1. Introduction
Over the past few decades, there has been a worldwide increase in reports of diseases affecting marine organisms of different taxa [
1]. Climate change is affecting the marine ecosystem, which is already subjected to many anthropogenic disturbances, such as overfishing, pollution and habitat destruction. Climate warming can increase pathogen development and their survival rates, disease transmission and host susceptibility [
2]. Environmental conditions play a crucial role not only in pathogen transmission, but also as risk factors for the occurrence of clinical diseases. Unlike mammals that regulate their internal environments, most fish are poikilotherms with little ability to regulate their core body temperature. In this situation, both the microbe and the host are physiologically tied to the environment they live in and have an optimal temperature range for survival. Extended periods outside the optimal range usually results in death [
3]. This is especially true in the Mediterranean Sea, which is one of the biggest reservoirs of biodiversity in the world [
4]. Diverse cold and tropical marine fauna combine and mix due to the basin’s oceanographic and biogeographical properties and, therefore, might serve as “miniature model” for the world’s oceans and provide insights into global patterns of marine ecosystems [
5].
Although aquaculture production has increased dramatically, fish consumption still largely depends on fisheries [
6]. Wild fish have an important ecological role in the ecosystem and economic role as a major protein source for humans [
7]. A wide range of marine pathogens from aquaculture is well documented, but there remains a lack of baseline data and information regarding pathogenic agents’ prevalence in the wild fish population [
8]. On the other hand, aquaculture is a fast-growing industry for the production of high protein-sourced foods, including fish production [
9]. This growth is accompanied by concerns from both the public and private sectors [
9], as fish production is commonly associated with serious environmental impacts (e.g., water pollution, pathogen transmission, and temperature changes). Mariculture production is usually as crowded and intensive production ponds. Fish are routinely treated and harvested from the walkway or raft around the cages, and boats are used for transportation of feed and for harvest [
10]. In Israel, there are several offshore marine farms along the Israeli Mediterranean coastline. Gilthead seabream (
Sparus aurata) is the dominant mariculture species, with an annual production in 2018 of 2450 tons, and 2310 tons in 2019 [
11]. Global aquaculture production has constantly increased during the last five decades [
12]. Although aquaculture in the Mediterranean is considered a relatively young industry, finfish diseases have been reported to cause substantial problems and mortalities among farmed stocks [
13]. Two apparent reasons that play a central role in the transmission of infectious pathogens are the farming activity, and the open design of Mediterranean aquaculture systems. As such, the transport of infected farmed fish from hatcheries, infected equipment, staff, and vessels, as well as through water currents, has been the main focus of fish health and biosecurity programs [
13]. Infectious agents are also common in marine waters. Bacteria and viruses can be transmitted both horizontally and vertically, due to oceanographic conditions in nearshore environments that are strongly influenced by local conditions [
14], and they can reduce commercial species’ growth and survivorship or decrease seafood quality. These impacts seem most problematic in the stressful and crowded conditions of aquaculture, which increasingly dominates seafood production as wild fishery production plateaus [
15]. It is often difficult to accurately estimate the impacts of diseases on wild populations, especially those of pelagic and subtidal fish species. However, there are a few quantitative data demonstrating that wild species near farms suffer more from infectious diseases than those in other areas. The movement of exotic infectious agents to new areas continues to be a great concern. It is important to note that the exposure of a host to a pathogen does not always result in infection, which, in turn, does not always lead to disease [
16]. Moreover, infection may occur geographically far from the immediate vicinity where infection is either detected or disease first becomes evident. These variables make diagnosis and/or pathogen evasion highly problematic.
Zoonotic microorganisms present a global public health concern. Those pathogens are known to cause protracted illness, especially in immuno-compromised individuals [
17]. As aquaculture production and the consumption of aquaculture products increase, the possibility of contracting zoonotic infections from either handling or ingesting these products also increases. Disease outbreaks are often related to management factors, such as the quality and quantity of nutrients in the water and high stocking density, which can increase bacterial loading on the external surface of the fish. As a result, diseased fish are more likely to transmit infection to humans [
18].
This research focused on two types of bacterial pathogens, namely
Vibrio and
Mycobacterium, which are known as zoonotic pathogens [
17] and are major causes of fish mortality [
19].
Vibrio species are Gram-negative curved rods that occur naturally in marine, estuarine, and freshwater systems worldwide. They occupy habitats ranging from the deep sea to shallow aquatic environments [
20]. More than 70 different
Vibrio species are known, 12 of which are recognized as human pathogens [
21]. Some species include human and animal pathogens capable of causing gastroenteritis, wound infections, cholera, and fatal septicemia [
22]. Infections caused by
Vibrio have been observed and documented in marine and estuarine-type fishes. The disease is known to have increased death rates (>50%) in fish farms soon after an outbreak [
23]. Fish vibriosis usually start with sluggishness and a loss of appetite. The disease may cause discoloration and eventual decay of the skin. Swollen sores may appear on the body, and in many cases protrude through the skin surface. Another symptom is redness around the fins and mouth. When the disease becomes systemic, it can cause exophthalmia, and the gut and rectum may be bloody and filled with fluid [
23].
Vibrio sp. cause some of the most significant infections of marine finfish. All marine fish are probably susceptible to at least one species.
Vibrio sp. have been infrequently isolated from freshwater aquarium fish and freshwater salmonids that have been fed marine offal [
24].
Vibrio sp. are typically facultative pathogens that can readily survive and multiply in the environment, although the relative pathogenicity of environmental versus fish isolates is uncertain.
Vibrio sp. are usually isolated from the mucosal surfaces and internal organs of clinically healthy fish. The highest environmental prevalence is in organically-polluted water and high salinity. A major predisposing risk factor for most types of fish-vibriosis is high water temperature. Crowding, organic pollution, and other stressors can also precipitate diseases outbreaks.
Vibrio strains also vary considerably in virulence, and some of them can cause disease without any predisposing stress. Some
Vibrio species produce hemolysins (which may cause anemia) and proteases (which may cause muscle damage) [
24].
Mycobacteria (Family
Mycobacteriaceae) are pleomorphic, Gram-positive, acid-fast, aerobic, non-motile rods [
17]. Fish mycobacteriosis is caused by non-tuberculous mycobacteria (NTM), and considered among the most chronic diseases occurring in aquatic animals [
25]. NTM is a group of more than 150 different species with distinct virulence features [
26]. In addition, fish mycobacteriosis is a chronic disease that has substantial economic consequences, as infections may significantly decrease production and trade. Some fish NTM are highly virulent and zoonotic [
17]. Typical signs of
Mycobacterium in fish are weight loss or emaciation, scale loss, ulcerations or hemorrhage along the body wall, granulomas, poor appetite and attitude, and often a history of reproductive problems [
27]. Both external and internal clinical signs caused by each pathogen are dependent on the host species, age of the fish, and stage of the disease (acute, chronic, and sub-clinic carrier); the signs are not always correlated or present at all [
28].
Mycobacterium marinum was first isolated in 1926 from several marine fish species [
24]; it is the most common etiological agent of fish mycobacteriosis [
24]. Other significant mycobacteria reported in marine fish are
Mycobacterium chelonae and
Mycobacterium fortuitum. NTM are saprophytes that reside in both soil and water, where they can survive for years. Commonly, gross examination of infected fish reveals grayish to white granulomatous nodules in various internal organs, mainly the spleen, liver, and kidney. However, in some cases, external signs may be present in the form of dermal ulceration [
24]. The objectives of this study were: (a) to evaluate the prevalence of
Vibrio sp. and
Mycobacterium sp. in wild and cultured marine fish species; (b) to compare pathogen prevalence between different years in farmed and wild fish; (c) to compare pathogen prevalence in different internal organs and maturity age; (d) to identify different strain variations of
Vibrio sp. and
Mycobacterium sp. in wild and cultured fish; and (e) to assess transmission of the pathogens from wild to cultured fish and vice versa.
4. Discussion
Vibrio sp. and
Mycobacterium sp. are widespread in nature, especially in the marine environment. Fish infected by these bacteria could be a source of zoonotic risk for human health [
26], and are known to cause infections in humans with different degrees of severity, especially in immunocompromised individuals [
22,
36]. Although
Vibrio sp. and
Mycobacterium sp. have been detected and studied in many fish species around the world, including various wild marine fish species [
21,
37,
38,
39,
40,
41], at the time of writing, this is the first study to identify
Vibrio sp. in the Lessepsian migrant species
Saurida lessepsianus and the endemic Mediterranean fish
Sardinella aurita. In this study, the presence of two important aquatic bacteria were investigated in four marine wild fish and one cultured farm fish in the eastern Mediterranean Sea. Both pathogens were detected in indigenous and Lessepsian species, and their prevalence varied greatly between fish species. In both pathogens, there were no statistically significant differences between the different wild species. However, the overall prevalence of
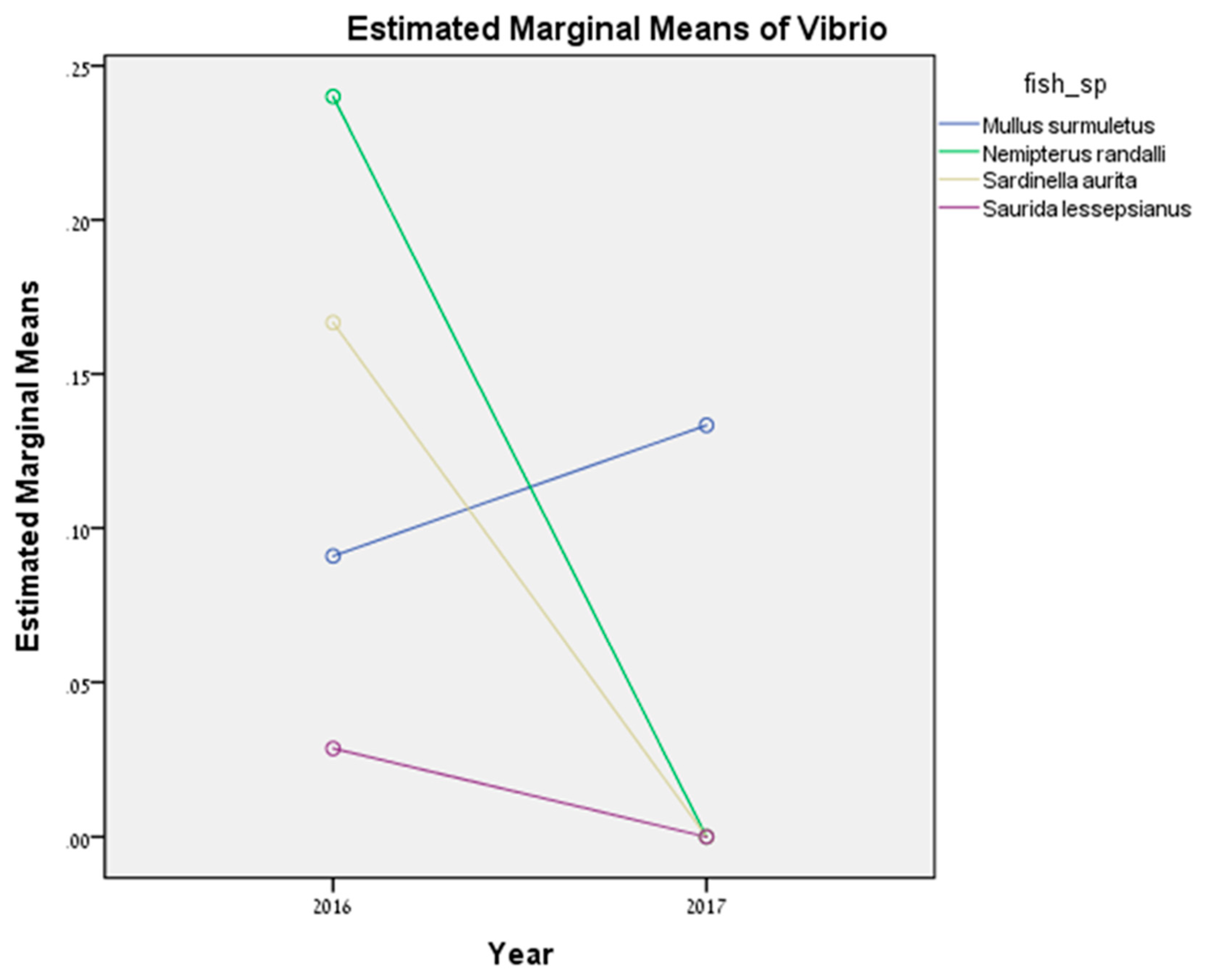
Vibrio sp. was significantly higher in 2016 compared to 2017.
Vibrio species are omnipresent and widely distributed in aquatic environments all over the world [
42]. The occurrence of
Vibrio sp. In fish, is commonly associated with elevated temperature, especially in temperate climes. Generally,
Vibrio species are detected in summer but are less common in winter, whereas the variations of
Vibrio sp. population size are lower in tropical and subtropical waters [
43]. Reports have shown a significant association between rising seawater temperature and an increase in the number of
Vibrio sp. infections, suggesting that global warming could be a factor in the emergence of vibriosis in temperate areas, due to its influence on resident bacterial communities [
43]. In 2016, during a continuous survey, most of the positive samples were detected in the summer period, which can explain the higher prevalence of
Vibrio sp. in 2016. However, most of the surveys in 2017 were between October and December, and the prevalence was lower. Within the cultured fish species, in 2018, the prevalence for
Mycobacterium sp. in
S. aurata was significantly higher compared to 2017, where no infection was detected at all. Due to logistical difficulties in field sampling (a seasonal ban on fishing vessels), we could not sample each season more than once for appropriate comparison. Thus, we cannot definitely determine if the prevalence of
Vibrio sp. is related to temporal changes in exposure or due to temperature shifts. Hence, this is a limitation of this study. The total prevalence for both pathogens was higher in the Lessepsian fish
N.
randalli and in the indigenous
M. surmuletus, with no reported clinical signs. High prevalence in an asymptomatic fish may indicate they can serve as carriers and horizontally infect other susceptible species living in proximity [
44].
N.
randalli is an invasive species, first reported in the Mediterranean in 2005 [
45], and has become a dominant fish species in the Israeli ichthyofauna within the past five years [
46]; the reasons for its successful establishment are unknown [
47]. Although
Vibrio sp. and
Mycobacterium sp. can be isolated from different organs (e.g., spleen and liver), it has been suggested that these pathogens have an affinity for, or are better detected, in kidney tissue [
48,
49]. This is in agreement with our results, where the prevalence for both pathogens in kidney was higher than in liver tissues in most of the examined fish species. In addition, the comparison of
Vibrio sp. and
Mycobacterium sp. prevalence in juvenile and adult fish was examined in four out of the five fish species (only mature
M. surmuletus were detected in this study and therefore were not included in the statistical analysis). In the wild fish, the highest prevalence was detected in adult-stage specimens. However, as for the cultured fish
S. aurata, positive results were obtained in both adult and juvenile specimens. It seems that at least the fish species tested in this study in different life stages are similarly susceptible for infection of
Vibrio sp. and
Mycobacterium sp. [
50].
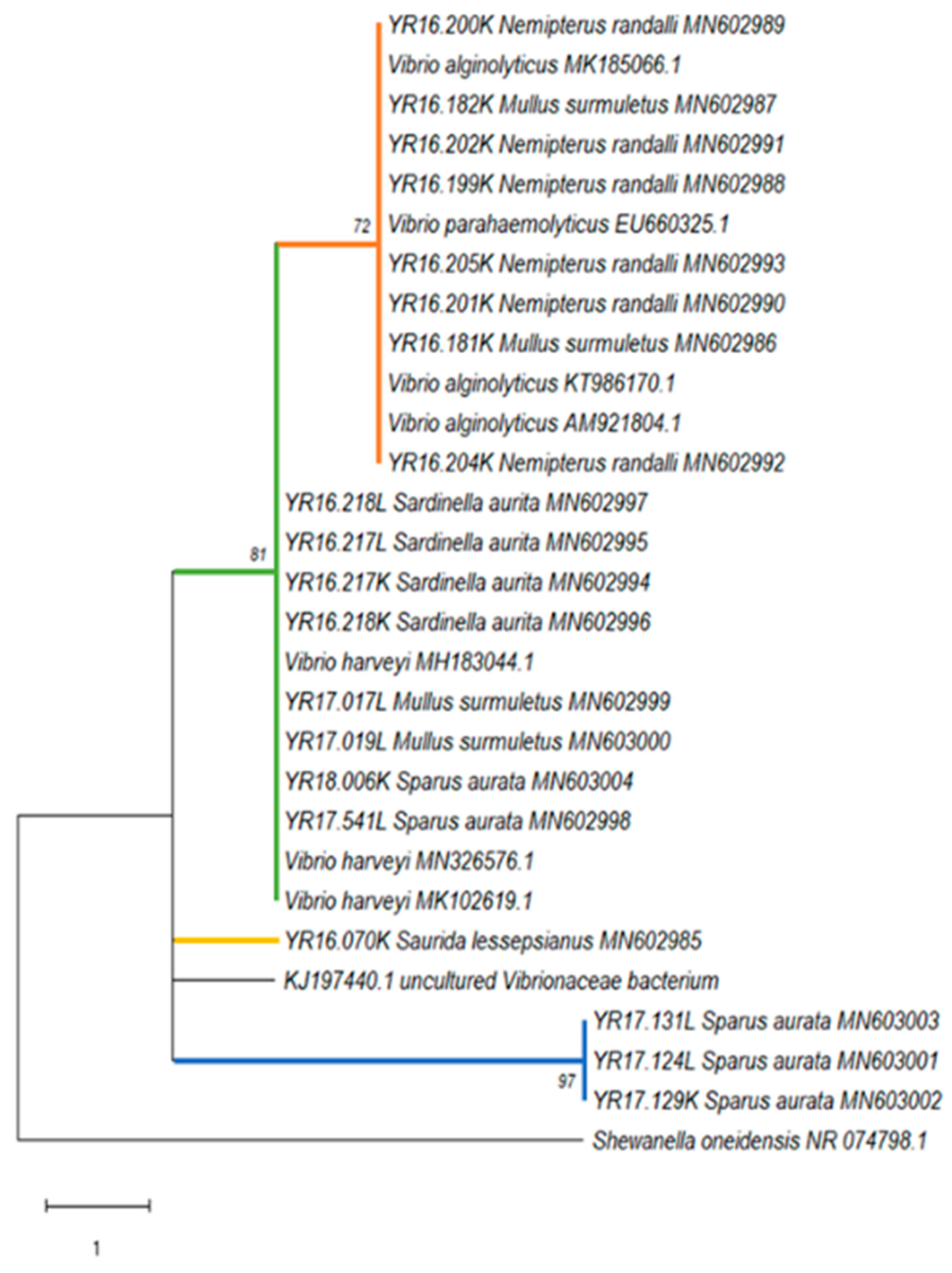
Phylogenetic analyses, based on the 16S rRNA gene sequences, revealed that all detected
Vibrio strains were divided into four different genogroups of
Vibrio sp., with an overlap in one group between the wild and the cultured species. This may suggest a spontaneous transmission between the wild and the farm fish. The first group showed high similarity to
V. parahaemolyticus and
V. alginolyticus. According to Montieri et al. [
35], these two
Vibrio species share nearly identical sequences in 16S rRNA gene (99.8% identity), and therefore this gene is inadequate for the separation between the two species. The second group showed high similarity to
V. harveyi. These species are known to be dangerous to humans, marine fish, and invertebrates and they can cause various diseases including vasculitis, gastroenteritis, septicemia, and skin infections [
51,
52,
53]. The third group contained only one strain that belonged to the Lessepsian fish
S. lessepsianus, which showed similarity to uncultured
Vibrionaceae bacterium isolated from pinfish (
Lagodon rhomboids) [
54], with a 95% homology. To the best of our knowledge, there were no records in the literature about the presence of
Vibrio sp. in this fish species. A possible explanation is that there is species-specific adaptation of some
Vibrio species [
55] and, therefore, this specific genotype might be a new
Vibrio genotype that is specific for
S. lessepsianus and possibly other marine fish. Further isolation and characterization of
Vibrio species is needed from this fish to understand its full genetic properties. In addition, the fourth group contained three identical strains all belonging to the cultured
S. aurata from 2017 without any similar references. It seems that this group has a unique insertion of 11 nucleotides which is common in 16S rRNA gene sequences, where insertion–deletion events are frequent and result in length differences among homologous sequences [
56]. The phylogenetic analysis of
Mycobacterium sp. reveals a clear separation between the wild and the cultured species: the wild species were positive across two main groups of
Mycobacterium sp. (with high similarity to
M. peregrinum and
M. neoaurum), while positive samples of the cultured fish
S. aurata were similar to
M. marinum. All these
Mycobacterium sp. are known as pathogenic species. Both
M. peregrinum and
M. neoaurum are fast growing mycobacteria that can cause bloodstream infections in immunocompromised hosts, and, unlike other NTM species [
57].
M. marinum is a recognized fish pathogen that can also infect endothermic organisms, including humans. In human infections,
M. marinum gains access to the body through skin abrasions and generally produces superficial and self-limiting lesions which involve the cooler parts of the body such as hands, forearms, elbows, and knees [
58,
59]. Although
M. marinum is a well-recognized pathogen of fish,
M. peregrinum has only recently been associated with diseases in fish [
60]. Even though mycobacteriosis cases have been reported previously in wild and cultured fish species [
39,
61], in this study, we documented Mycobacteria in
N. randalli for the first time. Over the last few decades, the 16S rRNA gene has emerged as a good standard for determining phylogenetic relationships of bacteria [
62]. By using PCR amplification and direct sequencing of 16S rRNA products, Knibb et al. [
63] identified
M. marinum directly from infected fish. This has allowed both proper taxonomic assignment and has opened the way to molecular epidemiologic analysis at the same time. However, even though this gene is still considered a key standard for bacterial identification [
64,
65], as more sequence information has accumulated over time, it has become evident that the resolution power of 16S rRNA sequences alone is often insufficient when closely related organisms are compared [
66]. Furthermore, Palys et al. [
66] suggested that protein-encoding genes may be more discriminative than those encoding rRNA, while the analysis of two or more unlinked loci would prevent bacterial misclassification due to possible homologous recombination with other taxa. Therefore, further molecular analyses are needed to understand the epidemiology and pathogenicity of the
Vibrio sp. and
Mycobacterium sp. identified in this study, both on fish and humans.
In the summer of 2018, there was an outbreak of
M. marinum in three out of the seven fish cages examined. The death rate ranged 5.7–13.5%, and some of the fish that survived showed clinical signs for the pathogen or no signs at all.
M. marinum was isolated from the kidney and spleen of both juvenile and adult fish. There was no evidence of the disease in 2017. A possible explanation is that in 2017 each cage was populated with ~350,000 fish, in contrast to 2018 where each cage was populated with ~470,000 fish. These conditions might have led to the disease outbreaks. As farmed fish are monitored regularly, they can be used as sentinels to evaluate pathogen exposure in the aquatic environment. However, this approach has utility only if farmed fish are susceptible to the pathogen and enter the marine environment free of the pathogen of interest. In addition, occurrence of disease in farmed fish populations does not necessarily imply occurrence of the same disease in wild populations. It is difficult to evaluate the health effect of escapees on the ecosystem without taking into consideration the qualitative aspects of wild fish assemblages around farms. Cross-contagion between farmed and wild fish species with shared pathogens may occur [
67] either through movements of individual fish or through species-specific migrations [
68,
69]. Connection among farms and other marine areas of interest through wild fish movements have been demonstrated both in Norwegian [
70] and in Mediterranean fish farms [
67]. Unlike parasitic pathogens, bacteria seem to exhibit higher potential to spread between wild and farmed fish. This is likely because the ecological barriers that exist for parasite transfer do not represent a great obstacle for bacteria [
13]. Firstly, bacteria are almost always present on the skin surface of fish. Secondly, bacterial diseases are usually treated by non-professional staff at the farms, and consequently involve increased risk of developing resistance and more pathogenic strains. Finally, bacteria are often generalists and do not need wild conspecifics to spread from farmed fish [
13]. There is a potential risk of pathogen transmission through movements of escaped and wild fish in Mediterranean fish farming areas, but actual transmission has been documented only in a handful of cases [
71]. Due to technical or operational malfunctions, infected farmed fish may escape and, in theory, spread pathogens to other cages/farms and wild fish as well. Furthermore, infected wild fish might also transfer pathogens to the farmed fish [
72]. This co-infection process leads to a large variety of shared pathogens among wild and farmed fish, while the various pathways of pathogen transmission increase the potential for infection and render epidemiological risk management difficult [
71]. Further research on molecular mechanisms of disease transmission in aquaculture and marine environments, as well more holistic analyses of pathogenic events in the Mediterranean Sea, are needed to clarify the potential of transmission of pathogens from aquaculture to the marine ecosystems.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}