Description of Three Novel Members in the Family Geobacteraceae, Oryzomonas japonicum gen. nov., sp. nov., Oryzomonas sagensis sp. nov., and Oryzomonas ruber sp. nov.

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Culture Conditions
2.2. Morphological and Physiological Analysis
2.3. Phylogenetic Analysis
2.4. Genome Sequencing, Assembly and Annotation
2.5. Genome Comparison
2.6. Genomic Fingerprints
2.7. Chemotaxonomic Characterization
3. Results and Discussion
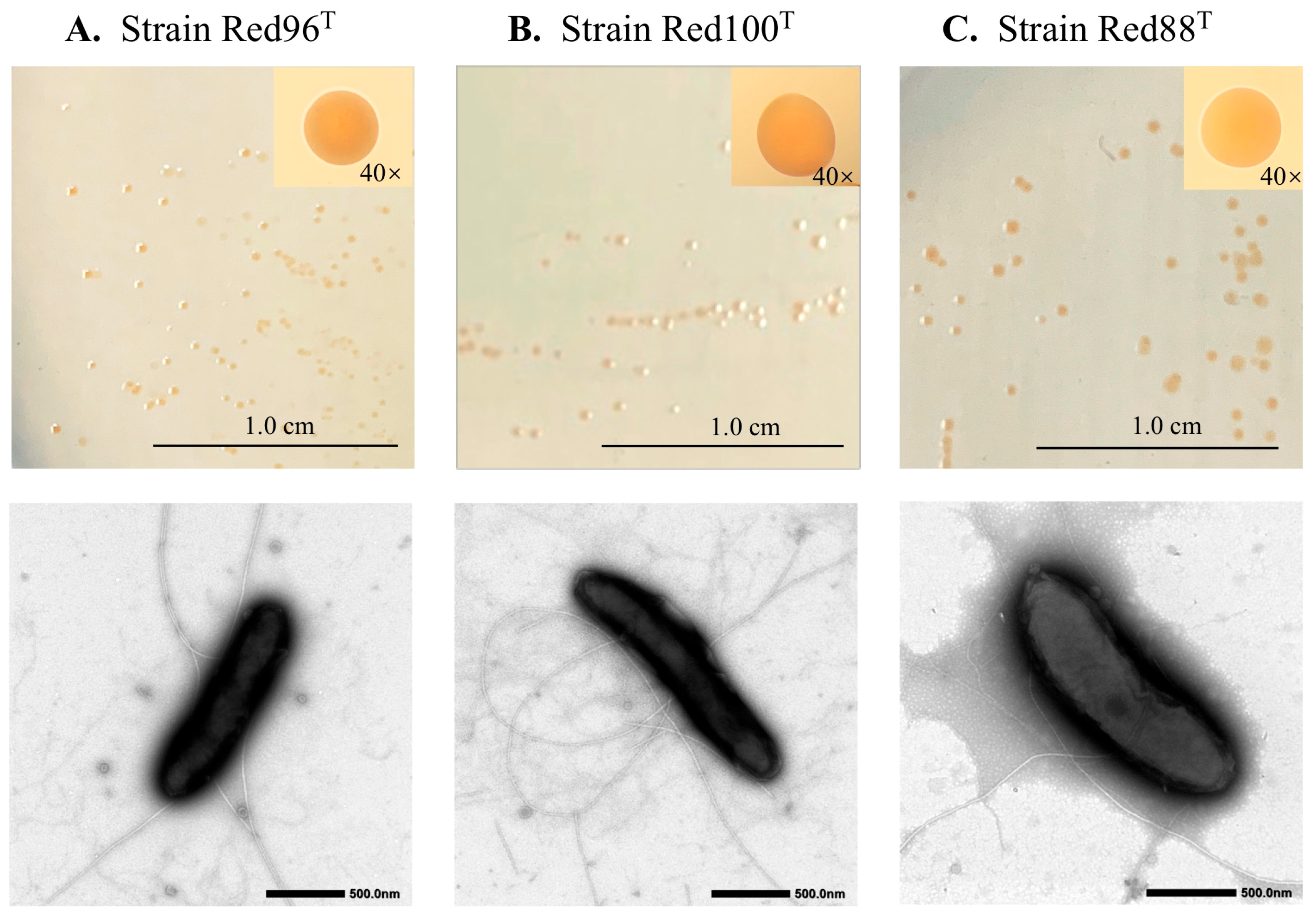
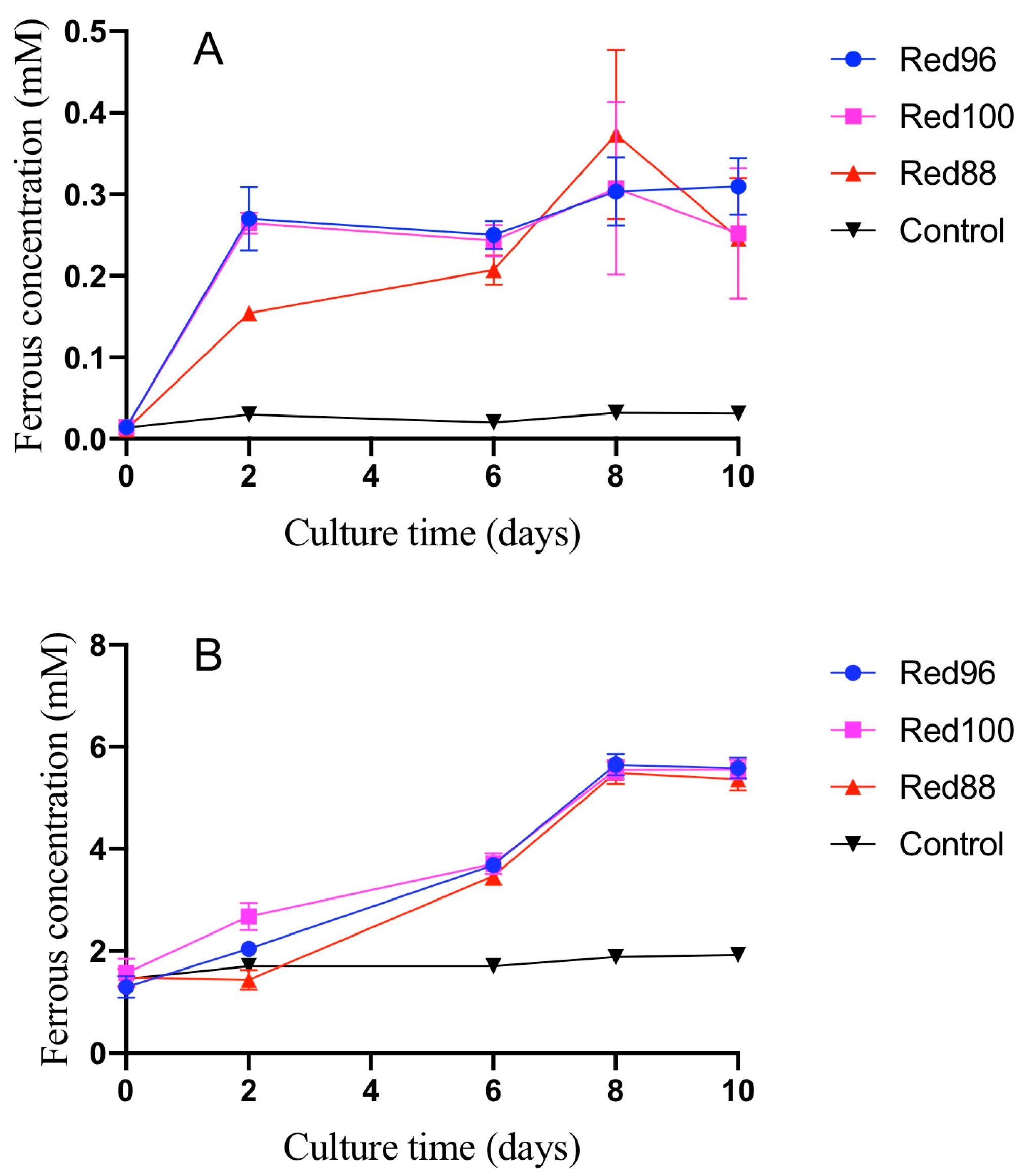
3.1. Morphological and Physiological Analysis
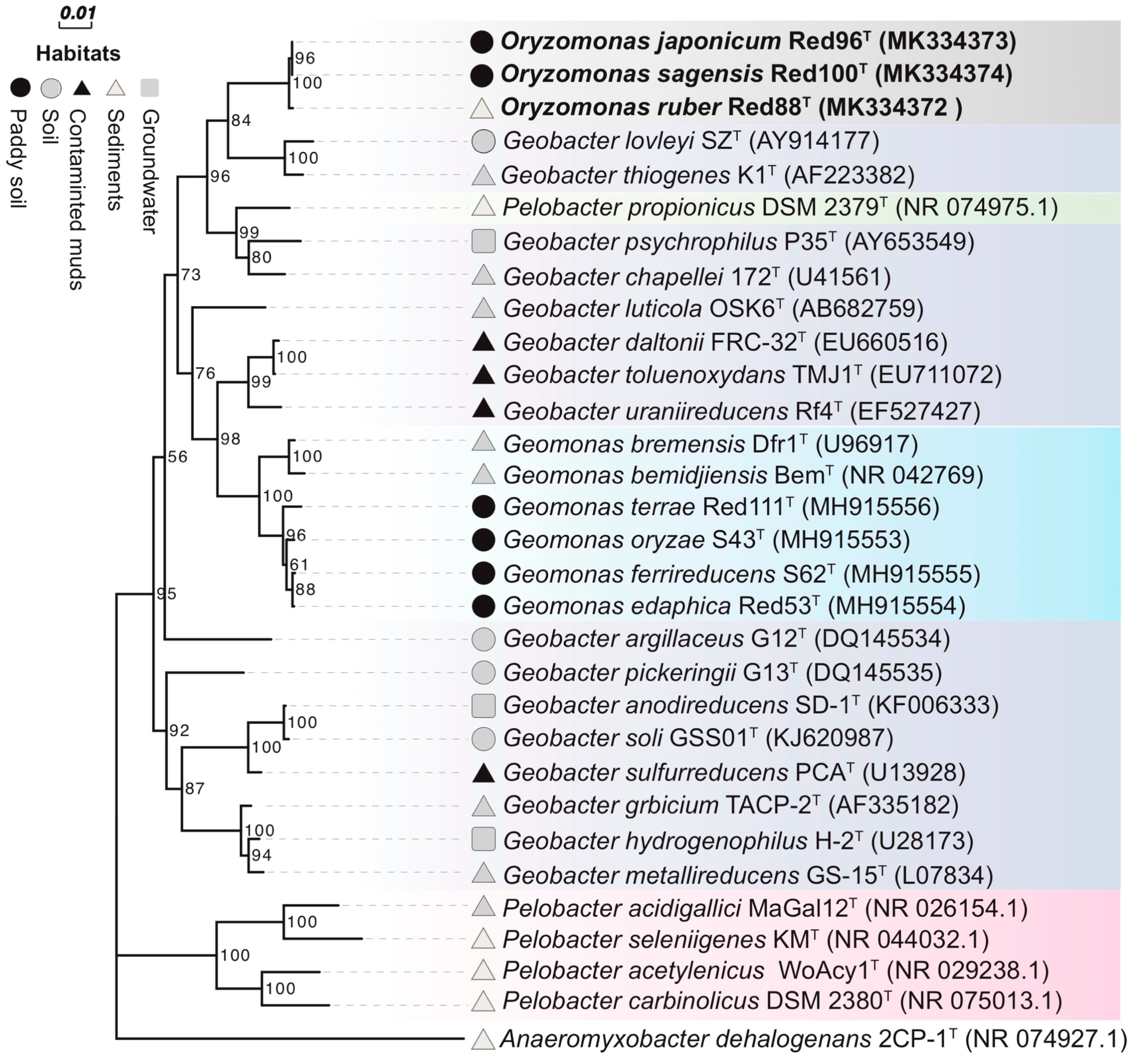
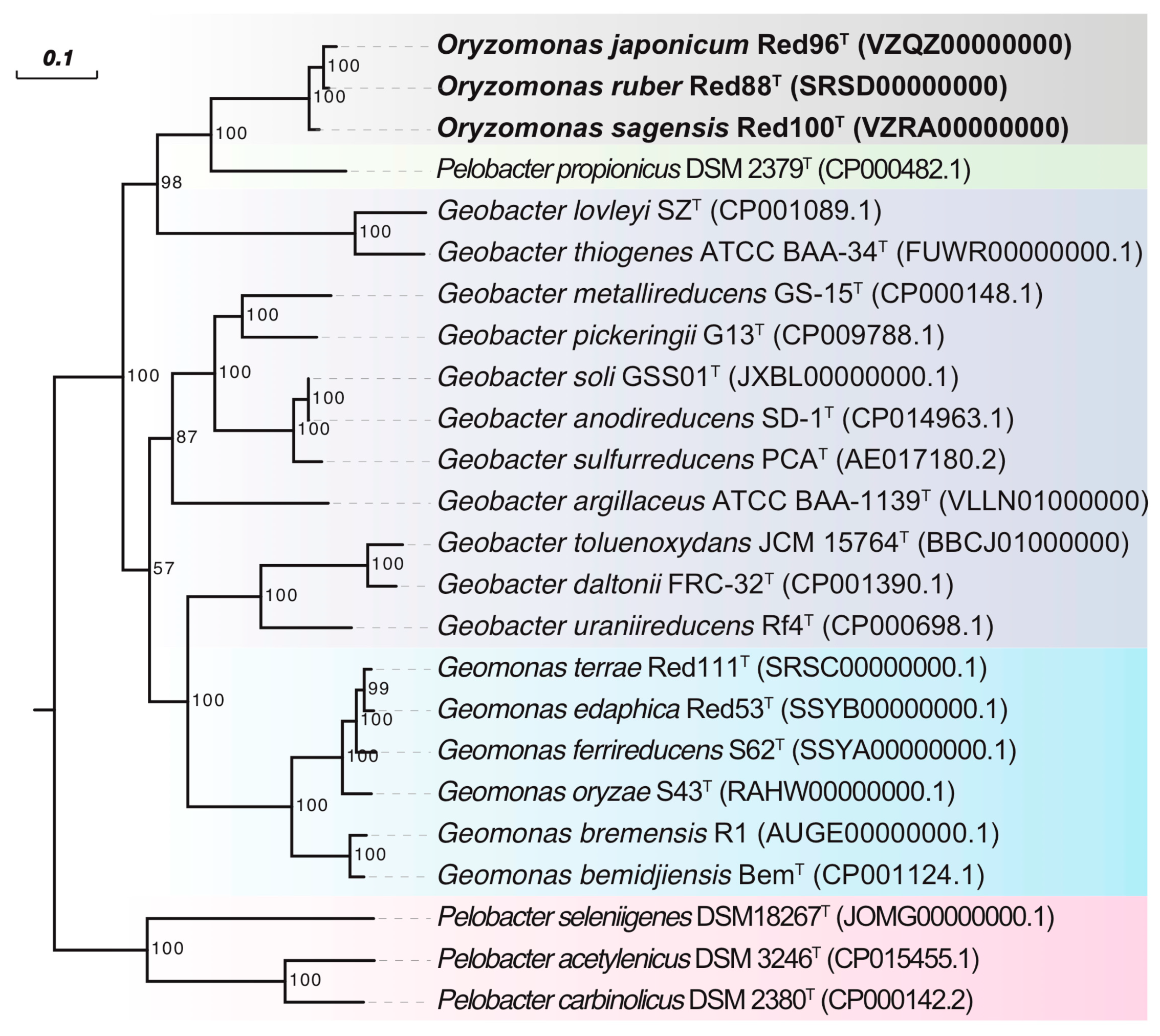
3.2. Phylogenetic Analysis
3.3. Genome Characteristics
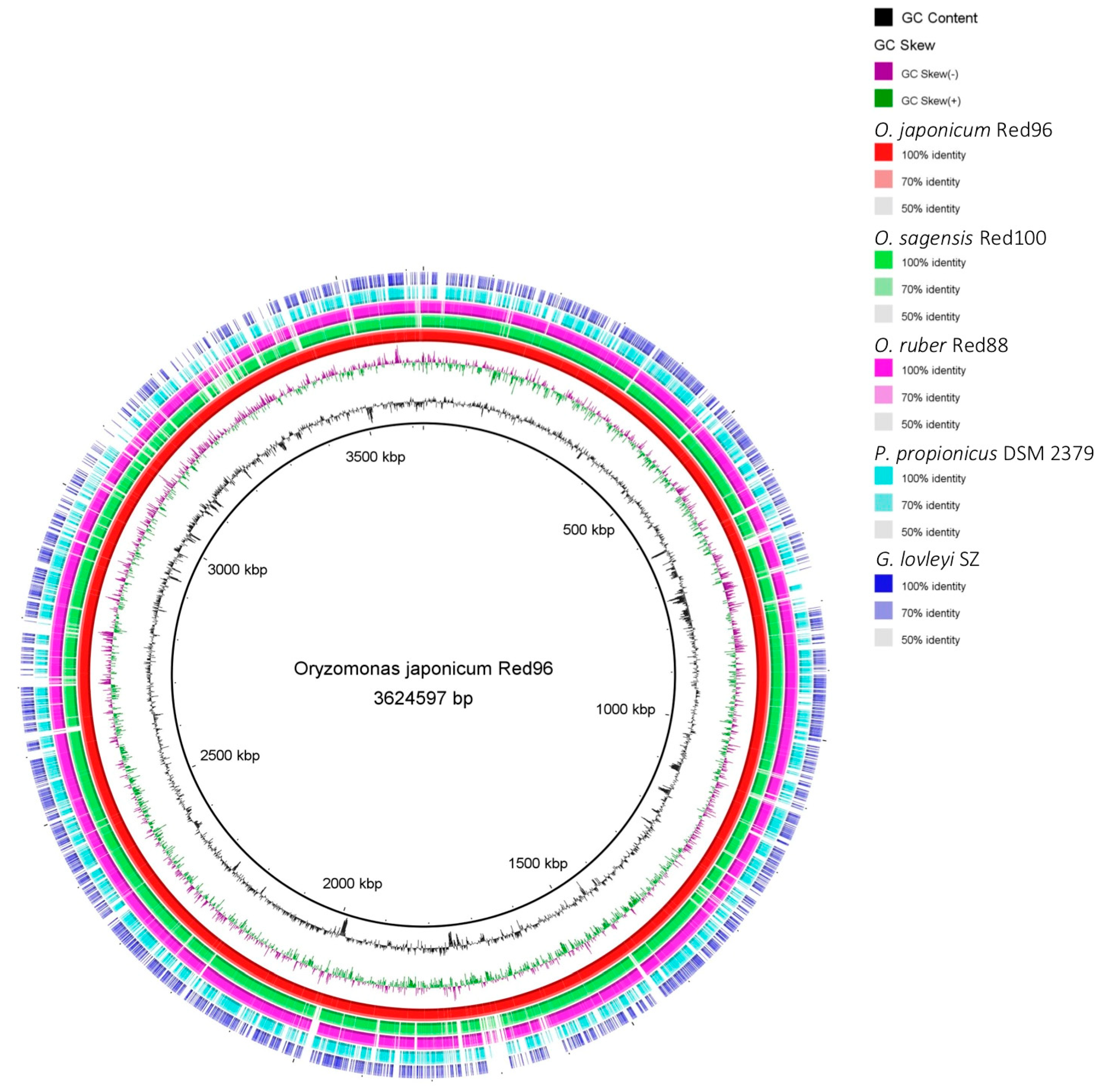
3.4. Genome Comparison
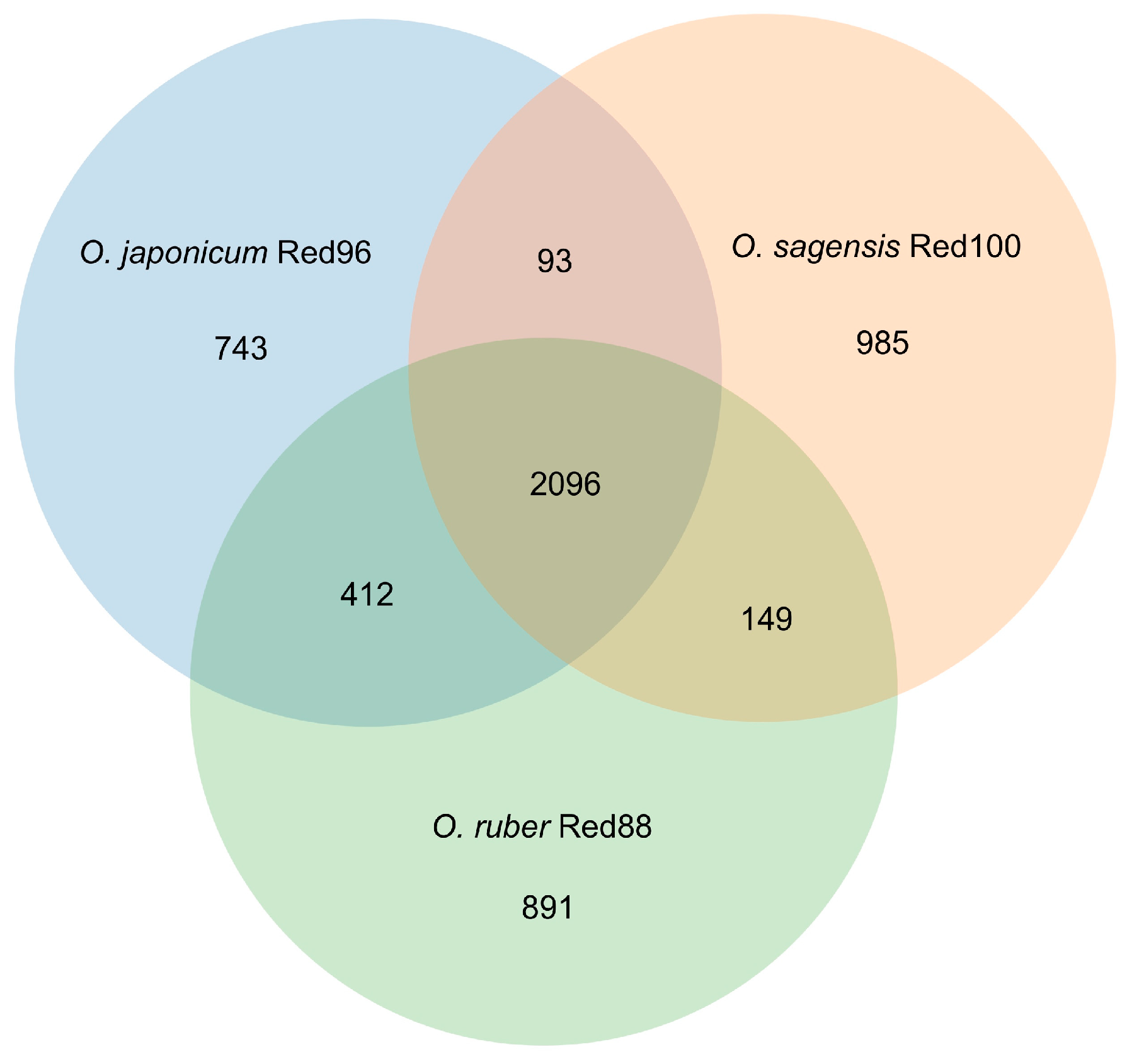
3.5. Genomic Fingerprints
3.6. Chemotaxonomic Characterization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- Röling, W.F.M. The Family Geobacteraceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Heidelberg, Germany, 2014; pp. 157–172. [Google Scholar]
- Xu, Z.; Masuda, Y.; Itoh, H.; Ushijima, N.; Shiratori, Y.; Senoo, K. Geomonas edaphica sp. nov., Geomonas ferrireducens sp. nov., Geomonas terrae sp. nov., four ferric-reducing bacteria isolated from paddy soil, and reclassification of three species of the genus Geobacter as members of the genus Geomonas gen. nov. Front. Microbiol. 2019, 10, 2201. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R.; Giovannoni, S.J.; White, D.C.; Champine, J.E.; Phillips, E.J.P.; Gorby, Y.A.; Goodwin, S. Geobacter metallireducens gen. nov. sp. nov., a microorganism capable of coupling the complete oxidation of organic compounds to the reduction of iron and other metals. Arch. Microbiol. 1993, 159, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R.; Ueki, T.; Zhang, T.; Malvankar, N.S.; Shrestha, P.M.; Flanagan, K.A.; Aklujkar, M.; Butler, J.E.; Giloteaux, L.; Rotaru, A.E.; et al. Geobacter: The microbe electric’s physiology, ecology, and practical applications. Adv. Microb. Physiol. 2011, 59, 1–100. [Google Scholar] [CrossRef] [PubMed]
- Viulu, S.; Nakamura, K.; Okada, Y.; Saitou, S.; Takamizawa, K. Geobacter luticola sp. nov., an Fe (III)-reducing bacterium isolated from lotus field mud. Int. J. Syst. Evol. Microbiol. 2013, 2, 442–448. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Yang, G.; Lu, Q.; Wu, M. Geobacter soli sp. nov., a dissimilatory Fe(III)-reducing bacterium isolated from forest soil. Int. J. Syst. Evol. Microbiol. 2014, 64, 3786–3791. [Google Scholar] [CrossRef] [Green Version]
- Holmes, D.E.; Giloteaux, L.; Chaurasia, A.K.; Williams, K.H.; Luef, B.; Wilkins, M.J.; Wrighton, K.C.; Thompson, C.A.; Comolli, L.R.; Lovley, D.R. Evidence of Geobacter-associated phage in a uranium-contaminated aquifer. ISME J. 2015, 9, 333–346. [Google Scholar] [CrossRef] [Green Version]
- Hori, T.; Müller, A.; Igarashi, Y.; Conrad, R.; Friedrich, M.W. Identification of iron-reducing microorganisms in anoxic rice paddy soil by 13C-acetate probing. ISME J. 2010, 4, 267–278. [Google Scholar] [CrossRef]
- Li, Z.; Suzuki, D.; Zhang, C.; Yang, S.; Nan, J.; Yoshida, N.; Wang, A.; Katayama, A. Anaerobic 4-chlorophenol mineralization in an enriched culture under iron-reducing conditions. J. Biosci. Bioeng. 2014, 118, 529–532. [Google Scholar] [CrossRef]
- Ding, L.J.; Su, J.Q.; Xu, H.J.; Jia, Z.J.; Zhu, Y.G. Long-term nitrogen fertilization of paddy soil shifts iron-reducing microbial community revealed by RNA-13C-acetate probing coupled with pyrosequencing. ISME J. 2015, 9, 721–734. [Google Scholar] [CrossRef]
- Itoh, H.; Ishii, S.; Shiratori, Y.; Oshima, K.; Otsuka, S.; Hattori, M.; Senoo, K. Seasonal transition of active bacterial and archaeal communities in relation to water management in paddy soils. Microbes Environ. 2013, 28, 370–380. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Liesack, W. Differential assemblage of functional units in paddy soil microbiomes. PLoS ONE 2015, 10, e0122221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, Y.; Itoh, H.; Shiratori, Y.; Isobe, K.; Otsuka, S.; Senoo, K. Predominant but previously-overlooked prokaryotic drivers of reductive nitrogen transformation in paddy soils, revealed by metatranscriptomics. Microbes Environ. 2017, 32, 180–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tipayno, S.C.; Truu, J.; Samaddar, S.; Truu, M.; Preem, J.K.; Oopkaup, K.; Espenberg, M.; Chatterjee, P.; Kang, Y.; Kim, K.; et al. The bacterial community structure and functional profile in the heavy metal contaminated paddy soils, surrounding a nonferrous smelter in South Korea. Ecol. Evol. 2018, 8, 6157–6168. [Google Scholar] [CrossRef] [PubMed]
- Holmes, D.E.; Shrestha, P.M.; Walker, D.J.F.; Dang, Y.; Nevin, K.P.; Woodard, T.L.; Lovley, D.R. Metatranscriptomic evidence for direct interspecies electron transfer between Geobacter and Methanothrix species in methanogenic rice paddy soils. Appl. Environ. Microbiol. 2017, 83, e00223-17. [Google Scholar] [CrossRef] [Green Version]
- Hori, T.; Aoyagi, T.; Itoh, H.; Narihiro, T.; Oikawa, A.; Suzuki, K.; Ogata, A.; Friedrich, M.W.; Conrad, R.; Kamagata, Y. Isolation of microorganisms involved in reduction of crystalline iron(III) oxides in natural environments. Front. Microbiol. 2015, 6, 386. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Su, J.Q.; Yang, X.R.; Zhou, G.W.; Lassen, S.B.; Zhu, Y.G. RNA stable isotope probing of potential feammox population in paddy soil. Environ. Sci. Technol. 2019, 53, 4841–4849. [Google Scholar] [CrossRef]
- Masuda, Y.; Itoh, H.; Shiratori, Y.; Senoo, K. Metatranscriptomic insights into microbial consortia driving methane metabolism in paddy soils. Soil Sci. Plant. Nutr. 2018, 64, 455–464. [Google Scholar] [CrossRef]
- Deng, D.; Zhang, Y.; Liu, Y. A Geobacter strain isolated from rice paddy soil with higher bioelectricity generation capability in comparison to Geobacter sulfurreducens PCA. RSC Adv. 2015, 5, 43978–43989. [Google Scholar] [CrossRef]
- Ohtsuka, T.; Yamaguchi, N.; Makino, T.; Sakurai, K.; Kimura, K.; Kudo, K.; Homma, E.; Dong, D.T.; Amachi, S. Arsenic dissolution from Japanese paddy soil by a dissimilatory arsenate-reducing bacterium Geobacter sp. OR-1. Environ. Sci. Technol. 2013, 47, 6263–6271. [Google Scholar] [CrossRef]
- Willems, A. The taxonomy of rhizobia: An overview. Plant. Soil 2006, 287, 3–14. [Google Scholar] [CrossRef]
- Lagier, J.C.; Dubourg, G.; Million, M.; Cadoret, F.; Bilen, M.; Fenollar, F.; Rolain, J.M.; Fournier, P.E.; Raoult, D. Culturing the human microbiota and culturomics. Nat. Rev. Microbiol. 2018, 16, 540–550. [Google Scholar] [CrossRef] [Green Version]
- Nevin, K.P.; Holmes, D.E.; Woodard, T.L.; Hinlein, E.S.; Ostendorf, D.W.; Lovley, D.R. Geobacter bemidjiensis sp. nov. and Geobacter psychrophilus sp. nov., two novel Fe(III)-reducing subsurface isolates. Int. J. Syst. Evol. Microbiol. 2005, 55, 1667–1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onley, J.R.; Ahsan, S.; Sanford, R.A. Denitrification by Anaeromyxobacter dehalogenans, a common soil bacterium lacking the nitrite reductase genes nirS and nirK. Appl. Environ. Microbiol. 2018, 84, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Ashida, N.; Ishii, S.; Hayano, S.; Tago, K.; Tsuji, T.; Yoshimura, Y.; Otsuka, S.; Senoo, K. Isolation of functional single cells from environments using a micromanipulator: Application to study denitrifying bacteria. Appl. Microbiol. Biotechnol. 2010, 85, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Glaeser, S.P.; Kämpfer, P. Multilocus sequence analysis (MLSA) in prokaryotic taxonomy. Syst. Appl. Microbiol. 2015, 38, 237–245. [Google Scholar] [CrossRef]
- Holmes, D.E.; Nevin, K.P.; Lovley, D.R. Comparison of 16S rRNA, nifD, recA, gyrB, rpoB and fusA genes within the family Geobacteraceae fam. nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 1591–1599. [Google Scholar] [CrossRef] [Green Version]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef] [Green Version]
- Boetzer, M.; Henkel, C.V.; Jansen, H.J.; Butler, D.; Pirovano, W. Scaffolding pre-assembled contigs using SSPACE. Bioinformatics 2010, 27, 578–579. [Google Scholar] [CrossRef] [Green Version]
- Boetzer, M.; Pirovano, W. Toward almost closed genomes with GapFiller. Genome Biol. 2012, 13, R56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG tools for functional characterization of genome and metagenome sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huerta-Cepas, J.; Forslund, K.; Coelho, L.P.; Szklarczyk, D.; Jensen, L.J.; von Mering, C.; Bork, P. Fast genome-wide functional annotation through orthology assignment by eggNOG-Mapper. Mol. Biol. Evol. 2017, 34, 2115–2122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Konstantinidis, K.T.; Tiedje, J.M. Towards a genome-based taxonomy for prokaryotes. J. Bacteriol. 2005, 187, 6258–6264. [Google Scholar] [CrossRef] [Green Version]
- Qin, Q.L.; Xie, B.B.; Zhang, X.Y.; Chen, X.L.; Zhou, B.C.; Zhou, J.; Oren, A.; Zhang, Y.Z. A proposed genus boundary for the prokaryotes based on genomic insights. J. Bacteriol. 2014, 196, 2210–2215. [Google Scholar] [CrossRef] [Green Version]
- Na, S.I.; Kim, Y.O.; Yoon, S.H.; Ha, S.; Baek, I.; Chun, J. UBCG: Up-to-date bacterial core gene set and pipeline for phylogenomic tree reconstruction. J. Microbiol. 2018, 56, 280–285. [Google Scholar] [CrossRef]
- Alikhan, N.F.; Petty, N.K.; Zakour, N.L.B.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genomics 2011, 12, 402. [Google Scholar] [CrossRef] [Green Version]
- Darling, A.C.E.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guy, L.; Kultima, J.R.; Andersson, S.G.E. genoPlotR: Comparative gene and genome visualization in R. Bioinformatics 2010, 26, 2334–2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Pauls, K.P. Optimization of the PCR program for RAPD analysis. Nucleic Acids Res. 1992, 20, 2606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Versalovic, J.; Schneider, M.; De Bruijn, F.J.; Lupski, J.R. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Methods Mol. Cell. Biol. 1994, 5, 25–40. [Google Scholar]
- Kuykendall, L.D.; Roy, M.A.; O’neill, J.J.; Devine, T.E. Fatty acids, antibiotic resistance, and deoxyribonucleic acid homology groups of Bradyrhizobium japonicum. Int. J. Syst. Bacteriol. 1988, 38, 358–361. [Google Scholar] [CrossRef] [Green Version]
- Gunina, A.; Dippold, M.A.; Glaser, B.; Kuzyakov, Y. Fate of low molecular weight organic substances in an arable soil: From microbial uptake to utilisation and stabilisation. Soil Biol. Biochem. 2014, 77, 304–313. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Afkar, E.; Fukumori, Y. Purification and characterization of triheme cytochrome c7 from the metal-reducing bacterium, Geobacter metallireducens. FEMS Microbiol. Lett. 1999, 175, 205–210. [Google Scholar] [CrossRef]
- Schink, B. Fermentation of 2,3-butanediol by Pelobacter carbinolicus sp. nov. and Pelobacter propionicus sp. nov., and evidence for propionate formation from C2 compounds. Arch. Microbiol. 1984, 137, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Sung, Y.; Fletcher, K.E.; Ritalahti, K.M.; Apkarian, R.P.; Ramos-Hernández, N.; Sanford, R.A.; Mesbah, N.M.; Löffler, F.E. Geobacter lovleyi sp. nov. strain SZ, a novel metal-reducing and tetrachloroethene-dechlorinating bacterium. Appl. Environ. Microbiol. 2006, 72, 2775–2782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nevin, K.P.; Holmes, D.E.; Woodard, T.L.; Covalla, S.F.; Lovley, D.R. Reclassification of Trichlorobacter thiogenes as Geobacter thiogenes comb. nov. Int. J. Syst. Evol. Microbiol. 2007, 57, 463–466. [Google Scholar] [CrossRef] [PubMed]
- Coates, J.D.; Bhupathiraju, V.K.; Achenbach, L.A.; Mcinerney, M.J.; Lovley, D.R. Geobacter hydrogenophilus, Geobacter chapellei and Geobacter grbiciae, three new, strictly anaerobic, dissimilatory Fe(III)-reducers. Int. J. Syst. Evol. Microbiol. 2001, 335182, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glöckner, F.O.; Ludwig, W.; Schleifer, K.H.; Whitman, W.B.; Euzéby, J.; Amann, R.; Rosselló-Móra, R. Uniting the classification of cultured and uncultured bacteria and archaea using 16S rRNA gene sequences. Nat. Rev. Microbiol. 2014, 12, 635. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Dong, H.; Reguera, G.; Beyenal, H.; Lu, A.; Liu, J.; Yu, H.Q.; Fredrickson, J.K. Extracellular electron transfer mechanisms between microorganisms and minerals. Nat. Rev. Microbiol. 2016, 14, 651–662. [Google Scholar] [CrossRef]
- Shi, L.; Fredrickson, J.K.; Zachara, J.M. Genomic analyses of bacterial porin-cytochrome gene clusters. Front. Microbiol. 2014, 5, 657. [Google Scholar] [CrossRef]
- Cai, X.; Huang, L.; Yang, G.; Yu, Z.; Wen, J.; Zhou, S. Transcriptomic, proteomic, and bioelectrochemical characterization of an exoelectrogen Geobacter soli grown with different electron acceptors. Front. Microbiol. 2018, 9, 1075. [Google Scholar] [CrossRef] [Green Version]
- Holmes, D.E.; Dang, Y.; Walker, D.J.F.; Lovley, D.R. The electrically conductive pili of Geobacter species are a recently evolved feature for extracellular electron transfer. Microb. genomics 2016, 2, e000072. [Google Scholar] [CrossRef]
- Lovley, D.R.; Walker, D. Geobacter protein nanowires. Front. Microbiol. 2019, 10, 2078. [Google Scholar] [CrossRef] [Green Version]
- Malvankar, N.S.; Lovley, D.R. Microbial nanowires for bioenergy applications. Curr. Opin. Biotechnol. 2014, 27, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.J.; Adhikari, R.Y.; Holmes, D.E.; Ward, J.E.; Woodard, T.L.; Nevin, K.P.; Lovley, D.R. Electrically conductive pili from pilin genes of phylogenetically diverse microorganisms. ISME J. 2018, 12, 48–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.W.; Meyer, S.D.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA. 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.; Rodriguez-R, L.M.; Konstantinidis, K.T. MyTaxa: An advanced taxonomic classifier for genomic and metagenomic sequences. Nucleic Acids Res. 2014, 42, e73. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
|---|---|---|---|---|---|---|---|---|---|
| Optimal temperature (°C) | 30–33 | 30–33 | 30–33 | 25 | 33 | 35 | 30 | 30–35 | 30–33 |
| Optimal pH | 5.5–6.5 | 6.0–7.0 | 5.5–6.5 | 6.5–7.0 | 7.0–8.0 | 6.8 | 7.0 | ca. 6.7 | 6.0–7.0 |
| G + C content (mol%) * | 59.0 | 59.7 | 58.4 | 50.2 a | 58.5 | 54.8 | 52.8 | 56.6 | 61.2 |
| Motility | + | + | + | − | − | + | − | − | + |
| Electron acceptor usage | |||||||||
| Nitrate | − | − | + | − | − | + | + | + | + |
| Fumarate | + | + | + | + | − | + | + | − | + |
| Sulfur | − | − | − | − | − | + | + | − | − |
| MnO2 | + | − | + | + | ND | + | − | + | − |
| Fe(III) citrate | + | + | + | + | ND | + | − | + | − |
| electron donor usage | |||||||||
| Succinate | + | + | + | + | ND | − | − | − | + |
| Butanol | − | − | − | − | + | ND | ND | + | − |
| Ethanol | + | − | + | − | + | − | − | + | + |
| Propionate | + | + | + | + | − | − | − | + | + |
| Malate | + | + | + | + | − | ND | − | − | + |
| Lactate | + | − | + | + | + | − | − | − | + |
| Methanol | + | + | + | + | − | − | ND | − | + |
| Pyruvate | + | + | + | + | + | + | − | + | + |
| Glucose | + | + | + | + | − | − | ND | − | + |
| Toluene | − | − | − | − | ND | − | ND | + | − |
| Features | Red96T | Red100T | Red88T |
|---|---|---|---|
| Assembled contigs | 30 | 17 | 16 |
| Genome length (bp) | 3,624,587 | 3,609,742 | 3,798,725 |
| N50 length (bp) | 336,264 | 620,468 | 353,860 |
| Average G + C content (mol%) | 59.0 | 59.7 | 58.4 |
| Number of predicted ORFs | 3363 | 3322 | 3523 |
| Number of rRNAs | 5 | 4 | 3 |
| Number of tRNAs | 49 | 51 | 50 |
| Number of ncRNAs | 3 | 3 | 3 |
| Depth of coverage (×) | 808 | 958 | 271 |
| Accession number | VZQZ00000000 | VZRA01000000 | SRSD01000000 |
| Reference Strains # | ANI Value (%) | GGDC Value (%) | AAI Value (%) | POCP (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Red96T | Red100T | Red88T | Red96T | Red100T | Red88T | Red96T | Red100 T | Red88 T | Red96 T | Red100 T | Red88 T | |
| Oryzomonas japonicum Red96T | 100 | 100 | 100 | 100 | ||||||||
| Oryzomonas sagensis Red100T | 94.2 | 100 | 58.2 | 100 | 95.0 | 100 | 86.0 | 100 | ||||
| Oryzomonas ruber Red88T | 95.6 | 94.6 | 100 | 67.5 | 61.7 | 100 | 96.3 | 95.2 | 100 | 85.0 | 83.5 | 100 |
| Pelobacter propionicus Ott Bd 1T | 73.8 | 74.0 | 73.9 | 20.6 | 21.1 | 21.1 | 66.6 | 66.6 | 66.4 | 52.6 | 51.8 | 50.8 |
| Geobacter lovleyi SZT | 70.7 | 70.6 | 70.8 | 19.8 | 19.9 | 19.8 | 63.6 | 63.6 | 63.4 | 56.3 | 55.4 | 54.8 |
| Geobacter thiogenes ATCC BAA-34T | 70.3 | 70.0 | 70.3 | 19.2 | 20.0 | 19.7 | 63.7 | 63.5 | 63.2 | 55.8 | 54.7 | 53.6 |
| Geobacter metallireducens GS-15T | 70.8 | 70.9 | 70.8 | 20.2 | 19.9 | 20.2 | 62.3 | 62.4 | 62 | 57.8 | 57.3 | 56.6 |
| Geobacter uraniireducens Rf4T | 70.7 | 70.6 | 70.4 | 20.2 | 20.6 | 20.3 | 62.8 | 62.1 | 62.3 | 54.3 | 53.7 | 53.0 |
| Geobacter toluenoxydans JCM 15764T | 69.7 | 69.8 | 69.5 | 20.0 | 20.5 | 19.3 | 61.1 | 60.8 | 60.5 | 49.4 | 49.0 | 47.8 |
| Geobacter daltonii FRC-32T | 69.6 | 69.5 | 69.3 | 20.3 | 22.2 | 20.6 | 61.3 | 61.0 | 60.7 | 54.9 | 54.5 | 53.1 |
| Geobacter sulfurreducens PCAT | 70.8 | 70.8 | 70.9 | 20.0 | 20.5 | 20.2 | 62.6 | 62.7 | 62.3 | 60.3 | 59.7 | 59.0 |
| Geobacter anodireducens SD-1T | 71.0 | 71.0 | 71.1 | 19.7 | 20.2 | 20.0 | 61.4 | 61.4 | 61.3 | 53.5 | 53.2 | 52.5 |
| Geobacter pickeringii G13T | 71.5 | 71.4 | 71.5 | 20.3 | 20.7 | 20.6 | 63.3 | 63.3 | 63.4 | 61.6 | 61.6 | 60.4 |
| Geobacter soli GSS01T | 71.0 | 71.0 | 71.0 | 19.8 | 20.3 | 20.1 | 62.4 | 62.3 | 62.3 | 59.4 | 58.9 | 58.4 |
| Geomonas oryzae S43T | 70.3 | 70.5 | 70.3 | 19.3 | 19.4 | 19.3 | 60.8 | 60.7 | 60.7 | 57.8 | 56.9 | 56.9 |
| Geomonas edaphica Red53T | 70.3 | 70.3 | 70.2 | 19.2 | 19.1 | 19.3 | 60.9 | 60.9 | 60.6 | 58.0 | 56.7 | 56.8 |
| Geomonas ferrireducens S62T | 70.3 | 70.3 | 70.3 | 19.3 | 19.2 | 19.5 | 60.9 | 60.6 | 60.5 | 57.4 | 56.0 | 55.8 |
| Geomonas terrae Red111T | 69.8 | 70.0 | 69.9 | 19.0 | 19.3 | 19.2 | 60.5 | 60.6 | 60.3 | 58.1 | 56.6 | 56.7 |
| Geomonas bemidjiensis BemT | 70.2 | 70.3 | 70.2 | 19.8 | 19.8 | 19.9 | 60.6 | 60.6 | 60.5 | 56.8 | 56.1 | 56.1 |
| Geomonas bremensis R1 | 70.4 | 70.2 | 70.3 | 19.9 | 19.9 | 19.9 | 60.4 | 60.4 | 60.1 | 56.4 | 55.7 | 55.5 |
| Fatty Acid | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| C12:0 | - | - | 0.1 | 1.9 |
| iso-C13:0 | 0.2 | 0.3 | 0.3 | 2.6 |
| C14:0 | 5.5 | 9.4 | 9.9 | 4.5 |
| iso-C15:0 | 52.1 | 40.4 | 33.5 | 19.1 |
| a-C15:0 | 0.7 | 0.5 | 0.5 | 0.8 |
| C15:0 | 0.4 | 0.6 | 0.4 | 0.4 |
| C15:1 B/F * | 1.4 | 1.3 | 1.3 | 0.4 |
| iso-C16:0 | 1.7 | 0.8 | 0.8 | 1.3 |
| C16:0 | 7.4 | 10.8 | 13.8 | 20.1 |
| C16:1 B/F * | 1.9 | 0.5 | 0.4 | 1.9 |
| C16:1 ω7c | 19.2 | 22.2 | 26.0 | 32.9 |
| C16:1 ω5c | 0.7 | 0.6 | 1.0 | 2.1 |
| C16:0 10-methyl | 0.9 | 0.6 | 0.8 | ND |
| C16:0 3-OH | 2.2 | 7.0 | 7.4 | 7.3 |
| C17:1 B/F * | 1.6 | 0.6 | 0.5 | 0.3 |
| iso-C17:0 | 1.5 | 0.9 | 0.8 | 0.9 |
| a-C17:0 | 0.1 | 0.1 | 0.1 | 0.2 |
| C18:0 | 0.3 | 0.6 | 0.6 | 1.1 |
| C18:1 ω7c | 1.5 | 1.1 | 1.0 | 0.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Z.; Masuda, Y.; Hayakawa, C.; Ushijima, N.; Kawano, K.; Shiratori, Y.; Senoo, K.; Itoh, H. Description of Three Novel Members in the Family Geobacteraceae, Oryzomonas japonicum gen. nov., sp. nov., Oryzomonas sagensis sp. nov., and Oryzomonas ruber sp. nov. Microorganisms 2020, 8, 634. https://doi.org/10.3390/microorganisms8050634
Xu Z, Masuda Y, Hayakawa C, Ushijima N, Kawano K, Shiratori Y, Senoo K, Itoh H. Description of Three Novel Members in the Family Geobacteraceae, Oryzomonas japonicum gen. nov., sp. nov., Oryzomonas sagensis sp. nov., and Oryzomonas ruber sp. nov. Microorganisms. 2020; 8(5):634. https://doi.org/10.3390/microorganisms8050634
Chicago/Turabian StyleXu, Zhenxing, Yoko Masuda, Chie Hayakawa, Natsumi Ushijima, Keisuke Kawano, Yutaka Shiratori, Keishi Senoo, and Hideomi Itoh. 2020. "Description of Three Novel Members in the Family Geobacteraceae, Oryzomonas japonicum gen. nov., sp. nov., Oryzomonas sagensis sp. nov., and Oryzomonas ruber sp. nov." Microorganisms 8, no. 5: 634. https://doi.org/10.3390/microorganisms8050634