A Review of the Functional Annotations of Important Genes in the AHPND-Causing pVA1 Plasmid
by
 ,
,
Hao-Ching Wang
1,2,3,* ,
,
Shin-Jen Lin
3,4,
Arpita Mohapatra
2,5,
Ramya Kumar
3,4 and
Han-Ching Wang
3,4,* 1
The Ph.D. Program for Translational Medicine, College of Medical Science and Technology, Taipei Medical University and Academia Sinica, Taipei 110, Taiwan
2
Graduate Institute of Translational Medicine, College of Medical Science and Technology, Taipei Medical University, Taipei 110, Taiwan
3
International Center for the Scientific Development of Shrimp Aquaculture, National Cheng Kung University, Tainan 701, Taiwan
4
Department of Biotechnology and Bioindustry Sciences, College of Bioscience and Biotechnology, National Cheng Kung University, Tainan 701, Taiwan
5
Mits School of Biotechnology, Utkal University, Bhubaneswar, Odisha 751004, India
*
Authors to whom correspondence should be addressed.
Microorganisms 2020, 8(7), 996; https://doi.org/10.3390/microorganisms8070996
Submission received: 19 June 2020
/
Revised: 30 June 2020
/
Accepted: 1 July 2020
/
Published: 3 July 2020
(This article belongs to the Special Issue Bacterial Responses to Environmental Stress and Their Specific Contribution to Escherichia coli and Vibrio spp. Survival and Virulence)
Abstract
:Acute hepatopancreatic necrosis disease (AHPND) is a lethal shrimp disease. The pathogenic agent of this disease is a special Vibrio parahaemolyticus strain that contains a pVA1 plasmid. The protein products of two toxin genes in pVA1, pirAvp and pirBvp, targeted the shrimp’s hepatopancreatic cells and were identified as the major virulence factors. However, in addition to pirAvp and pirBvp, pVA1 also contains about ~90 other open-reading frames (ORFs), which may encode functional proteins. NCBI BLASTp annotations of the functional roles of 40 pVA1 genes reveal transposases, conjugation factors, and antirestriction proteins that are involved in horizontal gene transfer, plasmid transmission, and maintenance, as well as components of type II and III secretion systems that may facilitate the toxic effects of pVA1-containing Vibrio spp. There is also evidence of a post-segregational killing (PSK) system that would ensure that only pVA1 plasmid-containing bacteria could survive after segregation. Here, in this review, we assess the functional importance of these pVA1 genes and consider those which might be worthy of further study.
1. Introduction
Acute hepatopancreatic necrosis disease (AHPND) is a bacterial disease that causes severe damage in shrimp farming [1]. In 2009, the first outbreak of AHPND was seen in China, and the disease quickly spread to Vietnam, Malaysia, Thailand, Philippines [1], and Bangladesh [2]. According to a recent report, it is now also present in the USA [3].
The original AHPND-causing bacteria was identified as a specific strain of Vibrio parahaemolyticus [4,5,6,7,8,9,10]. V. parahaemolyticus is a Gram-negative halophilic bacterium that can be found in marine environments [11]. In the past, V. parahaemolyticus was already recognized as a causal agent for human acute gastroenteritis in raw, contaminated sea foods [11], and it contains two hemolysin virulence factors, thermostable direct hemolysin (tdh), and TDH-related hemolysin (trh), that are both confirmed to cause damage to human cells [11]. Now, however, by acquiring a unique plasmid, pVA1, that encodes the binary pore-forming toxins PirAvp and PirBvp (often written as simply PirABvp), V. parahaemolyticus has also become a lethal pathogen to penaeid shrimps [4,5,6,7,8,9,10]. The major clinical AHPND symptom in V. parahaemolyticus-infected penaeid shrimps is a pale-to-white atrophied hepatopancreas (HP) [4,5,6,7,8,9,10]. Histological examinations further showed that PirABvp toxins caused shrimp HP cell death and led to the characteristic sloughing of the damaged epithelial cells into the HP Tubules [4,5,12].
Although the binary PirABvp toxins together seem to play the main role in damaging the HP cells [5,9,10], the pathogenesis of AHPND-causing V. parahaemolyticus is not yet fully understood. For example, we still do not know how V. parahaemolyticus colonizes the various shrimp organs, or how do shrimp cells respond to V. parahaemolyticus infection. In the course of the disease, apart from the PirABvp toxins, there is also very little that is known of the functional roles of other genes/proteins from the pVA1 plasmid, as well as those from the V. parahaemolyticus bacterium itself. To address these issues, advanced techniques, such as next generation sequencing, structural biology, and system biology have been applied [5,13,14]. These approaches have led to a recent report by Kumar et al., which found that crude bile acids affect the AHPND-causing V. parahaemolyticus by inducing biofilm formation and triggering an increased release of PirABvp toxins [14]. In addition, transcriptomic analysis of AHPND-causing V. parahaemolyticus treated with crude bile acid showed alterations in several metabolic and cellular signaling pathways [14]. However, although studies that look beyond the PirABvp toxins can provide valuable insights into AHPND, to date, there are still only a handful of reports that relate to other gene products of the pVA1 plasmid.
2. The AHPND-Causing Plasmid pVA1
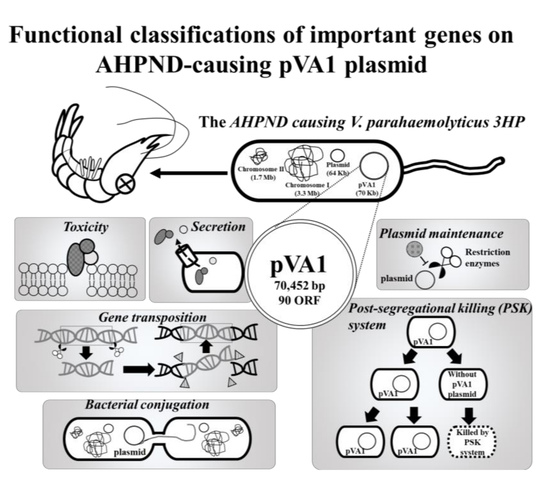
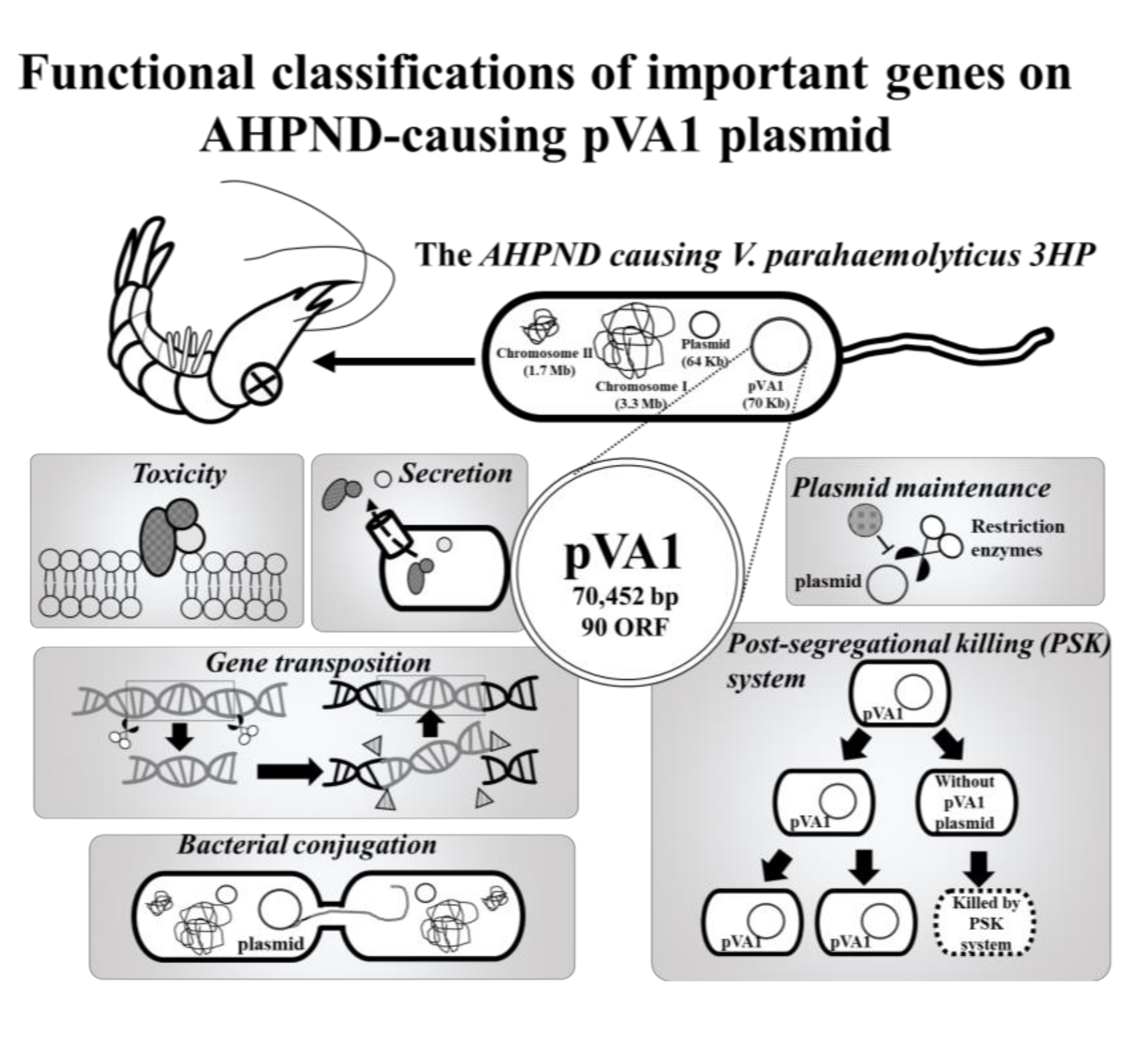
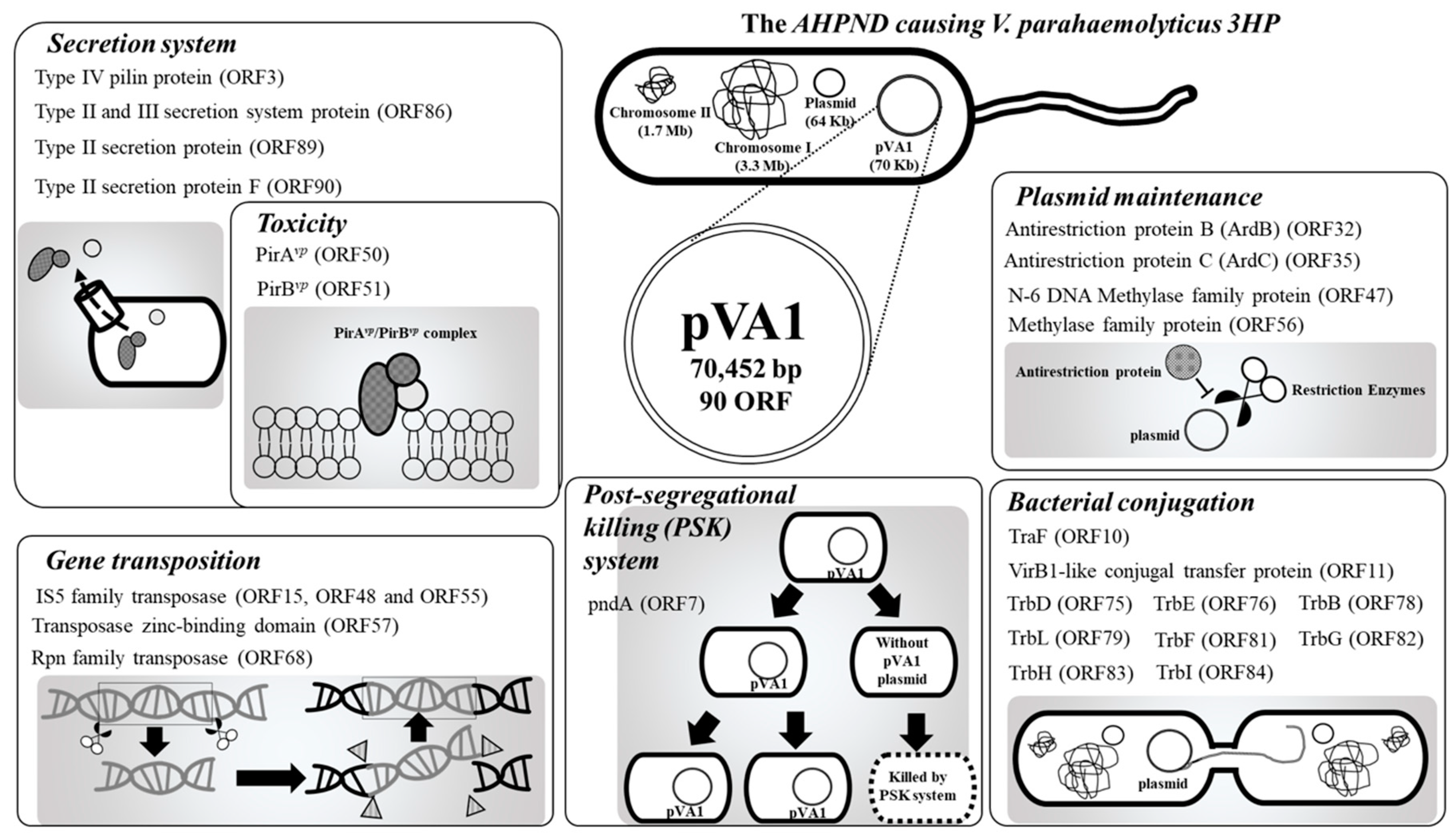
The AHPND-causing plasmid, pVA1, was first discovered by next-generation sequencing (NGS). By comparing the DNA sequences between V. parahaemolyticus strains that caused AHPND (3HP, 5HP, and China) and those that did not (S02), a 69-kb plasmid, pVA1, was found in the 3HP, 5HP, and China strains but not in the S02 strain [15,16,17,18,19]. Subsequently, the pVA1-encoded binary pore-forming PirABvp toxins were identified as the key factors in the AHPND pathology [5], and in addition to specific strains of V. parahaemolyticus, other Vibrio species, such as V. harveyi, V. campbellii, V. owensii, and V. punensis, have also become AHPND-causing agents [20,21,22,23]. All of these AHPND-causing Vibrio spp. harbor pVA1-related plasmids that contain the pirA and pirB toxin genes [5,20,21,22,23]. Recently, a non-Vibrio bacterium, Microccocus luteus, was also reported to carry the pirA and pirB toxin genes [24]. Another report confirmed that pVA1 pirABvp genes could be transferred from an AHPND-causing V. parahaemolyticus to a non-Vibrio and non-pathogenic Algoriphagus sp. [25]. These developments suggest that pVA1-related plasmids could be transmitted between different species, which would make it more difficult to control AHPND. It is already known that pVA1-related plasmids contain a number of genes for horizontal gene transfer, plasmid transmission, and maintenance, including the post-segregational killing (PSK) system. Meanwhile, components of type II and III secretion systems may further facilitate the toxic effects of pVA1-containing bacterial spp. Therefore, in addition to the toxic pirA and pirB genes, these plasmid-related genes (Figure 1; Table 1) are also worthy of further study. In this review, we will discuss the functional roles of these genes/proteins, and hopefully, open new areas for AHPND research.
2.1. PirAvp (ORF 51) and PirBvp (ORF50)
The products of these two genes were named based on their sequence homology to a binary toxin family: Photorhabdus insect-related (Pir)-like proteins. As mentioned above, PirAvp and PirBvp have been confirmed as the major factors that cause damage to the hepatopancreatic tissues of shrimps, and this is considered the key symptom of AHPND [4,5,6,7,8,9,10]. Although some reports showed PirBvp alone has the ability to cause cell damage [5,6], both PirAvp and PirBvp are necessary to achieve full toxicity. The pirAvp and pirBvp genes are both part of the same operon [5], which means that they are regulated and expressed synchronously.
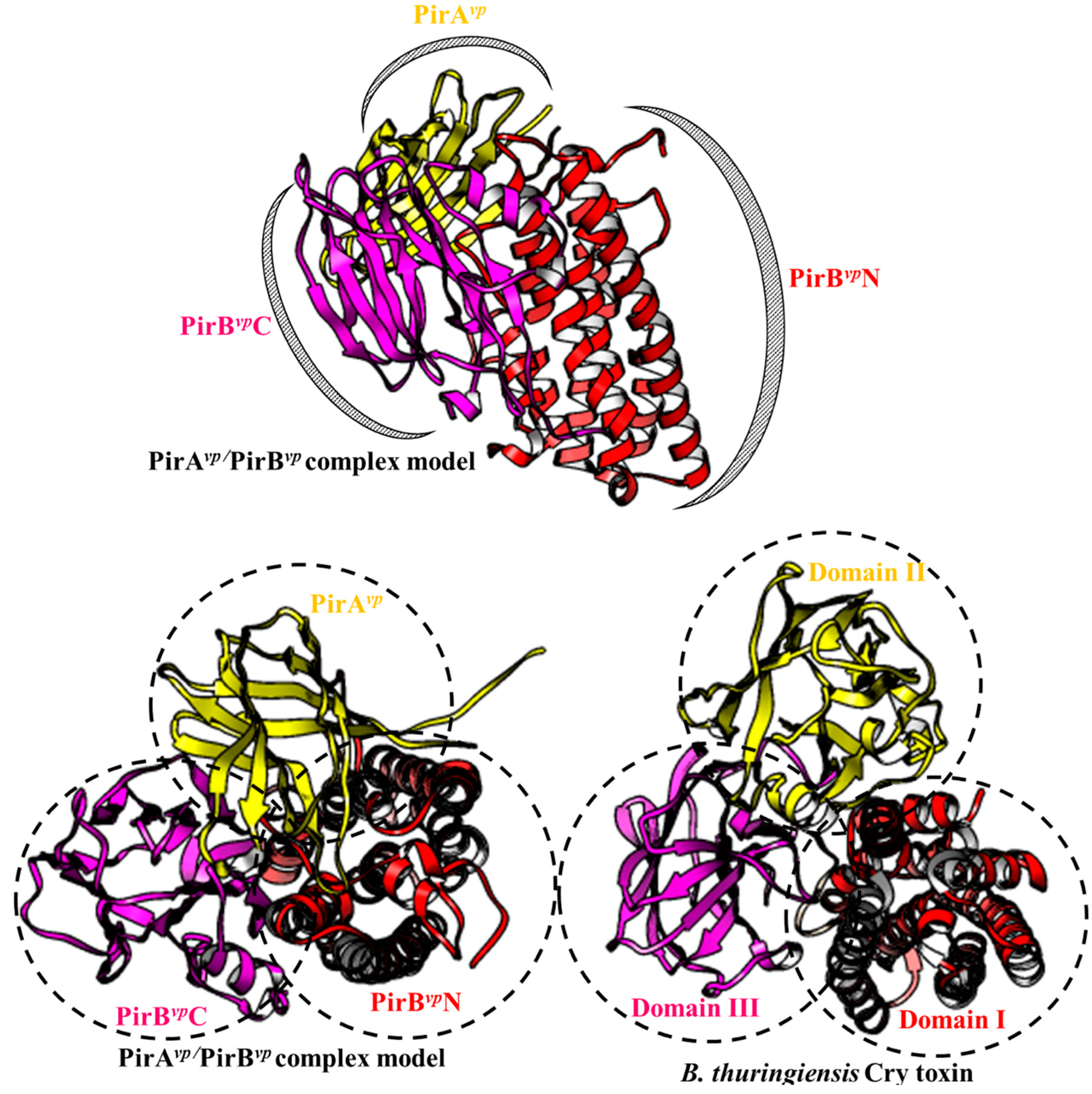
Structural analysis showed that there was a functional relationship between PirAvp/PirBvp and the Bacillus thuringiensis Cry toxin [5,9]. Functionally, Cry toxins act as pore-forming toxins and achieve cytotoxic effects via three domains: the N-terminal pore-forming domain I, the middle receptor binding domain II and C-terminal sugar/receptor binding domain III [43,44,45]. The typical cytotoxic mechanism of the Cry toxin is as follows: domain III first recognizes the N-Acetylgalactosamine (GalNAc) that is present on several receptors (e.g., aminopeptidase N, APN). Subsequently, domain II also binds to the recognized receptor. Then, the α1 helix of domain I is hydrolyzed, and this triggers the oligomerization of the Cry toxin to form pores on the cell membrane [43,44,45].
It was found that PirAvp and PirBvp have a Cry toxin-like folding [5,9]. The N-terminal and C-terminals domain of PirBvp (PirBvpN and PirBvpC) have a similar folding to the pore-forming domain I and receptor binding domain II of Cry toxin, respectively. The PirBvpN has an alpha-helical bundle structure, and this is called an inside-out membrane fold. The inside-out membrane fold consists of a hydrophobic α-helix surrounded by multiple amphipathic α-helices, and it is often found in other pore-forming toxins such as colicin and Cry toxins [9]. The structural characteristics of this domain allow it to switch between soluble (hydrophobic α-helix hides inside; inactivated) and transmembrane (hydrophobic α-helix exposed; activated and toxic) forms [9]. Meanwhile, the PirBvpC contains an antiparallel β-barrel jelly-roll topology. Despite the divergence of their amino acid sequences, PirBvpC and Cry domain II share a similar antiparallel β-barrel jelly-roll topology, suggesting that PirBvpC may also be involved in receptor binding. There is also a recent report which showed that alpha amylase-like protein, which is a 1,4-α-d-glucan glucanohydrolase targeted by the Cry toxin, has the ability to interact with PirBvp [46]. However, this result was based on far western and mass spectrometry analysis [46] and will need to be confirmed by other methods. Shrimp receptors for PirBvpC, thus still remain to be further investigated.
Like PirBvpC, PirAvp also has a jelly-roll topology. The structural analysis further shows that PirAvp has a substrate-binding pocket [9]. Although the precise targets of PirAvp have not yet been identified, the structural similarity of PirAvp to Cry domain III implies that these targets may include carbohydrates such as GalNAC [9].
Given their structural similarities, the cytotoxic activation of PirAvp/PirBvp should be similar to that of the Cry toxins. If so, then the first step for PirAvp/PirBvp would be to assemble into a Cry-like three-domain complex. In 2019, Lin et al. proposed binding models for PirAvp/PirBvp by using crosslinking based- and hydrogen-deuterium exchange-mass spectrometry. As shown in Figure 2, The PirAvp/PirBvpheterodimer model has all the functional domains that are necessary for its cytotoxicity: i.e., binding to the carbohydrate/receptor by PirAvp and PirBvpC would be followed by the creation of an uncontrollable pore by PirBvpN on the cell membrane to induce cell death. However, the binding affinity of PirAvp and PirBvp was found to be relatively low (~7 µM) in vitro [13], suggesting that PirAvp and PirBvp would not form a stable complex when the two proteins interact directly. It is possible, however, that the interaction with the receptor/sugar may improve the stability of the PirAvp/PirBvp complex. We also note that in contrast to the structural characteristics of PirAvp and PirBvpC discussed above, in this model, PirAvp and PirBvpC were matched respectively to the Cry receptor binding domain II and sugar-binding domain III (Figure 2 lower images). This suggests that the real functions of PirBvpC and PirAvp may be reversed. Interestingly, a recent report further supports this idea by finding that PirBvp, but not PirAvp, is able to target glycosaminoglycans like GalNH2 and GlcNH2 [47]. Clearly, more work needs to be done before we fully understand how these toxins, in fact, attach to the shrimp cell membrane.
2.2. Transposases (ORF15, ORF48, ORF55, ORF57 and ORF68)
DNA transposons are mobile DNA elements that can move from one DNA molecule to another. Transposases are a type of enzyme that can bind to the inverted sequences on both ends of a transposon, digest, and release the transposon, and promote translocation of this DNA fragment to another location on the genome [26,48]. This provides a mechanism to deliver genetic information into other chromosomes and confer new functions to a gene or replace a defective gene.
In pVA1, a total of five transposase genes were found (ORF15, 48, 55, 57, and 68). Apart from ORF57, which only contains a zinc-binding domain of the IS91 transposase family, the other four genes are complete transposases. ORF15, 48, and 55 share 100% sequence identity, and as members of the IS5 family of DDE transposases, they all contain a conserved DNA hydrolysis DDE (Asp-Asp-Glu) motif. The remaining transposase, ORF68, belongs to the Rpn family of recombination-promoting nuclease/putative transposases. All four of these transposases have the ability to trigger the transfer of DNA transposons.
In previous reports, it has been shown that the pirABvp genes can be deleted/transferred by gene transposition. For instance, in the pVA1 plasmid isolated from V. parahaemolyticus strain 3HP, it was found that the ORF49-54 gene cluster, which includes the pirABvp genes (ORF50 and ORF51) was flanked by two transposase genes in opposite directions (ORF 48 and ORF55) with inverted repeats in their terminals. It was also found that the gene cluster consisting of ORF 48-ORF54 was lost in the pVA1 type plasmid isolated from V. parahaemolyticus strain M2-36, leaving only ORF55 behind [5]. A gene cluster containing pirABvp has also been inserted into another AHPND-causing plasmid, pVH, isolated from V. owensii [49]. The same transposase genes with inverted repeats were also found up- and down-stream of this cluster [49]. Another report describes a pVA1 plasmid from a natural V. parahaemolyticus strain (XN87), which contains a mutated pirAvp gene that is interrupted by a transposon gene fragment [50]. This results in an out-of-frame insertion of pirAvp and further affects the downstream gene expression of pirBvp [50]. Furthermore, whole-genome sequencing of 40 V. parahaemolyticus isolated from shrimp hepatopancreas and aquaculture water in Malaysia indicated that pirABvp genes are prone to deletion [51]. Taken together, these results demonstrate the importance of this DNA transposon in the transfer, deletion, and mutation of the pirABvp-containing gene cluster. Screening other bacteria or plasmids for the presence of this gene cluster will also be useful for identifying potential AHPND-causing pathogens.
2.3. Conjugation Factors (ORF10, ORF11, ORF75, ORF76, ORF78, ORF79, ORF81, ORF82, ORF83, and ORF84)
Conjugation is a process used by bacteria to transfer their genetic material to a recipient bacterium. This horizontal gene transfer facilitates bacterial evolution, including the dissemination of antibiotic resistance genes [52]. During conjugation, a single-stranded DNA molecule is first produced, and the plasmid undergoes DNA replication and formation of the protein-DNA complex, which is processed by the DNA transfer and replication (Dtr) system. The coupling protein (CP) further delivers this protein-DNA intermediate to the trans-membrane channel, where it is actively secreted through the channel and the exocellular pili, which are produced by the mating pair formation (Mpf) system. The Mpf/CP conjugation system is a kind of type-IV secretion system (T4SS), and there are twelve components essential for the Mpf system, TraF, TrbB, TrbC, TrbD, TrbE, TrbF, TrbG, TrbH, TrbI, TrbJ, TrbK, and TrbL [30,31]. On the pVA1 plasmid, all of these genes have been found except for trbJ and trbK. In addition, ORF11 is annotated as a VirB1-like gene, which is an important component that facilitates the formation of type-IV secretion systems (T4SS) [32]. This mechanism is, therefore, available for pVA1-related plasmids or genes to use for transferring between different bacterial cells.
Recently, conjugation inhibitors were considered as a potential means of preventing the spread of antibiotic resistance genes among bacteria [53]. For example, in the type-IV secretion system, bacterial conjugation could be inhibited by using unsaturated fatty acids to block VirB11′s activity [54]. Another report also showed that two compounds isolated from medicinal plants, rottlerin [5,7-dihydroxy-2,2-dimethyl-6-(2,4,6-trihydroxy-3-methyl-5-acetylbenzyl)-8-cinnamoyl-1,2-chromene] and the red compound (8-cinnamoyl-5,7-dihydroxy-2,2,6-trimethylchromene), both have the ability to inhibit conjugal transfer of plasmids pKM101, TP114, pUB307, and R6K among Gram-positive bacteria [55]. It may, therefore, be worth investigating if any compounds are able to block the pVA1-derived conjugation system. By preventing the spread of pVA1, the damage produced by AHPND might be limited.
2.4. Antirestriction Proteins (ORF32 and ORF35)
Among different species of bacteria, conjugative plasmids play a crucial role in spreading a variety of genes between various bacterial species. However, the transfer of DNA in naturally occurring vectors is tightly regulated by the immigration system, or restriction-modification (R-M) system, of the host [56]. The bacterial R-M system acts as an immune system that attacks the foreign DNA in the cell [56]. To overcome this restriction barrier of the host, some donor plasmids encode antirestriction proteins such as Ard (alleviation of restriction of DNA). On the pVA1 plasmid, there are two putative antirestriction protein genes, ORF32 and ORF35, which respectively have deduced amino acid sequences with similarity to ArdB and ArdC.
Ard genes are commonly found in transposons and conjugative plasmids in various prokaryotes [57]. The Ard antirestriction proteins are of three types, ArdA, ArdB, and ArdC, all of which act as inhibitors of the type I R-M system [33,34,58,59,60,61]. ArdA and ArdB proteins are small and acidic, and they both include a small region of similarity that consists of 14 residues designated as the “antirestriction” domain [58]. However, while ArdA has a DNA-like shape and charge distribution, suggesting it acts as a DNA-mimic [59,62], ArdB proteins have a novel structural fold, and do not show any DNA-like properties [60]. Meanwhile, ArdC proteins can also bind to single-stranded DNA, and they have been observed to protect single-stranded but not double-stranded plasmid DNA against the activity of type II restriction endonuclease (HhaI) in vitro [34]. In their role as type I R-M system inhibitors, ArdB and ArdC presumably act to prevent pVA1 from being digested during plasmid transmission. Conversely, if their anti-restriction activity can be blocked, the transferred pVA1 plasmid should then become susceptible to attack by the bacterium’s type I R-M system. This is an issue that deserves further investigation.
2.5. Secretion System (ORF3, 86, 89, and 90)
In addition to the type IV secretion system mentioned above (i.e., ORF11, VirB1-like gene), four other ORFs (ORF3, 86, 89, and 90) belonging to type II and III secretion systems are also found on the pVA1 plasmid. V. parahaemolyticus has been reported to contain not only two type-III secretion systems (T3SS1 and T3SS2) and two type-VI secretion systems (T6SS1 and T6SS2) [63,64], but also two T2SSs and two T2/4SSs [15]. These secretion systems are all related to the virulence of V. parahaemolyticus. For example, T3SS1 contributes to the secretion of effector proteins that are cytotoxic to HeLa cells, and T3SS2 is related to the enterotoxicity in the rabbit ileal loop test [65]. T6SS1 has been reported as an antibacterial system and is associated with the AHPND-causative strains [66]. In addition, T6SSs also contribute to the adhesion of V. parahaemolyticus to host cells [67], while T2SS is commonly employed by Gram-negative bacteria to transport a variety of molecular cargos [68]. Since PirAvp and PirBvp are both secreted toxins [5], the pVA1 secretion systems may play an important role during toxin release.
The secretion systems of drug-resistant bacteria were considered as the drug targets [69,70]. For example, Acinetobacter baumannii T2SS is responsible for the secretion of multiple enzymes, and its inactivation reduces the in vivo fitness of A. baumannii, as well as increasing its sensitivity to the human complementary system [70]. Recently, several small molecules, such as salicylidene acyl hydrazides and N-Hydroxybenzimidazoles, have been confirmed as T3SS inhibitors [69]. Moreover, high-throughput screening (HTS) based on secreted lipase activity was also developed to identify small molecule inhibitors of the T2SS [70]. Similarly, a search for inhibitors of the secretion systems of AHPND-causing V. parahaemolyticus might also identify molecules with the potential for controlling the damage caused by PirABvp toxins.
2.6. pndA (ORF7)
A bacterial toxin-antitoxin system is a class of the plasmid addiction system (PAS) or post-segregational killing (PSK) system that is used to prevent the loss of plasmids from bacterial populations. For examples, in the hok-sok system of R1 plasmid, the srnB-srnC system of F plasmid and the pndA-pndB system of R438 plasmid, highly stable “killer” mRNAs that encode toxin proteins (Hok, SrnB, and PndA) are synthesized while small, unstable antisense RNAs (sok, srnC and pndB) are also produced. When coupled with host RNase II, these antisense RNAs facilitate the degradation of the killer mRNAs, and suppress the expression of toxin proteins [35]. However, after cell segregation, the production of antisense RNAs in the plasmid-free cells ceases, and due to their low stability, the levels of antisense RNAs rapidly decreases. Meanwhile, the killer mRNAs are still present. This results in the production of the encoded toxin proteins, each of which contains a single transmembrane domain, which is used to kill the plasmid-free cells from the inside [71,72]. The presence of a pndA gene (ORF7) in the pVA1 plasmid suggests that the PSK system is being used to ensure the presence of pVA1 in daughter cells. However, to date, the antisense RNA sequences for the pndA gene (i.e., pndB) have not been determined. Clearly, this will need to be further investigated to ascertain whether or not AHPND-causing bacteria actually include a viable pVA1-derived PSK system.
2.7. DNA Methyltransferase (ORF47 and ORF56)
DNA methylation is catalyzed by DNA methyltransferases (MTases, or methylases), which can transfer the methyl group from S-adenosyl methionine to the C-5 or N-4 positions of cytosine, or the N-6 position of adenine [73]. In bacteria, it is well known that DNA methylation protects the host’s own DNA from degradation by restriction-modification systems that are used to digest foreign DNA such as plasmids, transposons, and viral DNA [74,75]. In these cases, each MTase has its cognate restriction enzyme. However, some MTases (e.g., Dam and Dcm) do not have corresponding restriction enzymes, and instead of participating in restriction-modification systems, they are involved in the regulation of gene expression and virulence (for a review, please see [76]). ORF47 on the pVA1 plasmid is predicted to be an N-6 DNA MTase that is similar to Dam. Dam is essential for Vibrio cholerae virulence [36]. It also regulates the finP gene on the pSLT virulence plasmid of Salmonella and affects the formation of F-type pilus that is responsible for plasmid transfer via conjugation [77]. It should, therefore, be worth investigating whether this predicted N-6 DNA MTase of pVA1 works like Dam, and further affects the virulence of the plasmid-harboring bacteria.
3. Conclusion Remarks
By incorporating the pVA1 plasmid, Vibrio parahaemolyticus becomes an AHPND-causing pathogen that is fatal to shrimps. Although the major virulence of the pVA1 plasmid is due to the PirAvp/PirBvp toxins, there are still ~40 important ORFs with functional annotations (Table 1). The ORFs discussed here are involved in toxin secretion, gene transposition, plasmid maintenance, bacterial conjugation, and a post-segregational killing (PSK) system. As discussed above, some gene products, like the conjugation factor and the secretion system, could be potential drug targets for combatting AHPND. Other genes, such as those for anti-restriction proteins, should also be worth investigating for their functional roles in the spread of the AHPND-causing plasmid among Vibrio spps. Taken together, we hope the information reviewed here will be useful for suggesting new research avenues for increasing our understanding of the mechanism of this disease.
Author Contributions
Conceptualization, H.-C.W. (Hao-Ching Wang) and H.-C.W. (Han-Ching Wang); Writing—Original Draft Preparation, H.-C.W. (Hao-Ching Wang), S.-J.L., A.M. and R.K.; Writing—Review & Editing, H.-C.W. (Hao-Ching Wang) and H.-C.W. (Han-Ching Wang); Funding, H.-C.W. (Hao-Ching Wang) and H.-C.W. (Han-Ching Wang). All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported financially by the Ministry of Science and Technology (MOST106-2313-B-038-004-MY3, MOST108-2314-B-006-096-MY3, MOST109-2634-F-006-022) and University System of Taipei Joint Research Program (NTOU-TMU-109-01).
Acknowledgments
We are indebted to Paul Barlow for his helpful criticism.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shinn, A.P.; Pratoomyot, J.; Griffiths, D.; Trong, T.Q.; Vu, N.T.; Jiravanichpaisal, P.; Briggs, M. Asian shrimp production and economic costs of disease. Asian Fish. Sci. 2018, 31S, 29–58. [Google Scholar]
- Eshik, M.M.E.; Abedin, M.M.; Punom, N.J.; Begum, M.K.; Rahman, M.S. Molecular identification of AHPND positive Vibrio parahaemolyticus causing an outbreak in south-west shrimp farming regions of Bangladesh. J. Bangladesh Acad. Sci. 2017, 41, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Dhar, A.K.; Piamsomboon, P.; Caro, L.F.A.; Kanrar, S.; Adami, R.J.; Juan, Y.S. First report of acute hepatopancreatic necrosis disease (AHPND) occurring in the USA. Dis. Aquat. Org. 2019, 132, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Tran, L.; Nunan, L.; Redman, R.; Mohney, L.; Pantoja, C.; Fitzsimmons, K.; Lightner, D.V. Determination of the infectious nature of the agent of acute hepatopancreatic necrosis syndrome affecting penaeid shrimp. Dis. Aquat. Org. 2013, 105, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.T.; Chen, I.T.; Yang, Y.T.; Ko, T.P.; Huang, Y.T.; Huang, J.Y.; Huang, M.F.; Lin, S.J.; Chen, C.Y.; Lin, S.S.; et al. The opportunistic marine pathogen Vibrio parahaemolyticus becomes virulent by acquiring a plasmid that expresses a deadly toxin. Proc. Natl. Acad. Sci. USA 2015, 112, 10798–10803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, H.C.; Ng, T.H.; Ando, M.; Lee, C.T.; Chen, I.T.; Chuang, J.C.; Mavichak, R.; Chang, S.H.; Yeh, M.D.; Chiang, Y.A.; et al. Pathogenesis of acute hepatopancreatic necrosis disease (AHPND) in shrimp. Fish. Shellfish Immunol. 2015, 47, 1006–1014. [Google Scholar] [CrossRef] [PubMed]
- Tinwongger, S.; Nochiri, Y.; Thawonsuwan, J.; Nozaki, R.; Kondo, H.; Awasthi, S.P.; Hinenoya, A.; Yamasaki, S.; Hirono, I. Virulence of acute hepatopancreatic necrosis disease PirAB-like relies on secreted proteins not on gene copy number. J. Appl. Microbiol. 2016, 121, 1755–1765. [Google Scholar] [CrossRef]
- Theethakaew, C.; Nakamura, S.; Motooka, D.; Matsuda, S.; Kodama, T.; Chonsin, K.; Suthienkul, O.; Iida, T. Plasmid dynamics in Vibrio parahaemolyticus strains related to shrimp Acute Hepatopancreatic Necrosis Syndrome (AHPNS). Infect. Genet. Evol. 2017, 51, 211–218. [Google Scholar] [CrossRef]
- Lin, S.J.; Hsu, K.C.; Wang, H.C. Structural insights into the cytotoxic mechanism of Vibrio parahaemolyticus PirAvp and PirBvp toxins. Mar. Drugs. 2017, 15, 373. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Ng, T.H.; Wang, H.C. Acute hepatopancreatic necrosis disease in penaeid shrimp. Rev. Aquacult. 2020. [Google Scholar] [CrossRef] [Green Version]
- Letchumanan, V.; Chan, K.G.; Lee, L.H. Vibrio parahaemolyticus: A review on the pathogenesis, prevalence, and advance molecular identification techniques. Front. Microbiol. 2014, 5, 705. [Google Scholar] [CrossRef] [Green Version]
- Lightner, D.V.; Redman, R.M.; Pantoja, C.R.; Noble, B.L.; Tran, L. Early mortality syndrome affects shrimp in Asia. Glob. Aquac. Advocate 2012, 15, 40. [Google Scholar]
- Lin, S.J.; Chen, Y.F.; Hsu, K.C.; Chen, Y.L.; Ko, T.P.; Lo, C.F.; Wang, H.C.; Wang, H.C. Structural insights to the heterotetrameric interaction between the Vibrio parahaemolyticus PirAvp and PirBvp toxins and activation of the Cry-like pore-forming domain. Toxins 2019, 11, 233. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Ng, T.H.; Chang, C.C.; Lin, S.S.; Lo, C.F.; Wang, H.C. Bile acid and bile acid transporters are involved pathogenesis of acute hepatopancreatic necrosis disease in white shrimp Litopenaeus vannamei. Cel. Microbiol. 2019, 22, e13127. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Gil, B.; Soto-Rodriguez, S.; Lozano, R.; Betancourt-Lozano, M. Draft genome sequence of Vibrio parahaemolyticus strain M0605, which causes severe mortalities of shrimps in Mexico. Genome Announc. 2014, 2, e00055-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.T.; Chen, I.T.; Lee, C.T.; Chen, C.Y.; Lin, S.S.; Hor, L.I.; Tseng, T.C.; Huang, Y.T.; Sritunyalucksana, K.; Thitamadee, S.; et al. Draft genome sequences of four strains of Vibrio parahaemolyticus, three of which cause early mortality syndrome/acute hepatopancreatic necrosis disease in shrimp in China and Thailand. Genome Announc. 2014, 2, e00816-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, H.; Tinwongger, S.; Proespraiwong, P.; Mavichak, R.; Unajak, S.; Nozaki, R.; Hirono, I. Draft genome sequences of six strains of Vibrio parahaemolyticus isolated from early mortality syndrome/acute hepatopancreatic necrosis disease shrimp in Thailand. Genome Announc. 2014, 2, e00221-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Jimenez, S.; Noriega-Orozco, L.; Sotelo-Mundo, R.R.; Cantu-Robles, V.A.; Cobian-Guemes, A.G.; Cota-Verdugo, R.G.; Gamez-Alejo, L.A.; Del Pozo-Yauner, L.; Guevara-Hernandez, E.; Garcia-Orozco, K.D.; et al. High-quality draft genomes of two Vibrio parahaemolyticus strains aid in understanding acute hepatopancreatic necrosis disease of cultured shrimps in Mexico. Genome Announc. 2014, 2, e00800-14. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Tang, K.; Tran, L.; Lightner, D.V. Photorhabdus insect-related (Pir) toxin-like genes in a plasmid of Vibrio parahaemolyticus, the causative agent of acute hepatopancreatic necrosis disease (AHPND) of shrimp. Dis. Aquat. Org. 2015, 113, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Wang, H.; Xie, G.; Zou, P.; Guo, C.; Liang, Y.; Huang, J. An isolate of Vibrio campbellii carrying the pirVP gene causes acute hepatopancreatic necrosis disease. Emerg. Microbes Infect. 2017, 6, e2. [Google Scholar] [CrossRef] [Green Version]
- Kondo, H.; Van, P.T.; Dang, L.T.; Hirono, I. Draft genome sequence of non-Vibrio parahaemolyticus acute hepatopancreatic necrosis disease strain KC13.17.5, isolated from diseased shrimp in Vietnam. Genome Announc. 2015, 3, e00978-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Xiao, J.; Xia, X.; Pan, Y.; Yan, S.; Wang, Y. Draft genome sequence of Vibrio owensii strain SH-14, which causes shrimp acute hepatopancreatic necrosis disease. Genome Announc. 2015, 3, e01395-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Restrepo, L.; Bayot, B.; Arciniegas, S.; Bajaña, L.; Betancourt, I.; Panchana, F.; Reyes Muñoz, A. PirVPgenes causing AHPND identified in a new Vibrio species (Vibrio punensis) within the commensal Orientalis clade. Sci. Rep. 2018, 8, 13080. [Google Scholar] [CrossRef] [PubMed]
- Durán-Avelar, M.J.; Vázquez-Reyes, A.; González-Mercado, A.L.; Zambrano-Zaragoza, J.F.; Ayón-Pérez, M.F.; Agraz-Cibrián, J.M.; Gutiérrez-Franco, J.; Vibanco-Pérez, N. pirA- and pirB-like gene identification in Micrococcus luteus strains in Mexico. J. Fish. Dis. 2018, 41, 1667–1673. [Google Scholar] [CrossRef] [PubMed]
- Muthukrishnan, S.; Defoirdt, T.; Shariff, M.; Ina-Salwany, M.Y.; Yusoff, F.M.; Natrah, I. Horizontal gene transfer of the pirAB genes responsible for Acute Hepatopancreatic Necrosis Disease (AHPND) turns a non-Vibrio strain into an AHPND-positive pathogen. BioRxiv 2019. [Google Scholar] [CrossRef]
- Siguier, P.; Gourbeyre, E.; Chandler, M. Bacterial insertion sequences: Their genomic impact and diversity. FEMS Microbiol. Rev. 2014, 38, 865–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Kingston, A.W.; Ponkratz, C.; Raleigh, E.A. Rpn (YhgA-Like) proteins of Escherichia coli K-12 and their contribution to RecA-independent horizontal transfer. J. Bacteriol. 2017, 199, e00787-16. [Google Scholar] [CrossRef] [Green Version]
- Green, E.R.; Mecsas, J. Bacterial secretion systems: An overview. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Haase, J.; Lurz, R.; Grahn, A.M.; Bamford, D.H.; Lanka, E. Bacterial conjugation mediated by plasmid RP4: RSF1010 mobilization, donor-specific phage propagation, and pilus production require the same Tra2 core components of a proposed DNA transport complex. J. Bacteriol. 1995, 177, 4779–4791. [Google Scholar] [CrossRef] [Green Version]
- Schröder, G.; Lanka, E. The mating pair formation system of conjugative plasmids-A versatile secretion machinery for transfer of proteins and DNA. Plasmid 2005, 54, 1–25. [Google Scholar] [CrossRef]
- Höppner, C.; Liu, Z.; Domke, N.; Binns, A.N.; Baron, C. VirB1 orthologs from Brucella suis and pKM101 complement defects of the lytic transglycosylase required for efficient type IV secretion from Agrobacterium tumefaciens. J. Bacteriol. 2004, 186, 1415–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belogurov, A.A.; Delver, E.P.; Rodzevich, O.V. Plasmid pKM101 encodes two nonhomologous antirestriction proteins (ArdA and ArdB) whose expression is controlled by homologous regulatory sequences. J. Bacteriol. 1993, 175, 4843–4850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belogurov, A.A.; Delver, E.P.; Agafonova, O.V.; Belogurova, N.G.; Lee, L.Y.; Kado, C.I. Antirestriction protein Ard (Type C) encoded by IncW plasmid pSa has a high similarity to the “protein transport” domain of TraC1 primase of promiscuous plasmid RP4. J. Mol. Biol. 2000, 296, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, K.; Nielsen, A.; Thorsted, P.; Wagner, E.G. Mechanism of killer gene activation. Antisense RNA-dependent RNase III cleavage ensures rapid turn-over of the stable hok, srnB and pndA effector messenger RNAs. J. Mol. Biol. 1992, 226, 637–649. [Google Scholar] [CrossRef]
- Julio, S.M.; Heithoff, D.M.; Provenzano, D.; Klose, K.E.; Sinsheimer, R.L.; Low, D.A.; Mahan, M.J. DNA adenine methylase is essential for viability and plays a role in the pathogenesis of Yersinia pseudotuberculosis and Vibrio cholerae. Infect. Immun. 2001, 69, 7610–7615. [Google Scholar] [CrossRef] [Green Version]
- McLuskey, K.; Harrison, J.A.; Schuttelkopf, A.W.; Boxer, D.H.; Hunter, W.N. Insight into the role of Escherichia coli MobB in molybdenum cofactor biosynthesis based on the high resolution crystal structure. J. Biol. Chem. 2003, 278, 23706–23713. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.M.; Wang, G.; Pertsinidis, A.; Marians, K.J. Topoisomerase III acts at the replication fork to remove precatenanes. J. Bacteriol. 2019, 2019 201, e00563-18. [Google Scholar] [CrossRef] [Green Version]
- Lecompte, O.; Ripp, R.; Thierry, J.C.; Moras, D.; Poch, O. Comparative analysis of ribosomal proteins in complete genomes: An example of reductive evolution at the domain scale. Nucleic Acids Res. 2002, 30, 5382–5390. [Google Scholar] [CrossRef]
- Tang, Q.; Liu, Y.P.; Shan, H.H.; Tian, L.F.; Zhang, J.Z.; Yan, X.X. ATP-dependent conformational change in ABC-ATPase RecF serves as a switch in DNA repair. Sci. Rep. 2018, 8, 2127. [Google Scholar] [CrossRef]
- Desai, B.V.; Morrison, D.A. An unstable competence-induced protein, CoiA, promotes processing of donor DNA after uptake during genetic transformation in Streptococcus pneumoniae. J. Bacteriol. 2006, 188, 5177–5186. [Google Scholar] [CrossRef] [Green Version]
- Surtees, J.A.; Funnell, B.E. Plasmid and chromosome traffic control: How ParA and ParB drive partition. Curr. Top. Dev. Biol. 2003, 56, 145–180. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, J.L. Bivalent sequential binding model of a Bacillus thuringiensis toxin to gypsy moth aminopeptidase N receptor. J. Biol. Chem. 2000, 275, 14423–14431. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Wang, B.C.; Yu, Z.; Sun, M. Structural Insights into Bacillus thuringiensis Cry, Cyt and Parasporin Toxins. Toxins 2014, 6, 2732–2770. [Google Scholar] [CrossRef] [Green Version]
- Soberón, M.; Pardo, L.; Muñóz-Garay, C.; Sánchez, J.; Gómez, I.; Porta, H.; Bravo, A. Pore formation by Cry toxins. Adv. Exp. Med. Biol. 2010, 677, 127–142. [Google Scholar] [CrossRef] [PubMed]
- Almanza-Martínez, N.; Francisco Martínez Díaz, S.; Flores-Ramírez, G.; Zuñiga-Navarrete, F.; Gómez, I.; Cardona-Félix, C.S. An α-amylase-like protein interacts with PirB toxin from Vibrio parahaemolyticus in digestive tract tissue of white shrimp Litopenaeus vannamei. Aquac. Res. 2020, 1–5. [Google Scholar] [CrossRef]
- Victorio-De Los Santos, M.; Vibanco-Pérez, N.; Soto-Rodriguez, S.; Pereyra, A.; Zenteno, E.; Cano-Sánchez, P. The B subunit of PirABvp toxin secreted from Vibrio parahaemolyticus causing AHPND is an amino sugar specific lectin. Pathogens 2020, 9, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz-López, M.; García-Pérez, J.L. DNA transposons: Nature and applications in genomics. Curr. Genom. 2010, 11, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Liu, L.; Ke, Y.; Li, X.; Liu, Y.; Pan, Y.; Yan, S.; Wang, Y. Shrimp AHPND-causing plasmids encoding the PirAB toxins as mediated by pirAB-Tn903 are prevalent in various Vibrio species. Sci. Rep. 2017, 7, 42177. [Google Scholar] [CrossRef]
- Phiwsaiya, K.; Charoensapsri, W.; Taengphu, S.; Dong, H.T.; Sangsuriya, P.; Nguyen, G.T.T.; Pham, H.Q.; Amparyup, P.; Sritunyalucksana, K.; Taengchaiyaphum, S.; et al. A natural Vibrio parahaemolyticusΔpirAvp PirBvp+ mutant kills shrimp but produces no Pirvp toxins or AHPND lesions. Appl. Environ. Microbiol. 2017, 83, e00680-17. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.Z.Y.; Austin, C.M.; Ayub, Q.; Rahman, S.; Gan, H.M. Genomic characterization of Vibrio parahaemolyticus from Pacific white shrimp and rearing water in Malaysia reveals novel sequence types and structural variation in genomic regions containing the Photorhabdus insect-related (Pir) toxin-like genes. FEMS Microbiol. Lett. 2019, 366, fnz211. [Google Scholar] [CrossRef]
- Waters, V.L. Conjugative transfer in the dissemination of beta-lactam and aminoglycoside resistance. Front. Biosci. 1999, 4, D433–D456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabezón, E.; de la Cruz, F.; Arechaga, I. Conjugation inhibitors and their potential use to prevent dissemination of antibiotic resistance genes in bacteria. Front. Microbiol. 2017, 8, 2329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ripoll-Rozada, J.; Garcia-Cazorla, Y.; Getino, M.; Machon, C.; Sanabria-Rios, D.; de la Cruz, F.; Cabezón, E.; Arechaga, I. Type IV traffic ATPase TrwD as molecular target to inhibit bacterial conjugation. Mol. Microbiol. 2016, 100, 912–921. [Google Scholar] [CrossRef] [PubMed]
- Oyedemi, B.O.; Shinde, V.; Shinde, K.; Kakalou, D.; Stapleton, P.D.; Gibbons, S. Novel R-plasmid conjugal transfer inhibitory and antibacterial activities of phenolic compounds from Mallotus philippensis (Lam.) Mull. Arg. J. Glob. Antimicrob. Resist. 2016, 5, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Vasu, K.; Nagaraja, V. Diverse functions of restriction-modification systems in addition to cellular defense. Microbiol. Mol. Biol. Rev. 2013, 77, 53–72. [Google Scholar] [CrossRef] [Green Version]
- Wilkins, B.M. Plasmid promiscuity: Meeting the challenge of DNA immigration control. Environ. Microbiol. 2002, 4, 495–500. [Google Scholar] [CrossRef]
- Belogurov, A.A.; Delver, E.P. A motif conserved among the type I restriction-modification enzymes and antirestriction proteins: A possible basis for mechanism of action of plasmid-encoded antirestriction functions. Nucleic Acids Res. 1995, 23, 785–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMahon, S.A.; Roberts, G.A.; Johnson, K.A.; Cooper, L.P.; Liu, H.; White, J.H.; Carter, L.G.; Sanghvi, B.; Oke, M.; Walkinshaw, M.D.; et al. Extensive DNA mimicry by the ArdA anti-restriction protein and its role in the spread of antibiotic resistance. Nucleic Acids Res. 2009, 37, 4887–4897. [Google Scholar] [CrossRef]
- Serfiotis-Mitsa, D.; Herbert, A.P.; Roberts, G.A.; Soares, D.C.; White, J.H.; Blakely, G.W.; Uhrín, D.; Dryden, D.T.F. The structure of the KlcA and ArdB proteins reveals a novel fold and antirestriction activity against Type I DNA restriction systems in vivo but not in vitro. Nucleic Acids Res. 2010, 38, 1723–1737. [Google Scholar] [CrossRef] [Green Version]
- Balabanov, V.P.; Pustovoit, K.S.; Zavilgelsky, G.B. Comparative analysis of antirestriction activity of the ArdA and ArdB proteins encoded by genes of the R64 transmissible plasmid (IncI1). Mol. Biol. 2012, 46, 244–249. [Google Scholar] [CrossRef]
- Wang, H.C.; Ho, C.H.; Hsu, K.C.; Yang, J.M.; Wang, A.H.J. DNA mimic proteins: Functions, structures and bioinformatic analysis. Biochemistry 2014, 42, 1354–1364. [Google Scholar] [CrossRef] [Green Version]
- Henke, J.M.; Bassler, B.L. Quorum sensing regulates type III secretion in Vibrio harveyi and Vibrio parahaemolyticus. J. Bacteriol. 2004, 186, 3794–3805. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhou, D.; Mao, P.; Zhang, Y.; Hou, J.; Hu, Y.; Li, J.; Hou, S.; Yang, R.; Wang, R.; et al. Cell density- and quorum sensing-dependent expression of type VI secretion system 2 in Vibrio parahaemolyticus. PLoS ONE 2013, 8, e73363. [Google Scholar] [CrossRef] [Green Version]
- Park, K.S.; Ono, T.; Rokuda, M.; Jang, M.H.; Okada, K.; Iida, T.; Honda, T. Functional characterization of two type III secretion systems of Vibrio parahaemolyticus. Infect. Immun. 2004, 72, 6659–6665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Kinch, L.N.; Ray, A.; Dalia, A.B.; Cong, Q.; Nunan, L.M.; Camilli, A.; Grishin, N.V.; Salomon, D.; Orth, K. Acute hepatopancreatic necrosis disease-causing Vibrio parahaemolyticus strains maintain an antibacterial type VI secretion system with versatile effector repertoires. Appl. Environ. Microbiol. 2017, 83, e00737-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Yang, H.; Li, J.; Zhang, P.; Wu, B.; Zhu, B.; Zhang, Y.; Fang, W. Putative type VI secretion systems of Vibrio parahaemolyticus contribute to adhesion to cultured cell monolayers. Arch. Microbiol. 2012, 194, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Filloux, A. Protein secretion systems in Pseudomonas aeruginosa: An essay on diversity, evolution, and function. Front. Microbiol. 2011, 2, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, L.; Zhou, S.; Zhu, L.; Liang, C.; Chen, X. Small-molecule inhibitors of the type III secretion system. Molecules 2015, 20, 17659–17674. [Google Scholar] [CrossRef] [PubMed]
- Waack, U.; Johnson, T.L.; Chedid, K.; Xi, C.; Simmons, L.A.; Mobley, H.; Sandkvist, M. Targeting the type II secretion system: Development, optimization, and validation of a high-throughput screen for the identification of small molecule inhibitors. Front. Cell Infect. Microbiol. 2017, 7, 380. [Google Scholar] [CrossRef]
- Nielsen, A.K.; Thorsted, P.; Thisted, T.; Wagner, E.G.; Gerdes, K. The rifampicin-inducible genes srnB from F and pnd from R483 are regulated by antisense RNAs and mediate plasmid maintenance by killing of plasmid-free segregants. Mol. Microbiol. 1991, 5, 1961–1973. [Google Scholar] [CrossRef]
- Thisted, T.; Nielsen, A.K.; Gerdes, K. Mechanism of post-segregational killing: Translation of Hok, SrnB and Pnd mRNAs of plasmids R1, F and R483 is activated by 3′-end processing. EMBO J. 1994, 13, 1950–1959. [Google Scholar] [CrossRef]
- Palmer, B.R.; Marinus, M.G. The dam and dcm strains of Escherichia coli—A review. Gene 1994, 143, 1–12. [Google Scholar] [CrossRef]
- Kobayashi, I.; Nobusato, A.; Kobayashi-Takahashi, N.; Uchiyama, I. Shaping the genome--restriction-modification systems as mobile genetic elements. Curr. Opin. Genet. Dev. 1999, 9, 649–656. [Google Scholar] [CrossRef]
- Roberts, R.J.; Macelis, D. REBASE—restriction enzymes and methylases. Nucleic Acids Res. 2000, 28, 306–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Low, D.A.; Weyand, N.J.; Mahan, M.J. Roles of DNA adenine methylation in regulating bacterial gene expression and virulence. Infect. Immun. 2001, 69, 7197–7204. [Google Scholar] [CrossRef] [Green Version]
- Torreblanca, J.; Marqués, S.; Casadesús, J. Synthesis of FinP RNA by plasmids F and pSLT is regulated by DNA adenine methylation. Genetics 1999, 152, 31–45. [Google Scholar]
Figure 1.
Functional classifications of the pVA1 genes discussed in this review. All information in this figure (e.g., open-reading frames (ORF) number) refers to the plasmid pVA1 of the V. parahaemolyticus strain 3HP (NCBI accession number: KP324996).
Figure 1.
Functional classifications of the pVA1 genes discussed in this review. All information in this figure (e.g., open-reading frames (ORF) number) refers to the plasmid pVA1 of the V. parahaemolyticus strain 3HP (NCBI accession number: KP324996).

Figure 2.
PirAvp/PirBvp binding model (top) and a comparison between the PirAvp/PirBvp binding model and the Cry toxins (bottom). The PirAvp/PirBvp binding model is based on our previous report [13]. The PDB (Protein Data Bank) number for Cry toxin is 1CIY.
Figure 2.
PirAvp/PirBvp binding model (top) and a comparison between the PirAvp/PirBvp binding model and the Cry toxins (bottom). The PirAvp/PirBvp binding model is based on our previous report [13]. The PDB (Protein Data Bank) number for Cry toxin is 1CIY.


{kind=link}
{kind=link}
{kind=link}
Table 1.
The predicted open reading frames (ORFs) and their annotations.
| Pore Forming Toxins (2 †) | ||||
| ORF | Gene/Protein Name | Accession No. * | Location and Direction | Function Annotation # and Related References |
| ORF50 | PirBvp | AKC05670 | 33699←35015 | Photorhabdus insect-related (Pir) B; delta endotoxin, N-terminal domain [5,9] |
| ORF51 | PirAvp | AKC05671 | 35028←35363 | Photorhabdus insect-related (Pir) A [5,9] |
| Transposition (5 †) | ||||
| ORF | Gene/Protein Name | Accession No. * | Location and Direction | Function Annotation # |
| ORF15 | Transposase | AKC05635 | 9224→10144 | IS5 family transposase; DDE domain transposase [26] |
| ORF48 | Transposase | AKC05668 | 32035←32955 | IS5 family transposase; DDE domain transposase [26] |
| ORF55 | Transposase | AKC05675 | 36541→37461 | IS5 family transposase; DDE domain transposase [26] |
| ORF57 | Transposase | AKC05677 | 38476→38607 | Transposase zinc-binding domain; NCBI conserved domain: Zn_Tnp_IS91 (pfam14319) [27] |
| ORF68 | Transposase | AKC05688 | 48532→49476 | Rpn family recombination-promoting nuclease/putative transposase [28] |
| Secretion System (4 †) | ||||
| ORF | Gene/Protein Name | Accession No. * | Location and Direction | Function Annotation # |
| ORF3 | Type IV pilin protein | AKC05623 | 1052→2011 | Type II secretory pathway, pseudopilin PulG (Cell motility, Intracellular trafficking, secretion, and vesicular transport, Extracellular structures) [29] |
| ORF86 | Type II and III secretion system protein | AKC05706 | 64495→65997 | Pilus (MSHA, mannose-sensitive hemagglutinin type) biogenesis protein MshL; Bacterial type II and III secretion system protein [29] |
| ORF89 | Type II secretion protein | AKC05709 | 67759→69381 | Type II secretion protein; P-loop containing Nucleoside Triphosphate Hydrolases; Walker A motif; ATP binding site (chemical binding); Walker B motif [29] |
| ORF90 | Type II secretion protein F | AKC05710 | 69378→70382 | Type II secretion protein F [29] |
| Conjugation Factor (11 †) | ||||
| ORF | Gene/Protein Name | Accession No. * | Location and Direction | Function Annotation # |
| ORF10 | TraF | AKC05630 | 5999→6511 | Conjugative transfer signal peptidase TraF; Signal peptidase, peptidase S26 [30,31] |
| ORF11 | VirB1-like conjugal transfer protein | AKC05631 | 6525→7040 | Conjugal transfer protein; VirB1-like subfamily; one of twelve proteins making up type IV secretion systems (T4SS); VirB1 transglycosylase activity facilitate T4SS assembly [32] |
| ORF74 | TrbC | AKC05694 | 54020→54349 | Conjugal transfer protein TrbC; TrbC/VIRB2 family [30,31] |
| ORF75 | TrbD | AKC05695 | 54360→54674 | Mating pair formation protein TrbD; Type IV secretory pathway, VirB3-like protein [30,31] |
| ORF76 | TrbE | AKC05696 | 54715→57129 | Mating pair formation protein TrbE; conjugal transfer protein TrbE; Provisional; CagE, TrbE, VirB family, component of type IV transporter system; P-loop containing nucleoside triphosphate hydrolases [30,31] |
| ORF78 | TrbB | AKC05698 | 57895→58842 | Conjugal transfer protein TrbB; P-loop containing Nucleoside Triphosphate Hydrolases [30,31] |
| ORF79 | TrbL | AKC05699 | 58839→60149 | Conjugal transfer protein; TrbL/VirB6 plasmid conjugal transfer protein [30,31] |
| ORF81 | TrbF | AKC05701 | 60380→61054 | Conjugal transfer protein TrbF; VirB8 protein; putative dimerization motif (polypeptide binding) [30,31] |
| ORF82 | TrbG | AKC05702 | 61063→61935 | Conjugal transfer protein TrbG; VirB9/CagX/TrbG, a component of the type IV secretion system; VirB7 interaction site [30,31] |
| ORF83 | TrbH | AKC05703 | 61935→62357 | Conjugal transfer protein TrbH [30,31] |
| ORF84 | TrbI | AKC05704 | 62350→63606 | TrbI; conjugal transfer protein TrbI; Provisional; Bacterial conjugation TrbI-like protein [30,31] |
| Antirestriction Protein (2 †) | ||||
| ORF | Gene/Protein Name | Accession No. * | Location and Direction | Function Annotation # |
| ORF32 | Antirestriction protein (ArdB) | AKC05652 | 21587←22051 | Antirestriction protein; ArdB [33] |
| ORF35 | DNA primase; antirestriction protein (ArdC) | AKC05655 | 22595←23488 | DNA primase; antirestriction protein (ArdC) [34] |
| Post-Segregational Killing System(1 †) | ||||
| ORF | Gene/protein Name | Accession No. * | Location and Direction | Function Annotation # |
| ORF7 | Protein hokC /pndA | AKC05627 | 4292→4444 | Hok/gef family; Associated with a post-segregational killing (PSK) system [35] |
| DNA methyltransferase (2 †) | ||||
| ORF | Gene/Protein Name | Accession No. * | Location and Direction | Function Annotation # |
| ORF47 | N-6 DNA Methylase family protein | AKC05667 | 30063←32042 | N-6 DNA Methylase family protein [36] |
| ORF56 | Methylase family protein | AKC05676 | 37446←37976 | Methylase family protein [36] |
| Known Function (15 †) | ||||
| ORF | Gene/Protein Name | Accession No. * | Location and Direction | Function Annotation # |
| ORF5 | Trypsin protease | AKC05625 | 2553→3638 | Trypsin-like protease; NCBI conserved domain: Trypsin_2 (pfam13365) [27] |
| ORF21 | DNA-binding protein | AKC05641 | 12652→13059 | Domain in histone-like proteins of HNS family; NCBI conserved domain: Histone_HNS (pfam00816) [27] |
| ORF23 | MobB | AKC05643 | 14053→16020 | Molybdopterin-guanine dinucleotide biosynthesis protein MobB; P-loop containing nucleoside triphosphate hydrolases [37] |
| ORF26 | Mobilization protein | AKC05646 | 16788→17528 | Mobilization protein [37] |
| ORF27 | DNA topoisomerase III | AKC05647 | 17595→19805 | DNA topoisomerase III. DNA Topoisomerase, subtype IA; DNA-binding, ATP-binding and catalytic domain of bacterial DNA topoisomerases I and III, and eukaryotic DNA topoisomerase III and eubacterial and archael reverse gyrases [38] |
| ORF40 | DNA-binding protein | AKC05660 | 24974←25270 | Putative DNA-binding protein (no available reference) |
| ORF46 | SSU ribosomal protein S2p | AKC05666 | 28045←29535 | SSU (small subunit of ribosome) ribosomal protein S2p [39] |
| ORF53 | Exported protein | AKC05673 | 35657←36133 | Exported protein of unknown function [no available reference] |
| ORF61 | Recombinase RecF | AKC05681 | 40846→43179 | DNA repair exonuclease SbcCD ATPase subunit [Replication, recombination and repair] [40] |
| ORF62 | Phosphatidylinositol kinase | AKC05682 | 43568→44392 | Phosphatidylinositol kinase, CoiA-like protein [41] |
| ORF63 | Cold-shock protein | AKC05683 | 44809←45483 | Ribosomal protein S1-like RNA-binding domain. Found in a wide variety of RNA-associated proteins; NCBI conserved domain: CSP_CDS (cd04458) [27] |
| ORF64 | Alpha-helical coiled coil protein | AKC05684 | 45537←46523 | SMC_prok_B; chromosome segregation protein SMC, common bacterial type; NCBI conserved domains: KfrA_N (pfam11740) and CCDC158 (pfam15921) [27] |
| ORF66 | Tyrosine-tRNA ligase | AKC05686 | 47258←47578 | Putative tyrosine-tRNA ligase [no available reference] |
| ORF67 | ParA | AKC05687 | 47562←48239 | Chromosome partitioning protein ParA; plasmid-partitioning protein RepA [42] |
| ORF71 | Rep | AKC05691 | 51712←52551 | RNA polymerase Rpb2, domain 4; NCBI conserved domain: RNA_pol_Rpb2_4 (pfam04566) [27] |
† The number of each functional classification indicates how many pVA1 genes/proteins are classified into this type. *: Database source accession number: KP324996, #: Functional annotations were based on the NCBI Protein BLAST (blastp; https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE=Proteins&). This analysis was achieved on February 2020.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, H.-C.; Lin, S.-J.; Mohapatra, A.; Kumar, R.; Wang, H.-C. A Review of the Functional Annotations of Important Genes in the AHPND-Causing pVA1 Plasmid. Microorganisms 2020, 8, 996. https://doi.org/10.3390/microorganisms8070996
AMA Style
Wang H-C, Lin S-J, Mohapatra A, Kumar R, Wang H-C. A Review of the Functional Annotations of Important Genes in the AHPND-Causing pVA1 Plasmid. Microorganisms. 2020; 8(7):996. https://doi.org/10.3390/microorganisms8070996
Chicago/Turabian StyleWang, Hao-Ching, Shin-Jen Lin, Arpita Mohapatra, Ramya Kumar, and Han-Ching Wang. 2020. "A Review of the Functional Annotations of Important Genes in the AHPND-Causing pVA1 Plasmid" Microorganisms 8, no. 7: 996. https://doi.org/10.3390/microorganisms8070996
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.