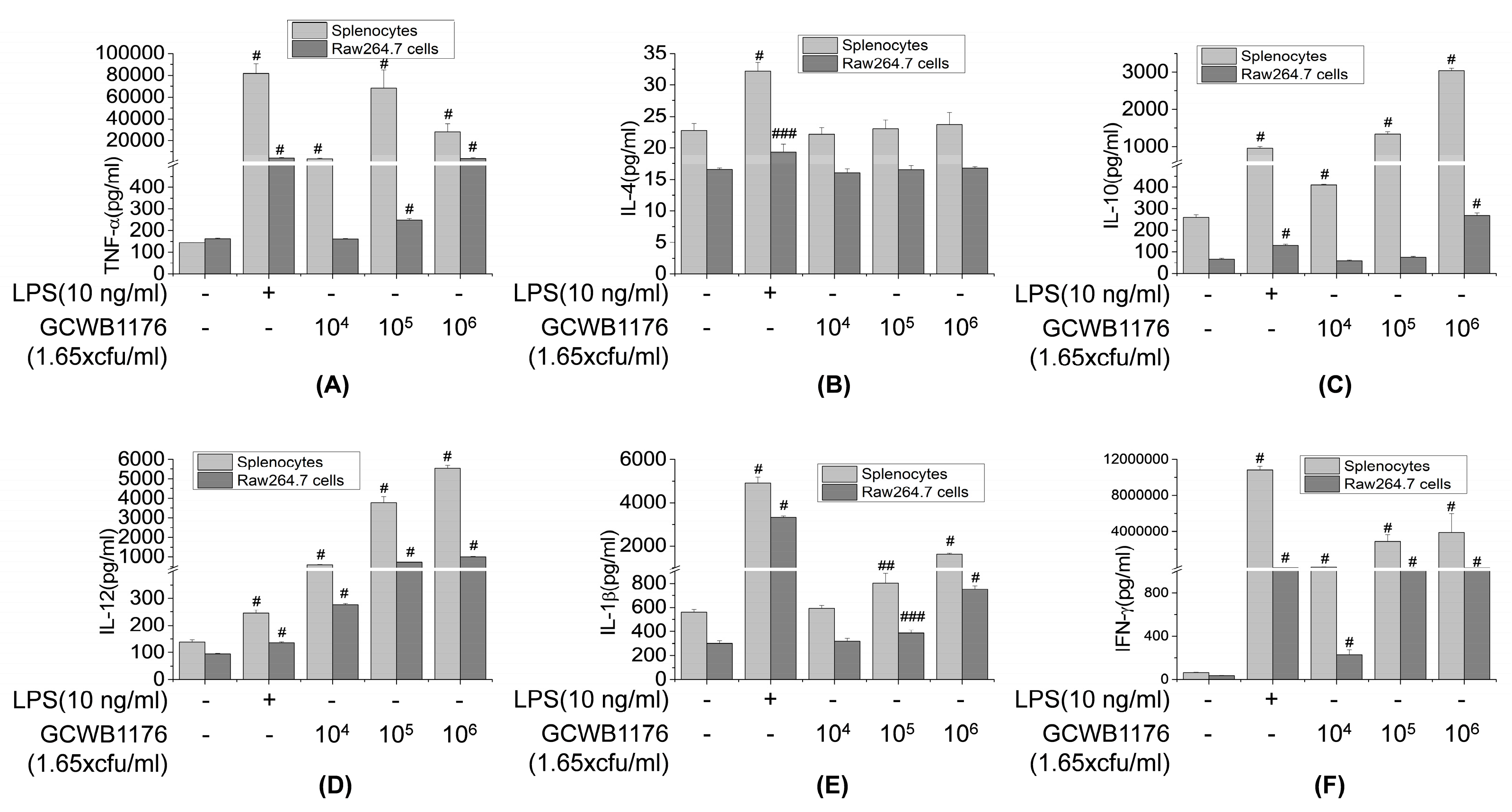
33]. Our results demonstrate that GCWB1176 induced the secretion of TNF-α, IFN-γ, IL-2, IL-4, IL-10 and IL-12p70 in mice with CTX-induced immunosuppression. In addition, our results demonstrate that GCWB1176 enhanced lymphocyte proliferative responses to T mitogen and NK cell activity in mice with CTX-induced immunosuppression.
Macrophages play an important role in host defense mechanisms and are an important part of the innate immune system. Many immunomodulators including LAB activate the immune response by activating macrophages [
13,
22,
33]. The function of macrophages in innate immunity is initiating, propagating and performing phagocytosis against pathogens [
29]. In addition, the activated macrophages produce a variety of immunomodulators including proinflammatory cytokines and NO [
30]. NO, which is synthesized from
l-arginine by inducible nitric oxide synthase, contributes to the killing of foreign pathogens and tumor cells and mediates a variety of biological functions as an intracellular messenger molecule [
7]. Activated macrophages stimulate the production of various immunomodulatory cytokines, such as TNF-α, IL-6, IL-10 and IL-12 [
35]. These immunomodulatory cytokines are involved in phagocytosis, inflammation, cell differentiation, proliferation, apoptosis and the promotion of immune cell functions [
5,
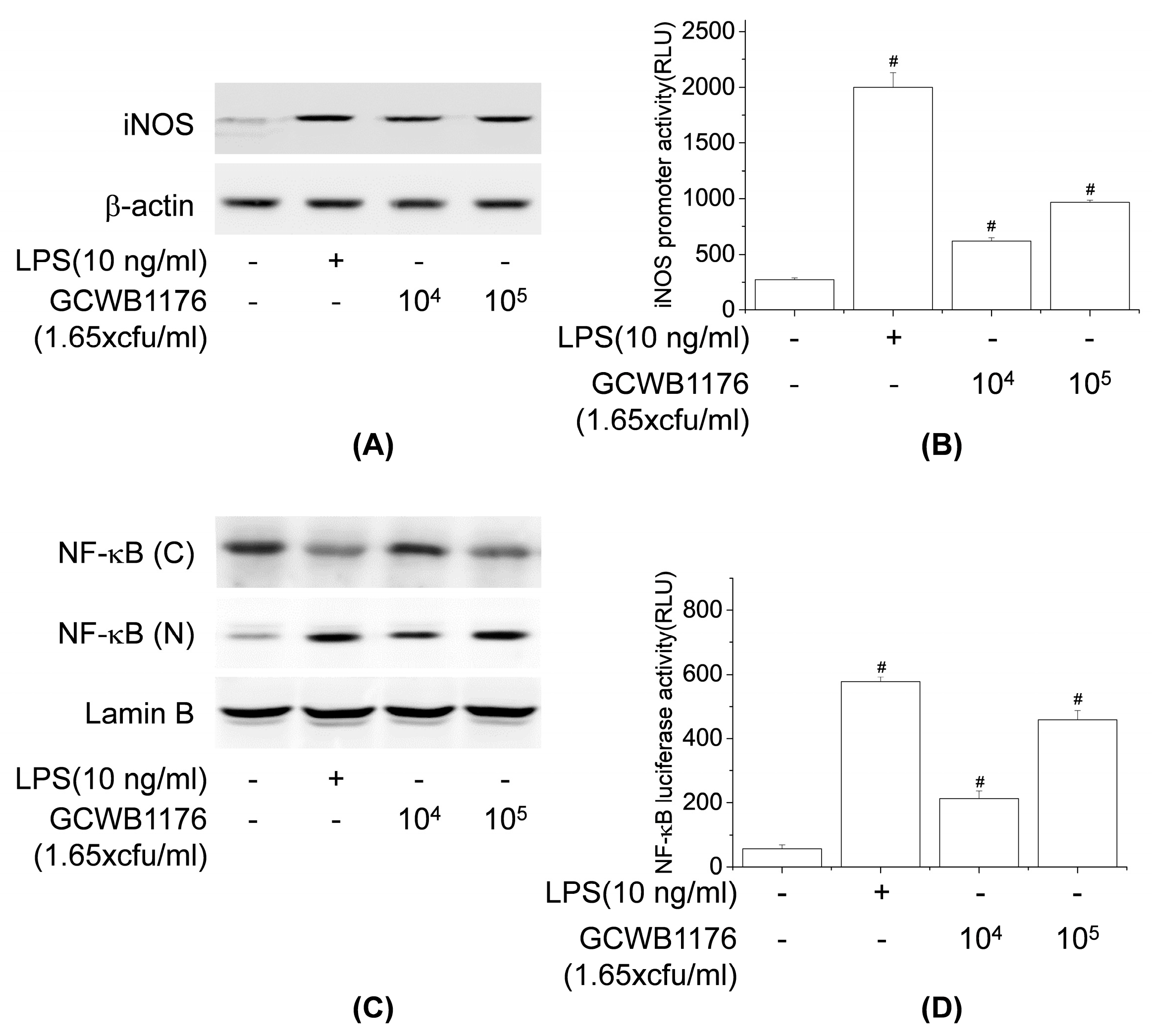
6]. In the present study, we found that GCWB1176 enhanced the phagocytic activity of RAW264.7 macrophages. In the present study, GCWB1176 increased the production of NO and macrophage-related cytokines TNF-α, IFN-γ, IL-1β, IL-4, IL-10 and IL-12p70 in mouse splenocytes and RAW264.7 macrophages, as well as enhanced iNOS protein expression and iNOS promoter luciferase activity in RAW264.7 macrophages. It was recently reported that the increase in immunomodulatory cytokines are associated with activation of NF-κB pathways [
36]. In macrophages, NF-κB is an important transcriptional factor of immune activation that upregulates the expression of many cytokines [
36]. It was recently reported that
Lactobacillus rhamnosus GG,
Lactobacillus helveticus IMAU70129,
Lactobacillus casei IMAU60214 and
Lactobacillus plantarum LM1004 induce NF-κB activation in human and mouse macrophages [
37,
38]. In the present study, we observed enhanced nuclear translocation of NF-κB and NF-κB promoter luciferase activity in RAW264.7 macrophages.
In summary, we evaluated the immunomodulatory effect of GCWB1176 in mouse primary splenocytes and RAW264.7 macrophages. GCWB1176 enhanced the phagocytic ability of macrophages, increased NK cell activity and increased the expression of immune modulators such as NO and cytokines. In addition, our results show that the NF-κB signaling pathways are responsible for these effects. More importantly, GCWB1176 improved spleen and thymus indices in CTX-induced immunosuppressed mouse model. These findings suggest that Lactococcus lactis GCWB1176 could function as an effective immunostimulatory agent in early innate immune responses.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}