
The FilZ Protein Contains a Single PilZ Domain and Facilitates the Swarming Motility of Pseudoalteromonas sp. SM9913
 , , and
, , and 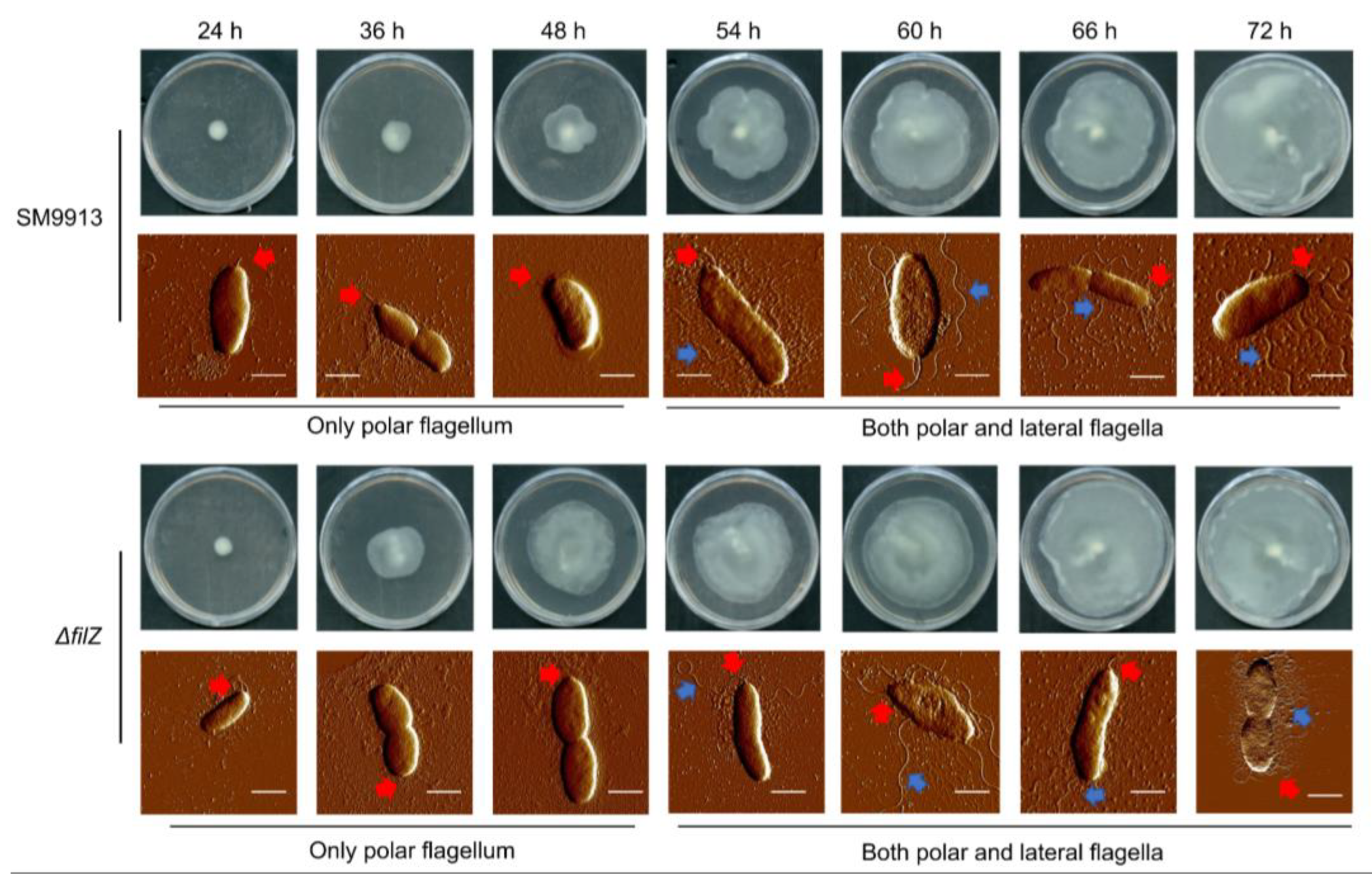{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Plasmids, and Growth Conditions
2.2. Gene Cloning, Mutation, and Protein Overexpression
2.3. Protein Purification
2.4. Isothermal Titration Calorimetry
2.5. Construction of the Mutants of Strain SM9913
2.6. Construction of Complementary Strains of SM9913
2.7. Bacterial Motility Assay
2.8. Total RNA Extraction and RT-qPCR
2.9. Atomic Force Microscopy Imaging
2.10. The Construction and Screening of E. coli BACTH Library and Two-Hybrid Assays
2.11. Bacterial Two-Hybrid Assays
2.12. Extraction and Detection of c-di-GMP
2.13. Pull-Down Experiment
2.14. Immunoblot
2.15. Chemotaxis Assays
3. Results
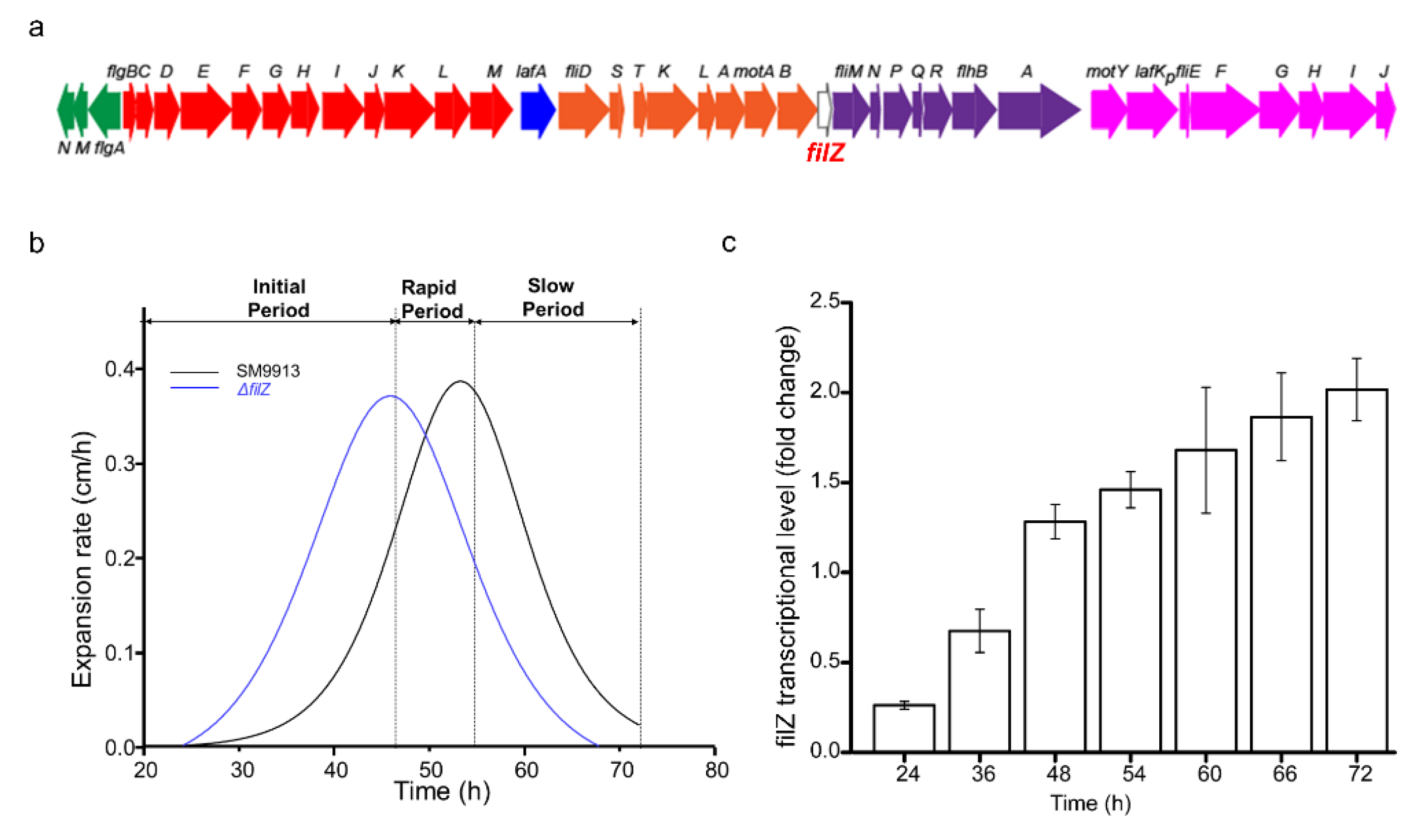
3.1. Effect of Protein FilZ on the Swarming of Strain SM9913
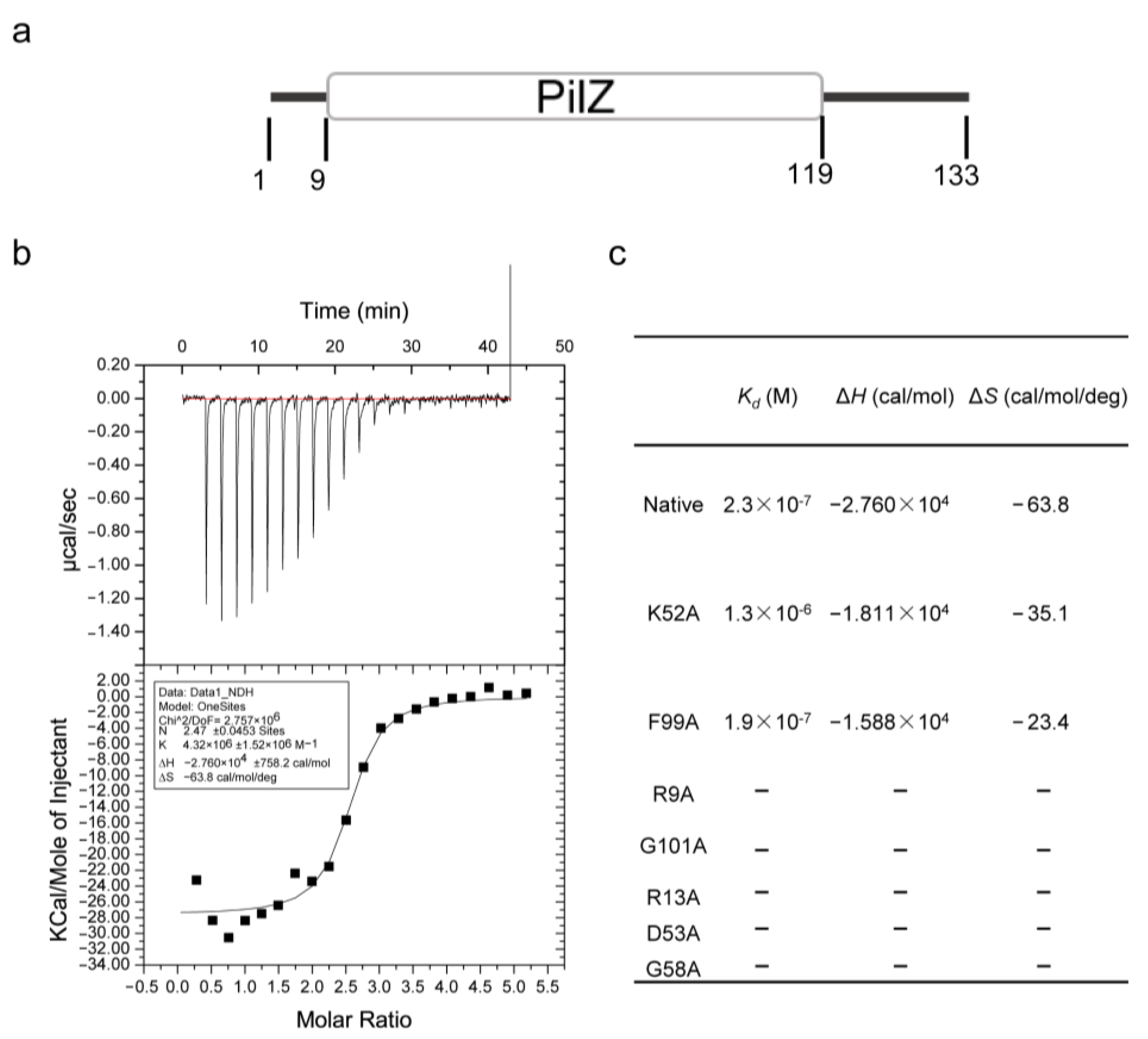
3.2. FilZ Has a Single PilZ Domain That Binds c-di-GMP
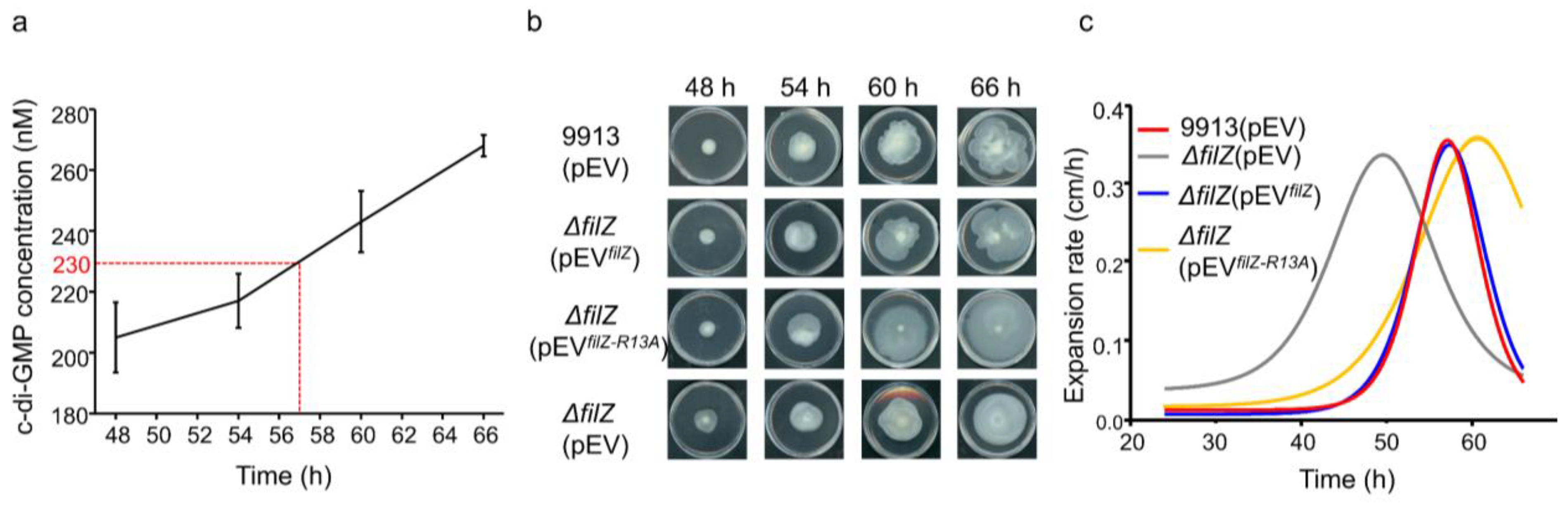
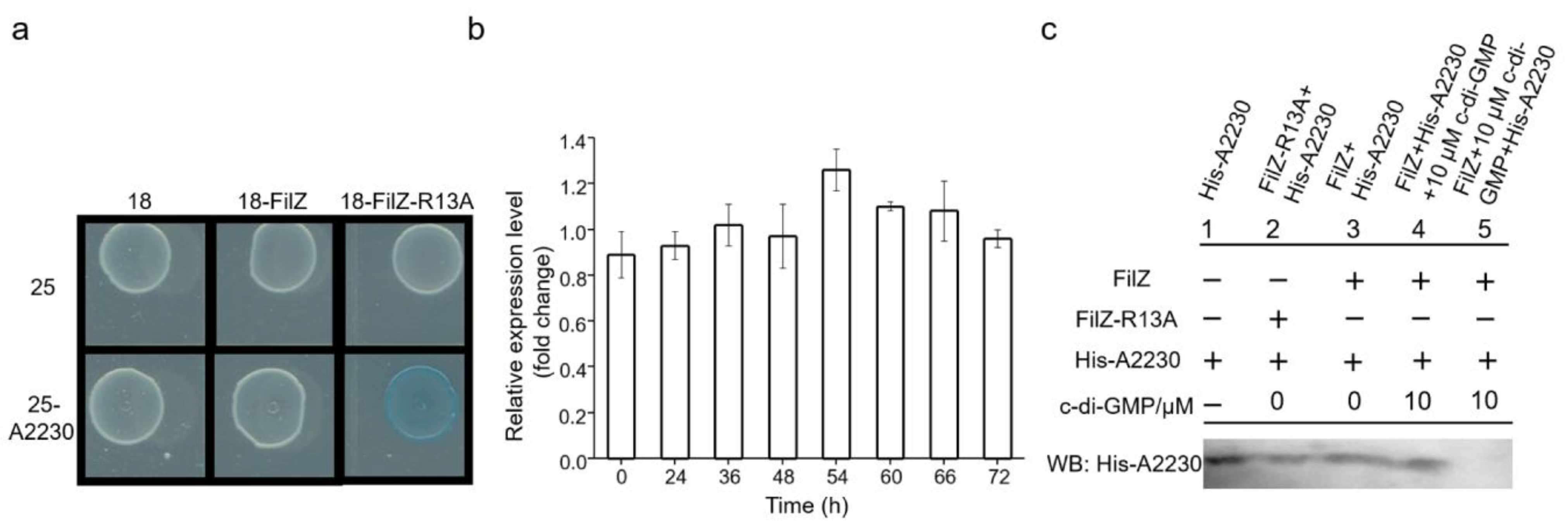
3.3. FilZ Activity Is Negatively Controlled by c-di-GMP In Vivo
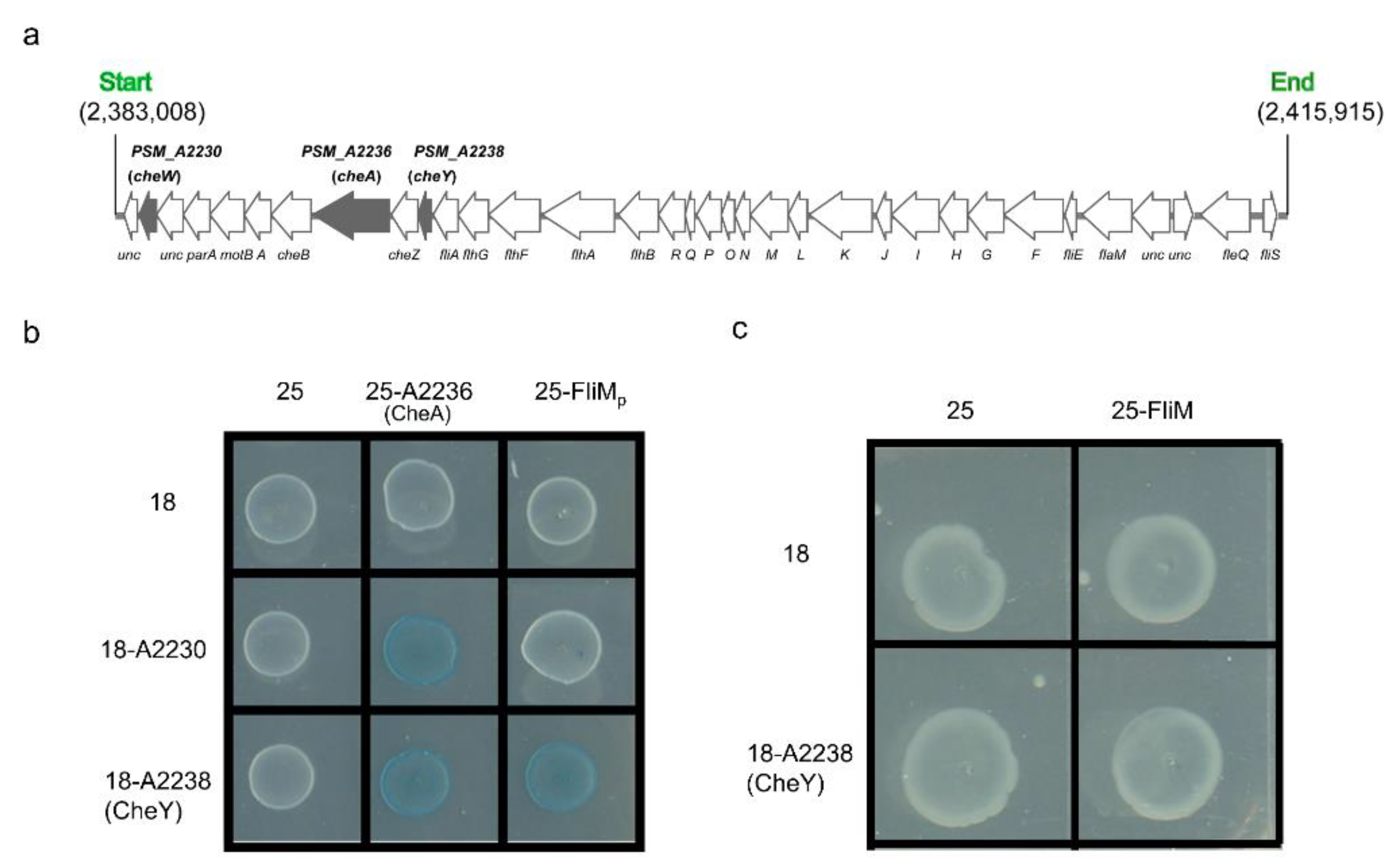
3.4. FilZ Interacts with the CheW-like Protein A2230
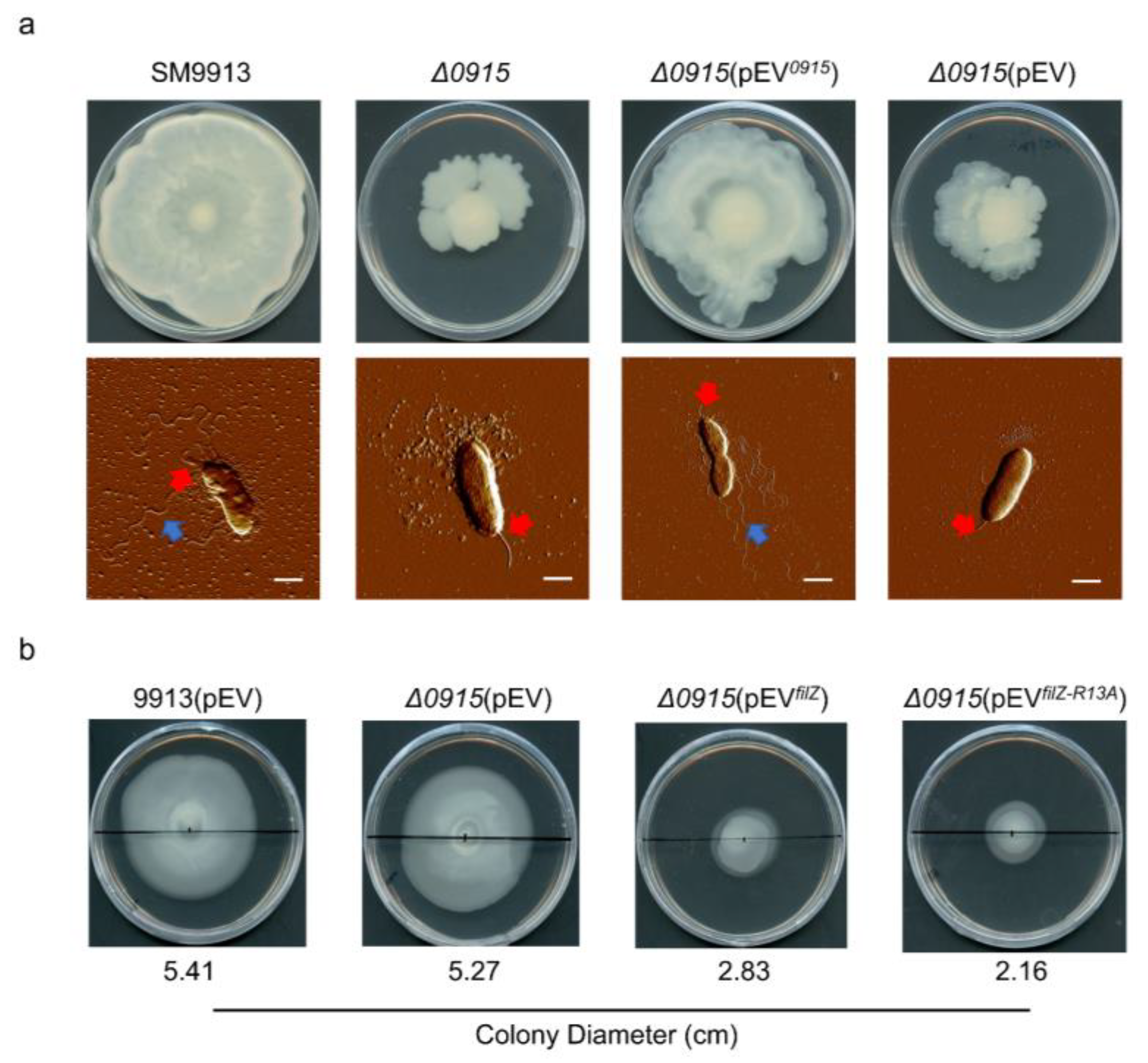
3.5. FilZ Interferes with the Polar Flagellar Motility
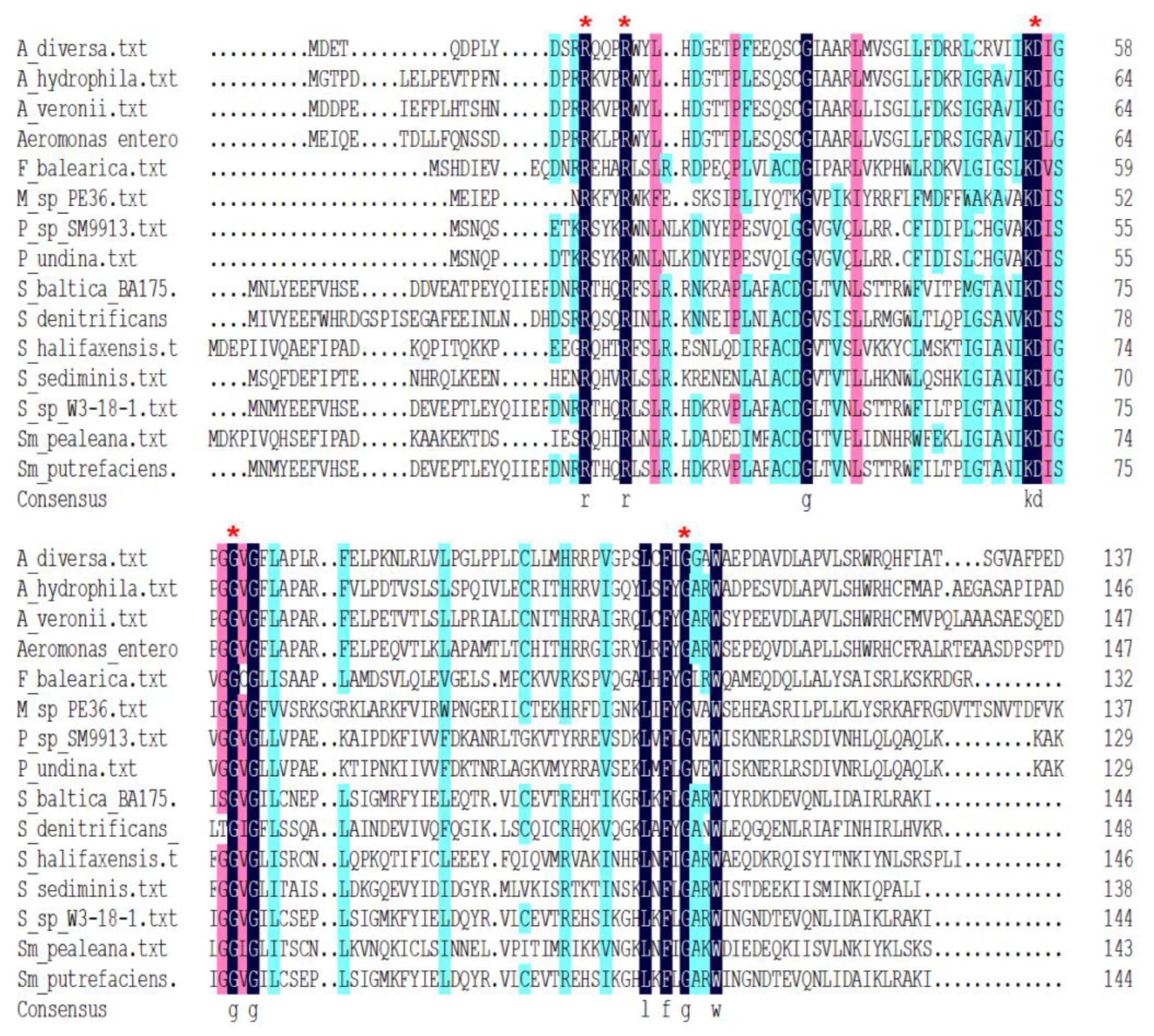
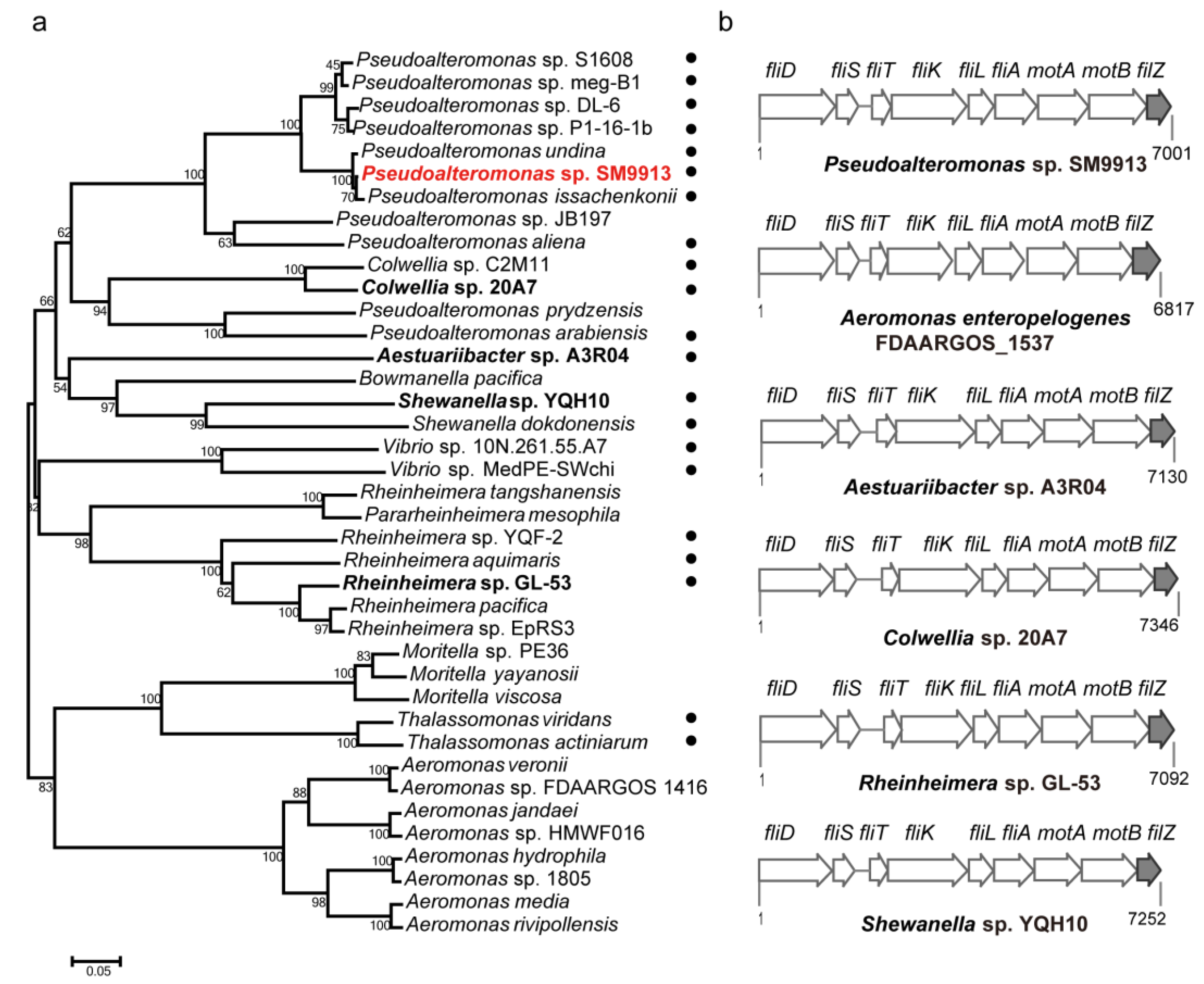
3.6. The Phylogenetic Distribution of FilZ
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nan, B.; Zusman, D.R. Novel mechanisms power bacterial gliding motility. Mol. Microbiol. 2016, 101, 186–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.; Carroll, B.L.; Liu, J. Structural basis of bacterial flagellar motor rotation and switching. Trends Microbiol. 2021, 29, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Kearns, D.B. A field guide to bacterial swarming motility. Nat. Rev. Microbiol. 2010, 8, 634–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wadhwa, N.; Berg, H.C. Bacterial motility: Machinery and mechanisms. Nat. Rev. Microbiol. 2022, 20, 161–173. [Google Scholar] [CrossRef]
- Partridge, J.D.; Harshey, R.M. Swarming: Flexible roaming plans. J. Bacteriol. 2013, 195, 909–918. [Google Scholar] [CrossRef] [Green Version]
- Harshey, R.M. Bees aren’t the only ones: Swarming in gram-negative bacteria. Mol. Microbiol. 1994, 13, 389–394. [Google Scholar] [CrossRef]
- Verstraeten, N.; Braeken, K.; Debkumari, B.; Fauvart, M.; Fransaer, J.; Vermant, J.; Michiels, J. Living on a surface: Swarming and biofilm formation. Trends Microbiol. 2008, 16, 496–506. [Google Scholar] [CrossRef]
- McCarter, L.L. Dual flagellar systems enable motility under different circumstances. J. Mol. Microbiol. Biotechnol. 2004, 7, 18–29. [Google Scholar] [CrossRef]
- Merino, S.; Shaw, J.G.; Tomas, J.M. Bacterial lateral flagella: An inducible flagella system. FEMS Microbiol. Lett. 2006, 263, 127–135. [Google Scholar] [CrossRef]
- Bubendorfer, S.; Held, S.; Windel, N.; Paulick, A.; Klingl, A.; Thormann, K.M. Specificity of motor components in the dual flagellar system of Shewanella putrefaciens CN-32. Mol. Microbiol. 2012, 83, 335–350. [Google Scholar] [CrossRef]
- Jian, H.; Wang, H.; Zeng, X.; Xiong, L.; Wang, F.; Xiao, X. Characterization of the relationship between polar and lateral flagellar structural genes in the deep-sea bacterium Shewanella piezotolerans WP3. Sci. Rep. 2016, 6, 39758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarter, L.; Silverman, M. Surface-induced swarmer cell differentiation of Vibrio parahaemolyticus. Mol. Microbiol. 1990, 4, 1057–1062. [Google Scholar] [CrossRef] [PubMed]
- McCarter, L.L.; Wright, M.E. Identification of genes encoding components of the swarmer cell flagellar motor and propeller and a sigma factor controlling differentiation of Vibrio parahaemolyticus. J. Bacteriol. 1993, 175, 3361–3371. [Google Scholar] [CrossRef] [Green Version]
- Stewart, B.J.; McCarter, L.L. Lateral flagellar gene system of Vibrio parahaemolyticus. J. Bacteriol. 2003, 185, 4508–4518. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, A.J.; Visick, K.L. Get the message out: Cyclic-di-GMP regulates multiple levels of flagellum-based motility. J. Bacteriol. 2008, 190, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Benach, J.; Swaminathan, S.S.; Tamayo, R.; Handelman, S.K.; Folta-Stogniew, E.; Ramos, J.E.; Forouhar, F.; Neely, H.; Seetharaman, J.; Camilli, A.; et al. The structural basis of cyclic diguanylate signal transduction by PilZ domains. EMBO J. 2007, 26, 5153–5166. [Google Scholar] [CrossRef] [Green Version]
- Cheang, Q.W.; Xin, L.; Chea, R.Y.F.; Liang, Z.X. Emerging paradigms for PilZ domain-mediated c-di-GMP signaling. Biochem. Soc. Trans. 2019, 47, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Paul, K.; Nieto, V.; Carlquist, W.C.; Blair, D.F.; Harshey, R.M. The c-di-GMP binding protein YcgR controls flagellar motor direction and speed to affect chemotaxis by a “backstop brake” mechanism. Mol. Cell. 2010, 38, 128–139. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, S.; Gao, X.; Dann, C.E., III; Kearns, D.B. MotI (DgrA) acts as a molecular clutch on the flagellar stator protein MotA in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2017, 114, 13537–13542. [Google Scholar] [CrossRef] [Green Version]
- Bense, S.; Bruchmann, S.; Steffen, A.; Stradal, T.E.B.; Haussler, S.; Duvel, J. Spatiotemporal control of FlgZ activity impacts Pseudomonas aeruginosa flagellar motility. Mol. Microbiol. 2019, 111, 1544–1557. [Google Scholar] [CrossRef]
- Kojima, S.; Yoneda, T.; Morimoto, W.; Homma, M. Effect of PlzD, a YcgR homolog of c-di-GMP binding protein, on polar flagellar motility in Vibrio alginolyticus. J. Biochem. 2019, 166, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Pecina, A.; Schwan, M.; Blagotinsek, V.; Rick, T.; Kluber, P.; Leonhard, T.; Bange, G.; Thormann, K.M. The stand-alone PilZ-domain protein MotL specifically regulates the activity of the secondary lateral flagellar system in Shewanella putrefaciens. Front. Microbiol. 2021, 12, 668892. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q.L.; Li, Y.; Zhang, Y.J.; Zhou, Z.M.; Zhang, W.X.; Chen, X.L.; Zhang, X.Y.; Zhou, B.C.; Wang, L.; Zhang, Y.Z. Comparative genomics reveals a deep-sea sediment-adapted life style of Pseudoalteromonas sp. SM9913. ISME J. 2011, 5, 274–284. [Google Scholar] [CrossRef] [Green Version]
- Eloe, E.A.; Lauro, F.M.; Vogel, R.F.; Bartlett, D.H. The deep-sea bacterium Photobacterium profundum SS9 utilizes separate flagellar systems for swimming and swarming under high-pressure conditions. Appl. Environ. Microbiol. 2008, 74, 6298–6305. [Google Scholar] [CrossRef] [Green Version]
- Stocker, R.; Seymour, J.R. Ecology and physics of bacterial chemotaxis in the ocean. Microbiol. Mol. Biol. Rev. 2012, 76, 792–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, Z.H.; Yu, Z.C.; Su, H.N.; Wang, L.; Chen, X.L.; Pang, X.; Qin, Q.L.; Xie, B.B.; Zhang, X.Y.; Zhou, B.C.; et al. Physiological and genetic analyses reveal a mechanistic insight into the multifaceted lifestyles of Pseudoalteromonas sp. SM9913 adapted to the deep-sea sediment. Environ. Microbiol. 2015, 17, 3795–3806. [Google Scholar] [CrossRef]
- Sheng, Q.; Liu, S.M.; Cheng, J.H.; Li, C.Y.; Fu, H.H.; Zhang, X.Y.; Song, X.Y.; McMinn, A.; Zhang, Y.Z.; Su, H.N.; et al. Lack of N-terminal segment of the flagellin protein results in the production of a shortened polar flagellum in the deep-sea sedimentary bacterium Pseudoalteromonas sp. strain SM9913. Appl. Environ. Microbiol. 2021, 87, e0152721. [Google Scholar] [CrossRef]
- Li, P.Y.; Yao, Q.Q.; Wang, P.; Zhang, Y.; Li, Y.; Zhang, Y.Q.; Hao, J.; Zhou, B.C.; Chen, X.L.; Shi, M.; et al. A novel subfamily esterase with a homoserine transacetylase-like fold but no transferase activity. Appl. Environ. Microbiol. 2017, 83, e00131-17. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Tian, F.; Li, X.; Fan, S.; Chen, H.; Wu, M.; Yang, C.H.; He, C. The degenerate EAL-GGDEF domain protein Filp functions as a cyclic di-GMP receptor and specifically interacts with the PilZ-domain protein PXO_02715 to regulate virulence in Xanthomonas oryzae pv. oryzae. Mol. Plant-Microbe Interact. MPMI 2014, 27, 578–589. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.; Ryu, K.S.; Kim, H.; Shin, J.S.; Lee, J.O.; Cheong, C.; Choi, B.S. Structure of PP4397 reveals the molecular basis for different c-di-GMP binding modes by Pilz domain proteins. J. Mol. Biol. 2010, 398, 97–110. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, P.; Zhang, J.; Wang, J.; Lu, Y.; Pang, X. Impact on multiple antibiotic pathways reveals MtrA as a master regulator of antibiotic production in Streptomyces spp. and potentially in other Actinobacteria. Appl. Environ. Microbiol. 2020, 86, e01201-20. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhang, P.; Zhang, J.; Xu, W.; Wang, X.; Wu, L.; Sheng, D.; Ma, W.; Cao, G.; Chen, X.L.; et al. The developmental regulator MtrA binds GlnR boxes and represses nitrogen metabolism genes in Streptomyces coelicolor. Mol. Microbiol. 2019, 112, 29–46. [Google Scholar] [CrossRef] [PubMed]
- de Bentzmann, S.; Bordi, C. Construction of Pseudomonas aeruginosa two-hybrid libraries for high-throughput assays. Methods Mol. Biol. 2014, 1149, 225–241. [Google Scholar] [CrossRef]
- Karimova, G.; Davi, M.; Ladant, D. The beta-lactam resistance protein Blr, a small membrane polypeptide, is a component of the Escherichia coli cell division machinery. J. Bacteriol. 2012, 194, 5576–5588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, S.W.; Li, B.Y.; Tang, K.H.; Yao, J.Y.; Wood, T.K.; Wang, P.X.; Wang, X.X. Conjugative plasmid-encoded toxin-antitoxin system PrpT/PrpA directly controls plasmid copy number. Proc. Natl. Acad. Sci. USA 2021, 118, e2011577118. [Google Scholar] [CrossRef]
- Petrova, O.E.; Sauer, K. High-performance liquid chromatography (HPLC)-based detection and quantitation of cellular c-di-GMP. Methods Mol. Biol. 2017, 1657, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.J.; Lan, J.F.; Shi, X.Z.; Yang, M.C.; Niu, G.J.; Ding, D.; Zhao, X.F.; Yu, X.Q.; Wang, J.X. Beta-arrestins negatively regulate the toll pathway in shrimp by preventing dorsal translocation and inhibiting dorsal transcriptional activity. J. Biol. Chem. 2016, 291, 7488–7504. [Google Scholar] [CrossRef] [Green Version]
- Tang, B.L.; Yang, J.; Chen, X.L.; Wang, P.; Zhao, H.L.; Su, H.N.; Li, C.Y.; Yu, Y.; Zhong, S.; Wang, L.; et al. A predator-prey interaction between a marine Pseudoalteromonas sp. and Gram-positive bacteria. Nat. Commun. 2020, 11, 285. [Google Scholar] [CrossRef] [Green Version]
- Caiazza, N.C.; O’Toole, G.A. SadB is required for the transition from reversible to irreversible attachment during biofilm formation by Pseudomonas aeruginosa PA14. J. Bacteriol. 2004, 186, 4476–4485. [Google Scholar] [CrossRef] [Green Version]
- Samanta, S.K.; Bhushan, B.; Chauhan, A.; Jain, R.K. Chemotaxis of a Ralstonia sp. SJ98 toward different nitroaromatic compounds and their degradation. Biochem. Biophys. Res. Commun. 2000, 269, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.J.; Yang, W.S.; Hong, Y.; Zhang, Y.; Wang, D.C.; Li, D.F. Structural insights into the mechanism of c-di-GMP-bound YcgR regulating flagellar motility in Escherichia coli. J. Biol. Chem. 2019, 295, 808–821. [Google Scholar] [CrossRef]
- Antani, J.D.; Gupta, R.; Lee, A.H.; Rhee, K.Y.; Manson, M.D.; Lele, P.P. Mechanosensitive recruitment of stator units promotes binding of the response regulator CheY-P to the flagellar motor. Nat. Commun. 2021, 12, 5442. [Google Scholar] [CrossRef] [PubMed]
- Manson, M.D.; Armitage, J.P.; Hoch, J.A.; Macnab, R.M. Bacterial locomotion and signal transduction. J. Bacteriol. 1998, 180, 1009–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belas, R.; Schneider, R.; Melch, M. Characterization of Proteus mirabilis precocious swarming mutants: Identification of rsbA, encoding a regulator of swarming behavior. J. Bacteriol. 1998, 180, 6126–6139. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Belas, R. Loss of FliL alters Proteus mirabilis surface sensing and temperature-dependent swarming. J. Bacteriol. 2015, 197, 159–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toguchi, A.; Siano, M.; Burkart, M.; Harshey, R.M. Genetics of swarming motility in Salmonella enterica serovar Typhimurium: Critical role for lipopolysaccharide. J. Bacteriol. 2000, 182, 6308–6321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soo, P.C.; Horng, Y.T.; Wei, J.R.; Shu, J.C.; Lu, C.C.; Lai, H.C. Regulation of swarming motility and flhDC(Sm) expression by RssAB signaling in Serratia marcescens. J. Bacteriol. 2008, 190, 2496–2504. [Google Scholar] [CrossRef] [Green Version]
- Soo, P.C.; Horng, Y.T.; Fu, Y.H.; Lu, C.C.; Lai, H.C. A potential acyltransferase regulates swarming in Serratia marcescens. Biochem. Biophys. Res. Commun. 2008, 371, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Soo, P.C.; Wei, J.R.; Horng, Y.T.; Hsieh, S.C.; Ho, S.W.; Lai, H.C. Characterization of the dapA-nlpB genetic locus involved in regulation of swarming motility, cell envelope architecture, hemolysin production, and cell attachment ability in Serratia marcescens. Infect. Immun. 2005, 73, 6075–6084. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.R.; Tsai, Y.H.; Soo, P.C.; Horng, Y.T.; Hsieh, S.C.; Ho, S.W.; Lai, H.C. Biochemical characterization of RssA-RssB, a two-component signal transduction system regulating swarming behavior in Serratia marcescens. J. Bacteriol. 2005, 187, 5683–5690. [Google Scholar] [CrossRef] [Green Version]
- Partridge, J.D.; Nhu, N.T.Q.; Dufour, Y.S.; Harshey, R.M. Tumble suppression is a conserved feature of swarming motility. MBio 2020, 11, e01189-20. [Google Scholar] [CrossRef] [PubMed]
- Paul, K.; Brunstetter, D.; Titen, S.; Blair, D.F. A molecular mechanism of direction switching in the flagellar motor of Escherichia coli. Proc. Natl. Acad. Sci. USA 2011, 108, 17171–17176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojima, M.; Kubo, R.; Yakushi, T.; Homma, M.; Kawagishi, I. The bidirectional polar and unidirectional lateral flagellar motors of Vibrio alginolyticus are controlled by a single CheY species. Mol. Microbiol. 2007, 64, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Yu, Z.; Li, B.; Cai, X.; Zeng, Z.; Chen, X.; Wang, X. Development of an efficient conjugation-based genetic manipulation system for Pseudoalteromonas. Microb. Cell Fact. 2015, 14, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheng, Q.; Liu, A.; Yang, P.; Chen, Z.; Wang, P.; Sun, H.; Li, C.; McMinn, A.; Chen, Y.; Zhang, Y.; et al. The FilZ Protein Contains a Single PilZ Domain and Facilitates the Swarming Motility of Pseudoalteromonas sp. SM9913. Microorganisms 2023, 11, 1566. https://doi.org/10.3390/microorganisms11061566
Sheng Q, Liu A, Yang P, Chen Z, Wang P, Sun H, Li C, McMinn A, Chen Y, Zhang Y, et al. The FilZ Protein Contains a Single PilZ Domain and Facilitates the Swarming Motility of Pseudoalteromonas sp. SM9913. Microorganisms. 2023; 11(6):1566. https://doi.org/10.3390/microorganisms11061566
Chicago/Turabian StyleSheng, Qi, Ang Liu, Peiling Yang, Zhuowei Chen, Peng Wang, Haining Sun, Chunyang Li, Andrew McMinn, Yin Chen, Yuzhong Zhang, and et al. 2023. "The FilZ Protein Contains a Single PilZ Domain and Facilitates the Swarming Motility of Pseudoalteromonas sp. SM9913" Microorganisms 11, no. 6: 1566. https://doi.org/10.3390/microorganisms11061566