sRNA OsiA Stabilizes Catalase mRNA during Oxidative Stress Response of Deincoccus radiodurans R1

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria Strain, Growth Conditions, and Oxidative Stress Treatment
2.2. Total RNA Extraction
2.3. Candidate sRNA Screen and the Prediction of sRNA Target Genes
2.4. Gene Ontology (GO) and KEGG Enrichment Analysis
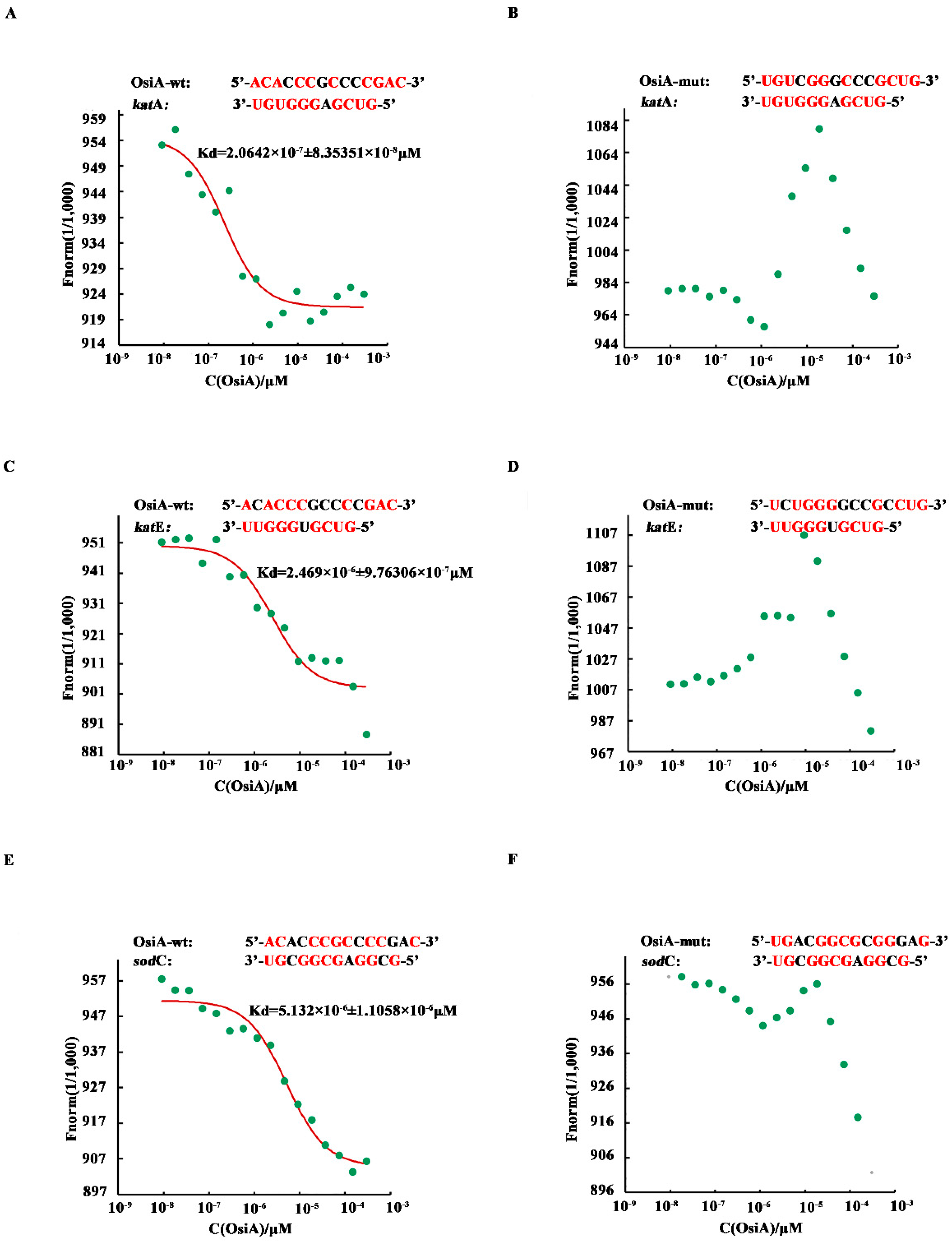
2.5. Microscale Thermophoresis Analysis
3. Results
3.1. Deep Sequencing sRNAs from D. radiodurans R1 under Oxidative Stress
3.2. 24 Oxidative Related sRNAs Were Identified by KEGG Analysis
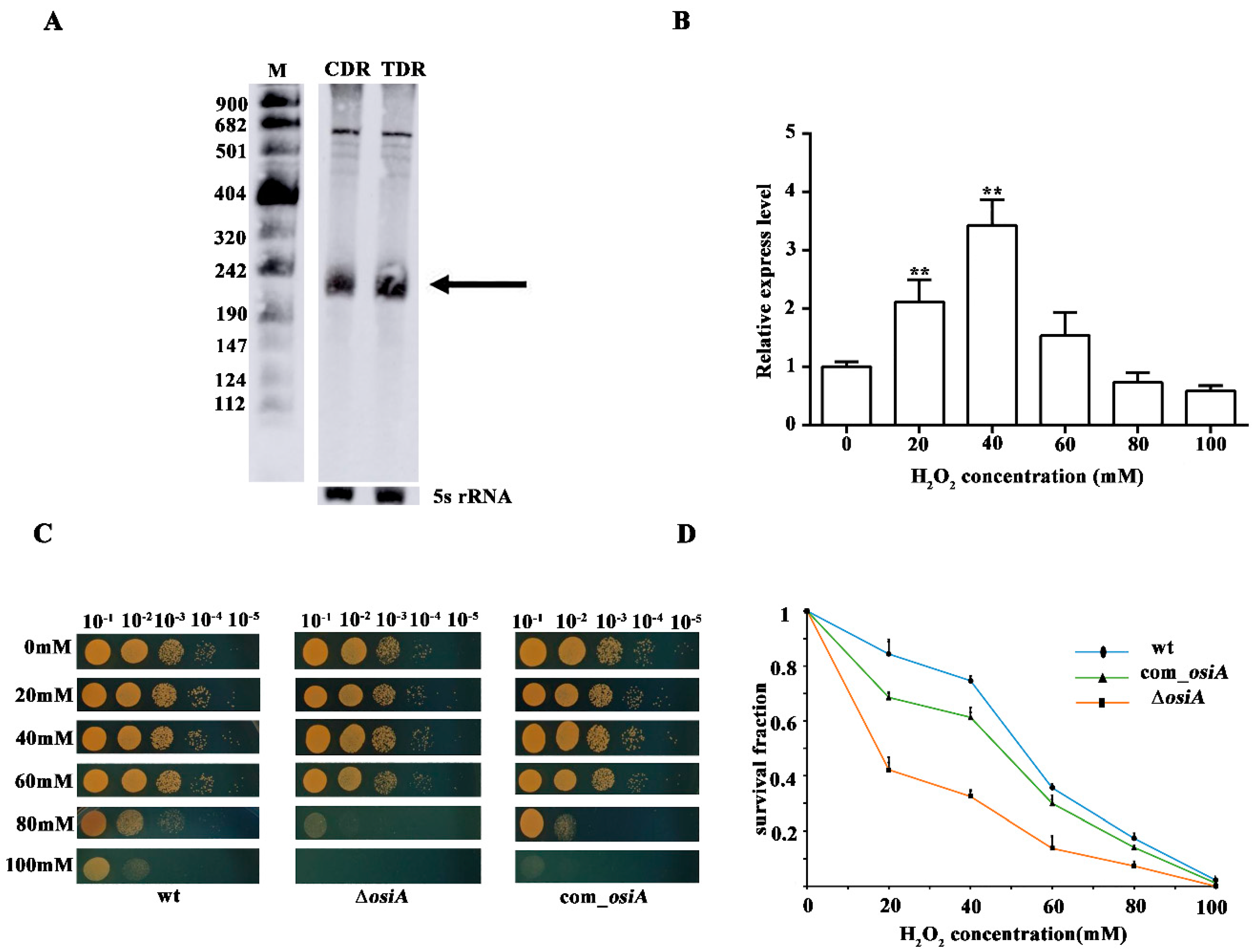
3.3. Characterization of OsiA, an Oxidative Response sRNA
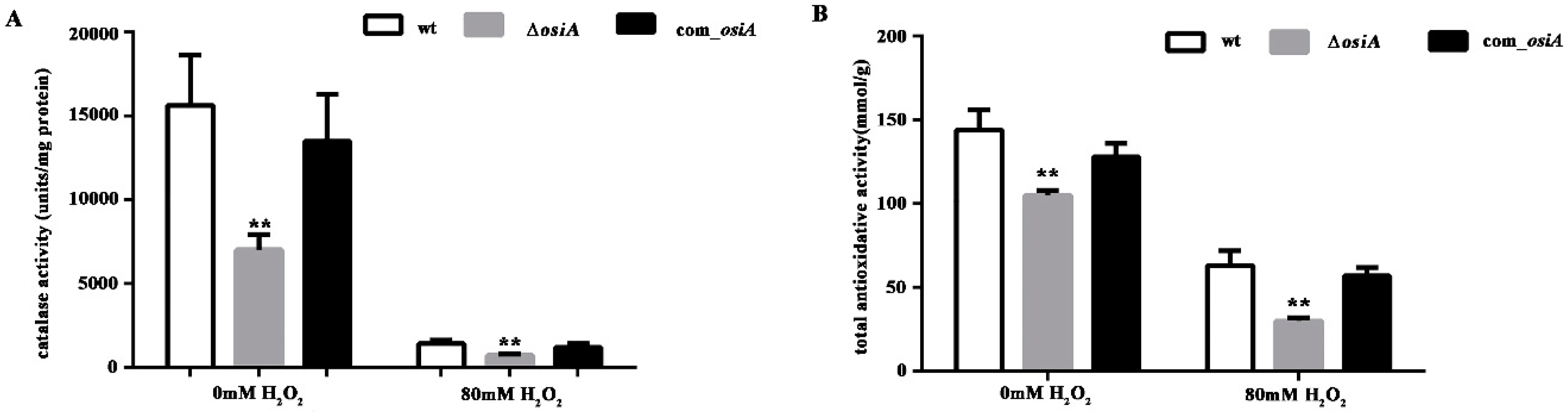
3.4. OsiA Enhances the Stability of Transcripts by Binding to the Target Genes through Multiple Bases
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cabiscol, E.; Tamarit, J.; Ros, J. Oxidative stress in bacteria and protein damage by reactive oxygen species. Int. Microbiol. 2000, 3, 3–8. [Google Scholar]
- Singh, O.V.; Gabani, P. Extremophiles: Radiation resistance microbial reserves and therapeutic implications. J. Appl. Microbiol. 2011, 110, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Q.; Ma, X.; Tian, B.; Li, T.; Yu, J.; Dai, S.; Weng, Y.; Hua, Y. Biosynthesis of gold nanoparticles by the extreme bacterium Deinococcus radiodurans and an evaluation of their antibacterial properties. Int. J. Nanomed. 2016, 11, 5931–5944. [Google Scholar] [CrossRef] [PubMed]
- White, O.; Eisen, J.A.; Heidelberg, J.F.; Hickey, E.K.; Peterson, J.D.; Dodson, R.J.; Haft, D.H.; Gwinn, M.L.; Nelson, W.C.; Richardson, D.L.; et al. Genome sequence of the radioresistant bacterium Deinococcus radiodurans R1. Science 1999, 286, 1571–1577. [Google Scholar] [CrossRef] [PubMed]
- Cox, M.M.; Battista, J.R. Deinococcus radiodurans - The consummate survivor. Nat. Rev. Microbiol. 2005, 3, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, L.; Li, T.; Lin, L.; Dai, S.; Tian, B.; Hua, Y. A PerR-like protein involved in response to oxidative stress in the extreme bacterium Deinococcus radiodurans. Biochem. Biophys. Res. Commun. 2014, 450, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Dai, S.; Tian, B.; Li, T.; Yu, J.; Liu, C.; Wang, L.; Xu, H.; Zhao, Y.; Hua, Y. DqsIR quorum sensing-mediated gene regulation of the extremophilic bacterium Deinococcus radiodurans in response to oxidative stress. Mol. Microbiol. 2016, 100, 527–541. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Sun, H.; Feng, Q.; Lu, H.; Zhao, Y.; Zhang, H.; Xu, X.; Jiao, J.; Wang, L.; Hua, Y. Extracellular dGMP Enhances Deinococcus radiodurans Tolerance to Oxidative Stress. PLoS ONE 2013, 8, e54420. [Google Scholar] [CrossRef]
- Ghosal, D.; Omelchenko, M.V.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Venkateswaran, A.; Zhai, M.; Kostandarithes, H.M.; Brim, H.; Makarova, K.S.; et al. How radiation kills cells: Survival of Deinococcus radiodurans and Shewanella oneidensis under oxidative stress. FEMS Microbiol. Rev. 2005, 29, 361–375. [Google Scholar]
- Slade, D.; Radman, M. Oxidative Stress Resistance in Deinococcus radiodurans. Microbiol. Mol. Biol. Rev. 2011, 75, 133–191. [Google Scholar] [CrossRef]
- Tian, B.; Sun, Z.; Shen, S.; Wang, H.; Jiao, J.; Wang, L.; Hu, Y.; Hua, Y. Effects of carotenoids from Deinococcus radiodurans on protein oxidation. Lett. Appl. Microbiol. 2009, 49, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Chen, H.; Xu, G.; Shah, A.M.U.H.; Hua, Y. DNA binding is essential for PprI function in response to radiation damage in Deinococcus radiodurans. DNA Repair (Amst.) 2012, 11, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Huang, L.; Hua, X.; Yin, L.; Hu, Y.; Wang, C.; Chen, W.; Yu, X.; Xu, Z.; Tian, B.; et al. Pleiotropic effects of RecQ in Deinococcus radiodurans. Genomics 2009, 94, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Devigne, A.; Ithurbide, S.; Bouthier de la Tour, C.; Passot, F.; Mathieu, M.; Sommer, S.; Servant, P. DdrO is an essential protein that regulates the radiation desiccation response and the apoptotic-like cell death in the radioresistant Deinococcus radiodurans bacterium. Mol. Microbiol. 2015, 96, 1069–1084. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, L.; Guérin, P.; Roche, D.; Cruveiller, S.; Pignol, D.; Vallenet, D.; Armengaud, J.; de Groot, A. Conservation and diversity of the IrrE/DdrO-controlled radiation response in radiation-resistant Deinococcus bacteria. Microbiologyopen 2017, 6, e477. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Arora, A.; Sairam, R.K.; Srivastava, G.C. Oxidative stress and antioxidative system in plants. Curr. Sci. 2002, 82, 1227–1238. [Google Scholar]
- Massé, E.; Vanderpool, C.K.; Gottesman, S. Effect of RyhB small RNA on global iron use in Escherichia coli. J. Bacteriol. 2005, 187, 6962–6971. [Google Scholar] [CrossRef]
- Zhan, Y.; Yan, Y.; Deng, Z.; Chen, M.; Lu, W.; Lu, C.; Shang, L.; Yang, Z.; Zhang, W.; Wang, W.; et al. The novel regulatory ncRNA, NfiS, optimizes nitrogen fixation via base pairing with the nitrogenase gene nifK mRNA in Pseudomonas stutzeri A1501. Proc. Natl. Acad. Sci. USA 2016, 113, E4348–E4356. [Google Scholar] [CrossRef]
- Zhang, H.; Zhan, Y.; Yan, Y.; Liu, Y.; Hu, G.; Wang, S.; Yang, H.; Qiu, X.; Liu, Y.; Li, J.; et al. The Pseudomonas stutzeri -specific regulatory ncRNA, NfiS, targets the katB mRNA encoding a catalase essential for optimal oxidative resistance and nitrogenase activity. J. Bacteriol. 2019, 201, e00334-19. [Google Scholar] [CrossRef]
- Romby, P.; Charpentier, E. An overview of RNAs with regulatory functions in gram-positive bacteria. Cell. Mol. Life Sci. 2010, 67, 217–237. [Google Scholar] [CrossRef] [PubMed]
- Citartan, M.; Raabe, C.A.; Hoe, C.H.; Rozhdestvensky, T.S.; Tang, T.H. Bacterial sRNAs: Regulation in Stress. In Stress and Environmental Regulation of Gene Expression and Adaptation in Bacteria; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2016. [Google Scholar]
- Pichon, C.; Felden, B. Small RNA gene identification and mRNA target predictions in bacteria. Bioinformatics 2008, 24, 2807–2813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, T.H.; Polacek, N.; Zywicki, M.; Huber, H.; Brugger, K.; Garrett, R.; Bachellerie, J.P.; Hüttenhofer, A. Identification of novel non-coding RNAs as potential antisense regulators in the archaeon Sulfolobus solfataricus. Mol. Microbiol. 2005, 55, 469–481. [Google Scholar] [CrossRef] [PubMed]
- Storz, G.; Vogel, J.; Wassarman, K.M. Regulation by Small RNAs in Bacteria: Expanding Frontiers. Mol. Cell 2011, 43, 880–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.H.; Liao, R.; Chou, B.; Contreras, L.M. Transcriptional analysis of Deinococcus radiodurans reveals novel small RNAs that are differentially expressed under ionizing radiation. Appl. Environ. Microbiol. 2015, 81, 1754–1764. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.; Kircher, M. Illumina sequencing library preparation for highly multiplexed target capture and sequencing. Cold Spring Harb. Protoc. 2010, 6. [Google Scholar] [CrossRef]
- Hua, X.; Hua, Y. Improved complete genome sequence of the extremely radioresistant bacterium Deinococcus radiodurans R1 obtained using PacBio singlemolecule sequencing. Genome Announc. 2016, 4, e00886-16. [Google Scholar] [CrossRef]
- Chen, Z.; Li, L.; Shan, Z.; Huang, H.; Chen, H.; Ding, X.; Guo, J.; Liu, L. Transcriptome sequencing analysis of novel sRNAs of Kineococcus radiotolerans in response to ionizing radiation. Microbiol. Res. 2016, 192, 122–129. [Google Scholar] [CrossRef]
- Busch, A.; Richter, A.S.; Backofen, R. IntaRNA: Efficient prediction of bacterial sRNA targets incorporating target site accessibility and seed regions. Bioinformatics 2008, 24, 2849–2856. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Boyle, E.I.; Weng, S.; Gollub, J.; Jin, H.; Botstein, D.; Cherry, J.M.; Sherlock, G. GO::TermFinder—Open source software for accessing Gene Ontology information and finding significantly enriched Gene Ontology terms associated with a list of genes. Bioinformatics 2004, 20, 3710–3715. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, F.M.; Wang, N.; Magee, G.B.; Nanduri, B.; Lawrence, M.L.; Camon, E.B.; Barrell, D.G.; Hill, D.P.; Dolan, M.E.; Williams, W.P.; et al. AgBase: A functional genomics resource for agriculture. BMC Genom. 2006, 7, 229. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Mao, X.; Cai, T.; Luo, J.; Wei, L. KOBAS server: A web-based platform for automated annotation and pathway identification. Nucleic Acids Res. 2006, 34, W720–W724. [Google Scholar] [CrossRef] [PubMed]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Beckert, B.; Kedrov, A.; Sohmen, D.; Kempf, G.; Wild, K.; Sinning, I.; Stahlberg, H.; Wilson, D.N.; Beckmann, R. Translational arrest by a prokaryotic signal recognition particle is mediated by RNA interactions. Nat. Struct. Mol. Biol. 2015, 22, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Jerabek-Willemsen, M.; André, T.; Wanner, R.; Roth, H.M.; Duhr, S.; Baaske, P.; Breitsprecher, D. MicroScale Thermophoresis: Interaction analysis and beyond. J. Mol. Struct. 2014, 1077, 101–113. [Google Scholar] [CrossRef] [Green Version]
- Buddeweg, A.; Sharma, K.; Urlaub, H.; Schmitz, R.A. sRNA41 affects ribosome binding sites within polycistronic mRNAs in Methanosarcina mazei Gö1. Mol. Microbiol. 2018, 107, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Kullapanich, C.; Dubbs, J.M.; Mongkolsuk, S. Inactivation of the Agrobacterium tumefaciens ActSR system affects resistance to multiple stresses with increased H2O2 sensitivity due to reduced expression of hemH. Microbiology 2019. [Google Scholar] [CrossRef]
- Navasa, N.; Ferrero, M.Á.; Rodríguez-Aparicio, L.B.; Monteagudo-Mera, A.; Gutiérrez, S.; Martínez-Blanco, H. The role of RcsA in the adaptation and survival of Escherichia coli K92. FEMS Microbiol. Lett. 2019, 366, 1–6. [Google Scholar] [CrossRef]
- Czégény, G.; Kőrösi, L.; Strid, Å.; Hideg, É. Multiple roles for Vitamin B6 in plant acclimation to UV-B. Sci. Rep. 2019, 9, 1259–1266. [Google Scholar] [CrossRef]
- Coutinho, L.G.; de Oliveira, A.H.S.; Witwer, M.; Leib, S.L.; Agnez-Lima, L.F. DNA repair protein APE1 is involved in host response during pneumococcal meningitis and its expression can be modulated by vitamin B6. J. Neuroinflamm. 2017, 14, 243. [Google Scholar] [CrossRef] [PubMed]
- Staerck, C.; Gastebois, A.; Vandeputte, P.; Calenda, A.; Larcher, G.; Gillmann, L.; Papon, N.; Bouchara, J.P.; Fleury, M.J.J. Microbial antioxidant defense enzymes. Microb. Pathog. 2017, 110, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Caro, A.A.; Cederbaum, A.I. Antioxidant properties of S-adenosyl-L-methionine in Fe2+-initiated oxidations. Free Radic. Biol. Med. 2004, 36, 1303–1316. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Ma, X.; Li, Z.; Jiao, Z.; Li, Y.; Ow, D.W. Maize OXIDATIVE STRESS2 homologs enhance cadmium tolerance in Arabidopsis through activation of a putative SAM-dependent methyltransferase gene. Plant Physiol. 2016, 171, 1675–1685. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function | sRNAs (+) | sRNAs (−) |
|---|---|---|
| Kinases | 15 | 17 |
| Transcription factor | 7 | 14 |
| Enzymes | 42 | 55 |
| Structural protein | 9 | 13 |
| sRNA Name | Pathway Mane | Target Genes Products |
|---|---|---|
| IGR_3053 | Peroxisome | KatA, SodC |
| Mismatch repair | RplV | |
| Two component system | methyl-accepting chemotaxis protein | |
| IGR_2205 | Peroxisome | class V aminotransferase |
| Mismatch repair | DNA ligase, DNA polymerase III, tau/gamma subunit, DR_2586 | |
| Glutathione metabolism | glucose-6-phosphate 1-dehydrogenase | |
| Carotenoid metabolism | phytoene dehydrogenase | |
| Two component system | NatA, DR_A0009, methyl-accepting chemotaxis-like protein | |
| Vitamin B6 metabolism | pyridoxamine 5-phosphate oxidase | |
| IGR_1662 | Peroxisome | KatE, |
| Mismatch repair | MutT, Ssb, MutS | |
| Carotenoid metabolism | phytoene synthase | |
| Two component system | 5-(carboxyamino)imidazole ribonucleotide mutase | |
| IGR-1449 | Mismatch repair | MutL, DNA helicase II |
| Glutathione metabolism | potassium-transporting ATPase subunit C | |
| Two component system | potassium-transporting ATPase subunit C, KdpD-related protein | |
| Vitamin B6 metabolism | RpsO, PdxT | |
| IGR_1916 | Peroxisome | long-chain fatty acid--CoA ligase |
| Mismatch repair | DR_1244 | |
| Glutathione metabolism | ValS | |
| IGR_2408 | Mismatch repair | 3-oxoacyl-acyl carrier protein reductase |
| Two component system | methyl-accepting chemotaxis protein, CheA-related protein | |
| Vitamin B6 metabolism | threonine synthase | |
| IGR_1612 | Mismatch repair | DNA polymerase III subunit epsilon, RuvA |
| IGR_2590 | Peroxisome | acyl-CoA synthetase |
| Mismatch repair | 3-isopropylmalate dehydratase large subunit | |
| Vitamin B6 metabolism | pyridoxamine kinase | |
| IGR_884 | Mismatch repair | XseA |
| Glutathione metabolism | diaminopimelate decarboxylase | |
| Two component system | potassium-transporting ATPase subunit B | |
| IGR_771 | Glutathione metabolism | nitrogen regulatory protein P-II, leucyl aminopeptidase |
| Carotenoid biosynthesis | lycopene cyclase | |
| IGR_2060 | Two component system | potassium-transporting ATPase subunit A |
| Vitamin B6 metabolism | RuvB, PdxS | |
| IGR_1174 | Peroxisome | isocitrate dehydrogenase |
| Glutathione metabolism | isocitrate dehydrogenase | |
| IGR_2150 | Mismatch repair | short chain dehydrogenase/reductase family oxidoreductase |
| Glutathione metabolism | cephalosporin acylase | |
| IGR_2479 | Mismatch repair | DNA polymerase III subunit alpha |
| Two-component system | succinate dehydrogenase, cytochrome subunit | |
| IGR_2389 | Peroxisome | Cu/Zn family superoxide dismutase |
| IGR_76 | Mismatch repair | single-stranded DNA-binding protein |
| IGR_585 | Mismatch repair | RpsS |
| IGR_2012 | Mismatch repair | single-stranded-DNA-specific exonuclease |
| IGR_2690 | Glutathione metabolism | Arginase, 6-phosphogluconate dehydrogenase-like protein |
| IGR_373 | Two-component system | NADH dehydrogenase II |
| IGR_826 | Two-component system | SdhB |
| IGR_1951 | Two-component system | DNA-binding response regulator |
| IGR_388 | Two-component system | PilH |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Xue, D.; Sun, W.; Han, J.; Li, J.; Gao, R.; Zhou, Z.; Zhang, W.; Chen, M.; Lin, M.; et al. sRNA OsiA Stabilizes Catalase mRNA during Oxidative Stress Response of Deincoccus radiodurans R1. Microorganisms 2019, 7, 422. https://doi.org/10.3390/microorganisms7100422
Chen Y, Xue D, Sun W, Han J, Li J, Gao R, Zhou Z, Zhang W, Chen M, Lin M, et al. sRNA OsiA Stabilizes Catalase mRNA during Oxidative Stress Response of Deincoccus radiodurans R1. Microorganisms. 2019; 7(10):422. https://doi.org/10.3390/microorganisms7100422
Chicago/Turabian StyleChen, Yun, Dong Xue, Wenjie Sun, Jiahui Han, Jiang Li, Ruyu Gao, Zhengfu Zhou, Wei Zhang, Ming Chen, Min Lin, and et al. 2019. "sRNA OsiA Stabilizes Catalase mRNA during Oxidative Stress Response of Deincoccus radiodurans R1" Microorganisms 7, no. 10: 422. https://doi.org/10.3390/microorganisms7100422