Whole-Genome Sequencing-Based Characteristics in Extended-Spectrum Beta-Lactamase-Producing Escherichia coli Isolated from Retail Meats in Korea

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Bacteria Isolation
2.2. Antimicrobial Susceptibility Testing and Confirmation of ESBL-ECs
2.3. Whole-Genome Sequencing and Phylogenetic Analysis
2.4. Nucleotide Sequence Accession Numbers
2.5. In silico Molecular Typing and Characterization
2.6. Multilocus Sequence Typing
2.7. Statistical Data Analysis
3. Results
3.1. Prevalence of ESBL-EC from Retail Raw Meat Samples
3.2. AMR of ESBL-ECs
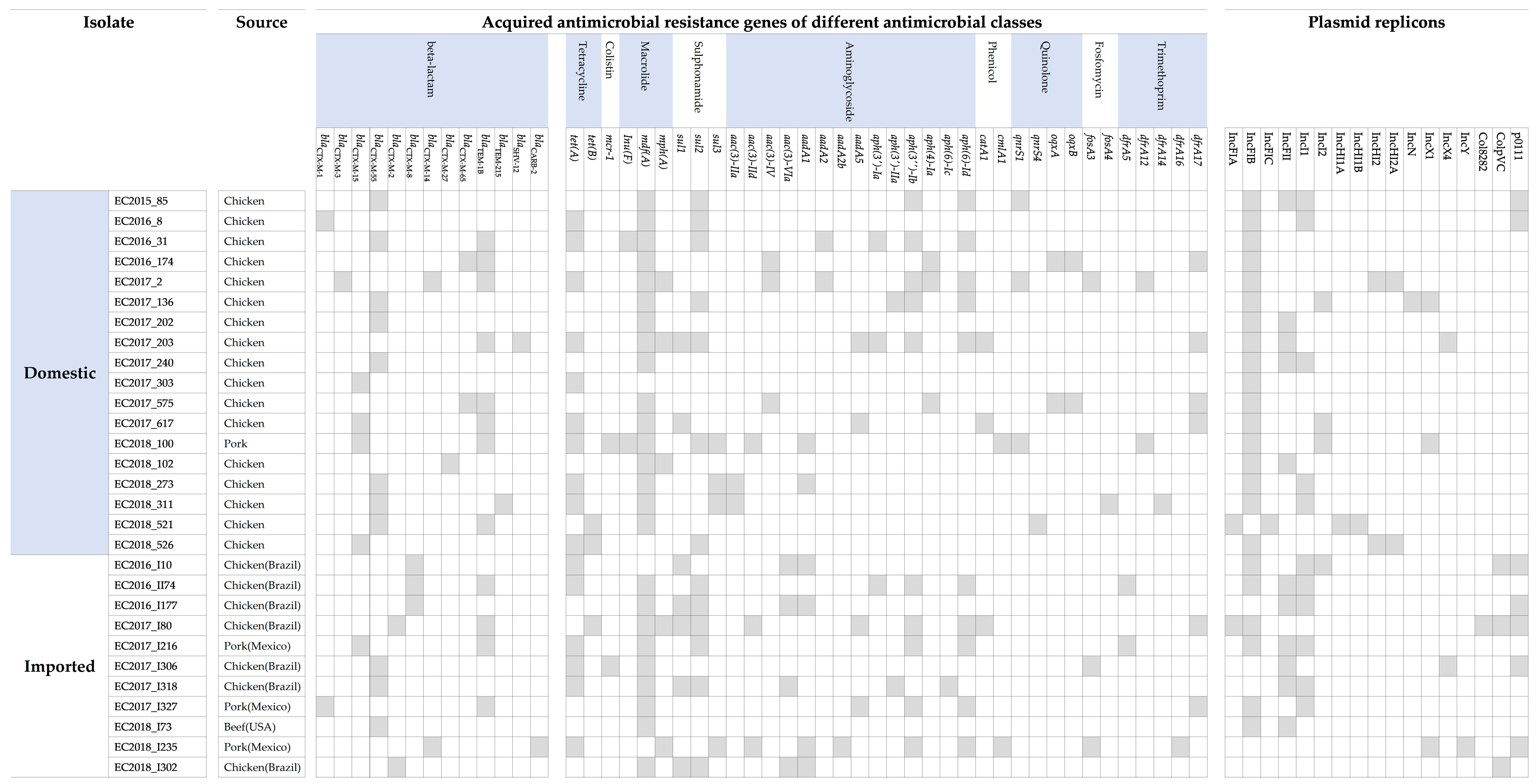
3.3. Distribution of Beta-Lactamase Genes and Plasmid Incompatibility Groups
3.4. STs of ESBL-EC
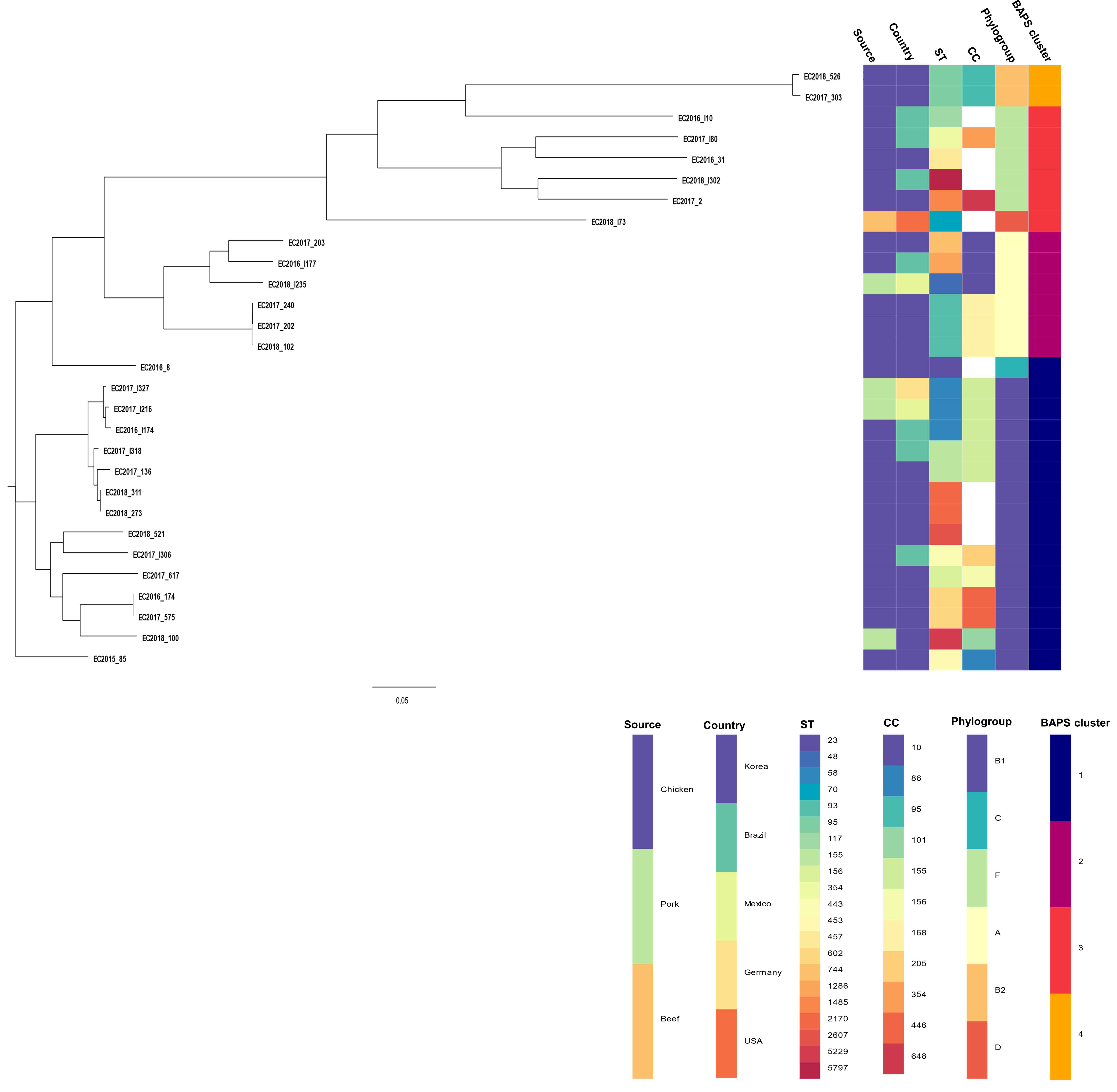
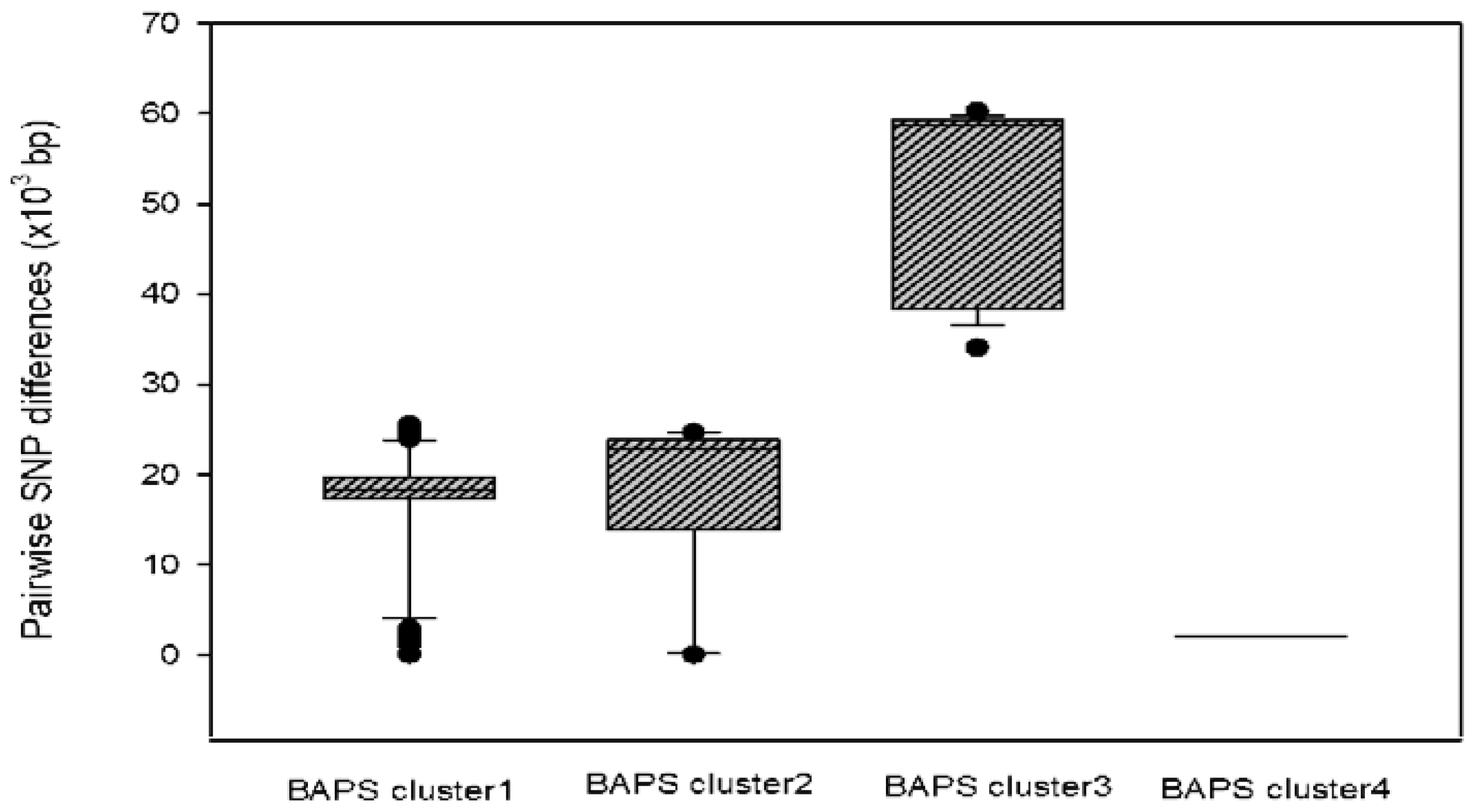
3.5. Pan-Genome, Population Structure and Phylogeny of ESBL-EC
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Croxen, M.A.; Finlay, B.B. Molecular mechanisms of Escherichia coli pathogenicity. Nat. Rev. Microbiol. 2009, 8, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Pouillot, F.; Chomton, M.; Blois, H.; Courroux, C.; Noelig, J.; Bidet, P.; Bingen, E.; Bonacorsi, S. Efficacy of bacteriophage therapy in experimental sepsis and meningitis caused by a clone O25b: H4-ST131 Escherichia coli strain producing CTX-M-15. Antimicrob. Agents Chemother. 2012, 56, 3568–3575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blake, D.; Hillman, K.; Fenlon, D.; Low, J. Transfer of antibiotic resistance between commensal and pathogenic members of the Enterobacteriaceae under ileal conditions. J. Appl. Microbiol. 2003, 95, 428–436. [Google Scholar] [CrossRef]
- Szmolka, A.; Nagy, B. Multidrug resistant commensal Escherichia coli in animals and its impact for public health. Front. Microbiol. 2013, 4, 258. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Integrated Surveillance of Antimicrobial Resistance in Foodborne Bacteria: Application of a One Health Approach: Guidance from the WHO Advisory Group on Integrated Surveillance of Antimicrobial Resistance (AGISAR); WHO: Geneva, Switzerland, 2017. [Google Scholar]
- World Health Organization. Global Priority of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Paterson, D.L.; Bonomo, R.A. Extended-spectrum β-lactamases: A clinical update. Clin. Microbiol. Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.; Uehara, T.; Bernhardt, T.G. Beta-lactam antibiotics induce a lethal malfunctioning of the bacterial cell wall synthesis machinery. Cell 2014, 159, 1300–1311. [Google Scholar] [CrossRef] [Green Version]
- Hawkey, P.M.; Warren, R.E.; Livermore, D.M.; McNulty, C.A.; Enoch, D.A.; Otter, J.A.; Wilson, A.P.R. Treatment of infections caused by multidrug-resistant gram-negative bacteria: Report of the British Society for Antimicrobial Chemotherapy/healthcare Infection Society/British Infection Association Joint Working Party. J. Antimicrob. Chemother. 2018, 73, iii2–iii78. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Baño, J.; Gutiérrez-Gutiérrez, B.; Machuca, I.; Pascual, A. Treatment of infections caused by extended-spectrum-beta-lactamase-, AmpC-, and carbapenemase-producing Enterobacteriaceae. Clin. Microbiol. Rev. 2018, 31. [Google Scholar] [CrossRef] [Green Version]
- Livermore, D.M.; Canton, R.; Gniadkowski, M.; Nordmann, P.; Rossolini, G.M.; Arlet, G.; Ayala, J.; Coque, T.M.; Kern-Zdanowicz, I.; Luzzaro, F.; et al. CTX-M: Changing the face of ESBLs in Europe. J. Antimicrob. Chemother. 2006, 59, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Dolejska, M.; Papagiannitsis, C.C. Plasmid-mediated resistance is going wild. Plasmid 2018, 99, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Doi, Y.; Iovleva, A.; Bonomo, R.A. The ecology of extended-spectrum β-lactamases (ESBLs) in the developed world. J. Travel Med. 2017, 24, S44–S51. [Google Scholar] [CrossRef]
- Valentin, L.; Sharp, H.; Hille, K.; Seibt, U.; Fischer, J.; Pfeifer, Y.; Michael, G.B.; Nickel, S.; Schmiedel, J.; Falgenhauer, L. Subgrouping of ESBL-producing Escherichia coli from animal and human sources: An approach to quantify the distribution of ESBL types between different reservoirs. Int. J. Med. Microbiol. 2014, 304, 805–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Been, M.; Lanza, V.F.; De Toro, M.; Scharringa, J.; Dohmen, W.; Du, Y.; Hu, J.; Lei, Y.; Li, N.; Tooming-Klunderud, A. Dissemination of cephalosporin resistance genes between Escherichia coli strains from farm animals and humans by specific plasmid lineages. PLoS Genet. 2014, 10. [Google Scholar] [CrossRef] [PubMed]
- Liebana, E.; Carattoli, A.; Coque, T.M.; Hasman, H.; Magiorakos, A.P.; Mevius, D.; Peixe, L.; Poirel, L.; Schuepbach-Regula, G.; Torneke, K. Public health risks of enterobacterial isolates producing extended-spectrum β-lactamases or AmpC β-lactamases in food and food-producing animals: An EU perspective of epidemiology, analytical methods, risk factors, and control options. Clin. Infect. Dis. 2012, 56, 1030–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leverstein-van Hall, M.; Dierikx, C.; Cohen Stuart, J.; Voets, G.; Van Den Munckhof, M.; Van Essen-Zandbergen, A.; Platteel, T.; Fluit, A.; Van de Sande-Bruinsma, N.; Scharinga, J. Dutch patients, retail chicken meat and poultry share the same ESBL genes, plasmids and strains. Clin. Microbiol. Infect. 2011, 17, 873–880. [Google Scholar] [CrossRef] [Green Version]
- Lazarus, B.; Paterson, D.L.; Mollinger, J.L.; Rogers, B.A. Do human extraintestinal Escherichia coli infections resistant to expanded-spectrum cephalosporins originate from food-producing animals? A systematic review. Clin. Infect. Dis. 2014, 60, 439–452. [Google Scholar] [CrossRef] [Green Version]
- Holmes, A.H.; Moore, L.S.; Sundsfjord, A.; Steinbakk, M.; Regmi, S.; Karkey, A.; Guerin, P.J.; Piddock, L.J. Understanding the mechanisms and drivers of antimicrobial resistance. Lancet 2016, 387, 176–187. [Google Scholar] [CrossRef]
- Schwarz, S.; Kehrenberg, C.; Walsh, T. Use of antimicrobial agents in veterinary medicine and food animal production. Int. J. Antimicrob. Agents 2001, 17, 431–437. [Google Scholar] [CrossRef]
- Oniciuc, E.A.; Likotrafiti, E.; Alvarez-Molina, A.; Prieto, M.; Santos, J.A.; Alvarez-Ordóñez, A. The present and future of whole genome sequencing (WGS) and whole metagenome sequencing (WMS) for surveillance of antimicrobial resistant microorganisms and antimicrobial resistance genes across the food chain. Genes 2018, 9, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, J.S.; Song, W.; Park, H.M.; Oh, J.Y.; Chae, J.C.; Han, J.I.; Jeong, S.H. First detection of New Delhi metallo-β-Lactamase-5-producing Escherichia coli from companion animals in Korea. Microb. Drug. Resist. 2019, 25, 344–349. [Google Scholar] [CrossRef]
- Lim, J.S.; Choi, D.S.; Kim, Y.J.; Chon, J.W.; Kim, H.S.; Park, H.J.; Moon, J.S.; Wee, S.H.; Seo, K.H. Characterization of Escherichia coli–producing extended-spectrum β-lactamase (ESBL) Isolated from chicken slaughterhouses in South Korea. Foodborne Pathog. Dis. 2015, 12, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Kim, J.; Kim, S.J.; Jeon, S.E.; Oh, K.H.; Cho, S.H.; Kang, Y.H.; Han, S.Y.; Chung, G.T. Characterization of CTX-M-type extended-spectrum beta-lactamase-producing diarrheagenic Escherichia coli isolates in the Republic of Korea during 2008–2011. J. Microbiol. Biotechnol. 2014, 24, 421–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamang, M.D.; Nam, H.M.; Kim, S.R.; Chae, M.H.; Jang, G.C.; Jung, S.C.; Lim, S.K. Prevalence and molecular characterization of CTX-M β-lactamase–producing Escherichia coli isolated from healthy swine and cattle. Foodborne Pathog. Dis. 2013, 10, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Tamang, M.D.; Nam, H.M.; Gurung, M.; Jang, G.C.; Kim, S.R.; Jung, S.C.; Park, Y.H.; Lim, S.K. Molecular characterization of CTX-M β-lactamase and associated addiction systems in Escherichia coli circulating among cattle, farm workers, and the farm environment. Appl. Environ. Microbiol. 2013, 79, 3898–3905. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Lim, Y.M.; Rheem, I.; Lee, Y.; Lee, J.C.; Seol, S.Y.; Lee, Y.C.; Cho, D.T. CTX-M and SHV-12 β-lactamases are the most common extended-spectrum enzymes in clinical isolates of Escherichia coli and Klebsiella pneumoniae collected from 3 university hospitals within Korea. FEMS Microbiol. Lett. 2005, 245, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.A.; Kim, H.; Choi, M.H.; Seo, Y.H.; Lee, H.; Lee, K. Whole-genome analysis of blaCTX-M-55-carrying Escherichia coli among pigs, farm environment, and farm workers. Ann. Lab. Med. 2020, 40, 180–183. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Kim, J.; Ryu, S.; Jeon, B. Predominance of blaCTX-M-65 and blaCTX-M-55 in extended-spectrum β-lactamase-producing Escherichia coli from raw retail chicken in South Korea. J. Glob. Antimicrob. Resist. 2019, 17, 216–220. [Google Scholar] [CrossRef]
- Kim, Y.J.; Moon, J.S.; Oh, D.H.; Chon, J.W.; Song, B.R.; Lim, J.S.; Heo, E.J.; Park, H.J.; Wee, S.H.; Sung, K. Genotypic characterization of ESBL-producing Escherichia coli from imported meat in South Korea. Food Res. Int. 2018, 107, 158–164. [Google Scholar] [CrossRef]
- Jo, S.J.; Woo, G.J. Molecular characterization of plasmids encoding CTX-M β-lactamases and their associated addiction systems circulating among Escherichia coli from retail chickens, chicken farms, and slaughterhouses in Korea. J. Microbiol. Biotechnol. 2016, 26, 270–276. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing. CLSI Supplement M100, 29th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019. [Google Scholar]
- EUCAST. The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 9.0. Available online: http://www.eucast.org (accessed on 30 January 2019).
- NARMS. The National Antimicrobial Resistance Monitoring System. Antimicrobial Agents Used for Susceptibility Testing for Escherichia coli Isolates. Available online: https://www.cdc.gov/narms/antibiotics-tested.html (accessed on 23 May 2019).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Tettelin, H.; Riley, D.; Cattuto, C.; Medini, D. Comparative genomics: The bacterial pan-genome. Curr. Opin. Microbiol. 2008, 11, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Rasko, D.A.; Rosovitz, M.; Myers, G.S.; Mongodin, E.F.; Fricke, W.F.; Gajer, P.; Crabtree, J.; Sebaihia, M.; Thomson, N.R.; Chaudhuri, R. The pangenome structure of Escherichia coli: Comparative genomic analysis of E. coli commensal and pathogenic isolates. J. Bacteriol. 2008, 190, 6881–6893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nourdin-Galindo, G.; Sánchez, P.; Molina, C.F.; Espinoza-Rojas, D.A.; Oliver, C.; Ruiz, P.; Vargas-Chacoff, L.; Cárcamo, J.G.; Figueroa, J.E.; Mancilla, M.; et al. Comparative pan-genome analysis of Piscirickettsia salmonis reveals genomic divergences within genogroups. Front. Cell. Infect. Microbiol. 2017, 7, 459. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Jia, X.; Yang, J.; Ling, Y.; Zhang, Z.; Yu, J.; Wu, J.; Xiao, J. PanGP: A tool for quickly analyzing bacterial pan-genome profile. Bioinformatics 2014, 30, 1297–1299. [Google Scholar] [CrossRef] [Green Version]
- Page, A.J.; Taylor, B.; Delaney, A.J.; Soares, J.; Seemann, T.; Keane, J.A.; Harris, S.R. SNP-sites: Rapid efficient extraction of SNPs from multi-FASTA alignments. Microb. Genom. 2016, 2. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Cheng, L.; Connor, T.R.; Sirén, J.; Aanensen, D.M.; Corander, J. Hierarchical and spatially explicit clustering of DNA sequences with BAPS software. Mol. Biol. Evol. 2013, 30, 1224–1228. [Google Scholar] [CrossRef]
- Hadfield, J.; Croucher, N.J.; Goater, R.J.; Abudahab, K.; Aanensen, D.M.; Harris, S.R. Phandango: An interactive viewer for bacterial population genomics. Bioinformatics 2017, 34, 292–293. [Google Scholar] [CrossRef] [PubMed]
- Beghain, J.; Bridier-Nahmias, A.; Le Nagard, H.; Denamur, E.; Clermont, O. ClermonTyping: An easy-to-use and accurate in silico method for Escherichia genus strain phylotyping. Microb. Genom. 2018, 4. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.; Ochman, H.; et al. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.W.; Jung, M.; Won, H.G.; Belaynehe, K.M.; Yoon, I.J.; Yoo, H.S. Characteristics of transmissible CTX-M- and CMY-type β-lactamase-producing Escherichia coli isolates collected from pig and chicken farms in South Korea. J. Microbiol. Biotechnol. 2017, 27, 1716–1723. [Google Scholar] [CrossRef]
- Kaesbohrer, A.; Bakran-Lebl, K.; Irrgang, A.; Fischer, J.; Kämpf, P.; Schiffmann, A.; Werckenthin, C.; Busch, M.; Kreienbrock, L.; Hille, K. Diversity in prevalence and characteristics of ESBL/pAmpC producing E. coli in food in Germany. Vet. Microbiol. 2019, 233, 52–60. [Google Scholar] [CrossRef]
- Randall, L.; Lodge, M.; Elviss, N.; Lemma, F.; Hopkins, K.; Teale, C.; Woodford, N. Evaluation of meat, fruit and vegetables from retail stores in five United Kingdom regions as sources of extended-spectrum beta-lactamase (ESBL)-producing and carbapenem-resistant Escherichia coli. Int. J. Food Microbiol. 2017, 241, 283–290. [Google Scholar] [CrossRef]
- Pehlivanlar Önen, S.; Aslantaş, Ö.; Şebnem Yılmaz, E.; Kürekci, C. Prevalence of β-lactamase producing Escherichia coli from retail meat in Turkey. J. Food Sci. 2015, 80, M2023–M2029. [Google Scholar] [CrossRef]
- Ojer-Usoz, E.; González, D.; Vitas, A.I.; Leiva, J.; García-Jalón, I.; Febles-Casquero, A.; De la Soledad Escolano, M. Prevalence of extended-spectrum β-lactamase-producing Enterobacteriaceae in meat products sold in Navarra, Spain. Meat Sci. 2013, 93, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Yoon, E.J.; Kim, D.; Jeong, S.H. Bloodstream infections and carbapenem-resistant Enterobacteriaceae in South Korea. Lancet Infect. Dis. 2019, 19, 931–932. [Google Scholar] [CrossRef]
- Ahn, J.Y.; Song, J.E.; Kim, M.H.; Choi, H.; Kim, J.K.; Ann, H.W.; Kim, J.H.; Jeon, Y.; Jeong, S.J.; Kim, S.B. Risk factors for the acquisition of carbapenem-resistant Escherichia coli at a tertiary care center in South Korea: A matched case-control study. Am. J. Infect. Control 2014, 42, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Van Boeckel, T.P.; Pires, J.; Silvester, R.; Zhao, C.; Song, J.; Criscuolo, N.G.; Gilbert, M.; Bonhoeffer, S.; Laxminarayan, R. Global trends in antimicrobial resistance in animals in low-and middle-income countries. Science 2019, 365, eaaw1944. [Google Scholar] [CrossRef] [Green Version]
- Bevan, E.R.; Jones, A.M.; Hawkey, P.M. Global epidemiology of CTX-M β-lactamases: Temporal and geographical shifts in genotype. J. Antimicrob. Chemother. 2017, 72, 2145–2155. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.S.; Song, W.; Park, H.M.; Oh, J.Y.; Chae, J.C.; Shin, S.; Jeong, S.H. Clonal spread of extended-spectrum cephalosporin-resistant Enterobacteriaceae between companion animals and humans in South Korea. Front. Microbiol. 2019, 10, 1371. [Google Scholar] [CrossRef] [Green Version]
- Na, S.H.; Moon, D.C.; Choi, M.J.; Oh, S.J.; Jung, D.Y.; Sung, E.J.; Kang, H.Y.; Hyun, B.H.; Lim, S.K. Antimicrobial resistance and molecular characterization of extended-spectrum β-lactamase-producing Escherichia coli isolated from ducks in South Korea. Foodborne Pathog. Dis. 2019, 16, 799–806. [Google Scholar] [CrossRef]
- He, D.; Chiou, J.; Zeng, Z.; Liu, L.; Chen, X.; Zeng, L.; Chan, E.W.C.; Liu, J.H.; Chen, S. Residues distal to the active site contribute to enhanced catalytic activity of variant and hybrid β-lactamases derived from CTX-M-14 and CTX-M-15. Antimicrob. Agents Chemother. 2015, 59, 5976–5983. [Google Scholar] [CrossRef] [Green Version]
- Kiratisin, P.; Apisarnthanarak, A.; Saifon, P.; Laesripa, C.; Kitphati, R.; Mundy, L.M. The emergence of a novel ceftazidime-resistant CTX-M extended-spectrum β-lactamase, CTX-M-55, in both community-onset and hospital-acquired infections in Thailand. Diagn. Microbiol. Infect. Dis. 2007, 58, 349–355. [Google Scholar] [CrossRef]
- Nadimpalli, M.; Fabre, L.; Yith, V.; Sem, N.; Gouali, M.; Delarocque-Astagneau, E.; Sreng, N.; Le Hello, S. CTX-M-55-type ESBL-producing Salmonella enterica are emerging among retail meats in Phnom Penh, Cambodia. J. Antimicrob. Chemother. 2018, 74, 342–348. [Google Scholar] [CrossRef]
- Rao, L.; Lv, L.; Zeng, Z.; Chen, S.; He, D.; Chen, X.; Wu, C.; Wang, Y.; Yang, T.; Wu, P. Increasing prevalence of extended-spectrum cephalosporin-resistant Escherichia coli in food animals and the diversity of CTX-M genotypes during 2003–2012. Vet. Microbiol. 2014, 172, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Zeng, Z.; Chen, S.; Liu, Y.; Yao, Q.; Deng, Y.; Chen, X.; Lv, L.; Zhuo, C.; Chen, Z. Prevalence and characterisation of CTX-M β-lactamases amongst Escherichia coli isolates from healthy food animals in China. Int. J. Antimicrob. Agents 2012, 39, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, W.; Ohsaki, Y.; Taniguchi, Y.; Koide, S.; Kawamura, K.; Suzuki, M.; Kimura, K.; Wachino, J.-I.; Nagano, Y.; Arakawa, Y. High prevalence of blaCTX-M-14 among genetically diverse Escherichia coli recovered from retail raw chicken meat portions in Japan. Int. J. Food Microbiol. 2018, 284, 98–104. [Google Scholar] [CrossRef]
- Mathers, A.J.; Peirano, G.; Pitout, J.D. The role of epidemic resistance plasmids and international high-risk clones in the spread of multidrug-resistant Enterobacteriaceae. Clin. Microbiol. Rev. 2015, 28, 565–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musicha, P.; Feasey, N.A.; Cain, A.K.; Kallonen, T.; Chaguza, C.; Peno, C.; Khonga, M.; Thompson, S.; Gray, K.J.; Mather, A.E. Genomic landscape of extended-spectrum β-lactamase resistance in Escherichia coli from an urban African setting. J. Antimicrob. Chemother. 2017, 72, 1602–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.C.; Lee, K.; Kim, Y.O.; Won, S.; Chun, J. Large-scale genomics reveals the genetic characteristics of seven species and importance of phylogenetic distance for estimating pan-genome size. Front. Microbiol. 2019, 10, 834. [Google Scholar] [CrossRef]
- Nguyen, V.T.; Jamrozy, D.; Matamoros, S.; Carrique-Mas, J.J.; Ho, H.M.; Thai, Q.H.; Nguyen, T.N.M.; Wagenaar, J.A.; Thwaites, G.; Parkhill, J. Limited contribution of non-intensive chicken farming to ESBL-producing Escherichia coli colonization in humans in Vietnam: An epidemiological and genomic analysis. J. Antimicrob. Chemother. 2019, 74, 561–570. [Google Scholar] [CrossRef] [Green Version]
- Clermont, O.; Christenson, J.K.; Daubié, A.S.; Gordon, D.M.; Denamur, E. Development of an allele-specific PCR for Escherichia coli B2 sub-typing, a rapid and easy to perform substitute of multilocus sequence typing. J. Microbiol. Methods 2014, 101, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Mora, A.; García-Peña, F.J.; Alonso, M.P.; Pedraza-Diaz, S.; Ortega-Mora, L.M.; Garcia-Parraga, D.; López, C.; Viso, S.; Dahbi, G.; Marzoa, J.; et al. Impact of human-associated Escherichia coli clonal groups in antarctic pinnipeds: Presence of ST73, ST95, ST141 and ST131. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Manges, A.R. Escherichia coli and urinary tract infections: The role of poultry-meat. Clin. Microbiol. Infect. 2016, 22, 122–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, M.J.; Hopkins, K.L.; Wareham, D.W.; Toleman, M.A.; Elviss, N.; Randall, L.; Teale, C.; Cleary, P.; Wiuff, C.; Doumith, M. Extended-spectrum β-lactamase-producing Escherichia coli in human-derived and foodchain-derived samples from England, Wales, and Scotland: An epidemiological surveillance and typing study. Lancet Infect. Dis. 2019, 19, 1325–1335. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.; Kim, J.; Seo, M.R.; Wie, S.-H.; Cho, Y.; Lim, S.K.; Lee, J.; Kwon, K.; Lee, H.; Cheong, H.J. Clinical characteristics of community-acquired acute pyelonephritis caused by ESBL-producing pathogens in South Korea. Infection 2013, 41, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Villa, M.F.L.; Salinas, L.; Villavivencio, F.; Rafael, T.; Salas, S.; Rivera, R.; Villacis, J.; Satan, C.; Ushiña, L.; Muñoz, O. Diverse Escherichia coli lineages, from domestic animals and humans in a household, carry colistin resistance gene mcr-1 in Ecuador. bioRxiv 2018. [Google Scholar] [CrossRef]
- Van Hoek, A.H.; Veenman, C.; Florijn, A.; Huijbers, P.M.; Graat, E.A.; De Greeff, S.; Dierikx, C.M.; Van Duijkeren, E. Longitudinal study of ESBL Escherichia coli carriage on an organic broiler farm. J. Antimicrob. Chemother. 2018, 73, 3298–3304. [Google Scholar] [CrossRef]
- Skurnik, D.; Clermont, O.; Guillard, T.; Launay, A.; Danilchanka, O.; Pons, S.; Diancourt, L.; Lebreton, F.; Kadlec, K.; Roux, D. Emergence of antimicrobial-resistant Escherichia coli of animal origin spreading in humans. Mol. Biol. Evol. 2016, 33, 898–914. [Google Scholar] [CrossRef] [Green Version]
- Choi, M.J.; Lim, S.K.; Jung, S.C.; Ko, K.S. Comparisons of CTX-M-producing Escherichia coli isolates from humans and animals in South Korea. J. Bacteriol. Virol. 2014, 44, 44–51. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Category | ESBL-EC Positive Rate, % (No. of ESBL-EC Confirmed Samples/ No. of Tested Samples) | |||
|---|---|---|---|---|
| Domestic | Imported | Total | p-Value ** | |
| Beef | 0.0 (0/612) | 0.1 (1/678) | 0.1 (1/ 1290) | 1.0000 |
| Pork | 0.2 (1/565) | 0.5 (3/561) | 0.4 (4/1126) | 0.6114 |
| Chicken | 3.0 (17/560) * | 2.7 (7/258) * | 2.9 (24/818) * | 0.9752 |
| Total | 1.0 (18/1737) | 0.7 (11/1497) | 0.9 (29/3234) | 0.4736 |
| Antibiotics | Resistance Rate, % (No. of Resistant Strains/ No. of Tested ESBL-ECs) | ||||
|---|---|---|---|---|---|
| Domestic | Imported | Total | p-Value * | ||
| Beta-lactams | AmC | 0.0 (0/18) | 0.0 (0/11) | 0.0 (0/29) | NA |
| AMP | 100.0 (18/18) | 100.0 (11/11) | 100.0 (29/29) | 1.0000 | |
| CAZ | 22.2 (4/18) | 0.0 (0/11) | 13.8 (4/29) | 0.2589 | |
| CTF | 100.0 (18/18) | 100.0 (11/11) | 100.0 (29/29) | 1.0000 | |
| FEP | 16.7 (3/18) | 9.1 (1/11) | 13.8 (4/29) | 0.9847 | |
| FOX | 0.0 (0/18) | 0.0 (0/11) | 0.0 (0/29) | NA | |
| MEM | 0.0 (0/18) | 0.0 (0/11) | 0.0 (0/29) | NA | |
| Non-beta-lactams | CHL | 50.0 (9/18) | 36.4 (4/11) | 44.8 (13/29) | 0.7401 |
| CIP | 50.0 (9/18) | 45.5 (5/11) | 48.3 (14/29) | 1.0000 | |
| COL | 11.1 (2/18) | 18.2 (2/11) | 13.8 (4/29) | 1.0000 | |
| GEN | 38.9 (7/18) | 45.5 (5/11) | 41.4 (12/29) | 1.0000 | |
| NAL | 83.3 (15/18) | 63.6 (7/11) | 75.9 (22/29) | 0.4499 | |
| STR | 55.6 (10/18) | 63.6 (7/11) | 58.6 (17/29) | 0.9679 | |
| SXT | 38.9 (7/18) | 54.5 (6/11) | 44.8 (13/29) | 0.6615 | |
| TET | 77.8 (14/18) | 63.6( 7/11) | 72.4 (21/29) | 0.6902 | |
| MDR | 100.0 (18/18) | 100.0 (11/11) | 100.0 (29/29) | 1.0000 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.; Kim, H.; Kim, Y.; Kim, M.; Kwak, H.; Ryu, S. Whole-Genome Sequencing-Based Characteristics in Extended-Spectrum Beta-Lactamase-Producing Escherichia coli Isolated from Retail Meats in Korea. Microorganisms 2020, 8, 508. https://doi.org/10.3390/microorganisms8040508
Kim S, Kim H, Kim Y, Kim M, Kwak H, Ryu S. Whole-Genome Sequencing-Based Characteristics in Extended-Spectrum Beta-Lactamase-Producing Escherichia coli Isolated from Retail Meats in Korea. Microorganisms. 2020; 8(4):508. https://doi.org/10.3390/microorganisms8040508
Chicago/Turabian StyleKim, Seokhwan, Hansol Kim, Yonghoon Kim, Migyeong Kim, Hyosun Kwak, and Sangryeol Ryu. 2020. "Whole-Genome Sequencing-Based Characteristics in Extended-Spectrum Beta-Lactamase-Producing Escherichia coli Isolated from Retail Meats in Korea" Microorganisms 8, no. 4: 508. https://doi.org/10.3390/microorganisms8040508