Complete Genome Sequence of Acidithiobacillus ferrooxidans YNTRS-40, a Strain of the Ferrous Iron- and Sulfur-Oxidizing Acidophile

Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions, Genomic DNA Isolation and Morphological Detection
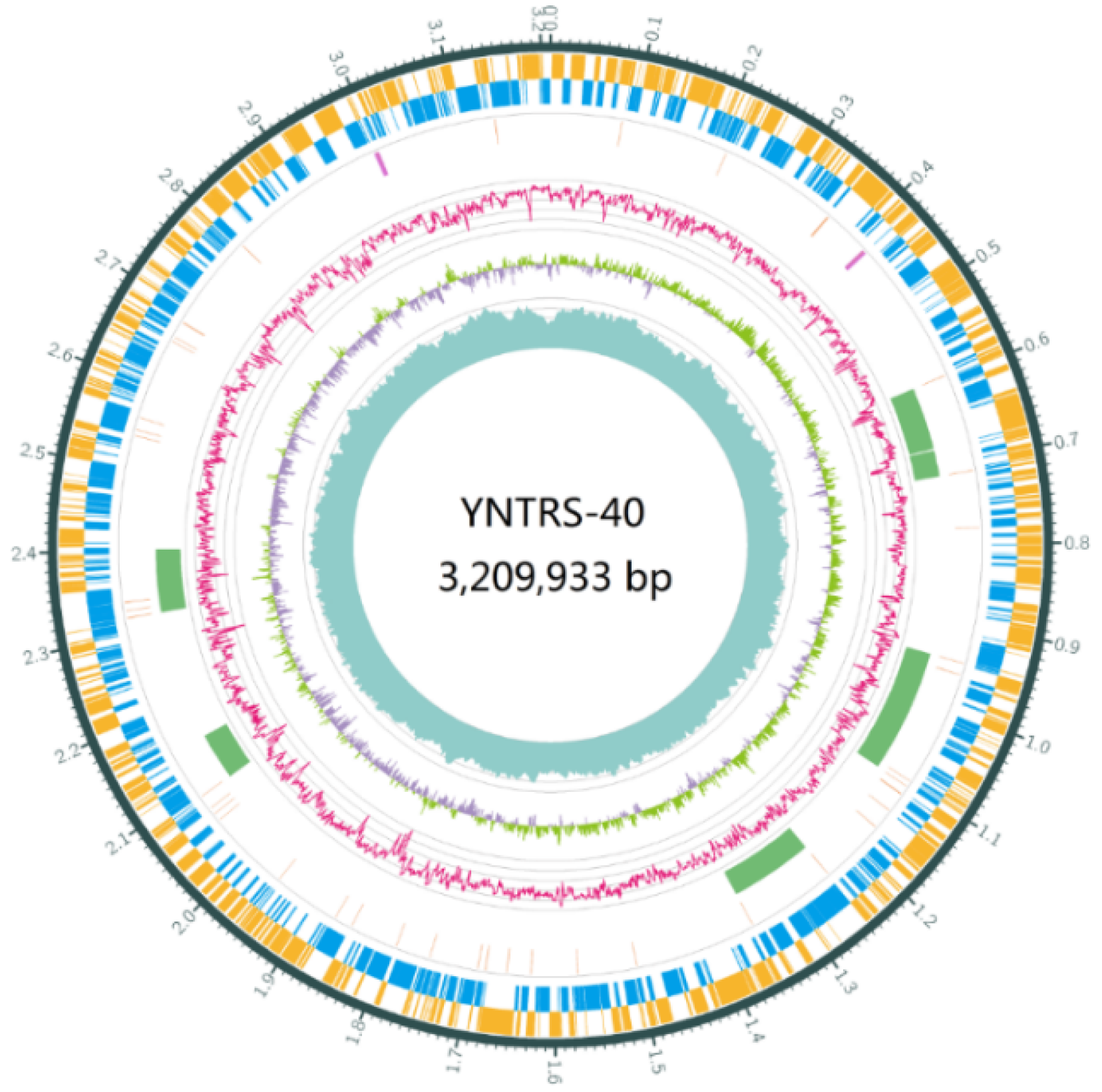
2.2. Genome Sequencing, Assembly and Annotation
3. Results and Discussion
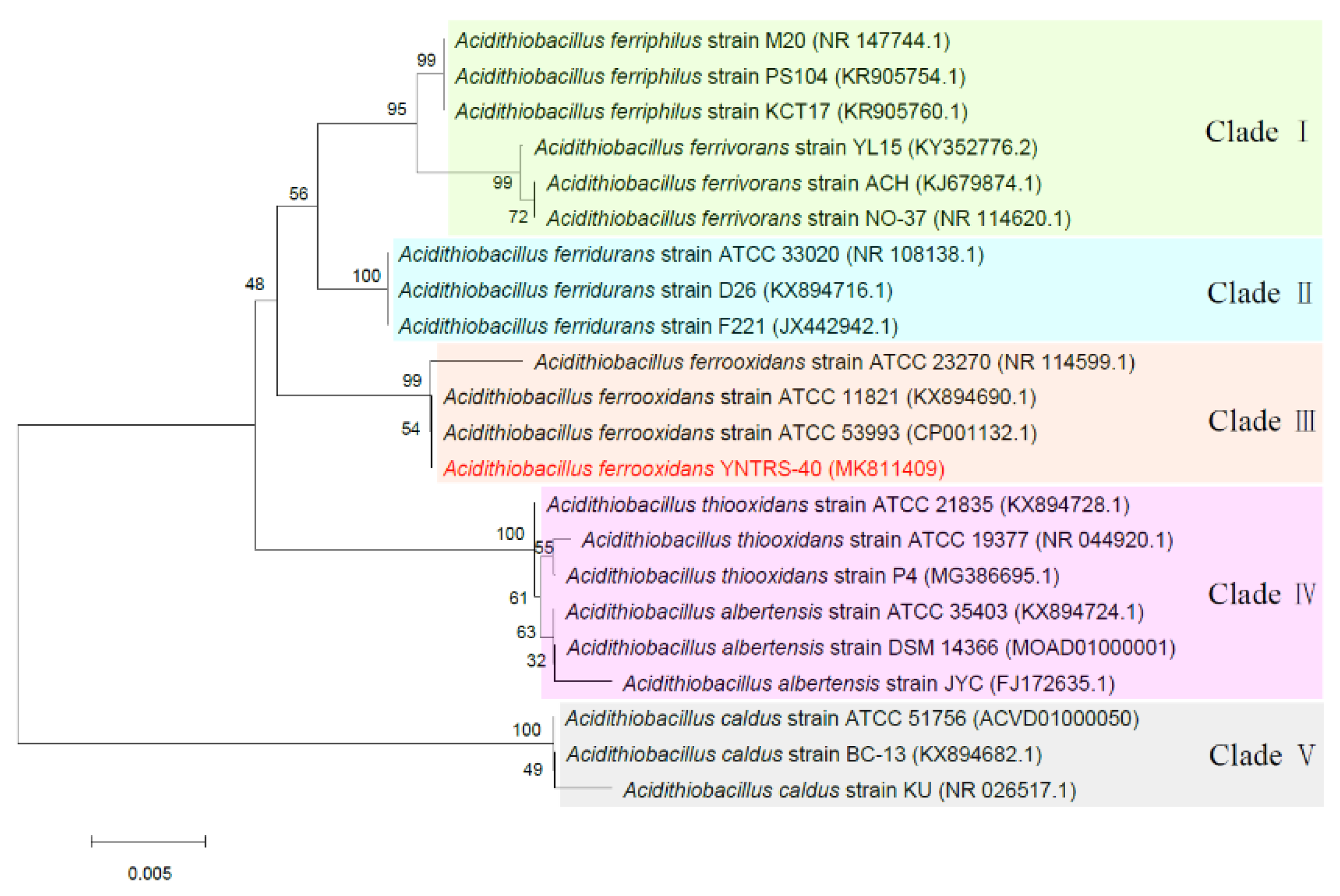
3.1. Genomic Features Related to Adaptation to Diverse Stresses
3.2. Genomic Features Related to the Oxidation of Ferrous Iron and Sulfur
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Colman, D.R.; Poudel, S.; Hamilton, T.L.; Havig, J.R.; Selensky, M.J.; Shock, E.L.; Boyd, E.S. Geobiological feedbacks and the evolution of thermoacidophiles. ISME J. 2018, 12, 225–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuñez, H.; Moya-Beltrán, A.; Covarrubias, P.C.; Issotta, F.; Cárdenas, J.P.; González, M.; Atavales, J.; Acuña, L.G.; Johnson, D.B.; Quatrini, R. Molecular systematics of the genus Acidithiobacillus: Insights into the phylogenetic structure and diversification of the taxon. Front. Microbiol. 2017, 8, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hongbo, Z.; Yisheng, Z.; Xian, Z.; Lu, Q.; Menglin, S.; Yu, Y.; Yansheng, Z.; Jun, W.; Hyunjung, K.; Guanzhou, Q. The dissolution and passivation mechanism of chalcopyrite in bioleaching: An overview. Miner. Eng. 2019, 136, 140–154. [Google Scholar]
- Xian, Z.; Xueduan, L.; Yili, L.; Fenliang, F.; Xiaoxia, Z.; Huaqun, Y. Metabolic diversity and adaptive mechanisms of iron- and/or sulfur-oxidizing autotrophic acidophiles in extremely acidic environments. Env. Microbiol. Rep. 2016, 8, 738–751. [Google Scholar]
- Temple, K.L.; Colmer, A.R. The autotrophic oxidation of iron by a new bacterium: Thiobacillus ferrooxidans. J. Bacteriol. 1951, 62, 605–611. [Google Scholar]
- Falagán, C.; Johnson, D.B. Acidithiobacillus ferriphilus sp. nov., a facultatively anaerobic iron- and sulfur-metabolizing extreme acidophile. Int. J. Syst. Evol. Microbiol. 2016, 66, 206–211. [Google Scholar] [CrossRef] [Green Version]
- Hallberg, K.B.; González-Toril, E.; Johnson, D.B. Acidithiobacillus ferrivorans, sp. nov.; facultatively anaerobic, psychrotolerant iron-, and sulfur-oxidizing acidophiles isolated from metal mine-impacted environments. Extremophiles 2010, 14, 9–19. [Google Scholar] [CrossRef]
- Hedrich, S.; Johnson, D.B. Acidithiobacillus ferridurans sp. nov., an acidophilic iron-, sulfur- and hydrogen-metabolizing chemolithotrophic gammaproteobacterium. Int. J. Syst. Evol. Microbiol. 2013, 63, 4018–4025. [Google Scholar] [CrossRef]
- Bryant, R.D.; McGroarty, K.M.; Costerton, J.W.; Laishley, E.J. Isolation and characterization of a new acidophilic Thiobacillus species (T. albertis). Can. J. Microbiol. 1983, 29, 1159–1170. [Google Scholar] [CrossRef]
- Waksman, S.A.; Joffe, J.S. Microorganisms concerned in the oxidation of sulfur in the soil: II. Thiobacillus Thiooxidans, a new sulfur-oxidizing organism isolated from the soil. J. Bacteriol. 1922, 7, 239–256. [Google Scholar]
- Hallberg, K.B.; Lindström, E.B. Characterization of Thiobacillus caldus sp. nov., a moderately thermophilic acidophile. Microbiology 1994, 140, 3451–3456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falagán, C.; Moya-Beltrán, A.; Castro, M.; Quatrini, R.; Johnson, D.B. Acidithiobacillus sulfuriphilus sp. nov.: An extremely acidophilic sulfur-oxidizing chemolithotroph isolated from a neutral pH environment. Int. J. Syst. Evol. Microbiol. 2019, 69, 2907–2913. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.P.; Wood, A.P. Reclassification of some species of Thiobacillus to the newly designated genera Acidithiobacillus gen. nov., Halothiobacillus gen. nov. and Thermithiobacillus gen. nov. Int. J. Syst. Evol. Microbiol. 2000, 50, 511–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitschke, W.; Bonnefoy, V. Energy acquisition in low pH environments. In Acidophiles: Life in Extremely Acidic Environments; Quatrini, R., Johnson, D.B., Eds.; Caister Academic Press: Norfolk, UK, 2016; pp. 19–48. [Google Scholar]
- Davis, R.A., Jr.; Welty, A.T.; Borrego, J.; Morales, J.A.; Pendon, J.G.; Ryan, J.G. Rio Tinto estuary (Spain): 5000 years of pollution. Environ. Geol. 2000, 39, 1107–1116. [Google Scholar] [CrossRef]
- González-Toril, E.; Llobet-Brossa, E.; Casamayor, E.O.; Amann, R.; Amils, R. Microbial ecology of an extreme acidic environment, the Tinto River. Appl. Environ. Microbiol. 2003, 69, 4853–4865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawlings, D.E. Heavy metal mining using microbes. Annu. Rev. Microbiol. 2002, 56, 65–91. [Google Scholar] [CrossRef]
- Rawlings, D.E. Characteristics and adaptability of iron- and sulfur-oxidizing microorganisms used for the recovery of metals from minerals and their concentrates. Microb. Cell Fact. 2005, 4, 13. [Google Scholar] [CrossRef] [Green Version]
- Sydow, A.; Krieg, T.; Mayer, F.; Schrader, J.; Holtmann, D. Electroactive bacteria—Molecular mechanisms and genetic tools. Appl. Microbiol. Biotechnol. 2014, 98, 8481–8495. [Google Scholar] [CrossRef]
- Valdés, J.; Pedroso, I.; Quatrini, R.; Dodson, R.J.; Tettelin, H.; Blake, R.; Eisen, J.A.; Holmes, D.S. Acidithiobacillus ferrooxidans metabolism: From genome sequence to industrial applications. BMC Genom. 2008, 9, 597. [Google Scholar] [CrossRef] [Green Version]
- Shuang, Z.; Lei, Y.; Weijia, X.; Peng, C.; Yu, Z.; Weidong, W. Acidithiobacillus ferrooxidans and its potential application. Extremophiles 2018, 22, 563–579. [Google Scholar]
- Lei, Y.; Shuang, Z.; Weidong, W.; Huixin, H.; Yanjie, W.; Gaobo, Y.; Peng, C. Draft genome sequence of Acidithiobacillus ferrooxidans YQH-1. Genom. Data 2015, 6, 269–270. [Google Scholar]
- Peng, C.; Lei, Y.; Zhengrong, W.; Ruixiang, X.; Suyue, L.; Ningbo, W.; Ning, L.; Hongyu, L. Draft genome sequence of extremely acidophilic bacterium Acidithiobacillus ferrooxidans DLC-5 isolated from acid mine drainage in Northeast China. Genom. Data 2015, 6, 267–268. [Google Scholar]
- Srivastava, S.; Briggs, B.R.; Dong, H. Abundance and taxonomic affiliation of molybdenum transport and utilization genes in Tengchong hot springs, China. Environ. Microbiol. 2018, 20, 2397–2409. [Google Scholar] [CrossRef] [PubMed]
- Senol Cali, D.; Kim, J.S.; Ghose, S.; Alkan, C.; Mutlu, O. Nanopore sequencing technology and tools for genome assembly: Computational analysis of the current state, bottlenecks and future directions. Brief. Bioinform. 2018, 20, 1542–1559. [Google Scholar] [CrossRef] [PubMed]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Hyatt, D.; Gwo-Liang, C.; LoCascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [Green Version]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, 54–57. [Google Scholar] [CrossRef]
- Lagesen, K.; Hallin, P.; Rødland, E.A.; Stærfeldt, H.-H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. Data, information, knowledge and principle: Back to metabolism in KEGG. Nucleic Acids Res. 2013, 42, 199–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galperin, M.Y.; Makarova, K.S.; Wolf, Y.I.; Koonin, E.V. Expanded microbial genome coverage and improved protein family annotation in the COG database. Nucleic Acids Res. 2014, 43, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics. 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Michael, R.; Ramon, R.M.; Frank, O.G.; Jörg, P. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar]
- Tatusov, R.L.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Kiryutin, B.; Koonin, E.V.; Krylov, D.M.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; et al. The COG database: An updated version includes eukaryotes. BMC Bioinform. 2003, 4, 41. [Google Scholar] [CrossRef] [Green Version]
- Xian, Z.; Xueduan, L.; Liangzhi, L.; Guanyun, W.; Danli, Z.; Yili, L.; Bo, M. Phylogeny, divergent evolution, and speciation of sulfur-oxidizing Acidithiobacillus populations. BMC Genom. 2019, 20, 438. [Google Scholar]
- Jensen, A.B.; Webb, C. Ferrous sulphate oxidation using thiobacillus ferrooxidans: A review. Process. Biochem. 1995, 30, 225–236. [Google Scholar] [CrossRef]
- Wenbin, N.; Dejuan, Z.; Feifan, L.; Lei, Y.; Peng, C.; Xiaoxuan, Y.; Hongyu, L. Quorum-sensing system in Acidithiobacillus ferrooxidans involved in its resistance to Cu2+. Lett. Appl. Microbiol. 2011, 53, 84–91. [Google Scholar] [CrossRef]
- Xu, Y.; Yang, M.; Yao, T.; Xiong, H. Isolation, identification and arsenic-resistance of Acidithiobacillus ferrooxidans HX3 producing schwertmannite. J. Environ. Sci. 2014, 26, 1463–1470. [Google Scholar] [CrossRef]
- Jones, P.; Binns, D.; Hsin-Yu, C.; Fraser, M.; Weizhong, L.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almárcegui, R.J.; Navarro, C.A.; Paradela, A.; Albar, J.P.; von Bernath, D.; Jerez, C.A. Response to copper of Acidithiobacillus ferrooxidans ATCC 23270 grown in elemental sulfur. Res. Microbiol. 2014, 165, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Sugio, T.; Iwahori, K.; Takeuchi, F.; Negishi, A.; Maeda, T.; Kamimura, K. Cytochrome c oxidase purified from a mercury-resistant strain of Acidithiobacillus ferrooxidans volatilizes mercury. J. Biosci. Bioeng. 2001, 92, 44–49. [Google Scholar] [CrossRef]
- Chengying, J.; Liu, L.; Xu, G.; Xiaoyan, Y.; Shuangjiang, L.; Poetsch, A. Resolution of carbon metabolism and sulfur-oxidation pathways of Metallosphaera cuprina Ar-4 via comparative proteomics. J. Proteom. 2014, 109, 276–289. [Google Scholar]
- Yue, Z.; Mengran, Y.; Shuang, Z.; Dan, Z.; Jiangong, D.; Weidong, W.; Lei, Y. Iron and sulfur oxidation pathways of Acidithiobacillus ferrooxidans. World J. Microbiol. Biotechnol. 2019, 35, 60. [Google Scholar]
- Ruiqi, Y.; Guangxiu, L.; Tuo, C.; Wei, Z.; Gaosen, Z.; Sijing, C. The complete genomic sequence of a novel cold-adapted bacterium, Planococcus maritimus Y42, isolated from crude oil-contaminated soil. Stand. Genom. Sci. 2018, 13, 23. [Google Scholar]
- Mochizuki, S.; Hiratsu, K.; Suwa, M.; Ishii, T.; Sugino, F.; Yamada, K.; Kinashi, H. The large linear plasmid pSLA2-L of Streptomyces rochei has an unusually condensed gene organization for secondary metabolism. Mol. Microbiol. 2003, 48, 1501–1510. [Google Scholar] [CrossRef]
- Barrie Johnson, D.; Hallberg, K.B. Carbon, iron and sulfur metabolism in acidophilic micro-organisms. Adv. Microbiol. Physiol. 2008, 54, 201–255. [Google Scholar]
- Ilbert, M.; Bonnefoy, V. Insight into the evolution of the iron oxidation pathways. Biochim. Biophys. Acta 2013, 1827, 161–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarzabal, A.; Appia-Ayme, C.; Ratouchniak, J.; Bonnefoy, V. Regulation of the expression of the Acidithiobacillus ferrooxidans rus operon encoding two cytochromes c, a cytochrome oxidase and rusticyanin. Microbiology 2004, 150, 2113–2123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quatrini, R.; Jedlicki, E.; Holmes, D.S. Genomic insights into the iron uptake mechanisms of the biomining microorganism Acidithiobacillus ferrooxidans. J. Ind. Microbiol. Biotechnol. 2005, 32, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Yarzábal, A.; Duquesne, K.; Bonnefoy, V. Rusticyanin gene expression of Acidithiobacillus ferrooxidans ATCC 33020 in sulfur- and in ferrous iron media. Hydrometallurgy 2003, 71, 107–114. [Google Scholar] [CrossRef]
- Brasseur, G.; Bruscella, P.; Bonnefoy, V.; Lemesle-Meunier, D. The bc1 complex of the iron-grown acidophilic chemolithotrophic bacterium Acidithiobacillus ferrooxidans functions in the reverse but not in the forward direction: Is there a second bc1 complex? Biochim. Biophys. Acta 2002, 1555, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Quatrini, R.; Appia-Ayme, C.; Denis, Y.; Jedlicki, E.; Holmes, D.S.; Bonnefoy, V. Extending the models for iron and sulfur oxidation in the extreme acidophile Acidithiobacillus ferrooxidans. BMC Genom. 2009, 10, 394. [Google Scholar] [CrossRef] [Green Version]
- Findlay, A.J.; Kamyshny, A. Turnover rates of intermediate sulfur species (Sx2-, S0, S2O32-, S4O62-, SO32-) in anoxic freshwater and sediments. Front. Microbiol. 2017, 8, 2551. [Google Scholar] [CrossRef]
- Mangold, S.; Valdés, J.; Holmes, D.; Dopson, M. Sulfur metabolism in the extreme acidophile Acidithiobacillus caldus. Front. Microbiol. 2011, 2, 17. [Google Scholar] [CrossRef] [Green Version]
- Quatrini, R.; Appia-Ayme, C.; Denis, Y.; Ratouchniak, J.; Veloso, F.; Valdes, J.; Lefimil, C.; Silver, S.; Roberto, F.; Orellana, O.; et al. Insights into the iron and sulfur energetic metabolism of Acidithiobacillus ferrooxidans by microarray transcriptome profiling. Hydrometallurgy 2006, 83, 263–272. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Attribute | Value | % of Total 1 |
|---|---|---|
| Genome size (bp) | 3,257,037 | 100.00 |
| DNA coding (bp) | 2,940,490 | 90.28 |
| DNA G + C (bp) | 1,905,651 | 58.51 |
| DNA scaffolds | 2 | 100.00 |
| Total genes | 3419 | 100.00 |
| Protein coding genes | 3349 | 97.95 |
| RNA genes | 70 | 2.05 |
| Pseudo genes | NA 2 | NA 2 |
| Genes in internal clusters | 8 | 16.21 |
| Genes with function prediction | 1692 | 50.52 |
| Genes assigned to COGs | 1793 | 53.54 |
| Genes with Pfam domains | 2539 | 75.81 |
| Genes with signal peptides | NA 2 | NA 2 |
| Genes with transmembrane helices | NA 2 | NA 2 |
| CRISPR repeats | 0 | 0 |
| Code | Value | % Age 1 | Description |
|---|---|---|---|
| J | 172 | 5.14 | Translation, ribosomal structure, and biogenesis |
| A | 0 | 0 | RNA processing and modification |
| K | 118 | 3.52 | Transcription |
| L | 126 | 3.76 | Replication, recombination, and repair |
| B | 1 | 0.03 | Chromatin structure and dynamics |
| D | 29 | 0.87 | Cell cycle control, Cell division, chromosome partitioning |
| Y | 0 | 0 | Nuclear structure |
| V | 90 | 2.69 | Defense mechanisms |
| T | 87 | 2.60 | Signal transduction mechanisms |
| M | 162 | 4.84 | Cell wall/membrane/envelope biogenesis |
| N | 20 | 0.60 | Cell motility |
| Z | 0 | 0 | Cytoskeleton |
| W | 14 | 0.42 | Extracellular structures |
| U | 62 | 1.85 | Intracellular trafficking, secretion, and vesicular transport |
| O | 96 | 2.87 | Posttranslational modification, protein turnover, chaperones |
| C | 129 | 3.85 | Energy production and conversion |
| G | 93 | 2.78 | Carbohydrate transport and metabolism |
| E | 137 | 4.09 | Amino acid transport and metabolism |
| F | 53 | 1.58 | Nucleotide transport and metabolism |
| H | 108 | 3.22 | Coenzyme transport and metabolism |
| I | 72 | 2.15 | Lipid transport and metabolism |
| P | 135 | 4.03 | Inorganic ion transport and metabolism |
| Q | 34 | 1.02 | Secondary metabolites biosynthesis, transport, and catabolism |
| R | 138 | 4.12 | General function prediction only |
| S | 51 | 1.52 | Function unknown |
| X | 88 | 2.63 | Mobilome: prophage, transposons |
| - | 1334 | 39.83 | Not in COGs |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhang, S.; Zhao, D.; Ni, Y.; Wang, W.; Yan, L. Complete Genome Sequence of Acidithiobacillus ferrooxidans YNTRS-40, a Strain of the Ferrous Iron- and Sulfur-Oxidizing Acidophile. Microorganisms 2020, 8, 2. https://doi.org/10.3390/microorganisms8010002
Zhang Y, Zhang S, Zhao D, Ni Y, Wang W, Yan L. Complete Genome Sequence of Acidithiobacillus ferrooxidans YNTRS-40, a Strain of the Ferrous Iron- and Sulfur-Oxidizing Acidophile. Microorganisms. 2020; 8(1):2. https://doi.org/10.3390/microorganisms8010002
Chicago/Turabian StyleZhang, Yu, Shuang Zhang, Dan Zhao, Yongqing Ni, Weidong Wang, and Lei Yan. 2020. "Complete Genome Sequence of Acidithiobacillus ferrooxidans YNTRS-40, a Strain of the Ferrous Iron- and Sulfur-Oxidizing Acidophile" Microorganisms 8, no. 1: 2. https://doi.org/10.3390/microorganisms8010002