
The Isolation of Anaerobic and Facultative Anaerobic Sulfate-Reducing Bacteria (SRB) and a Comparison of Related Enzymes in Their Sulfate Reduction Pathways
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Preparation of Bacterial Suspension of the Samples from the Rust Layer and Sediment
2.3. Separation, Purification, and Cryopreservation of Anaerobic SRB
2.4. Preliminary Identification of Strains
2.5. Screening of Facultative Anaerobic SRB
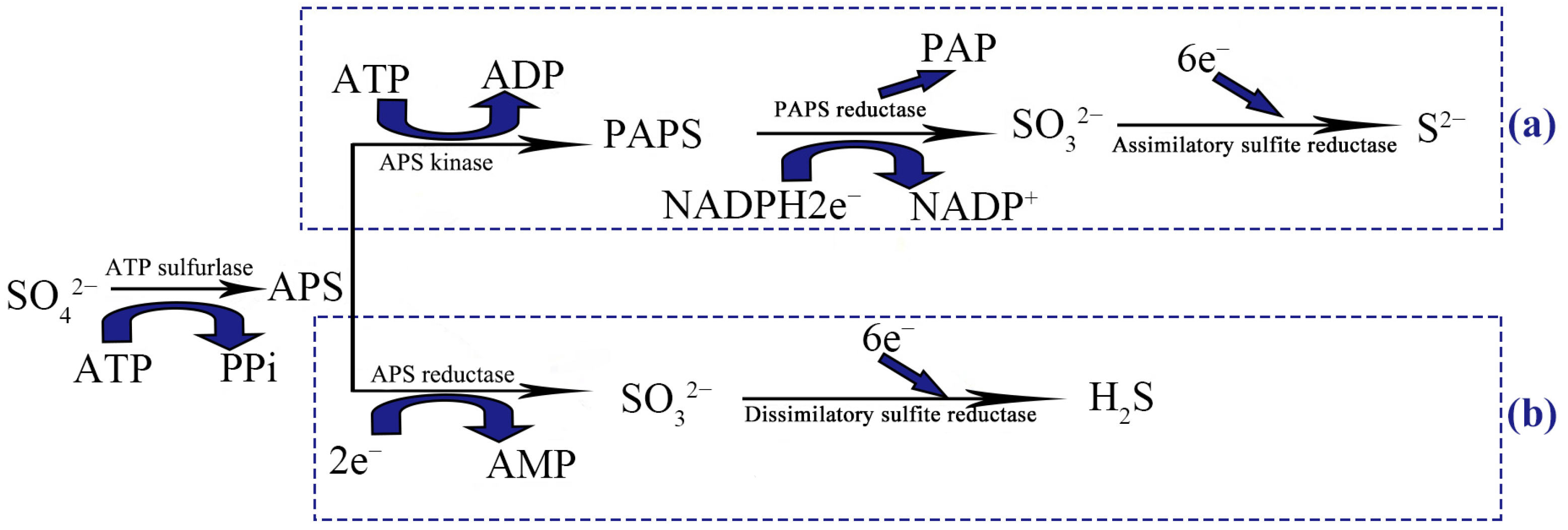
2.6. Distribution of Related Enzymes in the Sulfate Reduction Pathway
3. Results
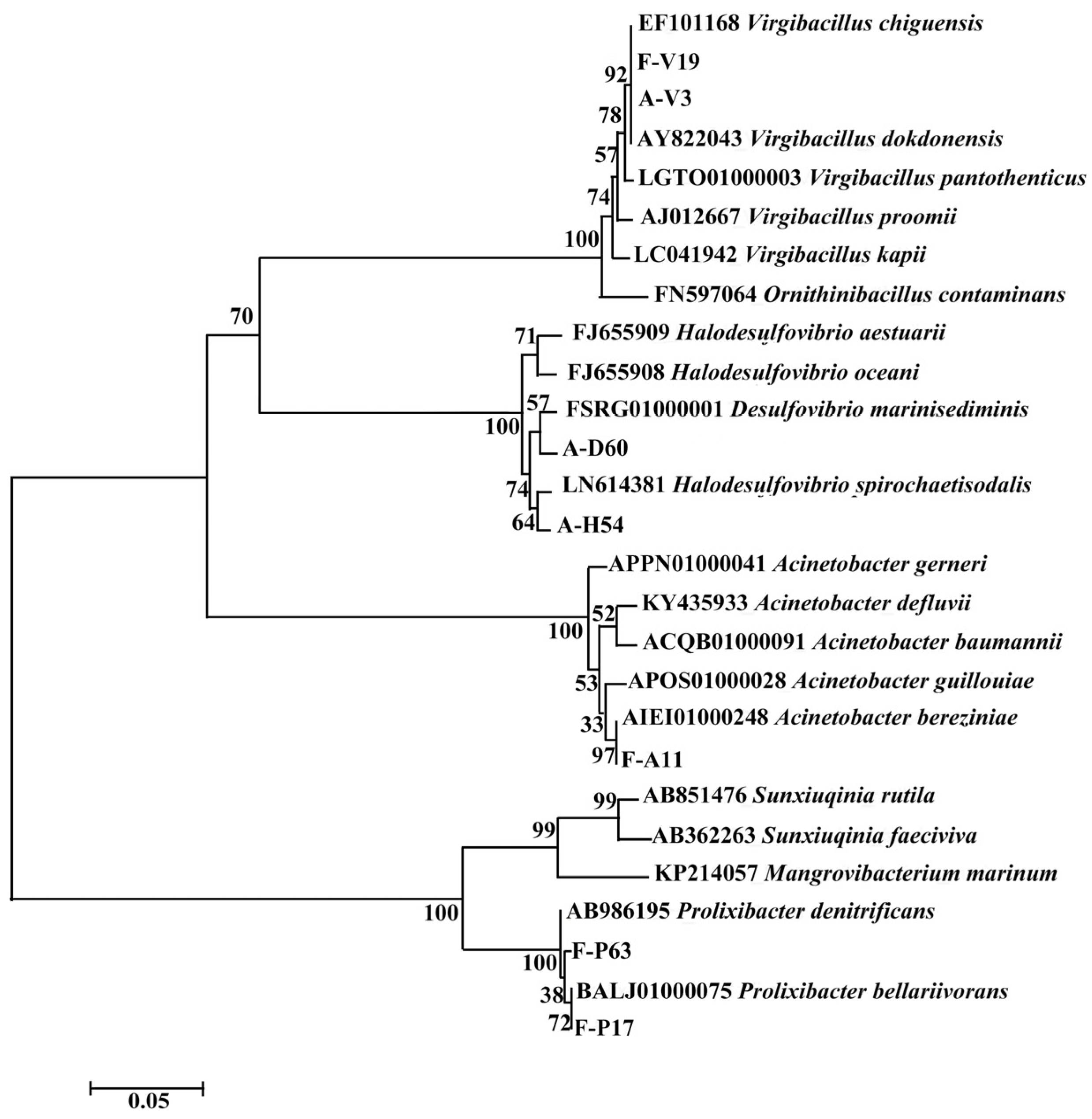
3.1. Identification of Isolated Bacteria
3.2. Screening of Facultative Anaerobic SRB
3.3. Isozyme Clustering Results
4. Discussion
4.1. Composition of Culturable SRB in Rust Layers and Sediments
4.2. Screening of Facultative Anaerobic Culturable SRB
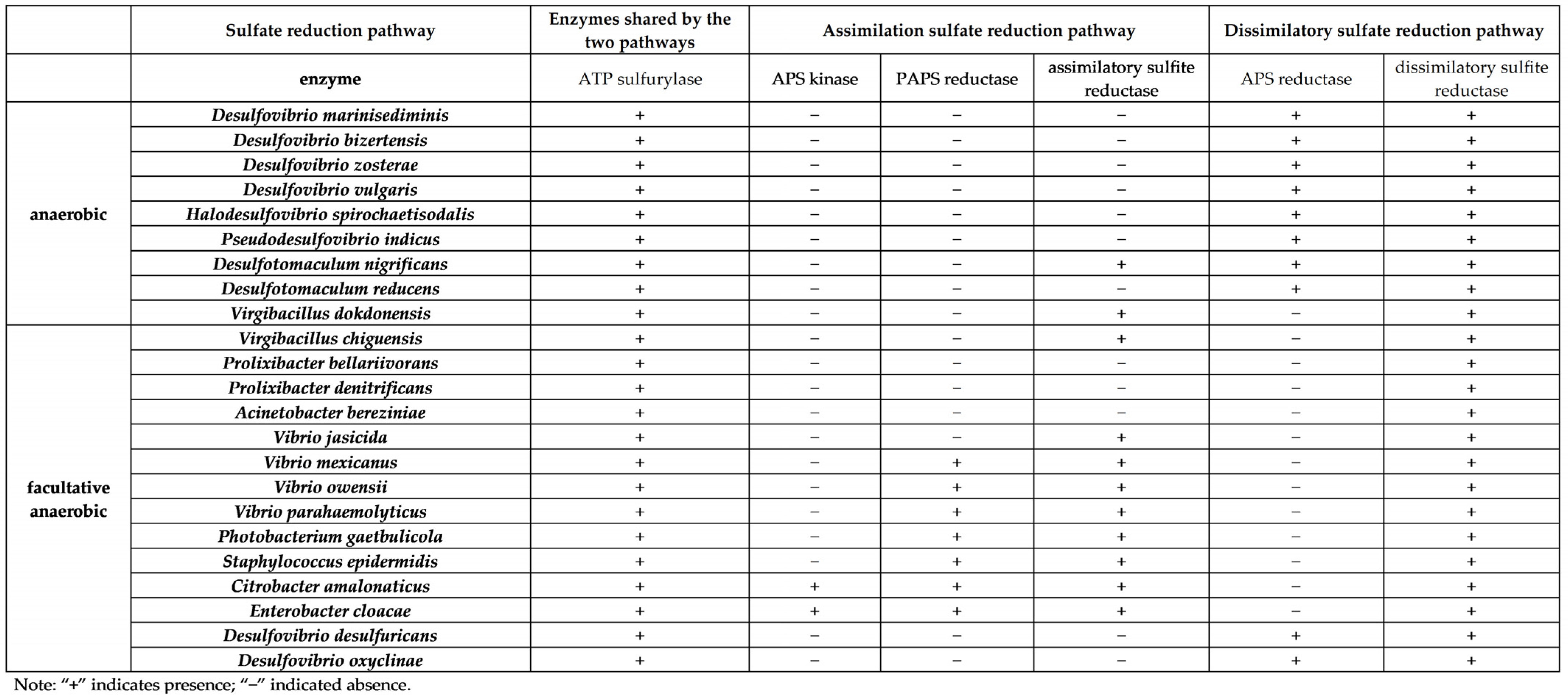
4.3. Distribution of Enzymes Involved in the Sulfate Reduction Pathway of Culturable SRB
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Barton, L.L.; Fauque, G.D. Biochemistry, physiology and biotechnology of sulfate-reducing bacteria. Adv. Appl. Microbiol. 2009, 68, 41–98. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, J.; Gao, Z.; Di, J.; Wang, D.; Guo, X.; Hu, Z.; Gao, X.; Wang, Y. Study on growth influencing factors and desulfurization performance of sulfate reducing bacteria based on the response surface methodology. ACS Omega 2023, 8, 4046–4059. [Google Scholar] [CrossRef]
- Gibson, G.R. Physiology and ecology of the sulphate-reducing bacteria. J. Appl. Microbiol. 1990, 69, 767–797. [Google Scholar] [CrossRef]
- Angell, P.; Urbanic, K. Sulphate-reducing bacterial activity as a parameter to predict localized corrosion of stainless alloys. Corros. Sci. 2000, 42, 897–912. [Google Scholar] [CrossRef]
- Hirano, S.I.; Ihara, S.; Wakai, S.; Dotsuta, Y.; Otani, K.; Kitagaki, T.; Ueno, F.; Okamoto, A. Novel Methanobacterium strain induces severe corrosion by retrieving electrons from Fe0 under a freshwater environment. Microorganisms 2022, 10, 270. [Google Scholar] [CrossRef]
- Martins, M.; Pereira, I.A.C. Sulfate-reducing bacteria as new microorganisms for biological hydrogen production. Int. J. Hydrogen Energy 2013, 38, 12294–12301. [Google Scholar] [CrossRef]
- Ma, X.; Jia, X.; Zhao, Y. Research and application of the processes of disposal of wastewater containing heavy metals by sulfate reducing bacteria. J. Microbiol. 2003, 23, 36–39. [Google Scholar] [CrossRef]
- Dong, Y.; Di, J.; Hu, H.; Zhou, J.; Ruan, Z.; Sun, X.; Zhao, Z. Experimental study in isolation of sulfate reducing bacteria and treatment of acid mine drainage process. J. Water Resour. Eng. 2019, 30, 25–30. [Google Scholar]
- Herman, J.P.; Redfern, L.; Teaf, C.; Covert, D.; Michael, P.R.; Missimer, T.M. Cumene contamination in groundwater: Observed concentrations, evaluation of remediation by sulfate enhanced bioremediation (SEB), and public health issues. Int. J. Environ. Res. Public Health 2020, 17, 8380. [Google Scholar] [CrossRef]
- Tripathi, A.K.; Saxena, P.; Thakur, P.; Rauniyar, S.; Samanta, D.; Gopalakrishnan, V.; Singh, R.N.; Sani, R.K. Transcriptomics and functional analysis of copper stress response in the sulfate-reducing bacterium Desulfovibrio alaskensis G20. Int. J. Mol. Sci. 2022, 23, 1396. [Google Scholar] [CrossRef]
- van den Brand, T.P.H.; Roest, K.; Chen, G.H.; Brdjanovic, D.; van Loosdrecht, M.C.M. Occurrence and activity of sulphate reducing bacteria in aerobic activated sludge systems. World J. Microbiol. Biotechnol. 2015, 31, 507–516. [Google Scholar] [CrossRef]
- Mohdali, M.K.F.; Ismail, M.; Bakar, A.A.; Noor, N.M.; Yahaya, N.; Zardasti, L.; Sam, A.R.M. Influence of environmental parameters on microbiologically influenced corrosion subject to different bacteria strains. Sains Malays. 2020, 49, 671–682. [Google Scholar] [CrossRef]
- Bitenieks, K.; Bārdule, A.; Eklöf, K.; Espenberg, M.; Ruņģis, D.E.; Kļaviņa, Z.; Kļaviņš, I.; Hu, H.; Lībiete, Z. The Influence of the degree of forest management on methylmercury and the composition of microbial communities in the sediments of boreal drainage ditches. Microorganisms 2022, 10, 1981. [Google Scholar] [CrossRef] [PubMed]
- Ramel, F.; Brasseur, G.; Pieulle, L.; Valette, O.; Hirschler-Réa, A.; Fardeau, M.L.; Dolla, A. Growth of the obligate anaerobe Desulfovibrio vulgaris Hildenborough under continuous low oxygen concentration sparging: Impact of the membrane-bound oxygen reductases. PLoS ONE 2015, 10, e0123455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Zhang, Z. Oxygen Tolerance of sulfate reducing bacteria in fresh and coastal sea qaters under enrichment culture conditions. Geol. J. China Univ. 2019, 25, 705–713. [Google Scholar] [CrossRef]
- Heggendorn, F.L.; Gonçalves, L.D.; Dias, E.P.; Heggendorn, C.; Lutterbach, M.T. Detection of sulphate-reducing bacteria and others cultivable facultative bacteria in dental tissues. Acta Stomatol. Croat. 2014, 48, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, X.L.; Li, Z.G.; Zhang, F.L. Experimental study on separation, identification and treatment ofc Chrome in metallurgical acid wastewater by SRB. J. Yanan Univ. (Nat. Sci. Ed.) 2020, 39, 65–70. [Google Scholar] [CrossRef]
- Chen, W. Research of Polyphasic Taxonomy and Integrated Control to Sulfate-Reducing Bacteria. Ph.D. Thesis, Huazhong University of Science and Technology, Wuhan, China, 2006; pp. 1–132. (In Chinese). [Google Scholar]
- Ma, Q.L.; Du, H.; Liu, Y.; Li, M. Sulfate-reducing prokaryotes in mangrove wetlands: Diversity and role in driving element coupling. Acta Microbiol. Sin. 2022, 62, 4606–4627. [Google Scholar] [CrossRef]
- Zou, S.B.; Gao, Q.; Cheng, H.H.; Ni, M.; Xu, Q.; Liu, M.; Zhou, D.; Zhou, Z.M.; Yuan, J.L. Vertical distribution of bacterial, sulfate-reducing and sulfur-oxidizing bacterial communities in sediment cores from freshwater prawn (Macrobrachium rosenbergii) aquaculture pond. Acta Microbiol. Sin. 2022, 62, 2719–2734. [Google Scholar] [CrossRef]
- Cheng, Y.; Song, T.L.; Tian, X.G.; Wang, Y.H. Distribution characteristics and environmental significance of sulfate-reducing bacterial community in high arsenic groundwater from different depth of Hetao Plain, China. Acta Microbiol. Sin. 2022, 62, 2372–2388. [Google Scholar] [CrossRef]
- Zhan, P.F.; Huang, J.F.; Yu, C.X.; Tong, C. Effects of saltwater and Fe (III) pulses on community structure and abundance of methanogens and sulfate-reducing bacteria in tidal freshwater marsh of the Min River estuary. Acta Sci. Circumstantiae 2020, 40, 2599–2610. [Google Scholar] [CrossRef]
- Cui, Y.Y.; Qin, Y.X.; Ding, Q.M.; Gao, Y.N. Study on corrosion behavior of X80 steel under stripping coating by sulfate reducing bacteria. BMC Biotechnol. 2021, 21, 5. [Google Scholar] [CrossRef]
- Yan, L.; Zhao, H. Advances in enzyme properties and applications of sulfate-reducing bacteria. Chin. Agric. Sci. Bull. 2020, 36, 13–19. [Google Scholar] [CrossRef]
- Song, C.; Zheng, C.L.; Wang, J.Y. Microorganism sulfate assimilation pathway and its relationship with heavy metal resistance. J. Anhui Agric. Sci. 2012, 40, 6368–6370+6400. [Google Scholar] [CrossRef]
- Jin, Z.; Wang, W.; Li, X.; Zhou, H.; Yi, G.; Wang, Q.; Yu, F.; Xiao, X.; Liu, X. Structure and function of piezophilic hyperthermophilic Pyrococcus yayanosii pApase. Int. J. Mol. Sci. 2021, 22, 7159. [Google Scholar] [CrossRef]
- Schwarz, A.; Gaete, M.; Nancucheo, I.; Villa-Gomez, D.; Aybar, M.; Sbárbaro, D. High-rate sulfate removal coupled to elemental sulfur production in mining process waters based on membrane-biofilm technology. Front. Bioeng. Biotechnol. 2022, 10, 805712. [Google Scholar] [CrossRef]
- Schweitzer, H.D.; Smith, H.J.; Barnhart, E.P.; McKay, L.J.; Gerlach, R.; Cunningham, A.B.; Malmstrom, R.R.; Goudeau, D.; Fields, M.W. Subsurface hydrocarbon degradation strategies in low- and high-sulfate coal seam communities identified with activity-based metagenomics. NPJ Biofilms Microbiomes 2022, 8, 7. [Google Scholar] [CrossRef]
- Wei, Z.; Zhang, Z.; Zhao, W.; Yin, T.; Liu, X.; Zhang, H. Overexpression of MET4 leads to the upregulation of stress-related genes and enhanced sulfite tolerance in Saccharomyces uvarum. Cells 2022, 11, 636. [Google Scholar] [CrossRef]
- Li, X.H. Screening of Anaerobic and Facultative Anaerobic Sulfate-Reducing Bacteria in Corrosive Steel Layer and Comparison of Their Corrosion Properties. Ph.D. Thesis, Ocean University of China, Qingdao, China, 2019; pp. 1–113. (In Chinese). [Google Scholar]
- Lin, W.; Yu, Z.; Zhang, H.; Thompson, I.P. Diversity and dynamics of microbial communities at each step of treatment plant for potable water generation. Water Res. 2014, 52, 218–230. [Google Scholar] [CrossRef]
- Zhang, Y.; Duan, J.; Li, X.; Wang, J.; Hou, B. Analysis of marine microbial communities colonizing various metallic materials and rust layers. Biofouling 2019, 35, 429–442. [Google Scholar] [CrossRef]
- Païssé, S.; Ghiglione, J.F.; Marty, F.; Abbas, B.; Gueuné, H.; Amaya, J.M.S.; Muyzer, G.; Quillet, L. Sulfate-reducing bacteria inhabiting natural corrosion deposits from marine steel structures. Appl. Microbiol. Biotechnol. 2013, 97, 7493–7504. [Google Scholar] [CrossRef] [PubMed]
- Postgate, J.R. The Sulphate Reducing Bacteria, 2nd ed.; Cambridge University Press: New York, NY, USA, 1983; p. 224. [Google Scholar]
- Li, X.H.; Xiao, H.; Zhang, W.; Li, Y.; Tang, X.; Duan, J.; Yang, Z.; Wang, J.; Guan, F.; Ding, G. Analysis of cultivable aerobic bacterial community composition and screening for facultative sulfate-reducing bacteria in marine corrosive steel. J. Oceanol. Limnol. 2019, 37, 600–614. [Google Scholar] [CrossRef]
- Pan, J.; Shao, Z.; Cao, H.; Sheng, Y. Review on the isolation and purification methods of sulfate reducing bacteria. J. Microbiol. 2007, 27, 79–83. [Google Scholar] [CrossRef]
- Liu, N. Characterization and Identification of SRB and Its Control Method. Master’s Thesis, Huazhong University of Science and Technology, Wuhan, China, 2012; pp. 1–72. (In Chinese). [Google Scholar]
- Plugge, C.M.; Zhang, W.; Scholten, J.C.; Stams, A.J. Metabolic flexibility of sulfate-reducing bacteria. Front. Microbiol. 2011, 2, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visser, M.; Stams, A.J.M.; Frutschi, M.; Bernier-Latmani, R. Phylogenetic comparison of Desulfotomaculum species of subgroup 1a and description of Desulfotomaculum reducens sp nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 762–767. [Google Scholar] [CrossRef] [PubMed]
- Sun, W. Isolation Identification and Phylogenetic Analysis of Three Anaerobic Functional Strains. Master’s Thesis, Harbin Institute of Technology, Harbin, China, 2008; pp. 1–91. [Google Scholar] [CrossRef]
- Zhang, X.L.; Chen, Z.X.; Liu, H.H.; Guo, S.W.; Chen, K.X. Effect of environment factors on the growth of sulfate-reducing bacteria. J. Chin. Soc. Corros. Prot. 2000, 20, 224–229. [Google Scholar]
- Sigalevich, P.; Meshorer, E.; Helman, Y.; Cohen, Y. Transition from anaerobic to aerobic growth conditions for the sulfate-reducing bacterium Desulfovibrio oxyclinae results in flocculation. Appl. Environ. Microb. 2000, 66, 5005–5012. [Google Scholar] [CrossRef] [Green Version]
- McBeth, J.M.; Emerson, D. In situ microbial community succession on mild steel in estuarine and marine environments: Exploring the role of iron-oxidizing bacteria. Front. Microbiol. 2016, 7, 767. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Tian, Y.; Kang, M.; Chen, C.; Song, Y.; Li, H. Effects of chlorination/chlorine dioxide disinfection on biofilm bacterial community and corrosion process in a reclaimed water distribution system. Chemosphere 2019, 215, 62–73. [Google Scholar] [CrossRef]
- Vigneron, A.; Alsop, E.B.; Chambers, B.; Lomans, B.P.; Head, I.M.; Tsesmetzis, N. Complementary microorganisms in highly corrosive biofilms from an offshore oil production facility. Appl. Environ. Microb. 2016, 82, 2545–2554. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Xu, D.; Li, Y.; Yang, K.; Gu, T. Electron mediators accelerate the microbiologically influenced corrosion of 304 stainless steel by the Desulfovibrio vulgaris biofilm. Bioelectrochemistry 2015, 101, 14–21. [Google Scholar] [CrossRef]
- Zhang, Y.; Pei, G.; Chen, L.; Zhang, W. Metabolic dynamics of Desulfovibrio vulgaris biofilm grown on a steel surface. Biofouling 2016, 32, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Häggblom, M.M. Genome guided identification of organohalide-respiring deltaproteobacteria from the marine environment. MBio 2018, 9, 1110–1128. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.R.; Hosokawa, H.; Kuroda, M.; Inoue, D.; Ike, M. Microbial antimonate reduction and removal potentials in river sediments. Chemosphere 2021, 266, 10. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Kang, S.J.; Lee, S.Y.; Lee, M.H.; Oh, T.K. Virgibacillus dokdonensis sp. nov. isolated from a Korean island, Dokdo, located at the edge of the east sea in Korea. Int. J. Syst. Evol. Microbiol. 2005, 55, 1833. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.Y.; Chang, C.C.; Chang, C.N.; Chen, T.W.; Shyu, Y.T. Virgibacillus chiguensis sp nov. a novel halophilic bacterium isolated from Chigu, a previously commercial saltern located in southern Taiwan. Int. J. Syst. Evol. Microbiol. 2008, 58, 341–345. [Google Scholar] [CrossRef]
- Chen, Y.H.; Shyu, Y.T.; Lin, S.S. Characterization of candidate genes involved in halotolerance using high-throughput omics in the halotolerant bacterium Virgibacillus chiguensis. PLoS ONE 2018, 13, e0201346. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.Y.; Lee, H.C.; Ko, N.Y.; Chang, C.M.; Shih, H.I.; Wu, C.J.; Ko, W.C. Clinical and economic impact of multidrug resistance in nosocomial Acinetobacter baumannii bacteremia. Infect. Control Hosp. Epidemiol. 2007, 28, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Zhao, Y.; Yan, H.; Zhao, H.; Han, M.; Sun, B.; Sun, X.; Hou, F.; Sun, H.; Han, L.; et al. Struvite precipitation induced by a novel sulfate-reducing bacterium Acinetobacter calcoaceticus SRB4 isolated from river sediment. Geomicrobiol. J. 2015, 32, 868–877. [Google Scholar] [CrossRef]
- Iino, T.; Sakamoto, M.; Ohkuma, M. Prolixibacter denitrificans sp. nov. an iron-corroding, facultatively aerobic, nitrate-reducing bacterium isolated from crude oil, and emended descriptions of the genus Prolixibacter and Prolixibacter bellariivorans. Int. J. Syst. Evol. Microbiol. 2015, 65, 2865–2869. [Google Scholar] [CrossRef]
- Iino, T.; Shono, N.; Ito, K.; Nakamura, R.; Sueoka, K.; Harayama, S.; Ohkuma, M. Nitrite as a causal factor for nitrate-dependent anaerobic corrosion of metallic iron induced by Prolixibacter strains. MicrobiologyOpen 2021, 10, e1225. [Google Scholar] [CrossRef] [PubMed]
- Beveridge, T.J.; Makin, S.A.; Kadurugamuwa, J.L.; Li, Z. Interactions between biofilms and the environment. FEMS Microbiol. Rev. 1997, 20, 291–303. [Google Scholar] [CrossRef]
- Dyksma, S.; Lenk, S.; Sawicka, J.E.; Mußmann, M. Uncultured Gammaproteobacteria and Desulfobacteraceae account for major acetate assimilation in a coastal marine sediment. Front. Microbiol. 2018, 9, 3124. [Google Scholar] [CrossRef] [PubMed]
- Ke, R. Effect of Aerobes and Facultative Aerobes on the Electrochemical Corrosion Behavior of 45 Steel in Seawater in Tropical Marine Environment. Master’s Thesis, Hainan University, Haikou, China, 2014; pp. 1–59. (In Chinese). [Google Scholar]
- Shivani, Y.; Subhash, Y.; Ch, S.; Chv, R. Halodesulfovibrio spirochaetisodalis gen. nov. sp. nov. and reclassification of four Desulfovibrio spp. Int. J. Syst. Evol. Microbiol. 2017, 67, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Nemec, A.; Musilek, M.; Sedo, O.; Baere, T.; Maixnerova, M.; Van, D.; Tanny, J.K.; Zdrahal, Z.; Vaneechoutte, M.; Dijkshoorn, L. Acinetobacter bereziniae sp. nov. and Acinetobacter guillouiae sp. nov. to accommodate Acinetobacter genomic species 10 and 11, respectively. Int. J. Syst. Evol. Microbiol. 2009, 60, 896–903. [Google Scholar] [CrossRef] [Green Version]
- Thakur, P.; Alaba, M.O.; Rauniyar, S.; Singh, R.N.; Saxena, P.; Bomgni, A.; Gnimpieba, E.Z.; Lushbough, C.; Goh, K.M.; Sani, R.K. Text-mining to identify gene sets involved in biocorrosion by sulfate-reducing bacteria: A Semi-Automated Workflow. Microorganisms 2023, 11, 119. [Google Scholar] [CrossRef]
- Wu, G.F.; Gao, H.C. Endogenous production and physiological functions of hydrogen sulfide in facultative anaerobic bacteria. Acta Microbiol. Sin. 2017, 57, 170–178. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order | Characteristic | Standard Strain | Accession | Reference |
|---|---|---|---|---|
| 1 | Anaerobic | Desulfovibrio bizertensis | Y18049 | [30] |
| 2 | Anaerobic | Desulfovibrio zosterae | DQ422859 | [30] |
| 3 | Anaerobic | Pseudodesulfovibrio indicus | CP014206 | [30] |
| 4 | Anaerobic | Desulfotomaculum nigrificans | PRJNA46699 | [37] |
| 5 | Anaerobic | Desulfovibrio vulgaris | PRJNA395924 | [38] |
| 6 | Anaerobic | Desulfotomaculum reducens | PRJNA13424 | [39] |
| 7 | Facultative anaerobic | Vibrio jasicida | AB562589 | [30] |
| 8 | Facultative anaerobic | Vibrio mexicanus | JQ434105 | [30] |
| 9 | Facultative anaerobic | Vibrio owensii | JPRD01000038 | [30] |
| 10 | Facultative anaerobic | Vibrio parahaemolyticus | BBQD01000032 | [30] |
| 11 | Facultative anaerobic | Photobacterium gaetbulicola | CP005974 | [30] |
| 12 | Facultative anaerobic | Staphylococcus epidermidis | L37605 | [30] |
| 13 | Facultative anaerobic | Citrobacter amalonaticus | PRJNA752145 | [17] |
| 14 | Facultative anaerobic | Enterobacter cloacae | PRJNA688591 | [40] |
| 15 | Facultative anaerobic | Desulfovibrio desulfuricans | PRJNA666287 | [41] |
| 16 | Facultative anaerobic | Desulfovibrio oxyclinae | PRJNA169818 | [42] |
| Phylum | Genera | Species | Q345 | Q235 | 921A | Sediment | Total |
|---|---|---|---|---|---|---|---|
| Proteobacteria | Desulfovibrio | Desulfovibrio marinisediminis | 23 | 5 | 11 | 8 | 47 |
| Halodesulfovibrio | Halodesulfovibrio spirochaetisodalis | 1 | 2 | 9 | 6 | 18 | |
| Acinetobacter | Acinetobacter bereziniae | 1 | / | 1 | 1 | 3 | |
| Firmicutes | Virgibacillus | Virgibacillus dokdonensis | 1 | / | / | / | 1 |
| Virgibacillus chiguensis | / | 1 | 3 | 2 | 6 | ||
| Bacteroidetes | Prolixibacter | Prolixibacter bellariivorans | 1 | / | / | / | 1 |
| Prolixibacter denitrificans | 1 | / | / | / | 1 | ||
| Sum | 28 | 8 | 24 | 17 | 77 |
| Characteristic | Q345 | Q235 | 921A | Sediment | Total | Species | Label |
|---|---|---|---|---|---|---|---|
| Facultative anaerobic | 3 | 1 | 4 | 3 | 11 | V. chiguensis | F-V19 |
| A. bereziniae | F-A11 | ||||||
| P. bellariivorans | F-P17 | ||||||
| P. denitrificans | F-P63 | ||||||
| Anaerobic | 25 | 7 | 20 | 14 | 66 | V. dokdonensis | A-V3 |
| D. marinisediminis | A-D60 | ||||||
| H. spirochaetisodalis | A-H54 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Li, X.; Guan, F.; Yang, Z.; Zhai, X.; Zhang, Y.; Tang, X.; Duan, J.; Xiao, H. The Isolation of Anaerobic and Facultative Anaerobic Sulfate-Reducing Bacteria (SRB) and a Comparison of Related Enzymes in Their Sulfate Reduction Pathways. Microorganisms 2023, 11, 2019. https://doi.org/10.3390/microorganisms11082019
Wang J, Li X, Guan F, Yang Z, Zhai X, Zhang Y, Tang X, Duan J, Xiao H. The Isolation of Anaerobic and Facultative Anaerobic Sulfate-Reducing Bacteria (SRB) and a Comparison of Related Enzymes in Their Sulfate Reduction Pathways. Microorganisms. 2023; 11(8):2019. https://doi.org/10.3390/microorganisms11082019
Chicago/Turabian StyleWang, Jing, Xiaohong Li, Fang Guan, Zhibo Yang, Xiaofan Zhai, Yimeng Zhang, Xuexi Tang, Jizhou Duan, and Hui Xiao. 2023. "The Isolation of Anaerobic and Facultative Anaerobic Sulfate-Reducing Bacteria (SRB) and a Comparison of Related Enzymes in Their Sulfate Reduction Pathways" Microorganisms 11, no. 8: 2019. https://doi.org/10.3390/microorganisms11082019