The Genetic Basis of Toxin Biosynthesis in Dinoflagellates
by
 , and
, and
Arjun Verma
1,* ,
,
Abanti Barua
1,2,
Rendy Ruvindy
1,
Henna Savela
3,
Penelope A. Ajani
1 and
Shauna A. Murray
1 1
Climate Change Cluster, University of Technology Sydney, Sydney 2007, Australia
2
Department of Microbiology, Noakhali Science and Technology University, Chittagong 3814, Bangladesh
3
Finnish Environment Institute, Marine Research Centre, 00790 Helsinki, Finland
*
Author to whom correspondence should be addressed.
Microorganisms 2019, 7(8), 222; https://doi.org/10.3390/microorganisms7080222
Submission received: 22 June 2019
/
Revised: 23 July 2019
/
Accepted: 27 July 2019
/
Published: 29 July 2019
(This article belongs to the Special Issue Dinoflagellate Biology in the Omics Era)
Abstract
:In marine ecosystems, dinoflagellates can become highly abundant and even dominant at times, despite their comparatively slow growth rates. One factor that may play a role in their ecological success is the production of complex secondary metabolite compounds that can have anti-predator, allelopathic, or other toxic effects on marine organisms, and also cause seafood poisoning in humans. Our knowledge about the genes involved in toxin biosynthesis in dinoflagellates is currently limited due to the complex genomic features of these organisms. Most recently, the sequencing of dinoflagellate transcriptomes has provided us with valuable insights into the biosynthesis of polyketide and alkaloid-based toxin molecules in dinoflagellate species. This review synthesizes the recent progress that has been made in understanding the evolution, biosynthetic pathways, and gene regulation in dinoflagellates with the aid of transcriptomic and other molecular genetic tools, and provides a pathway for future studies of dinoflagellates in this exciting omics era.
1. Introduction
Marine microbial eukaryotes are a diverse group of organisms comprising lineages that differ widely in their evolutionary histories, ecological niches, growth requirements, and nutritional strategies [1,2,3,4,5]. Among marine microbial eukaryotes (protists), dinoflagellates are of immense ecological and evolutionary significance [3,6,7,8]. Recent studies on protist species richness in the world’s oceans using genetic tools, such as metabarcoding, have shown that approximately half of 18S rDNA richness is made up of dinoflagellate sequences [9,10]. This widespread diversity is due to their ability to adapt to a wide variety of ecological niches, largely depending on their complex survival strategies (such as photoautotrophy, symbiosis, mixotrophy, and heterotrophy) [11,12]. They possess significant variability in morphology, pigment composition, and photosynthetic activity along with a broad spectrum of biological activities that serves various ecological niches. Their occupation of such diverse environments can be witnessed in their fossil records that date back several hundred million years [3,4,11].
Dinoflagellates are relatively inefficient at nutrient uptake, and exhibit slower growth rates compared to other protists such as chlorophytes, haptophytes, and diatoms [13,14]. Despite being poor competitors within their ecosystems, they can at times proliferate in large abundances, causing harmful algal blooms (HABs) [15,16]. Only about 2% of algal species have been reported as producing harmful blooms, of which 75% are dinoflagellates [14]. HABs are naturally occurring phenomena, however there is evidence that the frequency, geographic range, and intensity of these occurrences have increased over the past 30 years [17,18,19,20]. While the causes and consequences of HABs have been studied extensively, the production of toxic compounds by the culprit species and their interactions with co-occurring phytoplankton and potential predators remain to be fully understood [21,22]. These compounds display a wide variety of biological activity and may play distinct roles for producing organisms. One example of this is that the karlotoxins are produced by Karlodinium veneficum that may inhibit the growth of co-occurring plankton and immobilize potential prey species in close proximity [23,24]. Another example are the toxins produced by Alexandrium spp. that may play a role in grazing deterrence, with species-specific impacts found on copepods such as reduced grazing, reduced fecundity and delayed development [25,26,27,28,29]. Brevetoxins produced by Karenia brevis is also known to promote the survival of dinoflagellates by affecting grazer behavior [30,31,32]. Despite these studies, whether they are other as-yet-unidentified cellular functions of these compounds is currently unknown [32] (Figure 1).
Toxins produced during HABs can have widespread impacts on fisheries and aquaculture industries worldwide [17,33,34]. The most common effects of HABs are poisoning events, resulting in the deaths of marine organisms, and the accumulation of toxins in the marine food web, which may lead to eventual human poisoning via the consumption of contaminated seafood [35,36]. It is estimated that up to 60,000 human intoxications occur per year worldwide, with an overall mortality of approx. 1.5% [37,38]. The potential harm caused to commercially produced seafood by these marine toxins, and the associated human health risks are a concern for seafood safety regulatory bodies worldwide [39]. The greatest number of HAB related human health impacts have occurred due to toxic compounds that are classified as; saxitoxin (STX) and its analogues causing paralytic shellfish poisoning (PSP); brevetoxins (BTXs) and analogues causing neurotoxic shellfish poisoning (NSP); ciguatoxins (CTXs) and related compounds resulting in ciguatera fish poisoning (CFP); and okadaic acid (OA) and the dinophysis toxins (DTXs) causing diarrheic shellfish poisoning (DSP) [35,38]. Besides these well-known toxic compounds and their related illnesses, several new poisoning syndromes have recently appeared due to dinoflagellate toxins, such as azaspiracids (AZAs), yessotoxins (YTXs), and palytoxins (PLTXs) [38,40]. Approximately 20,000 marine natural products, with unique size, complexity, and biosynthetic pathways, have been discovered over the past 50 years, of which dinoflagellate toxins represent only a minor fraction, with many more that remain to be described [41]. Over the past 15 years, the first studies have begun to examine the genetic basis of bioactive compounds produced by algae, particularly the toxins synthesized by marine dinoflagellates. These studies have been challenging, largely due to the large size and the distinctive organization of dinoflagellate genomes compared to that of other eukaryotes [5,6].
2. Dinoflagellate Genomics
Dinoflagellate genomes are generally larger than the genomes of most other protists, ranging from 3–250 picograms of DNA per haploid genome, equating to around 1.2–112 × 109 base pairs [42]. This genetic material is encoded in 24–220 nuclear chromosomes that are permanently condensed in a liquid crystalline state throughout the cell cycle, attached to the nuclear envelope and lacking nucleosomal heterochromatin [43,44,45]. Instead of nucleosomes, dinoflagellate DNA is associated with histone like proteins (HLPs) that are similar to bacterial DNA binding proteins (HU proteins) [46,47,48]. Dinoflagellate genomes have a generally low protein content (10:1 DNA: protein in the chromatin) and their histones exhibit frequent losses of key residues that are highly conserved among all other eukaryotes [49,50,51]. These HLPs are expressed at low levels and play a limited role in chromatin packaging [50,52]. This role of chromosomal organization is seemingly filled by viral-derived proteins called dinoflagellate-viral-nucleoproteins (DVNPs) that were potentially transferred from viruses to dinoflagellate ancestors, where they have eventually replaced histones as the primary chromatin packaging proteins [53,54,55].
Dinoflagellate nuclear DNA has a relatively high guanine and cytosine (G+C) content varying from 45%–70% [52,56,57]. It also hosts several modified nucleotide building blocks such as 5-methyl cytosine, N6-methyl adenine, and 5-hydroxymethyluracil (5-meU) that make it extensively methylated [58,59]. It has been suggested that 5-meU, that substitutes around 70% of the thymine (T), is associated with a restriction-modification system for discriminating between dinoflagellate and foreign DNA sequences thereby promoting stability in the open reading frames of the dinoflagellate genes [49,58,59].
Dinoflagellate nuclear genomes have acquired plastid targeted genes via successive horizontal gene transfer from the peridinin plastid, the tertiary replacement plastid and its host nucleus, cyanobacteria, red algae, haptophyte, and even bacteria, due to which the genomes are highly chimeric [60,61,62,63]. Despite this, their gene content is lower than expected, varying from 37–87 × 103 protein coding genes, which accounts for only 0.05%–1.8% of the total genomic DNA [63]. However, this still remains larger than most other eukaryotes [63,64]. Two partial genomes from species belonging to the Symbiodiniaceae family, i.e., Fugacium kawaguti and Breviolum minutum, have been sequenced [56,65,66]. Recently the Amoebophrya ceratii (Syndiniales) genome was sequenced, which was substantially smaller than the Symbiodiniaceae genomes reported so far [67]. These studies show that dinoflagellates have uni-directionally aligned genes, forming cluster-like arrangements and have gene models that can be grouped into families [56,65]. Many of these genes are organized in multiple copies as tandem repeats, some of which may be present in up to ~105 copies [68,69]. Despite the high gene copy numbers, the dinoflagellate genomes are mostly repetitive non-coding DNA. However, some dinoflagellate genes may have a low intron density and occasionally even lack introns, and these may be genes that are highly expressed [68].
Several ‘intron-less’ genes have resulted from the incorporation of cDNA back into the dinoflagellate genome through a process of trans-regulatory elements, or retroposition [70,71]. In the dinoflagellate genomes sequenced, >20% of genes appeared to be the result of retroposition [56,65,67,72]. Such trans-regulatory processes are mRNA processing reactions by which exons from two separately transcribed pre-mRNAs are joined [73,74]. This process also adds a 22 bp sequence, known as the spliced leader (SL), at the 5’ end of transcribed mRNA, possibly making the mRNA pool for translation [73,74]. SL trans-splicing acts to convert polycistronic pre-mRNA to monocistronic mRNA and also acts as a gene expression regulator [73,74,75]. However, SL trans-splicing in dinoflagellates is different compared to that in other organisms since the conserved binding motifs occur in the exons instead of the introns [73,75]. Dinoflagellate genomes appear to lack the cis-regulatory elements such as the TATA box, but have appeared to display TTTT(G) motifs that might have replaced the TATA box [65]. As a result of such factors, most genes appear to be post-transcriptionally regulated, with only 5%–30% being regulated at the transcriptional level [6].
The organelle genomes in dinoflagellates (both plastid and mitochondrial) are also remarkably different compared to other eukaryotes [63,76,77,78]. Most eukaryotic plastids contain circular genomes encoding 100–200 genes. However, peridinin containing dinoflagellates have highly reduced plastid genomes containing only 3–4 proteins that are encoded into plasmid-like ‘minicircles’ [78,79]. Similar to their nuclear genome, dinoflagellate organelle genomes have several gene copies that are frequently disrupted by fragments of other genes, and many of these genes are transferred to the nucleus [80]. Therefore, the peridinin-containing dinoflagellates encode the smallest number of plastid genes for any photosynthetic eukaryote [78]. The mitochondrial genomes in dinoflagellates are also highly duplicated and recombined, similar to the nuclear genomes, and are comprised of abundant non-coding and repetitive sequences that are rich in AT content, yet these genomes are highly reduced compared to other closely related protists, due to gene transfers to the nucleus [63,67,77].
The complex genetic machinery of dinoflagellates, as briefly described above, makes it tedious to verify certain genetic pathways in dinoflagellates [6,63,81]. Furthermore, dinoflagellates have associated bacteria that cannot be eliminated from cultures, which makes it difficult to fully exclude the impact of these associated assemblages on biosynthetic pathway studies [6]. With the aid of sequencing cDNA libraries to generate expressed sequence tags (ESTs), it became possible to study gene expression and regulation in dinoflagellates for which little-to-no genomic information is available [63]. ESTs serve as markers for genes expressed under specific conditions and can be used as probes in the recovery of full-length cDNA or genomic sequences, recognition of exon and intron boundaries, delineation of protein families, and development of probes for genome wide expression profiling [52,63,82,83]. Furthermore, transcriptomic surveys by way of next-generation sequencing (NGS) RNA-Seq, in particular, have illuminated our understanding of the unique biology, metabolism, and ecology of dinoflagellates [82,84,85]. In this review, we summarize our current understanding of the genetic basis of toxin biosynthesis in dinoflagellates and discuss the future directions for this field.
3. Dinoflagellate Toxins
Dinoflagellate toxins are structurally and functionally diverse and possess unique biological activities, including ion channel modulation, phosphatase inhibition, hemolysis, mycotoxicity, and cytotoxicity [86,87]. In organisms including humans, the voltage-gated ion channels, such as the sodium, calcium, and potassium channels are electric signal generators that control muscle contractions, hormone secretion, sensing of the environment, information processing in the brain, and the reflex output to the peripheral tissues/muscles [38]. These channel pores can get blocked by toxin molecules, preventing ion conductance and altering voltage dependent gating [38]. Traditionally, the biosynthetic pathways for these complex compounds have been elucidated by isotope labelled studies on the cultured microorganisms [40,88]. While these studies have not been able to comprehensively elucidate the entire biosynthetic pathways involved, they have provided us great insights into the structural complexity of these compounds [87,88].
3.1. Dinoflagellate Toxins: Polyketides
Polyketides are a highly diverse group of natural products with large and complex carbon structures that are assembled from simple acyl building blocks [89,90]. Most toxins currently linked to NSP, CFP, and DSP, as well as AZAs, PLTXs, YTXs, and cyclic imines, have a polyketide backbone and are broadly classified into three main categories according to their chemical structures; (a) polycyclic polyketides (e.g. BTXs, CTXs, YTXs, PLTXs, and maitotoxins), (b) macrolides (e.g., amphidinolides, pectenotoxins, and spirolides), and (c) linear polyketides (e.g., DTXs and OA) [87,88,91]. Much of our current knowledge on polyketide biosynthesis is obtained from bacterial and fungal products and their biosynthetic pathways, which have enormous commercial value and remain among the most successful candidates among natural drug discovery [91,92]. The enzymes responsible for polyketide biosynthesis are large multi-domain enzyme complexes known as polyketide synthases (PKSs) that resemble fatty acid synthases (FASs) in both structure and function [91,93,94,95,96,97,98]. PKSs have unusual structures and catalytic reactions that are essential for understanding enzymatic catalysis and protein–protein interactions among fundamental biological processes in dinoflagellates. Such knowledge will improve our knowledge of their ecological and evolutionary significance, and will also aid in harnessing their biological potential for medical and other biotechnological advancements [92,99].
3.1.1. Polyketide Biosynthesis
Polyketide biosynthesis is a product of sequential condensation reactions of small carboxylic acid subunits into a growing acyl chain, similar to fatty acid biosynthesis, performed by β-ketosynthase (KS) domains [91,94,97,100]. The growing carbon chain resides on an acyl carrier protein (ACP) that presents it to the catalytic sites of the KS domains, while an acyl transferase (AT) presents the extender units to be added to the growing chain [91,100]. These carbon chains can undergo optional reduction of beta-ketone to an alcohol, dehydration of the alcohol, and saturation of the resultant double bond through the activities of ketoreductase (KR), dehydratase (DH), and enoyl-reductase (ER) domains, respectively [89,100]. In addition, non-ribosomal peptide synthases (NRPSs) are also linked to the biosynthesis of polyketide molecules, especially the ones that contain an amide or amine group in their structure [101,102]. These large multi-modular domains catalyze the incorporation of amino acids into the polyketide backbone in a manner analogous to PKSs [102]. A typical NRPS module comprises of an initiation module, i.e., an adenylation domain (A) that specifically activates an amino acid, a peptidyl carrier protein (PCP), and a condensation domain (C) that is responsible for chain elongation by creating a peptide bond between two PCP-bound amino acids [101,102]. Finally, a thioesterase domain (TE) releases the full-length molecule from the PKS complex [91,100,103] (Figure 2).
PKSs are generally classified into three groups according to structural and functional elements (domain organization) [94,98]. Type I PKSs are large proteins comprising a set of catalytic domains on a single protein that are used in a repetitive fashion for chain elongation, similar to fatty acid synthesis in animals and fungi [94,98]. Type II PKSs comprise different multi-protein complexes, where each catalytic domain is on a separate peptide domain which function as mono-functional proteins in an iterative fashion analogous to type II FASs in bacteria and plants [94,98]. Type III PKSs are self-contained homodimeric enzymes, where each monomer performs a specific function in an iterative manner without the use of ACP proteins and acts directly on the acyl unit precursor molecules [91,98,100]. These PKSs are typically associated with chalcone and stillbene synthases in higher plants, but have also been identified in several bacteria and are similar to Type II PKSs, but smaller in size [91,98,100]. Similarly, FASs are classified as type I multi-domain proteins synthesizing lipids in the cytosol [104], or as Type II FAS complexes that catalyse lipid biosynthesis in the chloroplast stroma of plants [105].
PKSs and FASs are well documented amongst bacterial and fungal secondary metabolites, and studies over the past 15 years have also begun to report them in toxic dinoflagellates [81]. The common evolutionary history of PKSs and FASs, and their possible bacterial origins have been the subject of immense discussion [81,94,95]. Multiple studies over the last decade examining their evolutionary history and functional diversity have been put forward to fully comprehend the complex toxin biosynthesis pathways in dinoflagellates.
3.1.1.1. The Eukaryotic Origin of Polyketide Toxin Biosynthesis
The first PKSs in dinoflagellates were identified by Snyder et al. [101] that reported partial type I, but not type II PKS genes from non-axenic cultures of six different dinoflagellate species using reverse transcription (RT) PCR. These species included OA producing Prorocentrum lima and P. hoffmanianum, BTX producing K. brevis and amphidinolide producing Amphidinium operculatum [106]. Partial type I PKSs were also identified from Symbiodinium sp. and Gymnodium catenatum, which were not known as polyketide producers at the time, thereby highlighting the potential to identify novel polyketides from these species in the future. This study did not demonstrate a clear phylogenetic origin as the KS domains were interspersed with bacterial and fungal type I KS sequences, and sequences from K. brevis branched into the clade with PKS encoding genes from the apicomplexan parasite Cryptosporidium parvum [106,107]. However, as 16S rDNA genes were amplified from the dinoflagellate cultures, it could not be ruled out that these were bacterial genes [106]. Snyder et al. [108] followed this study with the discovery of two putative PKS genes from K. brevis that were localized within the cell using both fluorescence in situ hybridization (FISH) and PCR screening, but also found these genes in the associated bacteria, as well as in Amphidinium strains that are non-brevetoxin producers [63]. This study presented the first evidence of resident PKS genes in any dinoflagellate, but could not comprehensively conclude if they were potentially derived from the associated bacteria [108].
The discovery of the conserved spliced leader sequence at the 5’ end of mRNA and the poly-A tail at the 3’ end from over 100 contigs from several different dinoflagellate species aided in identifying full length PKS transcripts from K. brevis [73,74,109]. This was pivotal in investigating toxin biosynthesis genes from dinoflagellates as it provided the most compelling evidence of dinoflagellate origin of these domains, as the process of trans-splicing on polycistronic mRNA does not take place in bacteria [73,74]. Since then numerous studies have reported PKSs from dinoflagellates with a 5’ spliced leader sequence and poly A tails, which phylogenetically group within the protist clade, thereby confirming their eukaryotic origin [81].
3.1.1.2. FAS vs PKS in Toxin Biosynthesis
The structural and functional similarities between FASs and PKSs have caused considerable debate over the biosynthesis of these molecules in dinoflagellates [77,89]. Early research on fatty acid biosynthesis genes in dinoflagellates and other closely related organisms identified cytosolic type I FASs from the dinoflagellate Crypthecodinium cohnii [110], in contrast to type II FASs that were reported from the apicomplexans Toxoplasma gondii and Plasmodium falciparum [111]. These findings gave rise to the hypothesis suggesting multi-functional Type I FAS/PKS genes could have evolved from individual Type II FAS genes through one or more gene fusion events. Van Dolah et al. [112] used 3H acetate labelling of fatty acids in K. brevis and reported cytosolic incorporation of these molecules, with little incorporation in the chloroplasts. Such feature is typical of type I FAS systems [112]. Along with the absence of Type II FAS transcripts from the transcriptomic assemblies of K. brevis, these findings could only be supported by the presence of a Type I FAS system in K. brevis. However, no sequences with Type I FAS conserved domains could be reported from the transcriptome, which raised the question of whether dinoflagellate fatty acid synthesis was carried out by PKSs [112]. Pawlowiez et al. [113] constructed a cDNA library from the CTX -producing G. polynesiensis and identified 33 PKS related sequences, but none of the KS domains reported in this study were mapped to the fatty acid biosynthesis pathway, which provided further evidence for fatty acid biosynthesis possibly being carried out by PKSs [113].
Kohli et al. [81] mined the transcriptomic libraries of 24 genera and 46 strains of dinoflagellates that were sequenced under the Marine Microbial Eukaryote Transcriptome Sequencing Project (MMETSP) and screened them for the seven key enzymes involved in fatty acid biosynthesis, i.e. 3-ketoacyl ACP synthase I, II and III, ACP S-malonyl-transacylase, trans-3-ketoacyl ACP reductase, 3-hydroxyacyl-ACP dehydratase and enoyl-ACP reductase. This study confirmed the eukaryotic origin of FASs as well as their type II like structure, by identifying 5’ spliced leaders and poly A tails on them as well as detecting transit peptides targeted towards the chloroplast [81]. The authors postulated that these type II FAS genes have transferred from the cytosolic plastids over to the nucleus as they are under a strong selection pressure owing to them coding for essential life processes [81]. This study clearly separated FASs from PKSs and thus provided a good framework to investigate the different functionality of these genes and the biosynthetic pathways in dinoflagellates [81].
3.1.1.3. KS Domains: Phylogenetic and Structural Diversity
Most studies investigating PKS toxin biosynthesis have targeted the KS domains as they are the most conserved among the PKS genes [114]. These domains have the greatest potential for revealing divergent homologs and thus provide the most information on the evolutionary history of PKS genes in dinoflagellates, and protists in general [114,115]. Early phylogenetic analysis performed on protists by John et al. [115] using the genomic data from Chromalveolate, Excavate and Plantae species, along with other known protist and bacterial PKS sequences, showed sequences with high similarity to type I PKS genes were present in only a few lineages. Besides the patchy distribution of type I PKSs, a new modular type I PKS gene cluster from protists was identified that differed from previously known type I PKS clusters [115]. Following studies on both toxic and non-toxic strains of K. brevis grouped the dinoflagellate derived KS contigs within the aforementioned type I PKS clade containing sequences from other protists [109,116]. While the K. brevis sequences clustered with type I PKSs, their structure and size were more similar to type II PKSs, with the presence of individual catalytic domains on separate transcripts [109,116]. Since that time, single domain PKSs have been reported in numerous toxic and non-toxic dinoflagellate strains/species [113,117,118,119,120,121,122,123]. Eichholz et al. [117] reported two dinoflagellate specific clades within the ‘protistan’ clade, one that was comprised of sequences from all three dinoflagellate with typical KS amino acid motifs and the other clade containing only sequences from K. brevis with alterations in their active sites [117].
With more comprehensive transcriptomic sequencing and genome wide surveys of several dinoflagellate species, numerous sub-clades containing both mono- and multi-functional domains have now been reported (Figure 3). These domains closely match with sequences from other dinoflagellates as well as species of apicomplexans, chlorophytes, and haptophytes among the type I ‘protistan’ clade. Such phylogenetic analysis suggest that these genes might be involved in the production of polyketide compounds that are produced by all species since no clear pattern based on chemical structure of the toxic compounds could be established [81,119,120,122,124,125]. Dinoflagellates have a much larger number of unique KS domains compared to other protists, and possibly all other eukaryotes [81] (Figure 4). Kohli et al. [81] extracted a total of 2577 KS domains from 24 genera comprising 46 strains of dinoflagellates, which were broadly clustered into three distinct type I single domain KS sub-clades within the dinoflagellate type I PKSs (Figure 3). Each of these clades included sequences from numerous species that are clearly not related to dinoflagellate species-based phylogeny suggestive of frequent, and possibly multiple, gene duplication events, horizontal gene transfer (HGT), domain shuffling and losses, and recombination have occurred in their genomes [81,125]. The conserved active site amongst KS domains (Cys-His-His), which is essential for their functionality, was observed in almost two-thirds of the domains, thereby suggesting novel functionality of these domains among dinoflagellate species [81].
Eichholz et al. [117] identified a previously uncharacterized conserved motif (ExExGYLG) at the N-terminal of the KS domains and speculated its function to be related to the mono-functional nature of KS domains and may play a role in structural rearrangements, substrate docking, or protein-protein interactions [117]. The conserved motif and its variants G(D/H/Y)YLG were observed at the 5’ end along with those containing another variant, G(A)LLG. These N terminal conserved sites have since been observed in other EST libraries [113,121,123]. The alteration of the conserved N terminal motif in the KS transcripts could explain the alternations/diversification in polyketide synthesis or the possibility that these copies might have diverged to play a role in other pathways via multiple gene duplication, loss, and selection events [113,117]. In contrast, no conserved domains were observed in the C-terminal region, thereby speculating that this region acts as a linker structure where the single domains could be separated to allow for the structural function of the complex enzymes [121].
3.1.1.4. Other PKS Genes
The minimal domains required for the PKS condensations reactions are the ACP, KS, and AT domains [87,88,91,100,106]. However, the work of auxiliary PKS domains yields a great diversity of polyketide compounds from dinoflagellates [88,106]. Kubota et al. [126] used degenerate PCR primer sets designed especially for type I KS domains and identified KS domains from an amphidinolide producing Amphidinium strain. In the same study, using shotgun sequencing, gene products showing similarity to AT, DH, KR, ACP, and TE were also identified. This study also highlighted the presence of introns among PKSs that caused difficulties in cloning the entire genes from genomic DNA library [126]. Bachvaroff and Place [68] also identified KS and KR domains, this time from the EST library of A. carterae, and reported up to 18 introns from the KR sequences that were surveyed. This suggests that PKS domains are intron-rich which was further confirmed with the genomic surveys of Symbiodiniaceae species [68,124,125]. All polyketide biosynthetic enzymes needed to synthesize the complex polyketide backbones, i.e. KS, KR, AT, ACP, ACP synthase (ACPS), TE, and ER, have been identified in several toxic dinoflagellates [119,120,121,123]. However, despite several transcriptomic and genomic surveys, only a handful of these domains have been investigated further for the better understanding of their evolutionary function and purpose in toxin biosynthesis.
Multi Domain PKSs
Deep sequencing of dinoflagellate transcriptomic libraries have yielded contigs encoding multiple PKS domains per transcript [113,122]. Initially, the dinoflagellate-specific spliced leader was not identified on these transcripts, and their clustering with bacterial KS domains suggested that these contigs were most likely part of the bacterial contamination within the dinoflagellate culture, since the cultures were not axenic [120,122]. Bachvaroff et al. [127] identified a hybrid PKS-NRPS transcript from A. carterae that encoded the dinoflagellate-specific spliced leader sequence and poly A tail that clustered with bacterial type I PKSs (Figure 3). Such domain architecture is reminiscent of the burA gene in bacteria Burkholderia that are PKS-NRPS hybrids and are involved in the synthesis of polyketide precursors in burkholderic acid biosynthesis [128]. These PKS-NRPS hybrids are potentially widespread among dinoflagellates as they have been identified from species that produce amine/amide-containing compounds, such as K. brevis and Ostreopsis spp., and also from species that do not produce any known products that would involve the NRPS pathway, e.g., Gambierdiscus spp. [122,123,129]. Such reports suggest that these domains were acquired prior to the diversification of dinoflagellates through horizontal gene transfer from cyanobacterial genes [118,122,123,124,130]. Dinoflagellate genomes are also subject to frequent recombination events and gene duplications, which play an important role in the evolution of modularity in PKSs. There is increasing evidence of multifunctional PKS domains in several dinoflagellates, indicating that multi-functionality coevolves with mono-functional domains [125]. However, it is still unclear whether the single domains PKSs have evolved from the decomposition of Type I multifunctional PKS clusters, or fusion of mono-functional PKS domains has led to multi-functionality [125].
Recent genomic surveys of PKS-NRPS domains in Symbiodiniaceae species demonstrated a phylogenetic trend specific to amino acid substrate utilization (amongst A- domains) and functional categories (in C-domains), as opposed to a species phylogeny [125]. This demonstrates that the chemical diversity in amine/amide-containing polyketide molecules is driven by the incorporated NRPS domain [125]. In such hybrids, a sequence of amino acids within the A-domain catalytic pocket governs the recognition and activation of amino acid substrates [125]. Therefore, any point mutations within the catalytic domain can drastically change the specificity of the A-domain. Single A- domains tend to incorporate polar and non-polar amino acids and interact with other NRPS components by transferring the substrate to the C- domain, which can perform epimerization or cyclization of the amino containing polyketide molecule [125].
Acyltransferase (AT) Domains
The acyltransferase (AT) domains recognize and attach carboxylic acid building blocks onto an ACP domain, which are then incorporated into the polyketide backbone by the catalytic reactions of the KS domains [89,96,100]. AT domains are broadly classified based upon specificity towards the building units [89,96,100,131]. Cis-AT PKSs are specific for methyl malonyl-CoA, hydroxyl malonyl-ACP, and methoxy malonyl-ACP, whereas trans-AT PKSs are more specific for malonyl-CoA only [131,132]. AT domains have been reported from several dinoflagellate species but have not been investigated in detail [120,121,123,125,126]. Recent genomic surveys of Symbiodiniaceae species by Beedessee et al. [125] reported a large number of trans-AT domains from these species and demonstrated a clear demarcation between cis- and trans- AT domains [125]. Trans ATs were first identified in bacteria and have since been reported from several microalgal species [131,133]. They are known to have unique catalytic mechanisms, highly unusual architectures with modules carrying either novel catalytic domains, or domain orders [132,134]. These domains have also been known to recombine via gene duplication, domain shuffling and retroposition, to form novel gene clusters in a mosaic like fashion, thereby promoting diversification of PKS biosynthesis. [125,134]. On the other hand, it is hypothesized that cis- AT domains may have been acquired via horizontal gene transfer of the entire PKS module, but remains to be investigated [125].
Ketoreductase (KR) Domains
Ketoreductase (KR) domains have been reported from several dinoflagellates to date, however the number of unique single KR domains identified from dinoflagellates are much smaller compared to the number of KS sequences in those species suggesting that the selection of KR domains is more conserved compared to the KS domains [121,122,123,129]. Recent phylogenetic investigations on KR domains placed the dinoflagellate derived KR domains into one large single domain protist clade, similar to KS domains, along with five distinct multi-KR domains sub-clades [123,129]. Sub-clade 1 comprised of KR domains from multi PKS modules with ‘KS-KR-ACP’ domain architecture, whereas sub-clade 2 contains KR domains from trans-AT PKS modules with ‘KS-DH-KR’ domains. Sub-clade 3 KR domains possess an ER domain inserted between the N-terminal and C-terminal, which has previously been described from FAS-like domains. Subclades 4 included all PKS-NRPS hybrids similar to burA-like sequences from dinoflagellates and Burkholderia spp. [123,129]. Van Dolah et al. [123] reported sub-clade 5 to include diverse multi PKS contigs with ‘KS-DH-KR’ architecture, along with PKS-NRPS hybrids and a multi-module PKS with highly amplified ACP domains and potential MT domains at their C-terminal ends [123]. Such phylogenetic analysis reveals that KR domains cluster according to their module architecture rather than grouping based on species or the toxic compound produced.
Functionally, KR domains have been classified based on the stereochemistry of the products they catalyze; i.e., A-type KRs produce an S-hydroxyl group and B-type KRs that produce the R-hydroxyl group [135]. An A-type KR would contain a conserved tryptophan residue (W motif) and generate an L-3-hydroxyacyl intermediate, whereas B-type KRs contain a conserved Leu-Asp-Asp (LDD motif) [136]. Among toxin producing dinoflagellates, several stereoisomeric analogues have been reported which do not share the same biological activity compared to other analogues. Such structure-activity relationships in toxin analogues could be driven by different KR domains that might be giving the molecules their varying toxigenic properties [129].
Methyl Transferase (MT) Domains
Both cyclic and non-cyclic polyketides constitute repetitive methyl groups that are derived from S-adenosyl methionine (SAM) [88,137,138]. MT domains catalyze the transfer of methyl group from SAM to either the carbon, nitrogen or oxygen atoms at various positions on the PKS-NRPS backbone and have been classified as C-MT, N-MT, and O-MT (carbon, nitrogen, and oxygen, respectively) depending upon their site of methylation [139,140]. MT domains have not been well documented in dinoflagellates. Only recent studies have unraveled multi-domain PKSs comprising KR-MT domains from several toxic and non-toxic dinoflagellate species [120,123,129]. These domains are known to act as linker regions that adopt a short chain reductase fold in the structure of large polyketides and might play a vital role in introducing folds in the structure of large toxin molecules such as PLTX and MTX and their analogues [129,141,142,143,144].
3.1.1.5. Other Genes
Through systematically assessing the functional capacity of 47 transcriptomic libraries from eight different dinoflagellate orders, Stephens et al. [145] identified a significant overrepresentation of membrane transporter gene families in toxin producing dinoflagellates, particularly the Voltage-gated Ion Channel (VIC) superfamily and the monovalent Cation: Proton Antiporter-1 (CPA1) family that are critical in the maintenance of ion concentrations and gradients across cell membranes. Even though these genes are not directly involved in toxin biosynthesis, they facilitate the secretion of toxins and make the producing cells resistant to their own toxins [145]. Most dinoflagellate toxins are known to target the sodium/calcium voltage-gated ion channels [38]. In eukaryotes, these channels are highly glycosylated with sialic acid, which is known to modulate the excitability of voltage-gated ion channels [146]. Stephens et al. [145] reported an overrepresentation of glycosyltransferase family and sialyl-transferase activity in toxic dinoflagellates. This indicates that the voltage-gated ion channels in toxin producing dinoflagellates are resistant to their own toxins due to processing and attachment of sialic acids to other macromolecules [145]. However, the use of sialic acid to either increase or recover the activity of toxin-resistant channels in dinoflagellates remain to be fully understood.
3.1.1.6. PKS Gene Regulation
Gene regulation and its role in toxin biosynthesis still remains unclear in dinoflagellates since the expression levels of most transcripts in toxic dinoflagellates remain unchanged despite varying culturing conditions or growth phases [147,148,149,150]. Transcription factors are less abundant among dinoflagellates in comparison to other protists, whereas post-transcriptional regulation remains the more dominant regulatory feature of gene expression in these organisms (for a review, see Roy et al. [151]). Most, if not all, of the transcripts in dinoflagellates are post-transcriptionally modified at their 5’ end by trans-splicing, which converts polycistronic pre-mRNAs into mature single gene messages [73,74]. The SL trans-splicing mechanism was first identified in trypanosomes, which carry out continuous transcription and rely entirely on post-transcriptional mechanisms for gene expression [152]. The SL trans-splicing mechanism has now been identified in numerous dinoflagellate species [73,74], and may be an essential requirement for various processes in dinoflagellates that are post-transcriptionally regulated such as bioluminescence [153], carbon fixation [154], photosynthesis [155], circadian regulated processes [147,150], acute stress responses [156], and responses to nitrogen or phosphorus limitation [157].
Microarray analysis in K. brevis showed the expression of PKSs did not change over the diel cycle despite evidence that toxin production is specific to certain diel phases or cell cycle stages in dinoflagellates [147]. Monroe et al. [116] compared the KS domain transcript levels between toxic vs. non-toxic strains of K. brevis and did not find any significant variation. However, at the protein level, KS domain proteins were 55%–70% less expressed in the non-toxic strain, suggesting that the ‘non-toxic’ phenotype in dinoflagellates might not be the result of gene mutations causing a non-functional PKS machinery, but rather a consequence of altered expression levels and/or activity of intact PKS protein complexes [116].
A number of studies have started to report a rich diversity of microRNA (miRNA) in dinoflagellates using a transcriptomic approach [65,158,159,160,161,162]. Additionally, small (or short) interfering RNA (siRNA) have been linked to the regulation of a large fraction of protein coding genes and processes [159]. EST libraries of Alexandrium tamarense have also reported miRNAs that are targeted towards housekeeping genes such as crystalline ribonucleoprotein complex and ribosomal proteins [160]. Genomic surveys of Symbiodiniaceae species have yielded numerous genes involved in primary metabolism of cells that are controlled by miRNAs [65,159]. However, the regulation of PKS toxin biosynthesis by miRNA in dinoflagellates remains to be established.
3.2. Dinoflagellate Toxins: Cyclic Imines
Cyclic imine (CI) toxins are a class of fast-acting neurotoxins that include spirolides, gymnodimines, pinnatoxins, as well as other minor compounds produced by a number dinoflagellate species: Alexandrium ostenfeldii, Karenia selliformis, K. mikimotoi, Prorocentrum lima, P. maculosum, and Vulcanodinium rugorum (for a review, see Stivala et al. [163]). Most of them are yet to undergo extensive transcriptomic studies to identify PKS or other toxin-related genes. A biosynthetic pathway for spirolide C has been proposed on the basis of results from studies using radiolabeled precursor molecules [164,165], and a hypothetical mechanism for pinnatoxin production has also been presented [166]. The structural similarities of spirolides, gymnodimines and pinnatoxins suggest at least a partially shared genetic basis, and it has been hypothesized that spirolides may represent the results of a horizontal gene transfer and recombination event involving gymnodimine and pinnatoxin genes [167]. The few transcriptomic studies have revealed the presence of several PKS genes in strains of spirolide-producing A. ostenfeldii [69,117]. However, the detailed genetic background of CI toxin production remains unknown and specific PKS genes have not yet been directly linked with SPX, GYM, or any of the other toxic CI natural products.
3.3. Dinoflagellate Toxins: Alkaloids
Paralytic shellfish toxins (PSTs) are alkaloid neurotoxins produced by dinoflagellates as well as by prokaryotic cyanobacteria. Saxitoxin (STX) is the parent PST compound originally purified from the clam Saxidomus giganteus [168]. PSTs have a tricyclic perhydropurine backbone, and structural variation occurs due to differing functional groups, resulting in compounds such as the C-toxins, the gonyautoxins (GTX), and the saxitoxin and neosaxitoxin group (e.g., STX and NEO) [169]. In vertebrates, PSTs prevent the transduction of neural signal by blocking voltage-gated ion (Na+, K+, Ca2+) channels, causing paralytic shellfish poisoning (PSP) [170,171,172,173]. Because of its high toxicity, STX has been enlisted as toxin relevant to bioterrorism [174].
PSTs reported in the dinoflagellates can be classified as non-sulfated (STX, NEO), mono-sulfated (GTX 1-6), di-sulfated (C 1-4), decarbamoylated (dcSTX, dcNEO, dcGTX1-4), deoxy-decarbamoyated (doSTX, doGTX1-2) and GC toxin analogues (only reported in Gymnodinium catenatum) [38,171,175,176,177] (Figure 5). The addition of one or more sulfate groups in the basic STX skeleton decreases the toxicity of the resulting molecule compared to the non-sulfated variants [171]. For this reason, STX and NEO are considered to be the most potent PSTs, followed by the gonyautoxins and C- toxins, which contain either one or two sulfate groups, respectively [171]. Approximately eight species of the dinoflagellate genus Alexandrium, as well as the species Gymnodinium catenatum and Pyrodinium bahamense var compressum, are known to produce PSTs [178,179,180,181].
3.3.1. sxt Genes and PST Biosynthesis Pathway in Dinoflagellates
The PST biosynthesis pathway has been studied extensively by investigating radioisotope labelled precursor incorporation since the 1980s [182], but the genes putatively involved in this process, i.e., the sxt gene cluster, were first described from cyanobacteria over 20 years later, and have since been reported from species of approx. seven cyanobacterial genera [148,183,184,185,186,187]. The prokaryotic cyanobacteria and eukaryotic dinoflagellates are only remotely related, but PSTs appear to be synthesized through a similar biosynthetic pathway using the same precursors in both groups of organisms [185,188] (Figure 5). Fourteen of the sxt genes (sxtA – sxtI, sxtP – sxtS and sxtU) have been identified as being common in PST-producing cyanobacteria and are considered “core genes” [185,189]. Amongst them, 8 sxt genes appear to be directly involved in the biosynthetic process [185] (see Figure 5).
The identification of the sxt genes in cyanobacteria provided a platform for the discovery of corresponding genes in dinoflagellates [57,190,191,192]. Transcriptomic annotation with the help of cyanobacterial sxt gene sequences as well as PCR and amplicon sequencing technologies have led to the identification and characterization of sxt homologs in dinoflagellate species [57,190,191,192]. The discovered sxt sequences display features characteristic of dinoflagellate genes; i.e., they have a high (~65%) GC content and are expressed as monocistronic transcripts with a 5’ splice leader sequence and a 3’eukaryotic poly-A tail [122,191] (Figure 6).
In both cyanobacteria and dinoflagellates, STX biosynthesis is initiated by the enzyme sxtA [185,188] (Figure 5). It contains four catalytic domains, encoded by sxtA1 (SAM-dependent methyltransferase, MT), sxtA2 (GCN 5-related N-acetyltransferase, ACT), sxtA3 (acyl carrier protein, ACP), and sxtA4 (8-amino-7-oxononanoate synthase like class II aminotransferase, AONS) [185]. The biosynthesis process starts with the binding of an acetyl-CoA derived acetyl with the ACP of sxtA to form acetyl-ACP, which is converted to propionyl-ACP after methylation of the acetate [185]. The AONS domain catalyzes a Claisen condensation between arginine and propionyl-ACP, thus producing ‘Compound A’ [185] (Figure 5).
In dinoflagellates, at least two sxtA mRNA isoforms have been reported as being simultaneously expressed [57]. Rather than a single homolog, sxtA appears to represent a gene family, with multiple paralogous copies, but only one of these copies appears to be linked to species that synthesize STX [190]. Phylogenetic analysis of sxtA1 reported three paralogs, or clades, of which two, i.e., clades 1 and 3, appear to be widely present in non-STX producing dinoflagellates indicating possible gene duplications events [190]. However, clade 2 of sxtA1 domain appeared to be exclusively made up of STX producing species [190]. The sxtA4 domain was found to be present and highly conserved in several strains of eight species of STX-producing dinoflagellates, but was absent from the non-producing species, thereby confirming the hypothesis that sxtA4 is an essential domain for STX biosynthesis [57,185,189,190]. Zhang et al. [193] used differential expression analysis to report that some paralogs are unrelated to STX biosynthesis, and further confirmed that the sxtA4 domain is essential for STX biosynthesis since no expression of the long sxtA transcript isoform (including the domain sxtA4, as represented in Figure 6) was found in a non-toxic strain of Alexandrium pacificum (Figure 6). However, these transcripts were strongly expressed in an STX-producing strain of the same species [193]. At the same time, no significant differential expression of the short sxtA transcript (excluding sxtA4, as represented in Figure 6) was detected between the two investigated strains [193]. Interestingly, neither sxtA1 nor sxtA4 were found in the recently discovered non-toxic dinoflagellate species Centrodinium punctatum, which appears to be closely related to Alexandrium spp. [194].
The second core gene in dinoflagellate STX biosynthesis is the sxtG, which has been characterized at both transcript and genomic levels (Figure 6), and encodes for an amidinotransferase [57,185,191]. sxtG catalyzes the transfer of a guanidine group from a second arginine to the growing SXT backbone forming Compound B [185] (Figure 5). Like sxtA, dinoflagellate sxtG is homologous to its cyanobacterial counterpart, but has acquired eukaryotic features over the course of its evolution [191]. Though introns are generally not common in dinoflagellate genes, introns of variable lengths have been reported in sxtG in some Alexandrium spp. [191] (Figure 6). Initially, it was reported that sxtG was present in all Alexandrium spp. independent of STX production [57,191,196]. However, using qPCR, sxtG could not be amplified in the STX-producing Alexandrium species [181]. Additionally, no sxtG homologs were found in non-STX producing dinoflagellates in a study examining transcriptomes [190]. The result suggests that if sxtG is present in non-STX producing species, it may be transcribed at extremely low levels [191].
Apart from sxtA and sxtG, other toxin-related genes have not been characterized at the genomic level, and are known from transcriptomic studies involving several PST-producing dinoflagellate species [57,190,191,192,193,195] (Table 1). These include the remaining core sxt genes, which catalyze reactions starting from the third step of the PST biosynthesis pathway, a heterocyclization reaction by sxtB [185] (Figure 5). The following double bond formation is catalyzed by sxtD, and the subsequent synthesis of the second and third heterocycles via epoxidation and aldehyde formation by sxtS [184,185] (Figure 5). Following these steps, the enzymes sxtU and sxtH/T work in sequence to produce dcSTX, the first complete PST analogue, while one further step, carbamoylation by sxtI, produces the parent compound STX [184,185] (Figure 5).
3.3.2. Concepts on sxt Gene Regulation
Complete understanding of toxin biosynthesis in dinoflagellates is challenging due to the unusual genomic features and gene regulation systems in dinoflagellates, as reviewed above. Large percentages of dinoflagellate DNA consists of non-coding repeat sequences with multiple tandem repeats of protein coding genes [64,198,199,200]. Mechanisms such as partial gene duplication and retroposition contribute to this copy number increase, and environmental stress may promote it [71,72,201]. An estimated 5%–30% of the dinoflagellate genes appear to be regulated at the transcriptional level, whereas most other genes may be regulated at the post-transcriptional or translational levels [6,202]. Dinoflagellates can take advantage of the abundant gene copies to help them produce more transcripts [200]. This mechanism is commonly called the ‘dosage effect’, and has also been reported from several other eukaryote organisms [75,203,204]. A study on A. minutum has suggested positive correlation between the genomic copy numbers of sxtA4 with a total cellular PST content [205].
The possibility of transcriptional control of PST biosynthesis has been investigated in Alexandrium spp., and the hypothesis has been supported by the discovery of putative sxtZ homologs, a gene which in cyanobacteria is thought to be involved in the transcriptional regulation of STX biosynthesis [193], as well as the significant positive correlation between sxtA4 expression levels and cellular PST quotas (Qt) under nutritional stress [206]. Whereas a less clear-cut correlation between the expression levels of sxtG and putative sxtI and Qt were reported, an evident effect of macronutrient availability on sxt gene expression was observed [206]. In contrast to these findings, other studies have reported no correlation between sxtA1 and sxtG mRNA quantities and toxin contents in strains of A. minutum [207], and no significant variation in expression levels of the toxin-related gene sxtA4 in A. pacificum or genes sxtA, sxtB, sxtD, sxtF/M, sxtG, sxtH/T, sxtI, sxtO, sxtP, sxtU, sxtW, sxtX, sxtZ, sxtPER, and sxtACT in A. pacificum, as observed between growth stages, while at the same time the intracellular toxin content varied significantly [195,208]. The presence of grazers, while increasing the toxin content and diversity of PST structural variants, did not increase the number of sxtA transcripts in Alexandrium catenella [209]. Together, these findings suggest that the PST biosynthesis genes in dinoflagellates are under a complex regulatory system that may involve genomic, transcriptional as well as post-transcriptional and translational elements.
Several proteomics studies have been performed to provide insight on the mechanisms and possible translational regulation of STX biosynthesis. Correlation between the PST biosynthesis pathway and other carbon and energy utilizing pathways has been observed [210,211]. Proteins involved in the translational machinery, photosynthetic pigment production, and toxin biosynthesis with linkages to arginine, which is an important PST precursor molecule, and glutamate biosynthesis, were found to be upregulated during the same stage of the cell cycle [211,212,213]. Proteins involved in bioluminescence have been observed to be upregulated during toxin production [211] and found to be downregulated in a non-toxic strain with toxin related proteins [210]. Hence, it can be concluded that the toxin biosynthesis might not be regulated as a single pathway rather than being regulated in concert with other biosynthetic pathways.
3.3.3. Applications for Detection and Monitoring of PST-Producing Dinoflagellates
The advances in characterization of the dinoflagellate STX biosynthesis pathway have made it possible to study the role of gene copy numbers and sxt gene expression in the regulation of PST biosynthesis [205], as well as to concentrate efforts on the development sxt-gene based monitoring assays [214,215]. The unprecedented socio-economic impacts of blooms of PST-producing dinoflagellates have driven the efforts to mitigate their impacts [34]. One approach is to use molecular methods such as quantitative polymerase chain reaction (qPCR) to detect and monitor PST-producing species. Several qPCR assays for the detection of Alexandrium spp. have been developed based on rRNA genes (e.g., References [216,217,218,219,220,221]), with specificities and sensitivities down to one cell per litre [221]. However, these assays do not provide any indication regarding the potential toxicity of a bloom as they only detect the presence of the Alexandrium cells.
Furthermore, studies have shown that rRNA gene copy numbers are highly variable [216,217,222], and this may lead to over- or underestimation of cell densities [221,223]. Characterization of sxt genes has made the development of PST-producing strain specific qPCR assays [214,215,224,225], and studies have shown that the genomic copy numbers of sxt genes vary less compared to rRNA genes, allowing for more accurate cell density estimates [197,223,224]. These assays have been successfully trialed to indicate the presence of toxic strains in seawater [215,224,225,226], and in commercially harvested oysters [227]. The detection of the toxic strains at low concentrations in seawater, enabled by sxt gene-based qPCR assays, provides a means for early warning systems designed to detect developing harmful blooms. In the future, combining qPCR with other analysis methods such as meta-transcriptomics could provide even more information on the active metabolic processes related to toxin production throughout bloom events [228].
4. Concluding Remarks
Immense advances have been made in the study of PST biosynthesis and in its application to the aquaculture industry. However, the studies on PKS domains and their relationships to polyketide-based toxins and cyclic imines have barely begun. Most toxin producing dinoflagellates are yet to undergo extensive transcriptomic studies to identify PKSs or other toxin-related genes. PKSs reported from the transcriptomic and genomic surveys of dinoflagellates have yielded divergent homologs, belonging to different phylogenetic sub-clades and producing a diverse range of secondary metabolites, however no toxin or species-specific sub-clades have been identified. Additionally, it is common for some dinoflagellate species to produce a variety of diverse PKS related compounds, impeding our ability to link specific pathways with particular compounds.
The large numbers of PKS domains in dinoflagellates have been linked to the production of numerous undetected and/or uncharacterized polyketide molecules [81]. In the screening hypothesis model [81,125,229], it is suggested that organisms may have selected specific evolutionary traits to increase the probability of developing a compound with potent biomolecular activity—one that enhances the generation and retention of chemical diversity, while also reducing the fitness costs [95]. Hence, organisms may produce numerous biomolecules simultaneously as a cost-effective way of generating chemical diversity, therefore increasing the likelihood of producing a rare molecule with a useful biological activity [95,229,230]. Such compounds could be synthesized as by-products due to the inherent reactivity of chemical intermediates accompanying core catalytic mechanisms [231]. These models and working hypotheses represent a useful lens by which to view the complex array of both polyketide toxic compounds and PKS genes that have begun to be discovered using genetic techniques. The development of targeted approaches, combined with advances in chemical identification of these complex compounds, will allow us to address this issue and advance the discovery of the genetic basis and regulation of dinoflagellate toxins in the future.
Author Contributions
Conceptualization, A.V. and S.A.M.; software, A.V. and P.A.A.; writing—original draft preparation, A.V., H.S., and R.R.; writing—review and editing, A.V., A.B., R.R., H.S., P.A.A, and S.A.M.; Supervision, P.A.A. and S.A.M.; Funding Acquisition, S.A.M.
Funding
This research was funded by Australian Research Council Future Fellowship project FT120100704 to S.A.M.
Acknowledgments
We sincerely thank Gurjeet Singh Kohli for preparing Figure 2 for the manuscript. We also thank the reviewers for constructive comments on an earlier version of this manuscript.
Conflicts of Interest
The authors acknowledge that there are no conflicts of interest.
References
- Caron, D.A.; Worden, A.Z.; Countway, P.D.; Demir, E.; Heidelberg, K.B. Protists are microbes too: a perspective. ISME J. 2009, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, P.G.; Katz, M.E.; Knoll, A.H.; Quigg, A.; Raven, J.A.; Schofield, O.; Taylor, F. The evolution of modern eukaryotic phytoplankton. Science 2004, 305, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Hackett, J.D.; Anderson, D.M.; Erdner, D.L.; Bhattacharya, D. Dinoflagellates: a remarkable evolutionary experiment. Am. J. Bot. 2004, 91, 1523–1534. [Google Scholar] [CrossRef] [PubMed]
- Taylor, F.; Hoppenrath, M.; Saldarriaga, J.F. Dinoflagellate diversity and distribution. Biodivers. Conserv. 2008, 17, 407–418. [Google Scholar] [CrossRef]
- Keeling, P.J.; Burki, F.; Wilcox, H.M.; Allam, B.; Allen, E.E.; Amaral-Zettler, L.A.; Armbrust, E.V.; Archibald, J.M.; Bharti, A.K.; Bell, C.J. The Marine Microbial Eukaryote Transcriptome Sequencing Project (MMETSP): illuminating the functional diversity of eukaryotic life in the oceans through transcriptome sequencing. PLoS Boil. 2014, 12, e1001889. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.A.; Suggett, D.J.; Doblin, M.A.; Kohli, G.S.; Seymour, J.R.; Fabris, M.; Ralph, P.J. Unravelling the functional genetics of dinoflagellates: a review of approaches and opportunities. Perspect. Phycol. 2016, 3, 37–52. [Google Scholar] [CrossRef]
- Ianora, A.; Bentley, M.G.; Caldwell, G.S.; Casotti, R.; Cembella, A.D.; Engström-Öst, J.; Halsband, C.; Sonnenschein, E.; Legrand, C.; Llewellyn, C.A. The relevance of marine chemical ecology to plankton and ecosystem function: an emerging field. Mar. Drugs 2011, 9, 1625–1648. [Google Scholar] [CrossRef] [PubMed]
- Gómez, F. A quantitative review of the lifestyle, habitat and trophic diversity of dinoflagellates (Dinoflagellata, Alveolata). Syst. Biodivers. 2012, 10, 267–275. [Google Scholar] [CrossRef]
- De Vargas, C.; Audic, S.; Henry, N.; Decelle, J.; Mahé, F.; Logares, R.; Lara, E.; Berney, C.; Le Bescot, N.; Probert, I. Eukaryotic plankton diversity in the sunlit ocean. Science 2015, 348, 1261605. [Google Scholar] [CrossRef] [Green Version]
- Le Bescot, N.; Mahé, F.; Audic, S.; Dimier, C.; Garet, M.J.; Poulain, J.; Wincker, P.; de Vargas, C.; Siano, R. Global patterns of pelagic dinoflagellate diversity across protist size classes unveiled by metabarcoding. Environ. Microbiol. 2016, 18, 609–626. [Google Scholar] [CrossRef]
- Schnepf, E.; Elbrächter, M. Nutritional strategies in dinoflagellates: a review with emphasis on cell biological aspects. Eur. J. Protistol. 1992, 28, 3–24. [Google Scholar] [CrossRef]
- Stoecker, D.K. Mixotrophy among Dinoflagellates. J. Eukaryot. Microbiol. 1999, 46, 397–401. [Google Scholar] [CrossRef]
- Litchman, E.; Klausmeier, C.A.; Schofield, O.M.; Falkowski, P.G. The role of functional traits and trade-offs in structuring phytoplankton communities: scaling from cellular to ecosystem level. Ecol. Lett. 2007, 10, 1170–1181. [Google Scholar] [CrossRef]
- Smayda, T.J.; Reynolds, C.S. Strategies of marine dinoflagellate survival and some rules of assembly. J. Sea Res. 2003, 49, 95–106. [Google Scholar] [CrossRef]
- Smayda, T.J. Harmful algal blooms: their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnol. Oceanogr. 1997, 42, 1137–1153. [Google Scholar] [CrossRef]
- Smayda, T.J. What is a bloom? A commentary. Limnol. Oceanogr. 1997, 42, 1132–1136. [Google Scholar] [CrossRef]
- Hallegraeff, G.M. A review of harmful algal blooms and their apparent global increase. Phycologia 1993, 32, 79–99. [Google Scholar] [CrossRef] [Green Version]
- Hallegraeff, G.M. Ocean climate change, phytoplankton community responses, and harmful algal blooms: a formidable predictive challenge. J. Phycol. 2010, 46, 220–235. [Google Scholar] [CrossRef]
- Smayda, T.J. Novel and nuisance phytoplankton blooms in the sea: Evidence for a global epidemic. In Toxic Marine Phytoplankton; Granéli, E., Sundström, B., Edler, L., Anderson, D.M., Eds.; Elsevier: New York, NY. USA, 1990; pp. 29–40. [Google Scholar]
- Zingone, A.; Enevoldsen, H.O. The diversity of harmful algal blooms: a challenge for science and management. Ocean Coast. Manag. 2000, 43, 725–748. [Google Scholar] [CrossRef]
- Cembella, A.D. Chemical ecology of eukaryotic microalgae in marine ecosystems. Phycologia 2003, 42, 420–447. [Google Scholar] [CrossRef]
- Shimizu, Y. Microalgal metabolites. Chem. Rev. 1993, 93, 1685–1698. [Google Scholar] [CrossRef]
- Adolf, J.E.; Krupatkina, D.; Bachvaroff, T.; Place, A.R. Karlotoxin mediates grazing by Oxyrrhis marina on strains of Karlodinium veneficum. Harmful Algae 2007, 6, 400–412. [Google Scholar] [CrossRef]
- Waggett, R.J.; Tester, P.A.; Place, A.R. Anti-grazing properties of the toxic dinoflagellate Karlodinium veneficum during predator–prey interactions with the copepod Acartia tonsa. Mar. Ecol. Prog. Ser. 2008, 366, 31–42. [Google Scholar] [CrossRef]
- Bagøien, E.; Miranda, A.; Reguera, B.; Franco, J.M. Effects of two paralytic shellfish toxin producing dinoflagellates on the pelagic harpacticoid copepod Euterpina acutifrons. Mar. Biol. 1996, 126, 361–369. [Google Scholar]
- Frangópulos, M.; Guisande, C.; Maneiro, I.; Riveiro, I.; Franco, J. Short-term and long-term effects of the toxic dinoflagellate Alexandrium minutum on the copepod Acartia Clausi. Mar. Ecol. Prog. Ser. 2000, 203, 161–169. [Google Scholar] [CrossRef]
- Colin, S.P.; Dam, H.G. Effects of the toxic dinoflagellate Alexandrium fundyense on the copepod Acartia hudsonica: a test of the mechanisms that reduce ingestion rates. Mar. Ecol. Prog. Ser. 2003, 248, 55–65. [Google Scholar] [CrossRef]
- Ianora, A.; Turner, J.T.; Esposito, F.; Carotenuto, Y.; Romano, G.; Fontana, A.; Guisande, C.; Miralto, A. Copepod egg production and hatching success is reduced by maternal diets of a non-neurotoxic strain of the dinoflagellate Alexandrium Tamarense. Mar. Ecol. Prog. Ser. 2004, 280, 199–210. [Google Scholar] [CrossRef]
- Selander, E.; Thor, P.; Toth, G.; Pavia, H. Copepods induce paralytic shellfish toxin production in marine dinoflagellates. Proc. R. Soc. B: Boil. Sci. 2006, 273, 1673–1680. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J.H.; Tester, P.A.; Forward Jr, R.B. Sublethal effects of the toxic dinoflagellate Karenia brevis on marine copepod behavior. J. Plankton Res. 2007, 29, 301–315. [Google Scholar] [CrossRef]
- Waggett, R.J.; Hardison, D.R.; Tester, P.A. Toxicity and nutritional inadequacy of Karenia brevis: synergistic mechanisms disrupt top-down grazer control. Mar. Ecol. Prog. Ser. 2012, 444, 15–30. [Google Scholar] [CrossRef]
- Sunda, W.G.; Burleson, C.; Hardison, D.R.; Morey, J.S.; Wang, Z.; Wolny, J.; Corcoran, A.A.; Flewelling, L.J.; Van Dolah, F.M. Osmotic stress does not trigger brevetoxin production in the dinoflagellate Karenia Brevis. Proc. Nat. Acad. Sci. USA 2013, 110, 10223–10228. [Google Scholar] [CrossRef]
- Davis, C.C. Gymnodinium brevis sp. nov., a cause of discolored water and animal mortality in the Gulf of Mexico. Bot. Gaz. 1948, 109, 358–360. [Google Scholar] [CrossRef]
- Anderson, D.M.; Glibert, P.M.; Burkholder, J.M. Harmful algal blooms and eutrophication: nutrient sources, composition, and consequences. Estuaries 2002, 25, 704–726. [Google Scholar] [CrossRef]
- Shumway, S.E. A review of the effects of algal blooms on shellfish and aquaculture. J. World Aquac. Soc. 1990, 21, 65–104. [Google Scholar] [CrossRef]
- Yasumoto, T.; Murata, M. Marine toxins. Chem. Rev. 1993, 93, 1897–1909. [Google Scholar] [CrossRef]
- Kantiani, L.; Llorca, M.; Sanchís, J.; Farré, M.; Barceló, D. Emerging food contaminants: A review. Anal. Bioanal. Chem. 2010, 398, 2413–2427. [Google Scholar] [CrossRef]
- Wang, D.-Z. Neurotoxins from marine dinoflagellates: a brief review. Mar. Drugs 2008, 6, 349–371. [Google Scholar] [CrossRef]
- Anderson, D.M.; Cembella, A.D.; Hallegraeff, G.M. Progress in understanding harmful algal blooms: paradigm shifts and new technologies for research, monitoring, and management. Annu. Rev. Mar. Sci. 2012, 4, 143–176. [Google Scholar] [CrossRef]
- Kalaitzis, J.A.; Chau, R.; Kohli, G.S.; Murray, S.A.; Neilan, B.A. Biosynthesis of toxic naturally-occurring seafood contaminants. Toxicon 2010, 56, 244–258. [Google Scholar] [CrossRef]
- Blunt, J.W.; Copp, B.R.; Hu, W.-P.; Munro, M.H.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2008, 25, 35–94. [Google Scholar] [CrossRef]
- Veldhuis, M.J.; Cucci, T.L.; Sieracki, M.E. Cellular DNA content of marine phytoplankton using two new fluorochromes: Taxomonic nad ecological implications. J. Phycol. 1997, 33, 527–541. [Google Scholar] [CrossRef]
- Rizzo, P.J. The enigma of the dinoflagellate chromosome. J. Protozool. 1991, 38, 246–252. [Google Scholar] [CrossRef]
- Rizzo, P.J. Those amazing dinoflagellate chromosomes. Cell Res. 2003, 13, 215. [Google Scholar] [CrossRef]
- Rizzo, P.J.; Noodén, L.D. Chromosomal proteins in the dinoflagellate alga Gyrodinium Cohnii. Sci. 1972, 176, 796–797. [Google Scholar] [CrossRef]
- Janouškovec, J.; Gavelis, G.S.; Burki, F.; Dinh, D.; Bachvaroff, T.R.; Gornik, S.G.; Bright, K.J.; Imanian, B.; Strom, S.L.; Delwiche, C.F. Major transitions in dinoflagellate evolution unveiled by phylotranscriptomics. Proc. Nat. Acad. Sci. USA 2017, 114, E171–E180. [Google Scholar]
- Allen, J.; Roberts, T.M.; Loeblich III, A.R.; Klotz, L.C. Characterization of the DNA from the dinoflagellate Crypthecodinium cohnii and implications for nuclear organization. Cell 1975, 6, 161–169. [Google Scholar] [CrossRef]
- Wong, J.T.; New, D.; Wong, J.; Hung, V. Histone-like proteins of the dinoflagellate Crypthecodinium cohnii have homologies to bacterial DNA-binding proteins. Eukaryot. Cell 2003, 2, 646–650. [Google Scholar] [CrossRef]
- Davies, W.; Jakobsen, K.; Nordby, Ø. Characterization of DNA from the Dinoflagellate Woloszynskia Bostoniensis. J. Protozool. 1988, 35, 418–422. [Google Scholar] [CrossRef]
- Roy, S.; Morse, D. A full suite of histone and histone modifying genes are transcribed in the dinoflagellate Lingulodinium. PLoS ONE 2012, 7, e34340. [Google Scholar] [CrossRef]
- Marinov, G.K.; Lynch, M. Diversity and divergence of dinoflagellate histone proteins. G3: Genes, Genomes, Genetics 2016, 6, 397–422. [Google Scholar] [CrossRef]
- Hackett, J.D.; Scheetz, T.E.; Yoon, H.S.; Soares, M.B.; Bonaldo, M.F.; Casavant, T.L.; Bhattacharya, D. Insights into a dinoflagellate genome through expressed sequence tag analysis. BMC Genom. 2005, 6, 80. [Google Scholar] [CrossRef] [Green Version]
- Gornik, S.G.; Ford, K.L.; Mulhern, T.D.; Bacic, A.; McFadden, G.I.; Waller, R.F. Loss of nucleosomal DNA condensation coincides with appearance of a novel nuclear protein in dinoflagellates. Curr. Boil. 2012, 22, 2303–2312. [Google Scholar] [CrossRef]
- Kato, K.H.; Moriyama, A.; Huitorel, P.; Cosson, J.; Cachon, M.; Sato, H. Isolation of the major basic nuclear protein and its localization on chromosomes of the dinoflagellate, Oxyrrhis Mar. Boil. Cell 1997, 89, 43–52. [Google Scholar] [CrossRef]
- Irwin, N.A.; Martin, B.J.; Young, B.P.; Browne, M.J.; Flaus, A.; Loewen, C.J.; Keeling, P.J.; Howe, L.J. Viral proteins as a potential driver of histone depletion in dinoflagellates. Nat. Commun. 2018, 9, 1535. [Google Scholar] [CrossRef]
- Shoguchi, E.; Shinzato, C.; Kawashima, T.; Gyoja, F.; Mungpakdee, S.; Koyanagi, R.; Takeuchi, T.; Hisata, K.; Tanaka, M.; Fujiwara, M. Draft assembly of the Symbiodinium minutum nuclear genome reveals dinoflagellate gene structure. Curr. Boil. 2013, 23, 1399–1408. [Google Scholar] [CrossRef]
- Stüken, A.; Orr, R.J.; Kellmann, R.; Murray, S.A.; Neilan, B.A.; Jakobsen, K.S. Discovery of nuclear-encoded genes for the neurotoxin saxitoxin in dinoflagellates. PLoS ONE 2011, 6, e20096. [Google Scholar] [CrossRef]
- Rae, P. Hydroxymethyluracil in eukaryote DNA: a natural feature of the pyrrophyta (dinoflagellates). Science 1976, 194, 1062–1064. [Google Scholar] [CrossRef]
- Rae, P.M.; Steele, R.E. Modified bases in the DNAs of unicellular eukaryotes: an examination of distributions and possible roles, with emphasis on hydroxymethyluracil in dinoflagellates. Biosystems 1978, 10, 37–53. [Google Scholar] [CrossRef]
- Hackett, J.D.; Yoon, H.S.; Soares, M.B.; Bonaldo, M.F.; Casavant, T.L.; Scheetz, T.E.; Nosenko, T.; Bhattacharya, D. Migration of the plastid genome to the nucleus in a peridinin dinoflagellate. Curr. Boil. 2004, 14, 213–218. [Google Scholar] [CrossRef]
- Yoon, H.S.; Hackett, J.D.; Van Dolah, F.M.; Nosenko, T.; Lidie, K.L.; Bhattacharya, D. Tertiary endosymbiosis driven genome evolution in dinoflagellate algae. Mol. Boil. Evol. 2005, 22, 1299–1308. [Google Scholar] [CrossRef]
- Patron, N.J.; Waller, R.F.; Archibald, J.M.; Keeling, P.J. Complex protein targeting to dinoflagellate plastids. J. Mol. Boil. 2005, 348, 1015–1024. [Google Scholar] [CrossRef]
- Lin, S. Genomic understanding of dinoflagellates. Res. Microbiol. 2011, 162, 551–569. [Google Scholar] [CrossRef]
- Hou, Y.; Lin, S. Distinct gene number-genome size relationships for eukaryotes and non-eukaryotes: Gene content estimation for dinoflagellate genomes. PLoS ONE 2009, 4, e6978. [Google Scholar] [CrossRef]
- Lin, S.; Cheng, S.; Song, B.; Zhong, X.; Lin, X.; Li, W.; Li, L.; Zhang, Y.; Zhang, H.; Ji, Z. The Symbiodinium kawagutii genome illuminates dinoflagellate gene expression and coral symbiosis. Science 2015, 350, 691–694. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Lambert, G.; Andersen, R.A.; Coffroth, M.A.; Galbraith, D.W. Symbiodinium (Pyrrhophyta) genome sizes (DNA content) are smallest amongst dinoflagelaltes. J. Phycol. 2005, 41, 880–886. [Google Scholar] [CrossRef]
- John, U.; Lu, Y.; Wohlrab, S.; Groth, M.; Janouškovec, J.; Kohli, G.S.; Mark, F.C.; Bickmeyer, U.; Farhat, S.; Felder, M. An aerobic eukaryotic parasite with functional mitochondria that likely lacks a mitochondrial genome. Sci. Adv. 2019, 5, eaav1110. [Google Scholar] [CrossRef] [Green Version]
- Bachvaroff, T.R.; Place, A.R. From stop to start: tandem gene arrangement, copy number and trans-splicing sites in the dinoflagellate Amphidinium Carterae. PLoS ONE 2008, 3, e2929. [Google Scholar] [CrossRef]
- Jaeckisch, N.; Yang, I.; Wohlrab, S.; Glöckner, G.; Kroymann, J.; Vogel, H.; Cembella, A.; John, U. Comparative Genomic and Transcriptomic Characterization of the Toxigenic Marine Dinoflagellate Alexandrium Ostenfeldii. PLoS ONE 2011, 6, e28012. [Google Scholar] [CrossRef]
- Beauchemin, M.; Roy, S.; Daoust, P.; Dagenais-Bellefeuille, S.; Bertomeu, T.; Letourneau, L.; Lang, B.F.; Morse, D. Dinoflagellate tandem array gene transcripts are highly conserved and not polycistronic. Proc. Nat. Acad. Sci. USA 2012, 109, 15793–15798. [Google Scholar] [CrossRef] [Green Version]
- Slamovits, C.H.; Keeling, P.J. Widespread recycling of processed cDNAs in dinoflagellates. Curr. Boil. 2008, 18, R550–R552. [Google Scholar] [CrossRef] [Green Version]
- Song, B.; Morse, D.; Song, Y.; Fu, Y.; Lin, X.; Wang, W.; Cheng, S.; Chen, W.; Liu, X.; Lin, S. Comparative genomics reveals two major bouts of gene retroposition coinciding with crucial periods of Symbiodinium evolution. Genome Boil. Evol. 2017, 9, 2037–2047. [Google Scholar] [CrossRef]
- Zhang, H.; Hou, Y.; Miranda, L.; Campbell, D.A.; Sturm, N.R.; Gaasterland, T.; Lin, S. Spliced leader RNA trans-splicing in dinoflagellates. Proc. Nat. Acad. Sci. USA 2007, 104, 4618–4623. [Google Scholar] [CrossRef] [Green Version]
- Lidie, K.B.; Van Dolah, F.M. Spliced leader RNA-mediated trans-splicing in a dinoflagellate, Karenia Brevis. J. Eukaryot. Microbiol. 2007, 54, 427–435. [Google Scholar] [CrossRef]
- Zhang, H.; Campbell, D.A.; Sturm, N.R.; Lin, S. Dinoflagellate spliced leader RNA genes display a variety of sequences and genomic arrangements. Mol. Boil. Evol. 2009, 26, 1757–1771. [Google Scholar] [CrossRef]
- Wisecaver, J.H.; Hackett, J.D. Dinoflagellate genome evolution. Annu. Rev. Microbiol. 2011, 65, 369–387. [Google Scholar] [CrossRef]
- Waller, R.F.; Jackson, C.J. Dinoflagellate mitochondrial genomes: stretching the rules of molecular biology. Bioessays 2009, 31, 237–245. [Google Scholar] [CrossRef]
- Howe, C.J.; Nisbet, R.E.R.; Barbrook, A.C. The remarkable chloroplast genome of dinoflagellates. J. Exp. Bot. 2008, 59, 1035–1045. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Green, B.; Cavalier-Smith, T. Single gene circles in dinoflagellate chloroplast genomes. Nature 1999, 400, 155. [Google Scholar] [CrossRef]
- Bachvaroff, T.R.; Concepcion, G.T.; Rogers, C.R.; Herman, E.M.; Delwiche, C.F. Dinoflagellate Expressed Sequence Tag data indicate massive transfer of chloroplast genes to the nuclear genome. Protist 2004, 155, 65–78. [Google Scholar] [CrossRef]
- Kohli, G.S.; John, U.; Van Dolah, F.M.; Murray, S.A. Evolutionary distinctiveness of fatty acid and polyketide synthesis in eukaryotes. ISME J. 2016, 10, 1877. [Google Scholar] [CrossRef]
- Akbar, M.; Ahmad, A.; Usup, G.; Bunawan, H. RNA-Seq as an emerging tool for marine dinoflagellate transcriptome analysis: process and challenges. Processes 2018, 6, 5. [Google Scholar] [CrossRef]
- Lidie, K.B.; Ryan, J.C.; Barbier, M.; Van Dolah, F.M. Gene expression in Florida red tide dinoflagellate Karenia brevis: analysis of an expressed sequence tag library and development of DNA microarray. Mar. Biotechnol. 2005, 7, 481–493. [Google Scholar] [CrossRef]
- Akbar, M.; Ahmad, A.; Usup, G.; Bunawan, H. Current Knowledge and Recent Advances in Marine Dinoflagellate Transcriptomic Research. J. Mar. Sci. Eng. 2018, 6, 13. [Google Scholar] [CrossRef]
- Hennon, G.M.; Dyhrman, S.T. Progress and promise of omics for predicting the impacts of climate change on harmful algal blooms. Harmful Algae 2019. [Google Scholar] [CrossRef]
- Camacho, F.G.; Rodríguez, J.G.; Mirón, A.S.; García, M.C.; Belarbi, E.; Chisti, Y.; Grima, E.M. Biotechnological significance of toxic marine dinoflagellates. Biotechnol. Adv. 2007, 25, 176–194. [Google Scholar] [CrossRef]
- Rein, K.S.; Borrone, J. Polyketides from dinoflagellates: origins, pharmacology and biosynthesis. Comp. Biochem. Physiol. Part B: Biochem. Mol. Boil. 1999, 124, 117–131. [Google Scholar] [CrossRef]
- Kellmann, R.; Stüken, A.; Orr, R.J.; Svendsen, H.M.; Jakobsen, K.S. Biosynthesis and molecular genetics of polyketides in marine dinoflagellates. Mar. Drugs 2010, 8, 1011–1048. [Google Scholar] [CrossRef]
- Staunton, J.; Weissman, K.J. Polyketide biosynthesis: a millennium review. Nat. Prod. Rep. 2001, 18, 380–416. [Google Scholar] [CrossRef]
- Hertweck, C. The biosynthetic logic of polyketide diversity. Angew. Chem. Int. Ed. 2009, 48, 4688–4716. [Google Scholar] [CrossRef]
- Hopwood, D.A. Genetic contributions to understanding polyketide synthases. Chem. Rev. 1997, 97, 2465–2498. [Google Scholar] [CrossRef]
- Shen, B. Polyketide biosynthesis beyond the type I, II and III polyketide synthase paradigms. Curr. Opin. Chem. Boil. 2003, 7, 285–295. [Google Scholar] [CrossRef]
- Donadio, S.; Staver, M.J.; McAlpine, J.B.; Swanson, S.J.; Katz, L. Modular organization of genes required for complex polyketide biosynthesis. Science 1991, 252, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Jenke-Kodama, H.; Sandmann, A.; Muüller, R.; Dittmann, E. Evolutionary implications of bacterial polyketide synthases. Mol. Boil. Evol. 2005, 22, 2027–2039. [Google Scholar] [CrossRef] [PubMed]
- Jenke-Kodama, H.; Dittmann, E. Evolution of metabolic diversity: insights from microbial polyketide synthases. Phytochemistry 2009, 70, 1858–1866. [Google Scholar] [CrossRef] [PubMed]
- Hopwood, D.A.; Sherman, D.H. Molecular genetics of polyketides and its comparison to fatty acid biosynthesis. Annu. Rev. Genet. 1990, 24, 37–62. [Google Scholar] [CrossRef] [PubMed]
- Smith, S. The animal fatty acid synthase: one gene, one polypeptide, seven enzymes. FASEB J. 1994, 8, 1248–1259. [Google Scholar] [CrossRef] [PubMed]
- Cane, D.E.; Walsh, C.T.; Khosla, C. Harnessing the biosynthetic code: combinations, permutations, and mutations. Science 1998, 282, 63–68. [Google Scholar] [CrossRef]
- Khosla, C. Harnessing the biosynthetic potential of modular polyketide synthases. Chem. Rev. 1997, 97, 2577–2590. [Google Scholar] [CrossRef]
- Khosla, C.; Gokhale, R.S.; Jacobsen, J.R.; Cane, D.E. Tolerance and specificity of polyketide synthases. Annu. Rev. Biochem. 1999, 68, 219–253. [Google Scholar] [CrossRef]
- Schwarzer, D.; Finking, R.; Marahiel, M.A. Nonribosomal peptides: from genes to products. Nat. Prod. Rep. 2003, 20, 275–287. [Google Scholar] [CrossRef]
- Marahiel, M.A.; Stachelhaus, T.; Mootz, H.D. Modular peptide synthetases involved in nonribosomal peptide synthesis. Chem. Rev. 1997, 97, 2651–2674. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A.; Marahiel, M.A. Genetic evidence for a role of thioesterase domains, integrated in or associated with peptide synthetases, in non-ribosomal peptide biosynthesis in Bacillus Subtilis. Arch. Microbiol. 1998, 169, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Wakil, S.J. Fatty acid synthase, a proficient multifunctional enzyme. Biochemistry 1989, 28, 4523–4530. [Google Scholar] [CrossRef] [PubMed]
- Harwood, J.L. Fatty acid metabolism. Annu. Rev. Plant Physiol. Plant Mol. Boil. 1988, 39, 101–138. [Google Scholar] [CrossRef]
- Snyder, R.; Gibbs, P.; Palacios, A.; Abiy, L.; Dickey, R.; Lopez, J.V.; Rein, K. Polyketide synthase genes from marine dinoflagellates. Mar. Biotechnol. 2003, 5, 1–12. [Google Scholar] [PubMed]
- Zhu, G.; LaGier, M.J.; Stejskal, F.; Millership, J.J.; Cai, X.; Keithly, J.S. Cryptosporidium parvum: the first protist known to encode a putative polyketide synthase. Gene 2002, 298, 79–89. [Google Scholar] [CrossRef]
- Snyder, R.V.; Guerrero, M.A.; Sinigalliano, C.D.; Winshell, J.; Perez, R.; Lopez, J.V.; Rein, K.S. Localization of polyketide synthase encoding genes to the toxic dinoflagellate Karenia Brevis. Phytochem. 2005, 66, 1767–1780. [Google Scholar] [CrossRef] [PubMed]
- Monroe, E.A.; Van Dolah, F.M. The toxic dinoflagellate Karenia brevis encodes novel type I-like polyketide synthases containing discrete catalytic domains. Protist 2008, 159, 471–482. [Google Scholar] [CrossRef]
- Sonnenborn, U.; Kunau, W.-H. Purification and properties of the fatty acid synthetase complex from the marine dinoflagellate, Crypthecodinium Cohnii. Biochim. Et Biophys. Acta (BBA)-Lipids Lipid Metab. 1982, 712, 523–534. [Google Scholar] [CrossRef]
- Waller, R.F.; Keeling, P.J.; Donald, R.G.K.; Striepen, B.; Handman, E.; Lang-Unnasch, N.; Cowman, A.F.; Besra, G.S.; Roos, D.S.; McFadden, G.I. Nuclear-encoded proteins target to the plastid in Toxoplasma gondii and Plasmodium Falciparum. Proc. Nat. Acad. Sci. USA 1998, 95, 12352–12357. [Google Scholar] [CrossRef]
- Van Dolah, F.M.; Zippay, M.L.; Pezzolesi, L.; Rein, K.S.; Johnson, J.G.; Morey, J.S.; Wang, Z.; Pistocchi, R. Subcellular localization of dinoflagellate polyketide synthases and fatty acid synthase activity. J. Phycol. 2013, 49, 1118–1127. [Google Scholar] [CrossRef] [PubMed]
- Pawlowiez, R.; Morey, J.; Darius, H.; Chinain, M.; Van Dolah, F. Transcriptome sequencing reveals single domain Type I-like polyketide synthases in the toxic dinoflagellate Gamb. Polyn. Harmful Algae 2014, 36, 29–37. [Google Scholar] [CrossRef]
- Kroken, S.; Glass, N.L.; Taylor, J.W.; Yoder, O.; Turgeon, B.G. Phylogenomic analysis of type I polyketide synthase genes in pathogenic and saprobic ascomycetes. Proc. Nat. Acad. Sci. USA 2003, 100, 15670–15675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- John, U.; Beszteri, B.; Derelle, E.; Van de Peer, Y.; Read, B.; Moreau, H.; Cembella, A. Novel insights into evolution of protistan polyketide synthases through phylogenomic analysis. Protist 2008, 159, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Monroe, E.A.; Johnson, J.G.; Wang, Z.; Pierce, R.K.; Van Dolah, F.M. Characterization and expression of nuclear-encoded polyketide synthases in the brevetoxin-producing dinoflagellate Karenia Brevis. J. Phycol. 2010, 46, 541–552. [Google Scholar] [CrossRef]
- Eichholz, K.; Beszteri, B.; John, U. Putative monofunctional type I polyketide synthase units: a dinoflagellate-specific feature? PLoS ONE 2012, 7, e48624. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, T.; Upadhyay, R.J.; Nagasaki, K.; Bhattacharya, D. Dozens of toxin-related genes are expressed in a nontoxic strain of the dinoflagellate Heterocapsa Circ. Mol. Boil. Evol. 2012, 29, 1503–1506. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Okuda, S.; Nakayama, K.; Shikata, T.; Takahashi, F.; Yamaguchi, H.; Skamoto, S.; Yamaguchi, M.; Tomaru, Y. RNA Sequencing Revealed Numerous Polyketide Synthase Genes in the Harmful Dinoflagellate Karenia Mikimotoi. PLoS ONE 2015, 10, e0142731. [Google Scholar] [CrossRef]
- Kohli, G.S.; John, U.; Figueroa, R.I.; Rhodes, L.L.; Harwood, D.T.; Groth, M.; Bolch, C.J.; Murray, S.A. Polyketide synthesis genes associated with toxin production in two species of Gambierdiscus (Dinophyceae). BMC Genom. 2015, 16, 410. [Google Scholar] [CrossRef]
- Meyer, J.M.; Rödelsperger, C.; Eichholz, K.; Tillmann, U.; Cembella, A.; McGaughran, A.; John, U. Transcriptomic characterisation and genomic glimps into the toxigenic dinoflagellate Azadinium spinosum, with emphasis on polykeitde synthase genes. BMC Genom. 2015, 16, 27. [Google Scholar] [CrossRef]
- Kohli, G.S.; Campbell, K.; John, U.; Smith, K.F.; Fraga, S.; Rhodes, L.L.; Murray, S.A. Role of Modular Polyketide Synthases in the Production of Polyether Ladder Compounds in Ciguatoxin-Producing Gambierdiscus polynesiensis and G. excentricus (Dinophyceae). J. Eukaryot. Microbiol. 2017, 64, 691–706. [Google Scholar] [CrossRef] [PubMed]
- Van Dolah, F.M.; Kohli, G.S.; Morey, J.S.; Murray, S.A. Both modular and single-domain Type I polyketide synthases are expressed in the brevetoxin-producing dinoflagellate, Karenia brevis (Dinophyceae). J. Phycol. 2017, 53, 1325–1339. [Google Scholar] [CrossRef] [PubMed]
- Beedessee, G.; Hisata, K.; Roy, M.C.; Satoh, N.; Shoguchi, E. Multifunctional polyketide synthase genes identified by genomic survey of the symbiotic dinoflagellate, Symbiodinium Minutum. BMC Genom. 2015, 16, 941. [Google Scholar] [CrossRef] [PubMed]
- Beedessee, G.; Hisata, K.; Roy, M.C.; Van Dolah, F.M.; Satoh, N.; Shoguchi, E. Diversified secondary metabolite biosynthesis gene repertoire revealed in symbiotic dinoflagellates. Sci. Rep. 2019, 9, 1204. [Google Scholar] [CrossRef] [PubMed]
- Kubota, T.; Iinuma, Y.; Kobayashi, J. Cloning of polyketide synthase genes from amphidinolide-producing dinoflagellate Amphidinium sp. Boil. Pharm. Bull. 2006, 29, 1314–1318. [Google Scholar] [CrossRef] [PubMed]
- Bachvaroff, T.R.W.; Jagus, R.; Place, A.R. A noncryptic noncanonical multi-module PKS/NRPS found in dinoflagellates. In Proceedings of the 16th International Conference on Harmful Algae, Wellington, New Zealand, 27–31 October 2014; pp. 101–104. [Google Scholar]
- Franke, J.; Ishida, K.; Hertweck, C. Genomics-driven discovery of burkholderic acid, a noncanonical, cryptic polyketide from human pathogenic Burkholderia species. Angew. Chem. Int. Ed. 2012, 51, 11611–11615. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Kohli, G.; Harwood, D.T.; Ralph, P.J.; Murray, S.A. Transcriptomic investigation into polyketide toxin synthesis in Ostreopsis (Dinophyceae) species. Environ. Microbiol. In review.
- López-Legentil, S.; Song, B.; DeTure, M.; Baden, D.G. Characterization and localization of a hybrid non-ribosomal peptide synthetase and polyketide synthase gene from the toxic dinoflagellate Karenia Brevis. Mar. Biotechnol. 2010, 12, 32–41. [Google Scholar] [CrossRef]
- Piel, J. Biosynthesis of polyketides by trans-AT polyketide synthases. Nat. Prod. Rep. 2010, 27, 996–1047. [Google Scholar] [CrossRef]
- Cheng, Y.-Q.; Tang, G.-L.; Shen, B. Type I polyketide synthase requiring a discrete acyltransferase for polyketide biosynthesis. Proc. Nat. Acad. Sci. USA 2003, 100, 3149–3154. [Google Scholar] [CrossRef] [Green Version]
- Shelest, E.; Heimerl, N.; Fichtner, M.; Sasso, S. Multimodular type I polyketide synthases in algae evolve by module duplications and displacement of AT domains in trans. BMC Genom. 2015, 16, 1015. [Google Scholar] [CrossRef]
- Nguyen, T.; Ishida, K.; Jenke-Kodama, H.; Dittmann, E.; Gurgui, C.; Hochmuth, T.; Taudien, S.; Platzer, M.; Hertweck, C.; Piel, J. Exploiting the mosaic structure of trans-acyltransferase polyketide synthases for natural product discovery and pathway dissection. Nat. Biotechnol. 2008, 26, 225. [Google Scholar] [CrossRef]
- Keatinge-Clay, A.T. A tylosin ketoreductase reveals how chirality is determined in polyketides. Chem. Boil. 2007, 14, 898–908. [Google Scholar] [CrossRef]
- Bonnett, S.A.; Whicher, J.R.; Papireddy, K.; Florova, G.; Smith, J.L.; Reynolds, K.A. Structural and stereochemical analysis of a modular polyketide synthase ketoreductase domain required for the generation of a cis-alkene. Chem. Boil. 2013, 20, 772–783. [Google Scholar] [CrossRef]
- Lee, M.S.; Qin, G.; Nakanishi, K.; Zagorski, M.G. Biosynthetic studies of brevetoxins, potent neurotoxins produced by the dinoflagellate Gymnodinium Breve. J. Am. Chem. Soc. 1989, 111, 6234–6241. [Google Scholar] [CrossRef]
- Nakanishi, K. The chemistry of brevetoxins: a review. Toxicon 1985, 23, 473–479. [Google Scholar] [CrossRef]
- Martin, J.L.; McMillan, F.M. SAM (dependent) I AM: the S-adenosylmethionine-dependent methyltransferase fold. Curr. Opin. Struct. Boil. 2002, 12, 783–793. [Google Scholar] [CrossRef]
- Miller, D.J.; Ouellette, N.; Evdokimova, E.; Savchenko, A.; Edwards, A.; Anderson, W.F. Crystal complexes of a predicted S-adenosylmethionine-dependent methyltransferase reveal a typical AdoMet binding domain and a substrate recognition domain. Protein Sci. 2003, 12, 1432–1442. [Google Scholar] [CrossRef] [Green Version]
- Murata, M.; Naoki, H.; Iwashita, T.; Matsunaga, S.; Sasaki, M.; Yokoyama, A.; Yasumoto, T. Structure of maitotoxin. J. Am. Chem. Soc. 1993, 115, 2060–2062. [Google Scholar] [CrossRef]
- Moore, R.E.; Bartolini, G. Structure of palytoxin. J. Am. Chem. Soc. 1981, 103, 2491–2494. [Google Scholar] [CrossRef]
- Keatinge-Clay, A.T. The structures of type I polyketide synthases. Nat. Prod. Rep. 2012, 29, 1050–1073. [Google Scholar] [CrossRef]
- Keatinge-Clay, A.T.; Stroud, R.M. The structure of a ketoreductase determines the organization of the β-carbon processing enzymes of modular polyketide synthases. Structure 2006, 14, 737–748. [Google Scholar] [CrossRef]
- Stephens, T.G.; Ragan, M.A.; Bhattacharya, D.; Chan, C.X. Core genes in diverse dinoflagellate lineages include a wealth of conserved dark genes with unknown functions. Sci. Rep. 2018, 8, 17175. [Google Scholar] [CrossRef]
- Marban, E.; Yamagishi, T.; Tomaselli, G.F. Structure and function of voltage-gated sodium channels. J. Physiol. 1998, 508, 647–657. [Google Scholar] [CrossRef]
- Van Dolah, F.M.; Lidie, K.B.; Morey, J.S.; Brunelle, S.A.; Ryan, J.C.; Monroe, E.A.; Haynes, B.L. microarray analysis of diurnal-and circadian-regulated genes in the florida red-tide dinoflagellate Karenia brevis (Dinophyceae). J. Phycol. 2007, 43, 741–752. [Google Scholar] [CrossRef]
- Moustafa, A.; Loram, J.E.; Hackett, J.D.; Anderson, D.M.; Plumley, F.G.; Bhattacharya, D. Origin of Saxitoxin Biosynthetic Genes in Cyanobacteria. PLoS ONE 2009, 4, e5758. [Google Scholar] [CrossRef]
- Yang, I.; John, U.; Beszteri, S.; Glöckner, G.; Krock, B.; Goesmann, A.; Cembella, A.D. Comparative gene expression in toxic versus non-toxic strains of the marine dinoflagellate Alexandrium Minutum. BMC Genom. 2010, 11, 248. [Google Scholar] [CrossRef]
- Brunelle, S.A.; Van Dolah, F.M. Post-transcriptional regulation of S-phase genes in the dinoflagellate, Karenia Brevis. J. Eukaryot. Microbiol. 2011, 58, 373–382. [Google Scholar] [CrossRef]
- Roy, S.; Jagus, R.; Morse, D. Translation and translational control in Dinoflagellates. Microorganisms 2018, 6, 30. [Google Scholar] [CrossRef]
- Palenchar, J.B.; Bellofatto, V. Gene transcription in trypanosomes. Mol. Biochem. Parasitol. 2006, 146, 135–141. [Google Scholar] [CrossRef]
- Mittag, M.; Li, L.; Hastings, J.W. The mRNA Level of the Circadian Regulated Gonyaulax Luciferase Remains Constant over the Cycle. Chronobiol. Int. 1998, 15, 93–98. [Google Scholar] [CrossRef]
- Fagan, T.; Morse, D.; Hastings, J.W. Circadian synthesis of a nuclear-encoded chloroplast glyceraldehyde-3-phosphate dehydrogenase in the dinoflagellate Gonyaulax polyedra is translationally controlled. Biochemistry 1999, 38, 7689–7695. [Google Scholar] [CrossRef]
- Le, Q.H.; Jovine, R.; Markovic, P.; Morse, D. Peridinin-chlorophyll a-protein is not implicated in the photosynthesis rhythm of the dinoflagellate Gonyaulax despite circadian regulation of its translation. Boil. Rhythm. Res. 2001, 32, 579–594. [Google Scholar] [CrossRef]
- Lidie, K.; Van Dolah, F. Characterization and regulation of gene expression networks in response to acute stress in Karenia brevis. In Proceedings of the Fourth Symposium on Harmful Algae in the US, Woods Hole, MA, USA, 28 October–1 November 2007; p. 54. [Google Scholar]
- Morey, J.S.; Monroe, E.A.; Kinney, A.L.; Beal, M.; Johnson, J.G.; Hitchcock, G.L.; Van Dolah, F.M. Transcriptomic response of the red tide dinoflagellate, Karenia brevis, to nitrogen and phosphorus depletion and addition. BMC Genom. 2011, 12, 346. [Google Scholar] [CrossRef]
- Dagenais-Bellefeuille, S.; Beauchemin, M.; Morse, D. miRNAs do not regulate circadian protein synthesis in the dinoflagellate Lingulodinium Polyedrum. PLoS ONE 2017, 12, e0168817. [Google Scholar] [CrossRef]
- Baumgarten, S.; Bayer, T.; Aranda, M.; Liew, Y.J.; Carr, A.; Micklem, G.; Voolstra, C.R. Integrating microRNA and mRNA expression profiling in Symbiodinium microadriaticum, a dinoflagellate symbiont of reef-building corals. BMC Genom. 2013, 14, 704. [Google Scholar] [CrossRef]
- Gao, D.; Qiu, L.; Hou, Z.; Zhang, Q.; Wu, J.; Gao, Q.; Song, L. Computational identification of microRNAs from the expressed sequence tags of toxic dinoflagellate Alexandrium Tamarense. Evol. Bioinform. 2013, 9. [Google Scholar] [CrossRef]
- Geng, H.; Sui, Z.; Zhang, S.; Du, Q.; Ren, Y.; Liu, Y.; Kong, F.; Zhong, J.; Ma, Q. Identification of microRNAs in the toxigenic Dinoflagellate Alexandrium catenella by high-throughput illumina sequencing and bioinformatic analysis. PLoS ONE 2015, 10, e0138709. [Google Scholar] [CrossRef]
- Shi, X.; Lin, X.; Li, L.; Li, M.; Palenik, B.; Lin, S. Transcriptomic and microRNAomic profiling reveals multi-faceted mechanisms to cope with phosphate stress in a dinoflagellate. ISME J. 2017, 11, 2209. [Google Scholar] [CrossRef]
- Stivala, C.E.; Benoit, E.; Araoz, R.; Servent, D.; Novikov, A.; Molgó, J.; Zakarian, A. Synthesis and biology of cyclic imine toxins, an emerging class of potent, globally distributed marine toxins. Nat. Prod. Rep. 2015, 32, 411–435. [Google Scholar] [CrossRef]
- MacKinnon, S.L.; Walter, J.A.; Quilliam, M.A.; Cembella, A.D.; LeBlanc, P.; Burton, I.W.; Hardstaff, W.R.; Lewis, N.I. Spirolides Isolated from Danish Strains of the Toxigenic Dinoflagellate Alexandrium o stenfeldii. J. Nat. Prod. 2006, 69, 983–987. [Google Scholar] [CrossRef]
- Anttila, M.; Strangman, W.; York, R.; Tomas, C.; Wright, J.L. Biosynthetic Studies of 13-Desmethylspirolide C Produced by Alexandrium ostenfeldii (= A. peruvianum): Rationalization of the Biosynthetic Pathway Following Incorporation of 13C-Labeled Methionine and Application of the Odd–Even Rule of Methylation. J. Nat. Prod. 2015, 79, 484–489. [Google Scholar] [CrossRef]
- Kuramoto, M.; Arimoto, H.; Uemura, D. Bioactive alkaloids from the sea: a review. Mar. Drugs 2004, 2, 39–54. [Google Scholar] [CrossRef]
- Van Wagoner, R.M.; Misner, I.; Tomas, C.R.; Wright, J.L. Occurrence of 12-methylgymnodimine in a spirolide-producing dinoflagellate Alexandrium peruvianum and the biogenetic implications. Tetrahedron Lett. 2011, 52, 4243–4246. [Google Scholar] [CrossRef]
- Schantz, E.J.; Mold, J.D.; Stanger, D.W.; Shavel, J.; Riel, F.J.; Bowden, J.P.; Lynch, J.M.; Wyler, R.S.; Riegel, B.; Sommer, H. Paralytic shellfish poison. VI. A procedure for the isolation and purification of the poison from toxic clam and mussel tissues. J. Am. Chem. Soc. 1957, 79, 5230–5235. [Google Scholar] [CrossRef]
- Schantz, E.J.; Ghazarossian, V.E.; Schnoes, H.K.; Strong, F.M.; Springer, J.P.; Pezzanite, J.O.; Clardy, J. Structure of saxitoxin. J. Am. Chem. Soc. 1975, 97, 1238–1239. [Google Scholar] [CrossRef]
- Cestèle, S.; Catterall, W.A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar] [CrossRef]
- Llewellyn, L.E. Saxitoxin, a toxic marine natural product that targets a multitude of receptors. Nat. Prod. Rep. 2006, 23, 200–222. [Google Scholar] [CrossRef]
- Su, Z.; Sheets, M.; Ishida, H.; Li, F.; Barry, W.H. Saxitoxin Blocks L-Type ICa. J. Pharmacol. Exp. Ther. 2004, 308, 324–329. [Google Scholar] [CrossRef]
- Wang, J.; Salata, J.J.; Bennett, P.B. Saxitoxin Is a Gating Modifier of hERG K+ Channels. J. Gen. Physiol. 2003, 121, 583–598. [Google Scholar] [CrossRef]
- Anderson, P.D. Bioterrorism: Toxins as Weapons. J. Pharm. Pr. 2012, 25, 121–129. [Google Scholar] [CrossRef]
- Wang, D.-Z.; Zhang, S.-F.; Zhang, Y.; Lin, L. Paralytic shellfish toxin biosynthesis in cyanobacteria and dinoflagellates: A molecular overview. J. Proteom. 2016, 135, 132–140. [Google Scholar] [CrossRef]
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic Alkaloids: Saxitoxin and Its Analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [Green Version]
- Suikkanen, S.; Kremp, A.; Hautala, H.; Krock, B. Paralytic shellfish toxins or spirolides? The role of environmental and genetic factors in toxin production of the Alexandrium ostenfeldii complex. Harmful Algae 2013, 26, 52–59. [Google Scholar] [CrossRef]
- Usup, G.; Kulis, D.M.; Anderson, D.M. Growth and toxin production of the toxic dinoflagellate Pyrodinium bahamense var. compressum in laboratory cultures. Nat. Toxins 1994, 2, 254–262. [Google Scholar] [CrossRef]
- Oshima, Y.; Blackburn, S.I.; Hallegraeff, G.M. Comparative study on paralytic shellfish toxin profiles of the dinoflagellate Gymnodinium catenatum from three different countries. Mar. Boil. 1993, 116, 471–476. [Google Scholar] [CrossRef]
- Anderson, D.M.; Alpermann, T.J.; Cembella, A.D.; Collos, Y.; Masseret, E.; Montresor, M. The globally distributed genus Alexandrium: Multifaceted roles in marine ecosystems and impacts on human health. Harmful Algae 2012, 14, 10–35. [Google Scholar] [CrossRef]
- John, U.; Litaker, R.W.; Montresor, M.; Murray, S.; Brosnahan, M.L.; Anderson, D.M. Formal revision of the Alexandrium tamarense species complex (Dinophyceae) taxonomy: the introduction of five species with emphasis on molecular-based (rDNA) classification. Protist 2014, 165, 779–804. [Google Scholar] [CrossRef]
- Shimizu, Y.; Norte, M.; Hori, A.; Genenah, A.; Kobayashi, M. Biosynthesis of saxitoxin analogs: the unexpected pathway. J. Am. Chem. Soc. 1984, 106, 6433–6434. [Google Scholar] [CrossRef]
- Mihali, T.K.; Carmichael, W.W.; Neilan, B.A. A Putative Gene Cluster from a Lyngbya wollei Bloom that Encodes Paralytic Shellfish Toxin Biosynthesis. PLoS ONE 2011, 6, e14657. [Google Scholar] [CrossRef]
- Mihali, T.K.; Kellmann, R.; Neilan, B.A. Characterisation of the paralytic shellfish toxin biosynthesis gene clusters in Anabaena circinalis AWQC131C and Aphanizomenon sp. NH-5. BMC Biochem. 2009, 10, 8. [Google Scholar] [CrossRef]
- Kellmann, R.; Mihali, T.K.; Jeon, Y.J.; Pickford, R.; Pomati, F.; Neilan, B.A. Biosynthetic Intermediate Analysis and Functional Homology Reveal a Saxitoxin Gene Cluster in Cyanobacteria. Appl. Environ. Microbiol. 2008, 74, 4044–4053. [Google Scholar] [CrossRef] [Green Version]
- Stucken, K.; John, U.; Cembella, A.; Murillo, A.A.; Soto-Liebe, K.; Fuentes-Valdés, J.J.; Friedel, M.; Plominsky, A.M.; Vásquez, M.; Glöckner, G. The Smallest Known Genomes of Multicellular and Toxic Cyanobacteria: Comparison, Minimal Gene Sets for Linked Traits and the Evolutionary Implications. PLoS ONE 2010, 5, e9235. [Google Scholar] [CrossRef]
- Cullen, A.; D’Agostino, P.M.; Mazmouz, R.; Pickford, R.; Wood, S.; Neilan, B.A. Insertions within the saxitoxin biosynthetic gene cluster result in differential toxin profiles. ACS Chem. Boil. 2018, 13, 3107–3114. [Google Scholar] [CrossRef]
- Tsuchiya, S.; Cho, Y.; Konoki, K.; Nagasawa, K.; Oshima, Y.; Yotsu-Yamashita, M. Synthesis and identification of proposed biosynthetic intermediates of saxitoxin in the cyanobacterium Anabaena circinalis (TA04) and the dinoflagellate Alexandrium tamarense (Axat-2). Org. Biomol. Chem. 2014, 12, 3016–3020. [Google Scholar] [CrossRef]
- Murray, S.A.; Mihali, T.K.; Neilan, B.A. Extraordinary Conservation, Gene Loss, and Positive Selection in the Evolution of an Ancient Neurotoxin. Mol. Boil. Evol. 2011, 28, 1173–1182. [Google Scholar] [CrossRef]
- Murray, S.A.; Diwan, R.; Orr, R.J.S.; Kohli, G.S.; John, U. Gene duplication, loss and selection in the evolution of saxitoxin biosynthesis in alveolates. Mol. Phylogenetics Evol. 2015, 92, 165–180. [Google Scholar] [CrossRef] [Green Version]
- Orr, R.J.S.; Stüken, A.; Murray, S.A.; Jakobsen, K.S. Evolutionary Acquisition and Loss of Saxitoxin Biosynthesis in Dinoflagellates: the Second “Core” Gene, sxtG. Appl. Environ. Microbiol. 2013, 79, 2128–2136. [Google Scholar] [CrossRef]
- Hackett, J.D.; Wisecaver, J.H.; Brosnahan, M.L.; Kulis, D.M.; Anderson, D.M.; Bhattacharya, D.; Plumley, F.G.; Erdner, D.L. Evolution of Saxitoxin Synthesis in Cyanobacteria and Dinoflagellates. Mol. Boil. Evol. 2012, 30, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, S.-F.; Lin, L.; Wang, D.-Z. Comparative Transcriptome Analysis of a Toxin-Producing Dinoflagellate Alexandrium catenella and Its Non-Toxic Mutant. Mar. Drugs 2014, 12, 5698–5718. [Google Scholar] [CrossRef]
- Li, Z.; Mertens, K.N.; Nézan, E.; Chomérat, N.; Bilien, G.; Iwataki, M.; Shin, H.H. Discovery of a New Clade Nested Within the Genus Alexandrium (Dinophyceae): Morpho-molecular Characterization of Centrodinium punctatum (Cleve) F.J.R. Taylor. Protist 2019, 170, 168–186. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, S.-F.; Lin, L.; Wang, D.-Z. Whole Transcriptomic Analysis Provides Insights into Molecular Mechanisms for Toxin Biosynthesis in a Toxic Dinoflagellate Alexandrium catenella (ACHK-T). Toxins 2017, 9, 213. [Google Scholar] [CrossRef]
- Orr, R.J.S.; Stüken, A.; Rundberget, T.; Eikrem, W.; Jakobsen, K.S. Improved phylogenetic resolution of toxic and non-toxic Alexandrium strains using a concatenated rDNA approach. Harmful Algae 2011, 10, 676–688. [Google Scholar] [CrossRef]
- Mendoza-Flores, A.; Leyva-Valencia, I.; Band-Schmidt, C.J.; Galindo-Sánchez, C.E.; Bustillos-Guzman, J.J. Identification of the gene sxtA (domains sxtA1 and sxtA4) in Mexican strains of Gymnodinium catenatum (Dinophyceae) and their evolution. Front. Mar. Sci. 2018, 5, 289. [Google Scholar] [CrossRef]
- McEwan, M.; Humanyun, R.; Slamovtis, C.H.; Keeling, P.J. Nuclear Genome Sequence Survey of the Dinoflagellate Heterocapsa Triquetra. J. Eukaryot. Microbiol. 2008, 55, 530–535. [Google Scholar] [CrossRef]
- Bertomeu, T.; Morse, D. Isolation of a dinoflagellate mitotic cyclin by functional complementation in yeast. Biochem. Biophys. Res. Commun. 2004, 323, 1172–1183. [Google Scholar] [CrossRef]
- Liu, L.; Hastings, J.W. Novel and rapidly diverging intergenic sequences between tandem repeats of the luciferase genes in seven dinoflagelalte species. J. Phycol. 2006, 42, 96–103. [Google Scholar] [CrossRef]
- Song, B.; Chen, S.; Chen, W. Dinoflagellates, a Unique Lineage for Retrogene Research. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Johnson, J.G.; Morey, J.S.; Neely, M.G.; Ryan, J.C.; Van Dolah, F.M. Transcriptome remodeling associated with chronological aging in the dinoflagellate, Karenia Brevis. Mar. Genom. 2012, 5, 15–25. [Google Scholar] [CrossRef]
- Perry, G.H.; Dominy, N.J.; Claw, K.G.; Lee, A.S.; Fiegler, H.; Redon, R.; Werner, J.; Villanea, F.A.; Mountain, J.L.; Misra, R.; et al. Diet and the evolution of human amylase gene copy number variation. Nat. Genet. 2007, 39, 1256. [Google Scholar] [CrossRef]
- Conrad, D.F.; Pinto, D.; Redon, R.; Feuk, L.; Gokcumen, O.; Zhang, Y.; Aerts, J.; Andrews, T.D.; Barnes, C.; Campbell, P.; et al. Origins and functional impact of copy number variation in the human genome. Nature 2009, 464, 704. [Google Scholar] [CrossRef]
- Stüken, A.; Riobó, P.; Franco, J.; Jakobsen, K.S.; Guillou, L.; Figueroa, R.I. Paralytic shellfish toxin content is related to genomic sxtA4 copy number in Alexandrium minutum strains. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef]
- Hii, K.S.; Lim, P.T.; Kon, N.F.; Takata, Y.; Usup, G.; Leaw, C.P. Physiological and transcriptional responses to inorganic nutrition in a tropical Pacific strain of Alexandrium minutum: Implications for the saxitoxin genes and toxin production. Harmful Algae 2016, 56, 9–21. [Google Scholar] [CrossRef]
- Perini, F.; Galluzzi, L.; Dell’Aversano, C.; Iacovo, E.D.; Tartaglione, L.; Ricci, F.; Forino, M.; Ciminiello, P.; Penna, A. SxtA and sxtG Gene Expression and Toxin Production in the Mediterranean Alexandrium minutum (Dinophyceae). Mar. Drugs 2014, 12, 5258–5276. [Google Scholar] [CrossRef]
- Wiese, M.; Murray, S.A.; Alvin, A.; Neilan, B.A. Gene expression and molecular evolution of sxtA4 in a saxitoxin producing dinoflagellate Alexandrium Catenella. Toxicon 2014, 92, 102–112. [Google Scholar] [CrossRef]
- Wohlrab, S.; Selander, E.; John, U. Predator cues reduce intraspecific trait variability in a marine dinoflagellate. BMC Ecol. 2017, 17, 8. [Google Scholar] [CrossRef]
- Wang, D.-Z.; Li, C.; Zhang, Y.; Wang, Y.-Y.; He, Z.-P.; Lin, L.; Hong, H.-S. Quantitative proteomic analysis of differentially expressed proteins in the toxicity-lost mutant of Alexandrium catenella (Dinophyceae) in the exponential phase. J. Proteom. 2012, 75, 5564–5577. [Google Scholar] [CrossRef]
- Zhang, S.-F.; Zhang, Y.; Lin, L.; Wang, D.-Z. iTRAQ-Based Quantitative Proteomic Analysis of a Toxigenic Dinoflagellate Alexandrium catenella at Different Stages of Toxin Biosynthesis during the Cell Cycle. Mar. Drugs 2018, 16, 491. [Google Scholar] [CrossRef]
- Shimizu, Y. Toxigenesis and biosynthesis of saxitoxin analogues. In Pure and Applied Chemistry; Walter de Gruyter GmbH: Berlin, Germany, 1986; Volume 58, p. 257. [Google Scholar]
- Taroncher-Oldenburg, G.; Anderson, D.M. Identification and Characterization of Three Differentially Expressed Genes, Encoding S-Adenosylhomocysteine Hydrolase, Methionine Aminopeptidase, and a Histone-Like Protein, in the Toxic Dinoflagellate Alexandrium Fundyense. Appl. Environ. Microbiol. 2000, 66, 2105–2112. [Google Scholar] [CrossRef]
- Murray, S.A.; Wiese, M.; Stüken, A.; Brett, S.; Kellmann, R.; Hallegraeff, G.; Neilan, B.A. sxtA-Based Quantitative Molecular Assay To Identify Saxitoxin-Producing Harmful Algal Blooms in Marine Waters. Appl. Environ. Microbiol. 2011, 77, 7050–7057. [Google Scholar] [CrossRef]
- Penna, A.; Perini, F.; Dell’Aversano, C.; Capellacci, S.; Tartaglione, L.; Giacobbe, M.G.; Casabianca, S.; Fraga, S.; Ciminiello, P.; Scardi, M. The sxt Gene and Paralytic Shellfish Poisoning Toxins as Markers for the Monitoring of Toxic Alexandrium Species Blooms. Environ. Sci. Technol. 2015, 49, 14230–14238. [Google Scholar] [CrossRef]
- Erdner, D.L.; Percy, L.; Keafer, B.; Lewis, J.; Anderson, D.M. A quantitative real-time PCR assay for the identification and enumeration of Alexandrium cysts in marine sediments. Deep. Sea Res. Part II: Top. Stud. Oceanogr. 2010, 57, 279–287. [Google Scholar] [CrossRef]
- Galluzzi, L.; Penna, A.; Bertozzini, E.; Vila, M.; Garcés, E.; Magnani, M. Development of a Real-Time PCR Assay for Rapid Detection and Quantification of Alexandrium minutum (a Dinoflagellate). Appl. Environ. Microbiol. 2004, 70, 1199–1206. [Google Scholar] [CrossRef]
- Hosoi-Tanabe, S.; Sako, Y. Species-Specific Detection and Quantification of Toxic Marine Dinoflagellates Alexandrium tamarense and A. catenella byReal-Time PCR Assay. Mar. Biotechnol. 2005, 7, 506–514. [Google Scholar] [CrossRef]
- Kamikawa, R.; Nagai, S.; Hosoi-Tanabe, S.; Itakura, S.; Yamaguchi, M.; Uchida, Y.; Baba, T.; Sako, Y. Application of real-time PCR assay for detection and quantification of Alexandrium tamarense and Alexandrium catenella cysts from marine sediments. Harmful Algae 2007, 6, 413–420. [Google Scholar] [CrossRef]
- Kon, N.F.; Teng, S.T.; Hii, K.S.; Yek, L.H.; Mujahid, A.; Lim, H.C.; Lim, P.T.; Leaw, C.P. Spatial distribution of toxic Alexandrium tamiyavanichii (Dinophyceae) in the southeastern South China Sea-Sulu Sea: A molecular-based assessment using real-time quantitative PCR (qPCR) assay. Harmful Algae 2015, 50, 8–20. [Google Scholar] [CrossRef]
- Ruvindy, R.; Bolch, C.J.; MacKenzie, L.; Smith, K.F.; Murray, S.A. qPCR Assays for the Detection and Quantification of Multiple Paralytic Shellfish Toxin-Producing Species of Alexandrium. Front. Microbiol. 2018, 9, 3153. [Google Scholar] [CrossRef]
- Galluzzi, L.; Bertozzini, E.; Penna, A.; Perini, F.; Garcés, E.; Magnani, M. Analysis of rRNA gene content in the Mediterranean dinoflagellate Alexandrium catenella and Alexandrium taylori: implications for the quantitative real-time PCR-based monitoring methods. J. Appl. Phycol. 2010, 22, 1–9. [Google Scholar] [CrossRef]
- Godhe, A.; Cusack, C.; Pedersen, J.; Andersen, P.; Anderson, D.M.; Bresnan, E.; Cembella, A.; Dahl, E.; Diercks, S.; Elbrächter, M.; et al. Intercalibration of classical and molecular techniques for identification of Alexandrium fundyense (Dinophyceae) and estimation of cell densities. Harmful Algae 2007, 6, 56–72. [Google Scholar] [CrossRef]
- Savela, H.; Harju, K.; Spoof, L.; Lindehoff, E.; Meriluoto, J.; Vehniäinen, M.; Kremp, A. Quantity of the dinoflagellate sxtA4 gene and cell density correlates with paralytic shellfish toxin production in Alexandrium ostenfeldii blooms. Harmful Algae 2016, 52, 1–10. [Google Scholar] [CrossRef]
- Stüken, A.; Dittami, S.M.; Eikrem, W.; McNamee, S.; Campbell, K.; Jakobsen, K.S.; Edvardsen, B. Novel hydrolysis-probe based qPCR assay to detect saxitoxin transcripts of dinoflagellates in environmental samples. Harmful Algae 2013, 28, 108–117. [Google Scholar] [CrossRef]
- Gao, Y.; Yu, R.-C.; Chen, J.-H.; Zhang, Q.-C.; Kong, F.-Z.; Zhou, M.-J. Distribution of Alexandrium fundyense and A. pacificum (Dinophyceae) in the Yellow Sea and Bohai Sea. Mar. Pollut. Bull. 2015, 96, 210–219. [Google Scholar] [CrossRef]
- Farrell, H.; O’Connor, W.; Seebacher, F.; Harwood, D.T.; Murray, S. Molecular Detection of the sxtA Gene from Saxitoxin-Producing Alexandrium minutum in Commercial Oysters. J. Shellfish. Res. 2016, 35, 169–177. [Google Scholar] [CrossRef]
- Zhuang, Y.; Zhang, H.; Hannick, L.; Lin, S. Metatranscriptome profiling reveals versatile N-nutrient utilization, CO2 limitation, oxidative stress, and active toxin production in an Alexandrium fundyense bloom. Harmful Algae 2015, 42, 60–70. [Google Scholar] [CrossRef]
- Firn, R.D.; Jones, C.G. Natural products–a simple model to explain chemical diversity. Nat. Prod. Rep. 2003, 20, 382–391. [Google Scholar] [CrossRef]
- Jenke-Kodama, H.; Müller, R.; Dittmann, E. Evolutionary mechanisms underlying secondary metabolite diversity. In Natural Compounds as Drugs Volume I; Birkhäuser: Basel, Switzerland, 2008; pp. 119–140. [Google Scholar]
- Weng, J.-K.; Philippe, R.N.; Noel, J.P. The rise of chemodiversity in plants. Science 2012, 336, 1667–1670. [Google Scholar] [CrossRef]
Figure 1.
Functional role of toxins produced by dinoflagellates. Schematic representing various biological roles played by toxins for their producing organisms. Photo credit: Diana Kleine, Dieter Tracey, Ian Hewson, Jane Hawkey, Jane Thomas and Tracey Saxby, Integration and Application Network, University of Maryland Center for Environmental Science (ian.umces.edu/imagelibrary/).
Figure 1.
Functional role of toxins produced by dinoflagellates. Schematic representing various biological roles played by toxins for their producing organisms. Photo credit: Diana Kleine, Dieter Tracey, Ian Hewson, Jane Hawkey, Jane Thomas and Tracey Saxby, Integration and Application Network, University of Maryland Center for Environmental Science (ian.umces.edu/imagelibrary/).

Figure 2.
Polyketide biosynthesis. Schematic representing the putative biosynthesis pathway of polyketide compound Amphidinol-9 by various modular PKS domains represented by ACP: acyl carrier protein, AT: acyl transferase, DH: dehydratase, KR: ketoreductase, and KS: ketosynthase. Figure credit: Gurjeet Singh Kohli.
Figure 2.
Polyketide biosynthesis. Schematic representing the putative biosynthesis pathway of polyketide compound Amphidinol-9 by various modular PKS domains represented by ACP: acyl carrier protein, AT: acyl transferase, DH: dehydratase, KR: ketoreductase, and KS: ketosynthase. Figure credit: Gurjeet Singh Kohli.

Figure 3.
Ketoacyl synthase (KS) domain phylogeny in dinoflagellates (modified from [81,122,129]). Phylogenetic tree representing the three sub-clades of KS domains within the dinoflagellate type I PKSs, multi-domain PKSs that cluster with other apicomplexans (condensed clade represented by 1), chlorophytes, haptophytes (condensed clade represented by 2) and the burA like PKS-NRPSs reported from dinoflagellates along with schematic representations of PKS domains represented by AT: acyl transferase, A: Non-Ribosomal Peptide Synthase, KS: ketosynthase, DH: dehydratase, KR: ketoreductase, TE: thioesterase, ACP: acyl carrier protein, ER: enoyl reductase, SL: 5’ splice leader, polyA: poly A tail. Other PKS groups are represented by 3: Modular type PKS cis AT clade, 4: fungal PKS non-reducing clade, 5: Animal type I FAS, 6: fugal PKS reducing; and 7: Type II PKSs.
Figure 3.
Ketoacyl synthase (KS) domain phylogeny in dinoflagellates (modified from [81,122,129]). Phylogenetic tree representing the three sub-clades of KS domains within the dinoflagellate type I PKSs, multi-domain PKSs that cluster with other apicomplexans (condensed clade represented by 1), chlorophytes, haptophytes (condensed clade represented by 2) and the burA like PKS-NRPSs reported from dinoflagellates along with schematic representations of PKS domains represented by AT: acyl transferase, A: Non-Ribosomal Peptide Synthase, KS: ketosynthase, DH: dehydratase, KR: ketoreductase, TE: thioesterase, ACP: acyl carrier protein, ER: enoyl reductase, SL: 5’ splice leader, polyA: poly A tail. Other PKS groups are represented by 3: Modular type PKS cis AT clade, 4: fungal PKS non-reducing clade, 5: Animal type I FAS, 6: fugal PKS reducing; and 7: Type II PKSs.

Figure 4.
Survey of ketosynthase (KS) domains from various marine microbial eukaryotic lineages (modified from [81]). The figure represents the large number of KS domains that have been reported from the analysis of MMETSP libraries of different dinoflagellate species in comparison with species of cryptophytes, haptophytes, viriplantae, ciliates, diatoms, and other strameopiles. Photo credit: Diana Kleine, Jane Thomas and Tracey Saxby, Integration and Application Network, University of Maryland Center for Environmental Science (ian.umces.edu/imagelibrary/).
Figure 4.
Survey of ketosynthase (KS) domains from various marine microbial eukaryotic lineages (modified from [81]). The figure represents the large number of KS domains that have been reported from the analysis of MMETSP libraries of different dinoflagellate species in comparison with species of cryptophytes, haptophytes, viriplantae, ciliates, diatoms, and other strameopiles. Photo credit: Diana Kleine, Jane Thomas and Tracey Saxby, Integration and Application Network, University of Maryland Center for Environmental Science (ian.umces.edu/imagelibrary/).

Figure 5.
Revised biosynthetic pathway of PSTs in dinoflagellates (modified from [22,184,185,193,195]). The reaction steps are as follows; 1: Claisen condensation; 2: amidino transfer; 3: heterocyclization; 4: desaturation (Double bond formation); 5: epoxidation of the new double bond; 6: aldehyde formation; 7: terminal aldehyde reduction; 8: dihydroxylation and 9: carbamoylation.
Figure 5.
Revised biosynthetic pathway of PSTs in dinoflagellates (modified from [22,184,185,193,195]). The reaction steps are as follows; 1: Claisen condensation; 2: amidino transfer; 3: heterocyclization; 4: desaturation (Double bond formation); 5: epoxidation of the new double bond; 6: aldehyde formation; 7: terminal aldehyde reduction; 8: dihydroxylation and 9: carbamoylation.

Figure 6.
The structure of sxtA and sxtG in dinoflagellates (modified from [57,191]). A. Transcript structure of sxtA short isoform. B. Transcript structure of sxtA long isoform. C. Transcript structure of sxtG. D. Genomic structure of sxtG.


{kind=link}
{kind=link}
{kind=link}
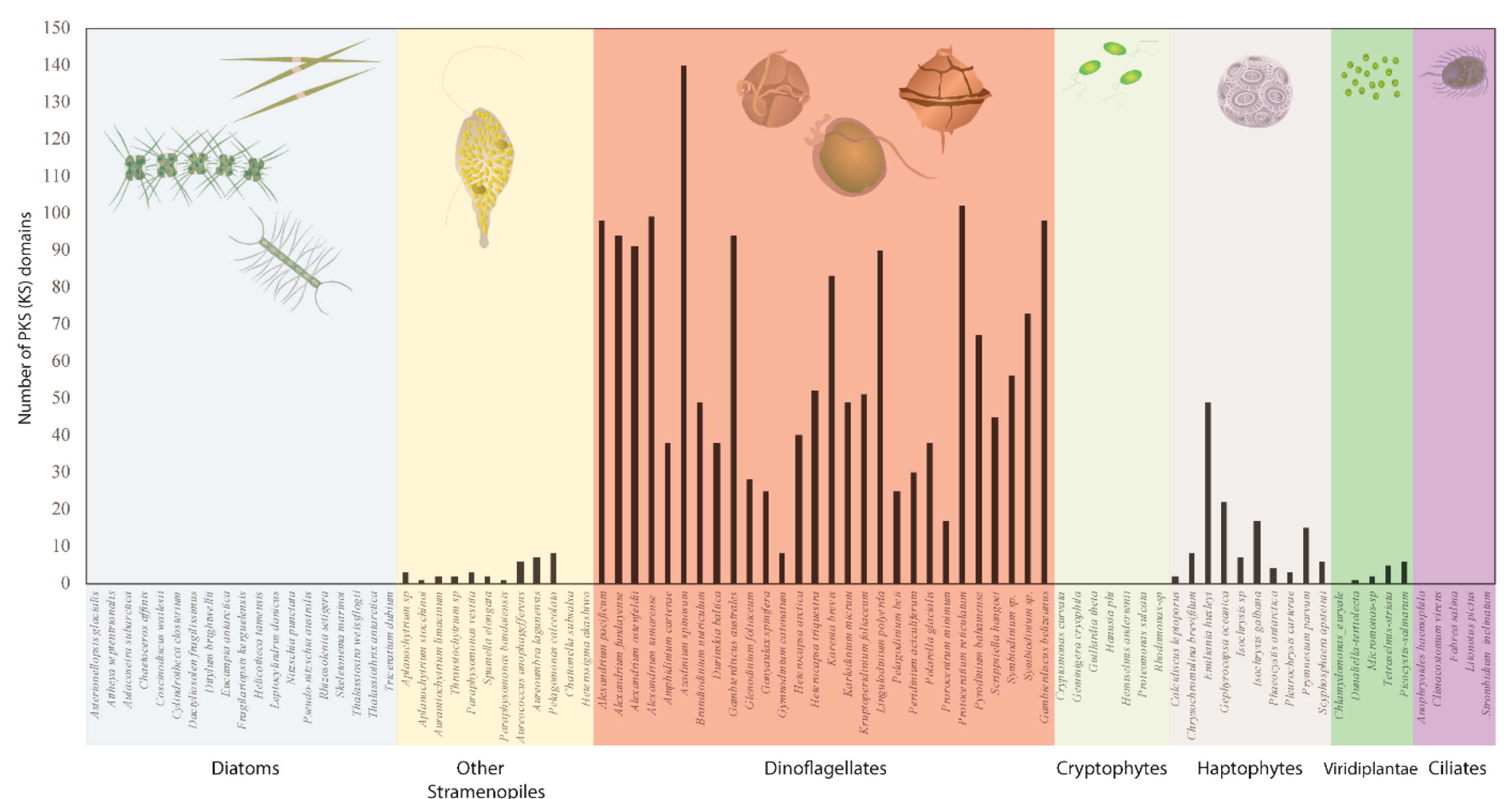{kind=link}
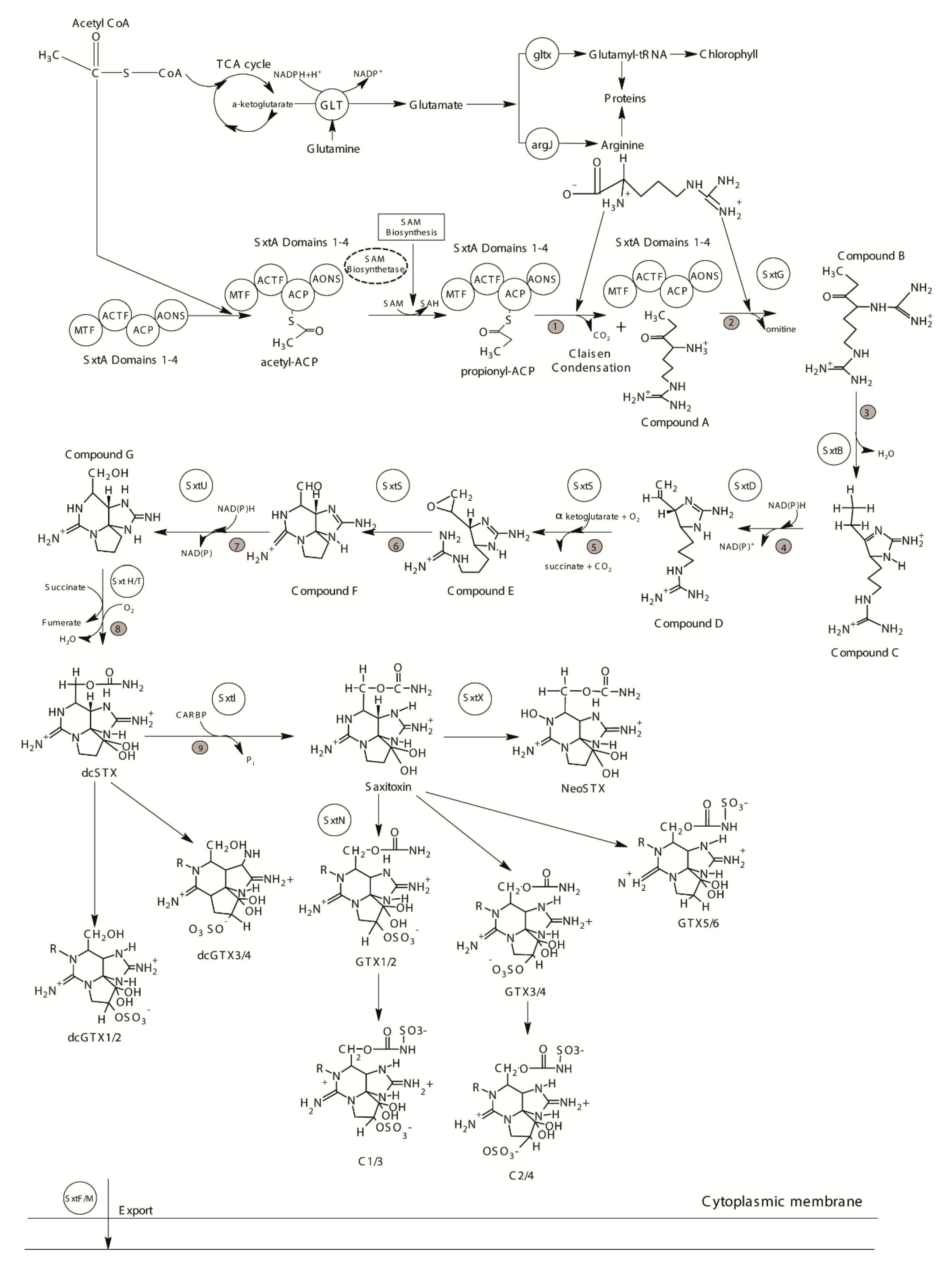{kind=link}
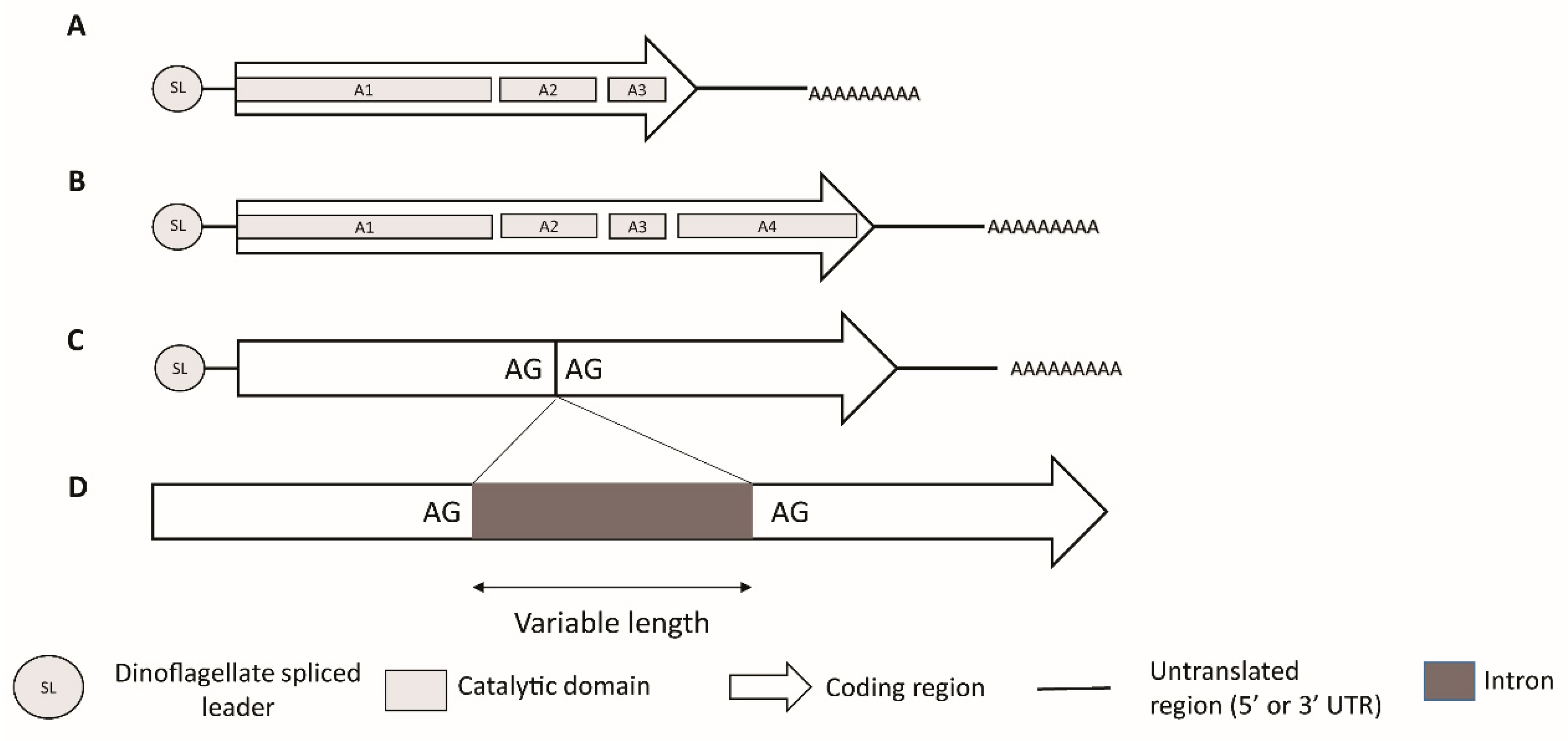{kind=link}
Table 1.
Putative sxt genes in the dinoflagellates.
| Gene | Function * | Dinoflagellate Species | Reported from Transcriptome | Reported from Genome |
|---|---|---|---|---|
| Core sxt Genes | ||||
| sxtA (sxtA4) | Aspartate aminotransferase | Alexandrium australiense Alexandrium catenella Alexandrium minutum Alexandrium ostenfeldii Alexandrium pacificum Alexandrium tamarense Gymnodinium catenatum Pyrodinium bahamense | na Yes [57,192] Yes [57] na Yes [190,192,193,195] No [181,192] Yes [190] Yes [190] | Yes [57,190] Yes [57,181,190] Yes [57,190] Yes [177] Yes [57,190] Yes [57]; No [181,190] Yes [57,197] na |
| sxtG | Amidinotransferase | Alexandrium affine Alexandrium andersoni Alexandrium australiense Alexandrium catenella Alexandrium insuetum Alexandrium minutum Alexandrium pacificum Gymnodinium catenatum Pyrodinium bahamense | No [191] No [191] Yes [191] Yes [57] No [191] Yes [57] Yes [191,192,193,195] Yes [190,192] Yes [190,192] | Yes [191] Yes [191] Yes [191] Yes [191] Yes [191] Yes [191] Yes [191] Yes [191] na |
| sxtB | Cytidine deaminase | Alexandrium catenella Alexandrium minutum Alexandrium pacificum | Yes [57] Yes [57] Yes [192,193,195] | na na na |
| sxtD | Sterole desaturase | Alexandrium pacificum | Yes [192,193,195] | na |
| sxtS | Phytanoyl-CoA dioxygenase | Alexandrium catenella Alexandrium minutum Alexandrium pacificum Gymnodinium catenatum Pyrodinium bahamense | No [57]; Yes [192] Yes [57] Yes [192] Yes [192] Yes [192] | na na na na na |
| sxtU | Alcohol dehydrogenase | Alexandrium catenella Alexandrium minutum Alexandrium pacificum Pyrodinium bahamense | Yes [57,192] Yes [57] Yes [192,193,195] Yes [192] | na na na na |
| sxtH/T | Phenylpropionate dioxygenase | Alexandrium catenella Alexandrium minutum Alexandrium pacificum | Yes [57,192] Yes [57] Yes [192,193]; Yes as H/T/DIOX [195] | na na na |
| sxtI | O-carbamoyl transferase | Alexandrium catenella Alexandrium minutum Alexandrium pacificum Pyrodinium bahamense | Yes [57] Yes [57] Yes [192,193,195] Yes [192] | na na na na |
| sxtF/M | Multidrug efflux protein | Alexandrium catenella Alexandrium minutum Alexandrium pacificum | Yes [57] Yes [57] Yes [192,193,195] | na na na |
| sxtP | STX binding protein | Alexandrium catenella Alexandrium pacificum | Yes [192] Yes [192,195] | na na |
| Other sxt Genes | ||||
| sxtL | GDSL lipase | Alexandrium pacificum | Yes [192] | na |
| sxtN | Sulfotransferase | Alexandrium pacificum | Yes [192] | na |
| sxtO | Adenylylsulfate kinase | Alexandrium pacificum | Yes [193,195] | na |
| sxtR | Acetyl CoA N-acyltransferase | Alexandrium catenella Alexandrium minutum | Yes [57] Yes [57] | na na |
| sxtW | Ferredoxin | Alexandrium pacificum | Yes [195] | na |
| sxtX | Cephalosporine hydroxylase | Alexandrium pacificum Pyrodinium bahamense | Yes [192,193,195] Yes [192] | na na |
| sxtZ | Histidine kinase | Alexandrium pacificum | Yes [193,195] | na |
| sxtPER | Permease | Alexandrium pacificum | Yes [195] | na |
| sxtACT | Acetylase | Alexandrium pacificum | Yes [195] | na |
* Function inferred according to [185]. na: not available.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Verma, A.; Barua, A.; Ruvindy, R.; Savela, H.; Ajani, P.A.; Murray, S.A. The Genetic Basis of Toxin Biosynthesis in Dinoflagellates. Microorganisms 2019, 7, 222. https://doi.org/10.3390/microorganisms7080222
AMA Style
Verma A, Barua A, Ruvindy R, Savela H, Ajani PA, Murray SA. The Genetic Basis of Toxin Biosynthesis in Dinoflagellates. Microorganisms. 2019; 7(8):222. https://doi.org/10.3390/microorganisms7080222
Chicago/Turabian StyleVerma, Arjun, Abanti Barua, Rendy Ruvindy, Henna Savela, Penelope A. Ajani, and Shauna A. Murray. 2019. "The Genetic Basis of Toxin Biosynthesis in Dinoflagellates" Microorganisms 7, no. 8: 222. https://doi.org/10.3390/microorganisms7080222
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.