Evidence for Common Horizontal Transmission of Wolbachia among Ants and Ant Crickets: Kleptoparasitism Added to the List

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Collection, DNA Extraction and PCR Conditions
2.2. Sequence Alignment and Phylogenetic Analyses
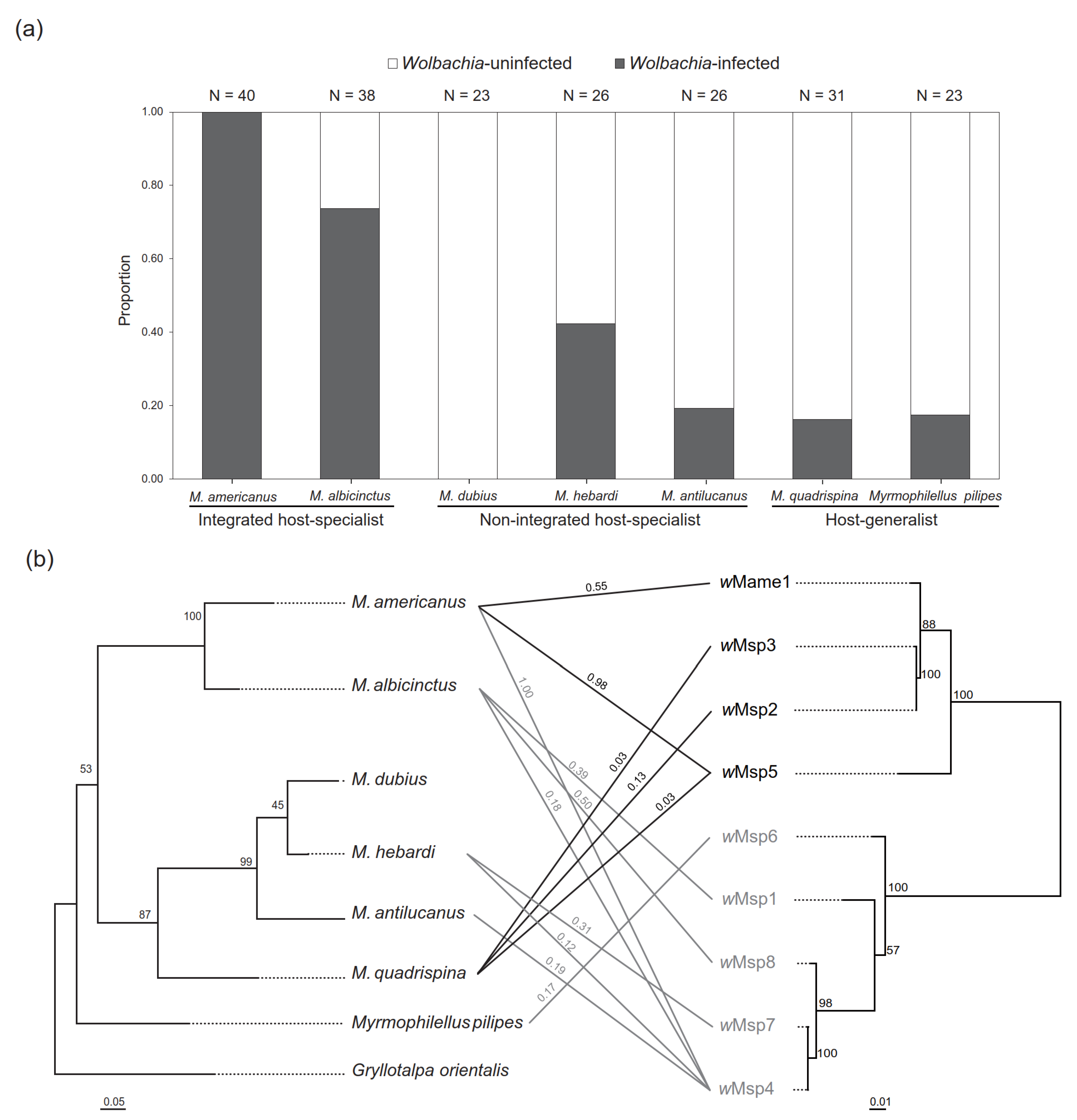
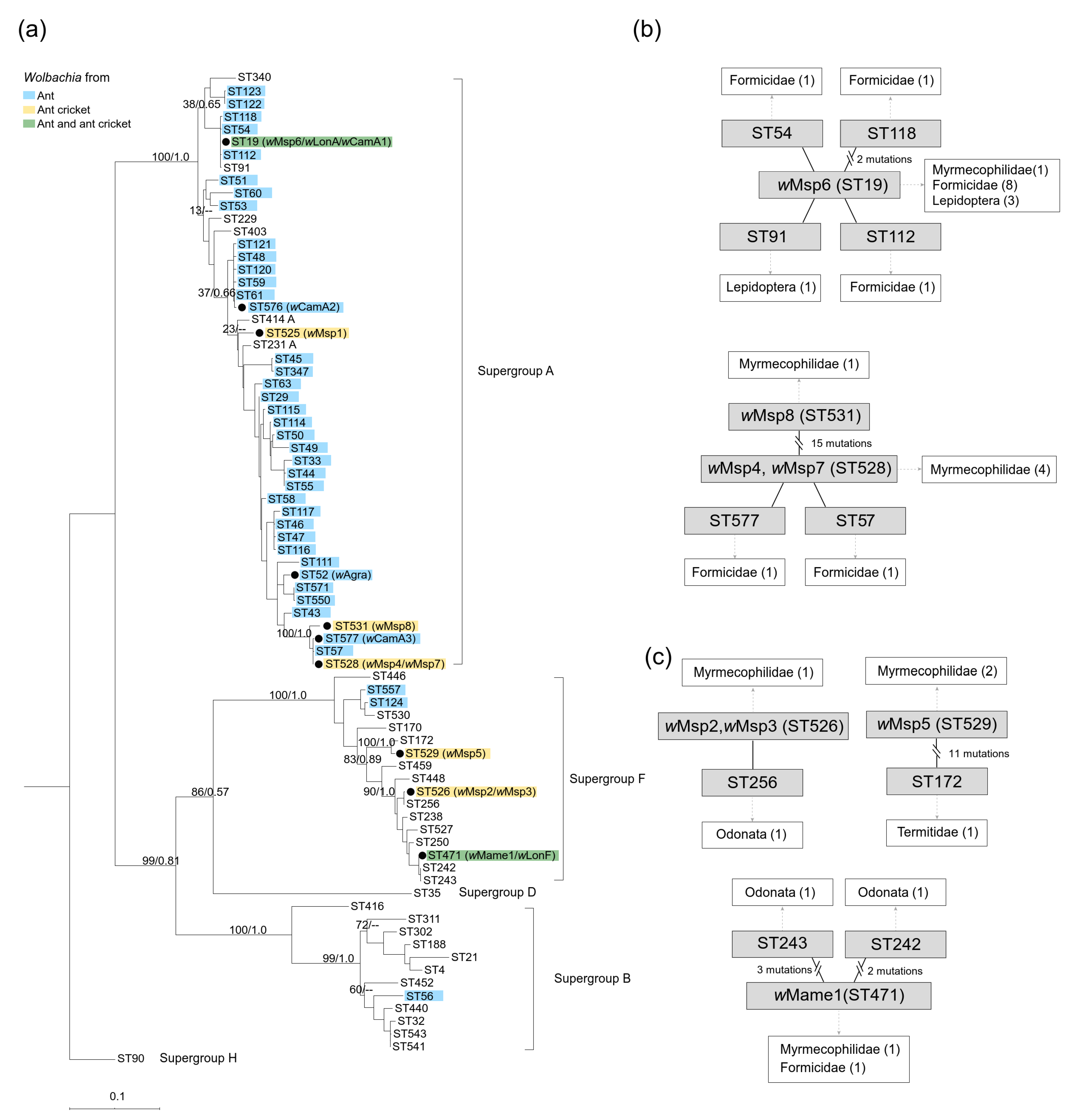
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zug, R.; Hammerstein, P. Still a host of hosts for Wolbachia: Analysis of recent data suggests that 40% of terrestrial arthropod species are infected. PLoS ONE 2012, 7, e38544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinert, L.A.; Araujo-Jnr, E.V.; Ahmed, M.Z.; Welch, J.J. The incidence of bacterial endosymbionts in terrestrial arthropods. Proc. R. Soc. B Biol. Sci. 2015, 282, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Baldo, L.; Hotopp, J.C.D.; Jolley, K.A.; Bordenstein, S.R.; Biber, S.A.; Choudhury, R.R.; Hayashi, C.; Maiden, M.C.J.; Tettelin, H.; Werren, J.H. Multilocus sequence typing system for the endosymbiont Wolbachia pipientis. Appl. Environ. Microbiol. 2006, 72, 7098–7110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldo, L.; Ayoub, N.A.; Hayashi, C.Y.; Russell, J.A.; Stahlhut, J.K.; Werren, J.H. Insight into the routes of Wolbachia invasion: High levels of horizontal transfer in the spider genus Agelenopsis revealed by Wolbachia strain and mitochondrial DNA diversity. Mol. Ecol. 2008, 17, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Raychoudhury, R.; Baldo, L.; Oliveira, D.C.S.G.; Werren, J.H. Modes of acquisition of Wolbachia: Horizontal transfer; hybrid introgression; and codivergence in the Nasonia species complex. Evolution 2009, 63, 165–183. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.Z.; Breinholt, J.W.; Kawahara, A.Y. Evidence for common horizontal transmission of Wolbachia among butterflies and moths. BMC Evol. Biol. 2016, 16, 118. [Google Scholar] [CrossRef] [Green Version]
- Bailly-bechet, M.; Martins-sim, P.; Szo, G.J.; Mialdea, G.; Charlat, S.; Evolutive, B. How long does Wolbachia remain on board? Mol. Biol. Evol. 2017, 34, 1183–1193. [Google Scholar] [CrossRef] [Green Version]
- Vavre, F.; Fouillet, P.; Fleury, F. Between and within host species selection on cytoplasmic incompatibility-inducing Wolbachia in haplodiploids. Evolution 2003, 57, 421–427. [Google Scholar] [CrossRef]
- Boivin, T.; Henri, H.; Vavre, F.; Gidoin, C.; Veber, P.; Candau, J.N.; Magnoux, E.; Roques, A.; Auger-Rozenberg, M.A. Epidemiology of asexuality induced by the endosymbiotic Wolbachia across phytophagous wasp species: Host plant specialization matters. Mol. Ecol. 2014, 23, 2362–2375. [Google Scholar] [CrossRef] [Green Version]
- Stahlhut, J.K.; Desjardins, C.A.; Clark, M.E.; Baldo, L.; Russell, J.A.; Werren, J.H.; Jaenike, J. The mushroom habitat as an ecological arena for global exchange of Wolbachia. Mol. Ecol. 2010, 19, 1940–1952. [Google Scholar] [CrossRef]
- Heath, B.D.; Butcher, R.D.; Whitfield, W.G.; Hubbard, S.F. Horizontal transfer of Wolbachia between phylogenetically distant insect species by a naturally occurring mechanism. Curr. Biol. 1999, 9, 313–316. [Google Scholar] [CrossRef] [Green Version]
- Hoy, M.A.; Jeyaprakash, A. Microbial diversity in the predatory mite Metaseiulus occidentalis (Acari: Phytoseiidae) and its prey; Tetranychus urticae (Acari: Tetranychidae). Biol. Control. 2005, 32, 427–441. [Google Scholar] [CrossRef]
- Huigens, M.E.; De Almeida, R.P.; Boons, P.A.H.; Luck, R.F.; Stouthamer, R. Natural interspecific and intraspecific horizontal transfer of parthenogenesis-inducing Wolbachia in Trichogramma wasps. Proc. R. Soc. B Biol. Sci. 2004, 271, 509–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kittayapong, P.; Jamnongluk, W.; Thipaksorn, A.; Milne, J.R.; Sindhusake, C. Wolbachia infection complexity among insects in the tropical rice-field community. Mol. Ecol. 2003, 12, 1049–1060. [Google Scholar] [CrossRef]
- LeClec’h, W.; Chevalier, F.D.; Genty, L.; Bertaux, J.; Bouchon, D.; Sicard, M. Cannibalism and predation as paths for horizontal passage of Wolbachia between terrestrial isopods. PLoS ONE 2013, 8, e60232. [Google Scholar] [CrossRef]
- Sintupachee, S.; Milne, J.R.; Poonchaisri, S.; Baimai, V.; Kittayapong, P. Closely related Wolbachia strains within the pumpkin arthropod community and the potential for horizontal transmission via the plant. Microb. Ecol. 2006, 51, 294–301. [Google Scholar] [CrossRef]
- Dedeine, F.; Ahrens, M.; Calcaterra, L.; Shoemaker, D.D. Social parasitism in fire ants (Solenopsis spp.): A potential mechanism for interspecies transfer of Wolbachia. Mol. Ecol. 2005, 14, 1543–1548. [Google Scholar] [CrossRef]
- Tolley, S.J.A.; Nonacs, P.; Sapountzis, P. Wolbachia horizontal transmission events in ants: What do we know and what can we learn? Front. Microbiol. 2019, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- VanBorm, S.; Wenseleers, T.; Billen, J.; Boomsma, J. Cloning and sequencing of wsp encoding gene fragments reveals a diversity of co-infecting Wolbachia strains in Acromyrmex leafcutter ants. Mol. Phylogenet. Evol. 2003, 26, 102–109. [Google Scholar] [CrossRef]
- Kronauer, D.J.C.; Pierce, N.E. Myrmecophiles. Curr. Biol. 2011, 21, 208–209. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, T.; Maruyama, M.; Itino, T. Behavioral differences between two ant cricket species in nansei islands: Host-specialist versus host-generalist. Insectes Soc. 2009, 56, 389–396. [Google Scholar] [CrossRef]
- Komatsu, T.; Maruyama, M.; Itino, T. Nonintegrated host association of Myrmecophilus tetramorii; a specialist myrmecophilous ant cricket (Orthoptera: Myrmecophilidae). Psyche 2013, 568536. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.W.; Hugel, S.; Wetterer, J.K.; Tseng, S.P.; Ooi, C.S.M.; Lee, Y.; Yang, C.C.S. Ant crickets (Orthoptera: Myrmecophilidae) associated with the invasive yellow crazy ant Anoplolepis gracilipes (Hymenoptera: Formicidae): Evidence for cryptic species and potential co-introduction with hosts. Myrmecol. News 2020, 30, 103–129. [Google Scholar] [CrossRef]
- Wetterer, J.K.; Hugel, S. Worldwide spread of the ant cricket Myrmecophilus americanus; a symbiont of the longhorn crazy ant; Paratrechina longicornis. Sociobiology 2008, 52, 157–165. [Google Scholar]
- Komatsu, T.; Maruyama, M. Additional records of the distribution and host ant species for the ant cricket Myrmophilellus pilipes. Insectes Soc. 2016, 63, 623–627. [Google Scholar] [CrossRef]
- Tseng, S.P.; Wetterer, J.K.; Suarez, A.V.; Lee, C.Y.; Yoshimura, T.; Shoemaker, D.; Yang, C.C.S. Genetic diversity and Wolbachia infection patterns in a globally distributed invasive ant. Front. Genet. 2019, 10, 838. [Google Scholar] [CrossRef]
- Penn, O.; Privman, E.; Ashkenazy, H.; Landan, G.; Graur, D.; Pupko, T. GUIDANCE: A web server for assessing alignment confidence scores. Nucleic Acids Res. 2010, 38, W23–W28. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Kozlov, A.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast; scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [Green Version]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. PartitionFinder: Combined Selection of Partitioning Schemes and Substitution Models for Phylogenetic Analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [Green Version]
- Didelot, X.; Falush, D. Inference of bacterial microevolution using multilocus sequence data. Genetics 2007, 175, 1251–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, J.A.; Goldman-Huertas, B.; Moreau, C.S.; Baldo, L.; Stahlhut, J.K.; Werren, J.H.; Pierce, N.E. Specialization and geographic isolation among Wolbachia symbionts from ants and lycaenid butterflies. Evolution 2009, 63, 624–640. [Google Scholar] [CrossRef]
- Komatsu, T.; Maruyama, M.; Itino, T. Differences in host specificity and behavior of two ant cricket species (Orthoptera: Myrmecophilidae) in Honshu; Japan. J. Entomol. Sci. 2010, 45, 227–238. [Google Scholar] [CrossRef]
- Komatsu, T.; Maruyama, M.; Hattori, M.; Itino, T. Morphological characteristics reflect food sources and degree of host ant specificity in four Myrmecophilus crickets. Insectes Soc. 2018, 65, 47–57. [Google Scholar] [CrossRef]
- Henderson, G.; Akre, R.D. Biology of the myrmecophilous cricket; Myrmecophila manni (Orthoptera: Gryllidae). J. Kansas Entomol. Soc. 1986, 59, 454–467. [Google Scholar]
- Hölldobler, B.; Wilson, E.O. The Ants; Harvard University Press: Cambridge, MA, USA, 1990; pp. 512–515. [Google Scholar]
- Treanor, D.; Hughes., W.O. Limited female dispersal predicts the incidence of Wolbachia across ants (Hymenoptera: Formicidae). J. Evol. Biol. 2019, 32, 1163–1170. [Google Scholar] [CrossRef]
- Shoemaker, D.D.; Ross, K.G.; Keller, L.; Vargo, E.L.; Werren, J.H. Wolbachia infections in native and introduced populations of fire ants (Solenopsis spp.). Insect Mol. Biol. 2000, 9, 661–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldo, L.; Werren, J.H. Revisiting Wolbachia supergroup typing based on WSP: Spurious lineages and discordance with MLST. Curr. Microbiol. 2007, 55, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Baldo, L.; Desjardins, C.A.; Russell, J.A.; Stahlhut, J.K.; Werren, J.H. Accelerated microevolution in an outer membrane protein (OMP) of the intracellular bacteria Wolbachia. BMC Evol. Biol. 2010, 10, 48. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Strain | Host Type | Host Species | wsp Accession No. | MLST id | ST a | Supergroup | gatB | coxA | hcpA | ftsZ | fbpA |
|---|---|---|---|---|---|---|---|---|---|---|---|
| wMsp1 | Ant cricket | M. albicinctus | MK995471 | 1926 | 525 | A | 294 | 291 | 331 | 251 | 459 |
| wMsp2 | Ant cricket | M. quadrispina | MK995472 | 1927 | 526 | F | 170 | 147 | 178 | 252 | 125 |
| wMsp3 | Ant cricket | M. quadrispina | MK995473 | 1928 | 526 | F | 170 | 147 | 178 | 252 | 125 |
| wMsp4 | Ant cricket | M. americanus, M. albicinctus, M. hebardi, M. antilucanus | MK995474 | 1929 | 528 | A | 49 | 44 | 297 | 42 | 49 |
| wMsp5 | Ant cricket | M. americanus, M. quadrispina | MK995475 | 1930 | 529 | F | 295 | 94 | 332 | 85 | 460 |
| wMsp6 | Ant cricket | Myrmophilellus pilipes | MK995476 | 1931 | 19 | A | 7 | 6 | 7 | 3 | 8 |
| wMsp7 | Ant cricket | M. hebardi | MK995477 | 1932 | 528 | A | 49 | 44 | 297 | 42 | 49 |
| wMsp8 | Ant cricket | M. albicinctus | MK995478 | 1933 | 531 | A | 49 | 44 | 333 | 253 | 49 |
| wMame1 | Ant cricket | M. americanus | MK995479 | 2024 | 471 a | F | 168 | 147 | 262 | 132 | 226 |
| wMame2 | Ant cricket | M. americanus | MK995480 | NA | NA | NA | NA | NA | NA | NA | NA |
| wLonA | Ant | Paratrechina longicornis | MN927214 | 1827 | 19 | A | 7 | 6 | 7 | 3 | 8 |
| wLonF | Ant | Paratrechina longicornis | MN927215 | 1828 | 471 | F | 168 | 147 | 262 | 132 | 226 |
| wAgra | Ant | Anoplolepis gracilipes | MN927216 | 2017 | 52 | A | 22 | 2 | 51 | 32 | 36 |
| wCamA1 | Ant | Camponotus sp. | MN927217 | 2021 | 19 b | A | 7 | 6 | 7 | 3 | 8 |
| wCamA2 | Ant | Camponotus sp. | MN927218 | 2022 | 576 b | A | 323 | 2 | 47 | 45 | 481 |
| wCamA3 | Ant | Camponotus sp. | MN967009 | 2023 | 577 b | A | 49 | 44 | 297 | 42 | 482 |
| Strain | GenBank Accession no./ PubMLST id | Percent Identity | Host (Strain) | Common Name |
|---|---|---|---|---|
| wMame1 | MN927215/ id: #1828 | 100% | Paratrechina longicornis (wLonF) | longhorn crazy ant |
| wMame2 | KC161941 | 97.22% | Tachinid sp. | tachinid fly |
| KC161936 | 97.22% | Pyralidid sp. | pyralid moth | |
| wMsp1 | MG797608 | 99.63% | Loxoblemmus equestris | hard-headed cricket |
| wMsp2 | MN044106 | 100% | Macrodinychus multispinosus | parasitoid mite |
| MN927215/ id: #1828 | 99.62% | Paratrechina longicornis (wLonF) | longhorn crazy ant | |
| wMsp3 | MN044106 | 99.62% | Macrodinychus multispinosus | parasitoid mite |
| MN927215/ id: #1828 | 99.23% | Paratrechina longicornis (wLonF) | longhorn crazy ant | |
| wMsp4 | MN967009 | 100% | Camponotus sp. (wCamA3) | carpenter ant |
| KU527484 | 100% | Tetramorium lanuginosum | wooly ant | |
| KC137165 | 100% | Odontomachus sp. | trap jaw ant | |
| GU236978 | 100% | Aulacophora nigripennis | leaf beetle | |
| MG551859 | 100% | Octodonta nipae | nipa palm hispid beetle | |
| id: #120 | 100% | Camponotus leonardi | carpenter ant | |
| wMsp5 | KM078883 | 94.64% | Chorthippus parallelus | meadow grasshopper |
| JN701984 | 94.41% | Chorthippus parallelus | meadow grasshopper | |
| wMsp6 | MN927214/ id: #1827 | 100% | Paratrechina longicornis (wLonA) | longhorn crazy ant |
| MN927217 | 100% | Camponotus sp. (wCamA1) | carpenter ant | |
| KU527480 | 100% | Tapinoma sessile | odorous house ant | |
| KU527478 | 100% | Tapinoma melanocephalum | ghost ant | |
| HQ602874 | 100% | Ceutorhynchus neglectus | weevil | |
| AB024571 | 100% | Ephestia cautella | almond moth | |
| id: #111 | 100% | Technomyrmex albipes | white-footed ant | |
| id: #115 | 100% | Leptomyrmex sp. | spider ant | |
| id: #141 | 100% | Pheidole sp. | big-headed ant | |
| id: #146 | 100% | Leptogenys sp. | razorjaw ant | |
| id: #116 | 100% | Myrmecorhynchus sp. | ant | |
| id: #124 | 100% | Pheidole plagiara | big-headed ant | |
| id: #125 | 100% | Pheidole sauberi | big-headed ant | |
| id: #135 | 100% | Ochetellus glaber | black household ant | |
| id: #13 | 100% | Ephestia kuehniella | mediterranean flour moth | |
| id: #123 | 100% | Ornipholidotos peucetia | glasswings | |
| id: #451 | 100% | Aricia artaxerxes | northern brown argus | |
| wMsp7 | MN967009 | 99.81% | Camponotus sp. (wCamA3) | carpenter ant |
| KU527484 | 99.81% | Tetramorium lanuginosum | wooly ant | |
| KC137165 | 99.81% | Odontomachus sp. | trap jaw ant | |
| GU236978 | 99.81% | Aulacophora nigripennis | leaf beetles | |
| MG551859 | 99.81% | Octodonta nipae | nipa palm hispid beetle | |
| id: #120 | 99.81% | Camponotus leonardi | carpenter ants | |
| wMsp8 | EF219194 | 95.47% | Ixodes ricinus | castor bean tick |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tseng, S.-P.; Hsu, P.-W.; Lee, C.-C.; Wetterer, J.K.; Hugel, S.; Wu, L.-H.; Lee, C.-Y.; Yoshimura, T.; Yang, C.-C.S. Evidence for Common Horizontal Transmission of Wolbachia among Ants and Ant Crickets: Kleptoparasitism Added to the List. Microorganisms 2020, 8, 805. https://doi.org/10.3390/microorganisms8060805
Tseng S-P, Hsu P-W, Lee C-C, Wetterer JK, Hugel S, Wu L-H, Lee C-Y, Yoshimura T, Yang C-CS. Evidence for Common Horizontal Transmission of Wolbachia among Ants and Ant Crickets: Kleptoparasitism Added to the List. Microorganisms. 2020; 8(6):805. https://doi.org/10.3390/microorganisms8060805
Chicago/Turabian StyleTseng, Shu-Ping, Po-Wei Hsu, Chih-Chi Lee, James K. Wetterer, Sylvain Hugel, Li-Hsin Wu, Chow-Yang Lee, Tsuyoshi Yoshimura, and Chin-Cheng Scotty Yang. 2020. "Evidence for Common Horizontal Transmission of Wolbachia among Ants and Ant Crickets: Kleptoparasitism Added to the List" Microorganisms 8, no. 6: 805. https://doi.org/10.3390/microorganisms8060805