Symbioflor2® Escherichia coli Genotypes Enhance Ileal and Colonic Gene Expression Associated with Mucosal Defense in Gnotobiotic Mice

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Concerning Animal Experimentation
2.2. Establishment of Gnotobiotic Mice
2.2.1. Mouse Inoculation
2.2.2. Animal Experimentations
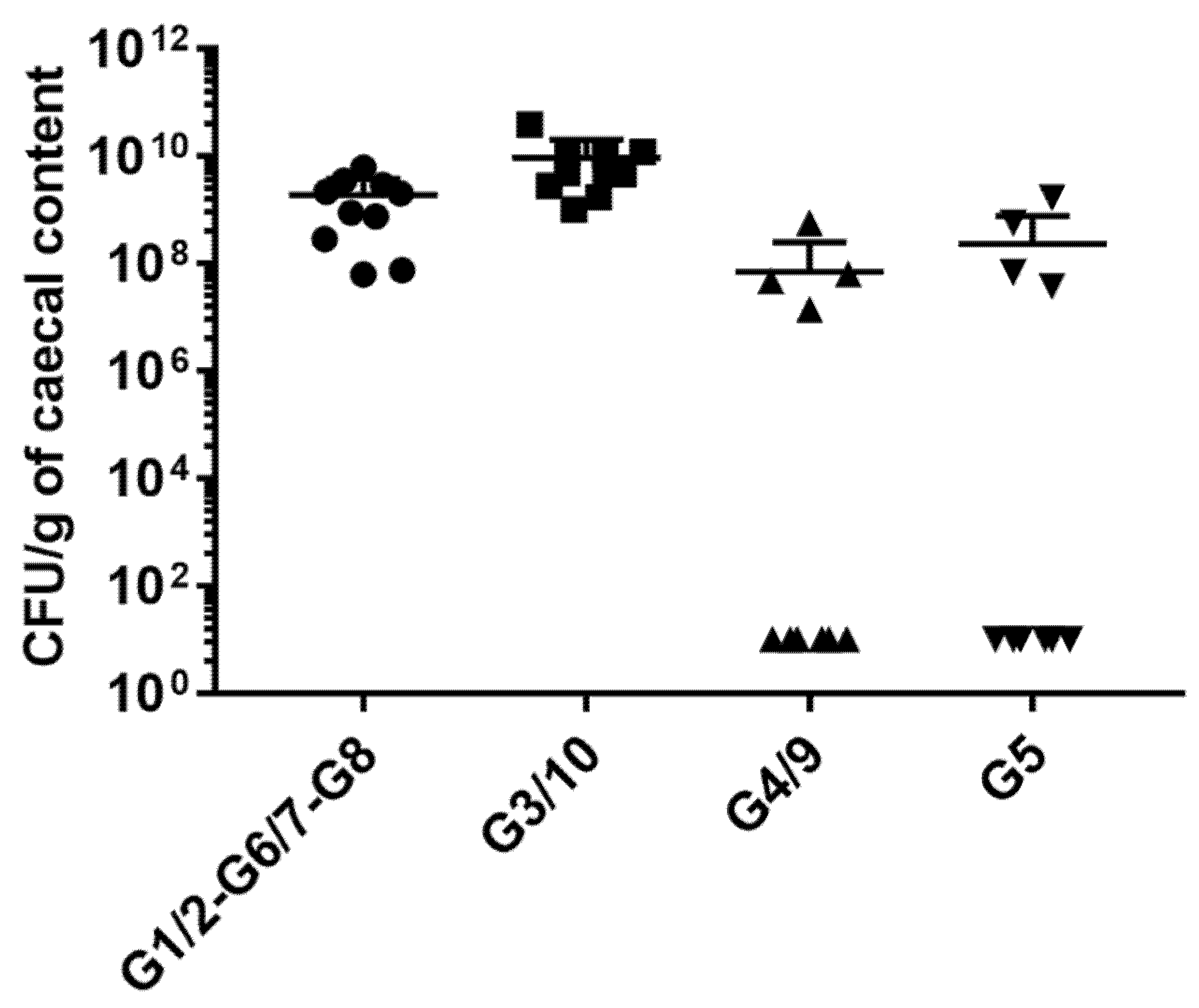
2.3. Monitoring of E. coli Colonization Levels in Cecal Samples of Monoxenic Mice by QPCR
2.4. Quantification of Colonization Level of E.coli Genotypes in Mice Inoculated with Symbioflor2®
2.5. Isolation of RNA from Intestinal Tissue and Preparation of cDNAs
2.6. Design of Ileal and Colonic Custom TaqMan® Array Card
2.7. Gene Expression Experiments
2.8. Gene Expression Analysis and Statistics
2.9. Single Real-Time Quantitative PCR Analyses of Gene Expression
2.10. Availability of Data and Materials
3. Results
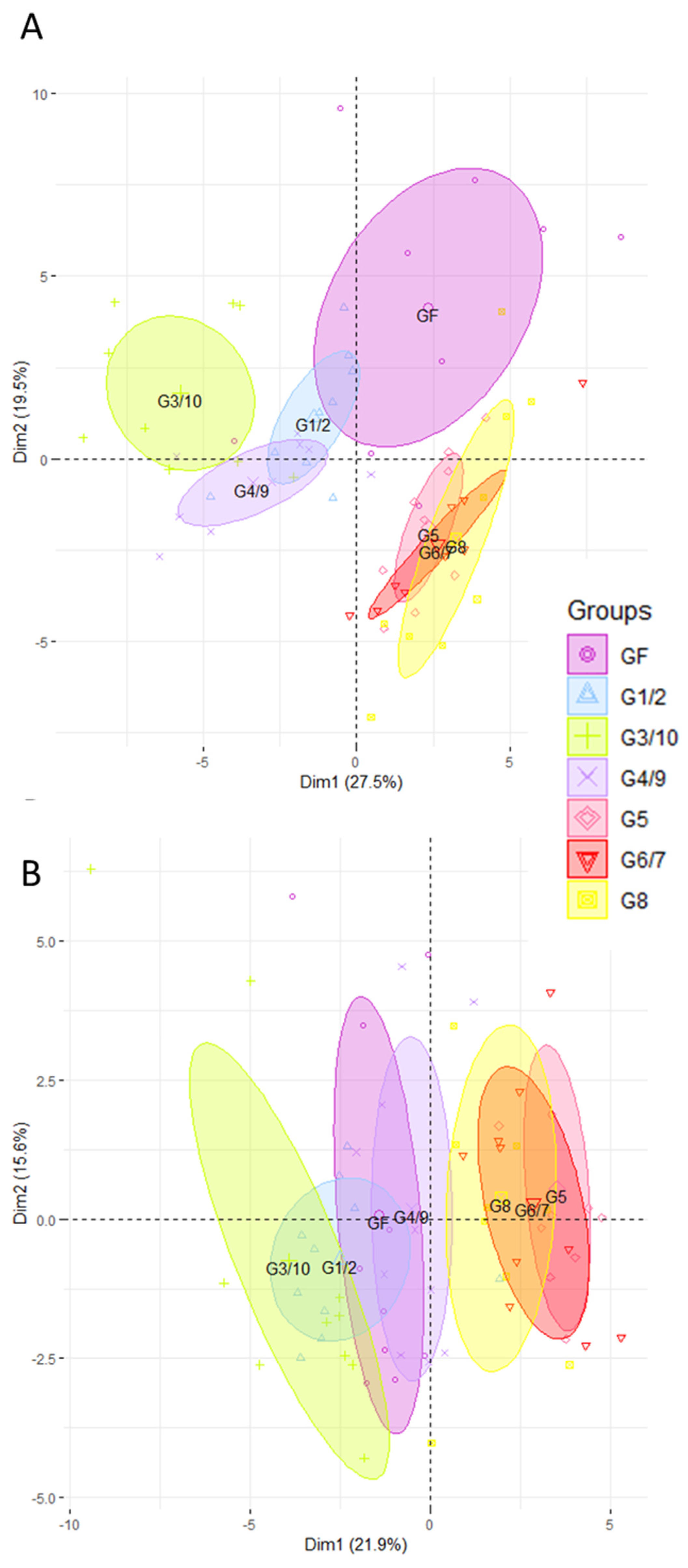
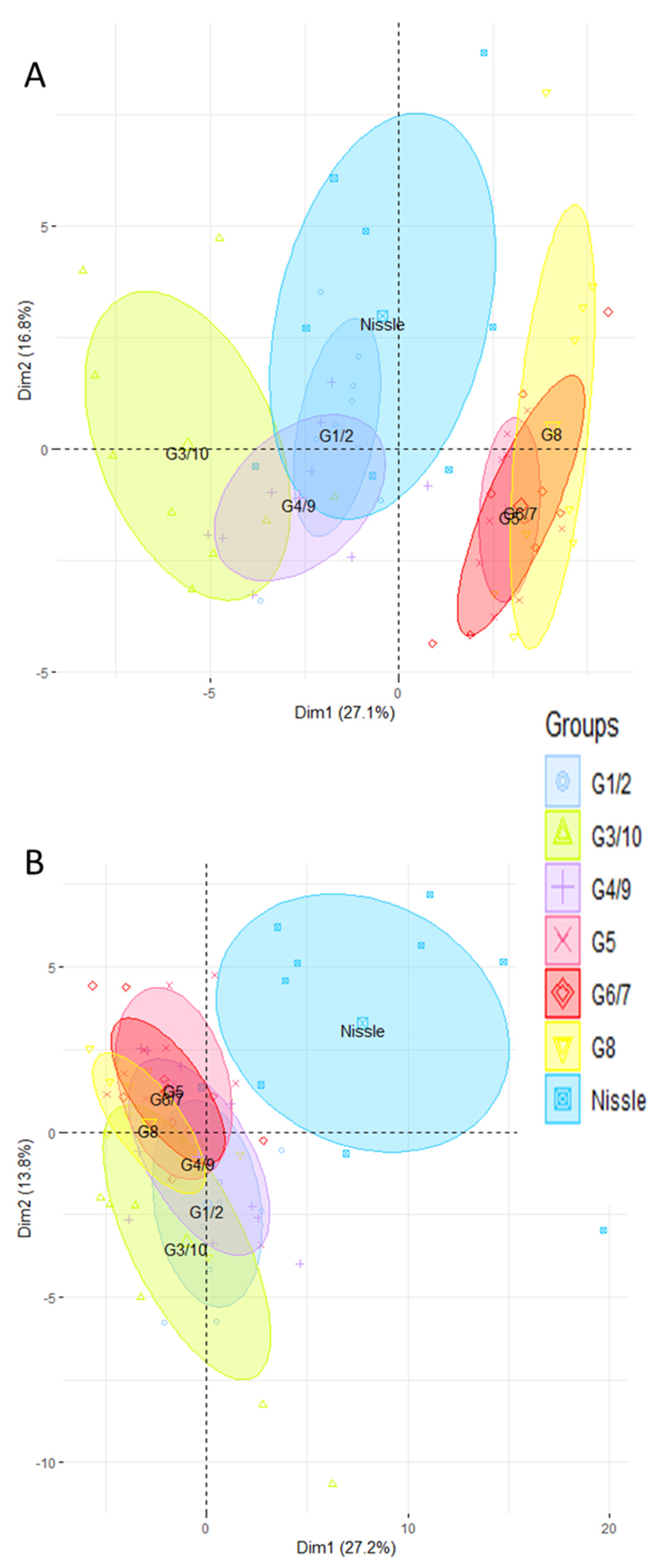
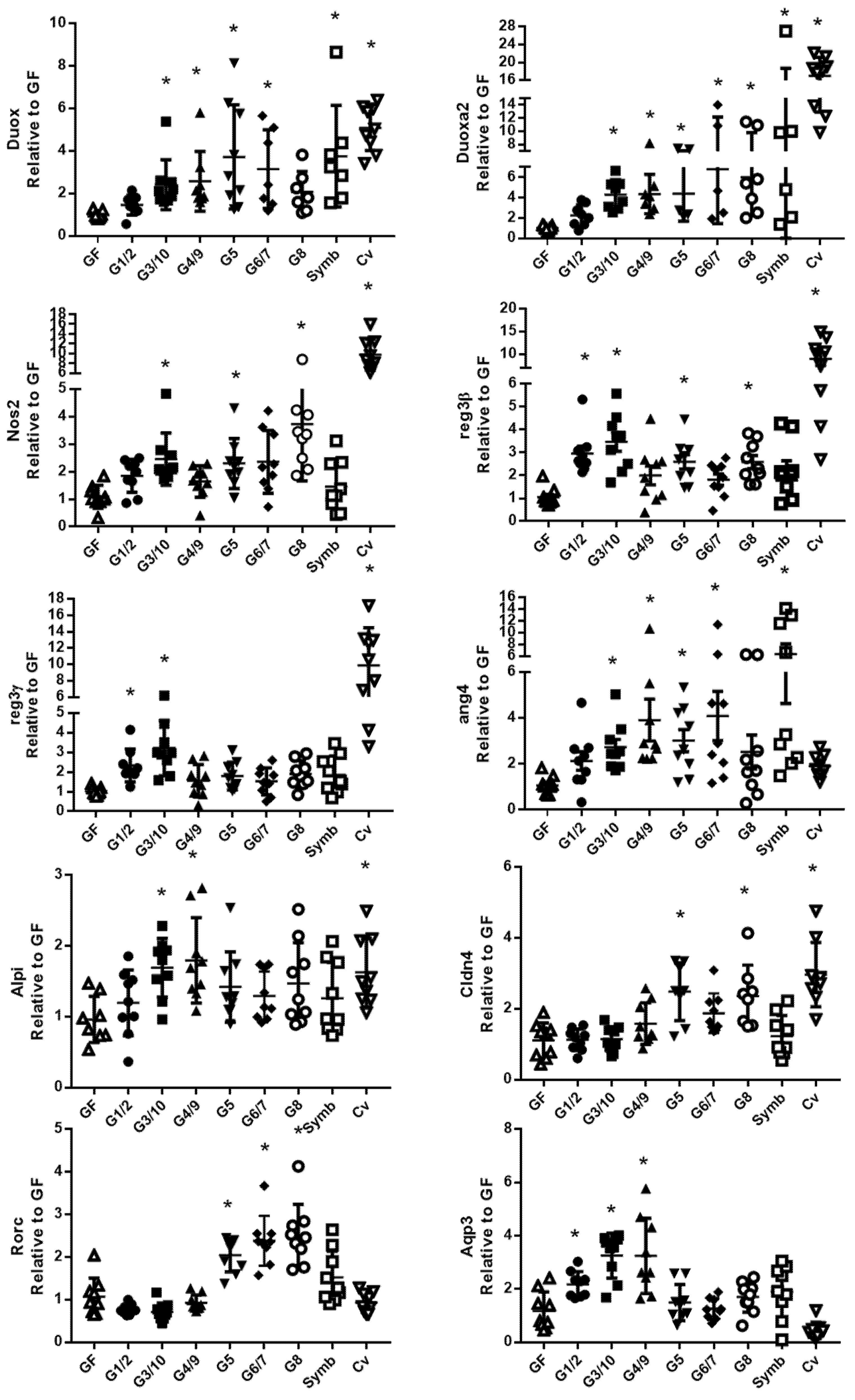
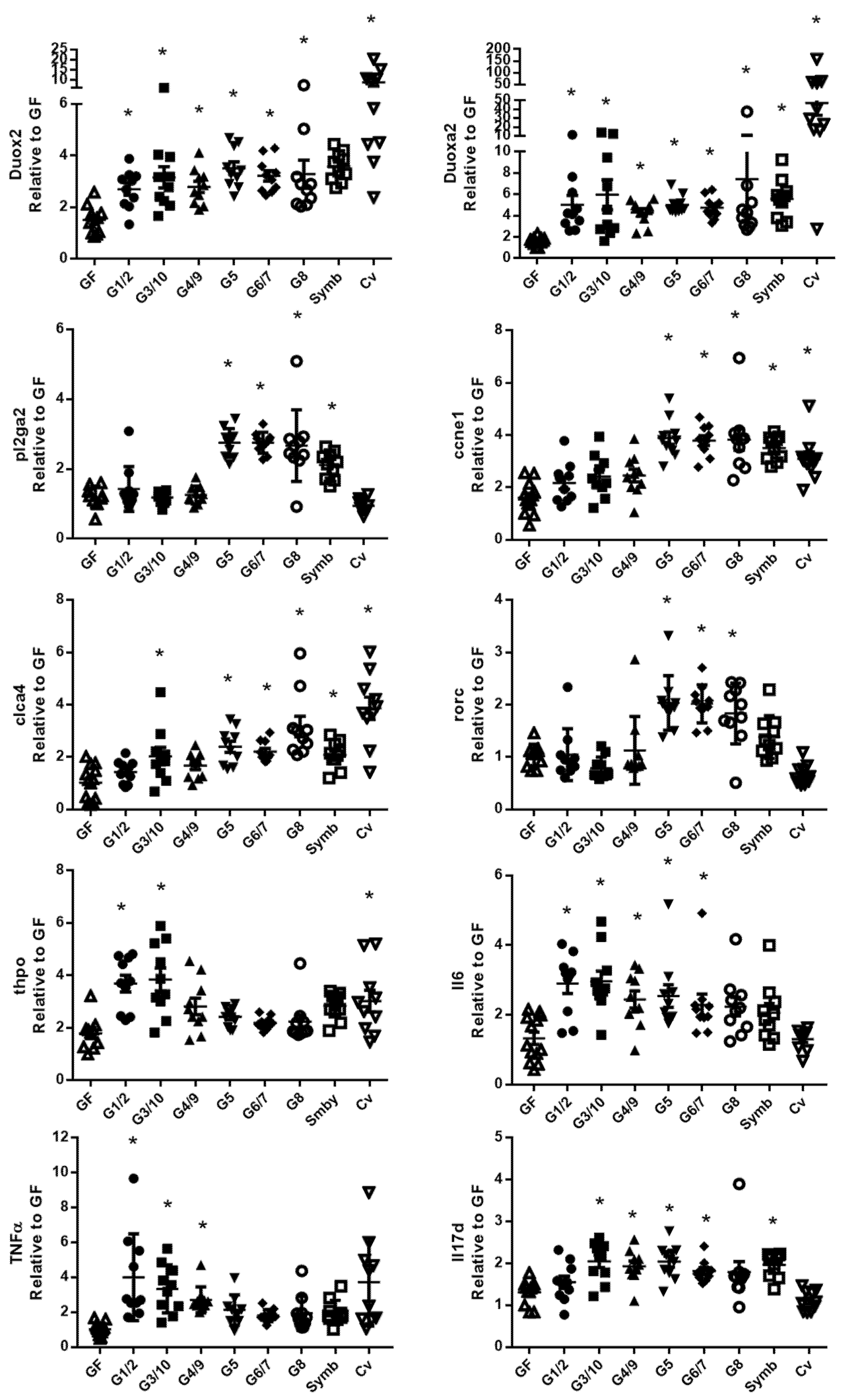
3.1. The Ileal and Colonic Gene Expression Differs according to Which Genotype of Symbioflor2® Is Used
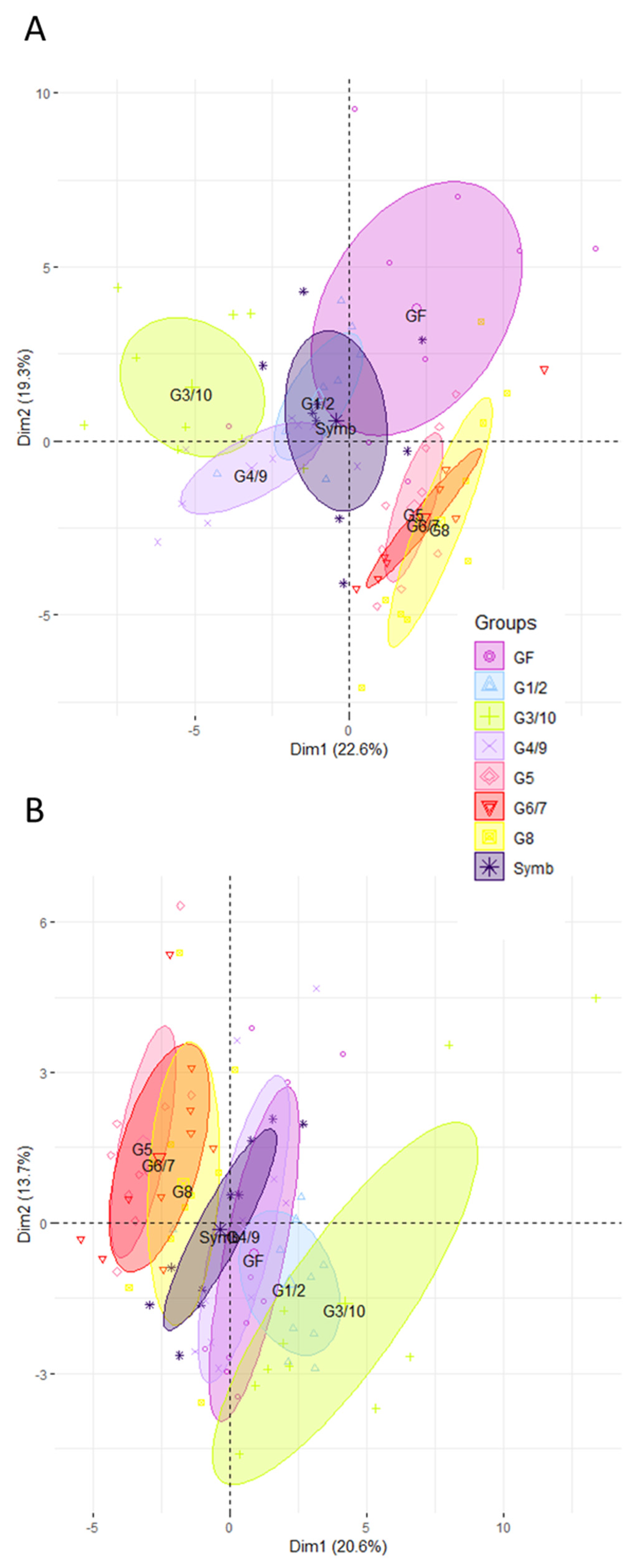
3.2. The Intestinal Gene Expression Profile of Mice Associated with the Symbioflor2® Preparation Is Distinct to That of GF but There Is No Predominance of Any Single Genotype
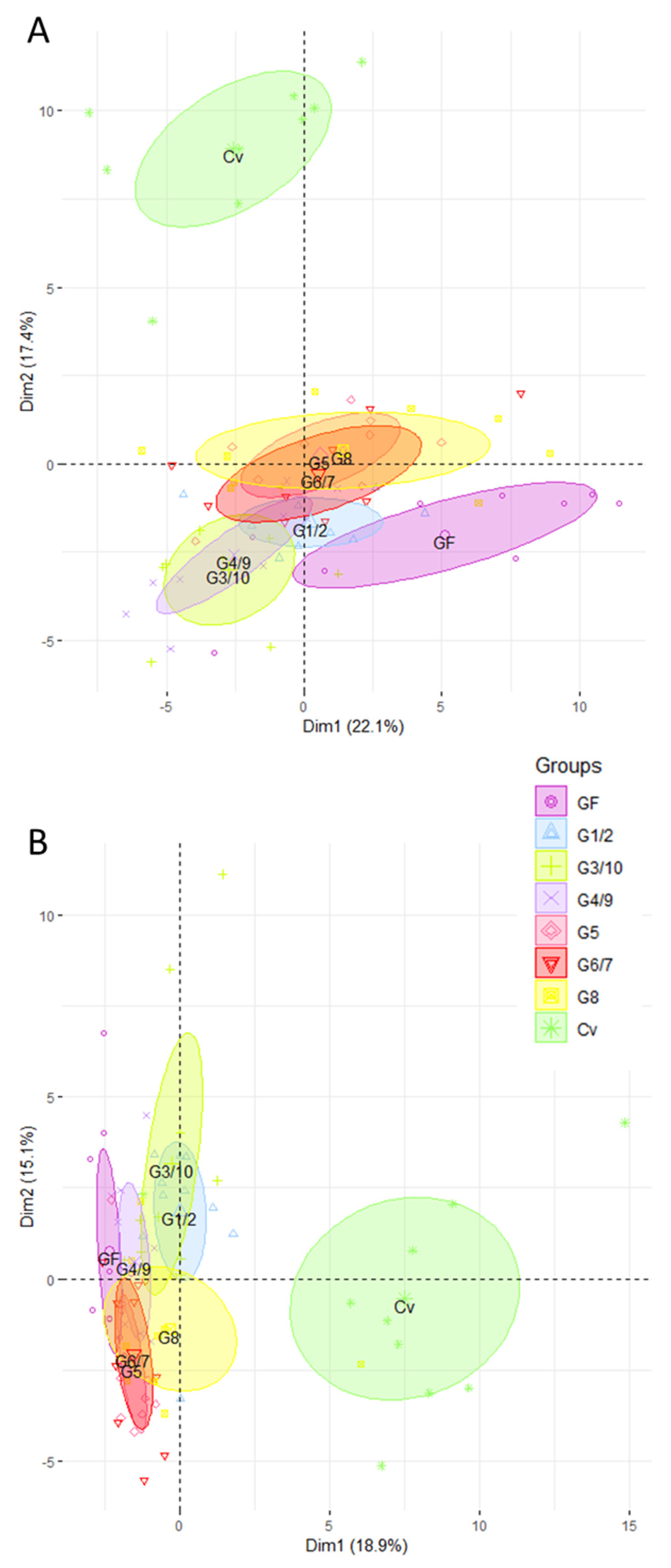
3.3. The Ileal and Colonic Gene Expression Profiles of Mono-Associated Mice Converge toward the CV Profile
3.4. The Nissle Strain and the Six Bacterial Genotypes of Symbioflor2® Have Different Effects on Intestinal Gene Expression Profile, Particularly in the Colon
3.5. The Six Bacterial Strains of Symbioflor® 2 Modulate Genes Involved in Several Cell Functions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beimfohr, C. A Review of Research Conducted with Probiotic E. coli Marketed as Symbioflor. Int. J. Bacteriol. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Wassenaar, T.M. Insights from 100 Years of Research with Probiotic E. Coli. Eur. J. Microbiol. Immunol. 2016, 6, 147–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henker, J.; Muller, S.; Laass, M.W.; Schreiner, A.; Schulze, J. Probiotic Escherichia coli Nissle 1917 (EcN) for successful remission maintenance of ulcerative colitis in children and adolescents: An open-label pilot study. Z Gastroenterol. 2008, 46, 874–875. [Google Scholar] [CrossRef]
- Faghihi, A.H.; Agah, S.; Masoudi, M.; Ghafoori, S.M.; Eshraghi, A. Efficacy of Probiotic Escherichia coli Nissle 1917 in Patients with Irritable Bowel Syndrome: A Double Blind Placebo-controlled Randomized Trial. Acta Med. Indones. 2015, 47, 201–208. [Google Scholar]
- Kruis, W.; Chrubasik, S.; Boehm, S.; Stange, C.; Schulze, J. A double-blind placebo-controlled trial to study therapeutic effects of probiotic Escherichia coli Nissle 1917 in subgroups of patients with irritable bowel syndrome. Int. J. Colorectal. Dis. 2012, 27, 467–474. [Google Scholar] [CrossRef] [Green Version]
- Kruis, W.; Fric, P.; Pokrotnieks, J.; Lukas, M.; Fixa, B.; Kascak, M.; Kamm, M.A.; Weismueller, J.; Beglinger, C.; Stolte, M.; et al. Maintaining remission of ulcerative colitis with the probiotic Escherichia coli Nissle 1917 is as effective as with standard mesalazine. Gut 2004, 53, 1617–1623. [Google Scholar] [CrossRef]
- Kruis, W.; Schutz, E.; Fric, P.; Fixa, B.; Judmaier, G.; Stolte, M. Double-blind comparison of an oral Escherichia coli preparation and mesalazine in maintaining remission of ulcerative colitis. Aliment. Pharmacol. Ther. 1997, 11, 853–858. [Google Scholar] [CrossRef]
- Rembacken, B.J.; Snelling, A.M.; Hawkey, P.M.; Chalmers, D.M.; Axon, A.T. Non-pathogenic Escherichia coli versus mesalazine for the treatment of ulcerative colitis: A randomised trial. Lancet 1999, 354, 635–639. [Google Scholar] [CrossRef]
- Harbord, M.; Eliakim, R.; Bettenworth, D.; Karmiris, K.; Katsanos, K.; Kopylov, U.; Kucharzik, T.; Molnar, T.; Raine, T.; Sebastian, S.; et al. Third European Evidence-based Consensus on Diagnosis and Management of Ulcerative Colitis. Part 2: Current Management. J. Crohns. Colitis. 2017, 11, 769–784. [Google Scholar] [CrossRef] [Green Version]
- Enck, P.; Zimmermann, K.; Menke, G.; Klosterhalfen, S. Randomized controlled treatment trial of irritable bowel syndrome with a probiotic E.-coli preparation (DSM17252) compared to placebo. Z Gastroenterol. 2009, 47, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Martens, U.; Enck, P.; Zieseniss, E. Probiotic treatment of irritable bowel syndrome in children. Ger. Med. Sci. 2010, 8. [Google Scholar] [CrossRef]
- Mondel, M.; Schroeder, B.O.; Zimmermann, K.; Huber, H.; Nuding, S.; Beisner, J.; Fellermann, K.; Stange, E.F.; Wehkamp, J. Probiotic E. coli treatment mediates antimicrobial human beta-defensin synthesis and fecal excretion in humans. Mucosal. Immunol. 2009, 2, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Willenbrock, H.; Hallin, P.F.; Wassenaar, T.M.; Ussery, D.W. Characterization of probiotic Escherichia coli isolates with a novel pan-genome microarray. Genome. Biol. 2007, 8, R267. [Google Scholar] [CrossRef] [Green Version]
- Tomas, J.; Wrzosek, L.; Bouznad, N.; Bouet, S.; Mayeur, C.; Noordine, M.L.; Honvo-Houeto, E.; Langella, P.; Thomas, M.; Cherbuy, C. Primocolonization is associated with colonic epithelial maturation during conventionalization. FASEB J. 2013, 27, 645–655. [Google Scholar] [CrossRef]
- Escribano-Vazquez, U.; Verstraeten, S.; Martin, R.; Chain, F.; Langella, P.; Thomas, M.; Cherbuy, C. The commensal Escherichia coli CEC15 reinforces intestinal defences in gnotobiotic mice and is protective in a chronic colitis mouse model. Sci. Rep. 2019, 9, 11431. [Google Scholar] [CrossRef]
- Wassenaar, T.M.; Beimfohr, C.; Geske, T.; Zimmermann, K. Voluntarily exposure to a single, high dose of probiotic Escherichia coli results in prolonged colonisation. Benef. Microbes. 2014, 5, 367–375. [Google Scholar] [CrossRef]
- Tomas, J.; Reygner, J.; Mayeur, C.; Ducroc, R.; Bouet, S.; Bridonneau, C.; Cavin, J.B.; Thomas, M.; Langella, P.; Cherbuy, C. Early colonizing Escherichia coli elicits remodeling of rat colonic epithelium shifting toward a new homeostatic state. ISME J. 2015, 9, 46–58. [Google Scholar] [CrossRef] [Green Version]
- Ha, E.M.; Oh, C.T.; Bae, Y.S.; Lee, W.J. A direct role for dual oxidase in Drosophila gut immunity. Science 2005, 310, 847–850. [Google Scholar] [CrossRef]
- Yao, Z.; Wang, A.; Li, Y.; Cai, Z.; Lemaitre, B.; Zhang, H. The dual oxidase gene BdDuox regulates the intestinal bacterial community homeostasis of Bactrocera dorsalis. ISME J. 2016, 10, 1037–1050. [Google Scholar] [CrossRef] [Green Version]
- Hoeven, R.; McCallum, K.C.; Cruz, M.R.; Garsin, D.A. Ce-Duox1/BLI-3 generated reactive oxygen species trigger protective SKN-1 activity via p38 MAPK signaling during infection in C. elegans. PLoS Pathog. 2011, 7, e1002453. [Google Scholar]
- Aerssens, J.; Camilleri, M.; Talloen, W.; Thielemans, L.; Gohlmann, H.W.; Van Den Wyngaert, I.; Thielemans, T.; De Hoogt, R.; Andrews, C.N.; Bharucha, A.E.; et al. Alterations in mucosal immunity identified in the colon of patients with irritable bowel syndrome. Clin. Gastroenterol. Hepatol. 2008, 6, 194–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haberman, Y.; Tickle, T.L.; Dexheimer, P.J.; Kim, M.O.; Tang, D.; Karns, R.; Baldassano, R.N.; Noe, J.D.; Rosh, J.; Markowitz, J.; et al. Pediatric Crohn disease patients exhibit specific ileal transcriptome and microbiome signature. J. Clin. Invest. 2014, 124, 3617–3633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanai, H.; Ben-Shachar, S.; Baram, L.; Elad, H.; Gitstein, G.; Brazowski, E.; Tulchinsky, H.; Pasmanik-Chor, M.; Dotan, I. Gene expression alterations in ulcerative colitis patients after restorative proctocolectomy extend to the small bowel proximal to the pouch. Gut 2015, 64, 756–764. [Google Scholar] [CrossRef] [PubMed]
- Grasberger, H.; Gao, J.; Nagao-Kitamoto, H.; Kitamoto, S.; Zhang, M.; Kamada, N.; Eaton, K.A.; El-Zaatari, M.; Shreiner, A.B.; Merchant, J.L.; et al. Increased Expression of DUOX2 Is an Epithelial Response to Mucosal Dysbiosis Required for Immune Homeostasis in Mouse Intestine. Gastroenterology 2015, 149, 1849–1859. [Google Scholar] [CrossRef] [Green Version]
- Birchenough, G.M.; Nystrom, E.E.; Johansson, M.E.; Hansson, G.C. A sentinel goblet cell guards the colonic crypt by triggering Nlrp6-dependent Muc2 secretion. Science 2016, 352, 1535–1542. [Google Scholar] [CrossRef] [Green Version]
- Thiagarajah, J.R.; Chang, J.; Goettel, J.A.; Verkman, A.S.; Lencer, W.I. Aquaporin-3 mediates hydrogen peroxide-dependent responses to environmental stress in colonic epithelia. Proc. Natl. Acad. Sci USA 2017, 114, 568–573. [Google Scholar] [CrossRef] [Green Version]
- Bates, J.M.; Akerlund, J.; Mittge, E.; Guillemin, K. Intestinal alkaline phosphatase detoxifies lipopolysaccharide and prevents inflammation in zebrafish in response to the gut microbiota. Cell Host Microbe. 2007, 2, 371–382. [Google Scholar] [CrossRef] [Green Version]
- Ramasamy, S.; Nguyen, D.D.; Eston, M.A.; Alam, S.N.; Moss, A.K.; Ebrahimi, F.; Biswas, B.; Mostafa, G.; Chen, K.T.; Kaliannan, K.; et al. Intestinal alkaline phosphatase has beneficial effects in mouse models of chronic colitis. Inflamm. Bowel. Dis. 2011, 17, 532–542. [Google Scholar] [CrossRef]
- Miquel, S.; Leclerc, M.; Martin, R.; Chain, F.; Lenoir, M.; Raguideau, S.; Hudault, S.; Bridonneau, C.; Northen, T.; Bowen, B.; et al. Identification of metabolic signatures linked to anti-inflammatory effects of Faecalibacterium prausnitzii. mBio 2015, 6, e00300–e00315. [Google Scholar] [CrossRef] [Green Version]
- Wassenaar, T.M.; Zschuttig, A.; Beimfohr, C.; Geske, T.; Auerbach, C.; Cook, H.; Zimmermann, K.; Gunzer, F. Virulence genes in a probiotic E. coli product with a recorded long history of safe use. Eur. J. Microbiol. Immunol. 2015, 5, 81–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zschuttig, A.; Auerbach, C.; Meltke, S.; Eichhorn, C.; Brandt, M.; Blom, J.; Goesmann, A.; Jarek, M.; Scharfe, M.; Zimmermann, K.; et al. Complete Sequence of Probiotic Symbioflor 2 Escherichia coli Strain G3/10 and Draft Sequences of Symbioflor 2 E. coli Strains G1/2, G4/9, G5, G6/7, and G8. Genome. Announc. 2015, 3, e01330–e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escribano-Vazquez, U.; Beimfohr, C.; Bellet, D.; Thomas, M.; Zimmermann, K.; Langella, P.; Cherbuy, C. Symbioflor2® Escherichia coli Genotypes Enhance Ileal and Colonic Gene Expression Associated with Mucosal Defense in Gnotobiotic Mice. Microorganisms 2020, 8, 512. https://doi.org/10.3390/microorganisms8040512
Escribano-Vazquez U, Beimfohr C, Bellet D, Thomas M, Zimmermann K, Langella P, Cherbuy C. Symbioflor2® Escherichia coli Genotypes Enhance Ileal and Colonic Gene Expression Associated with Mucosal Defense in Gnotobiotic Mice. Microorganisms. 2020; 8(4):512. https://doi.org/10.3390/microorganisms8040512
Chicago/Turabian StyleEscribano-Vazquez, Unai, Claudia Beimfohr, Deborah Bellet, Muriel Thomas, Kurt Zimmermann, Philippe Langella, and Claire Cherbuy. 2020. "Symbioflor2® Escherichia coli Genotypes Enhance Ileal and Colonic Gene Expression Associated with Mucosal Defense in Gnotobiotic Mice" Microorganisms 8, no. 4: 512. https://doi.org/10.3390/microorganisms8040512