
A Higher Abundance of Actinomyces spp. in the Gut Is Associated with Spontaneous Preterm Birth

 ,
,  , ,
, ,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Questionnaire
2.3. Food Intake Frequency Evaluation
2.4. Microbial Analysis
2.4.1. Microbial DNA Extraction and Sequencing
2.4.2. Bioinformatic Analysis
2.5. Statistics
3. Results
3.1. Demographic and Anthropometric Data between Mothers and Newborns with or without Preterm Delivery
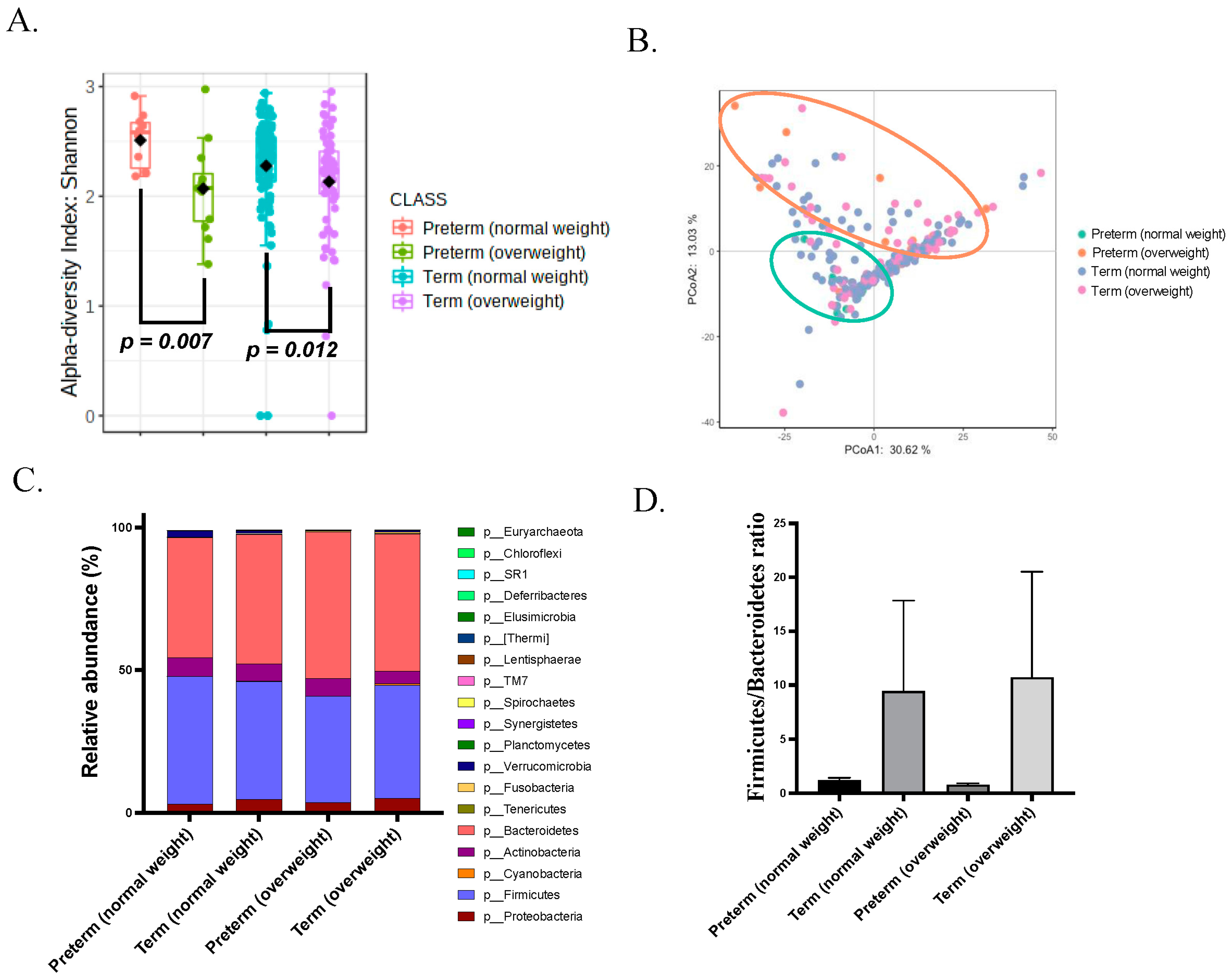
3.2. Gut Microbiota of Overweight Mothers before Pregnancy Is Dysregulated
3.3. Gut Dysbiosis in Mothers with sPTB before Delivery
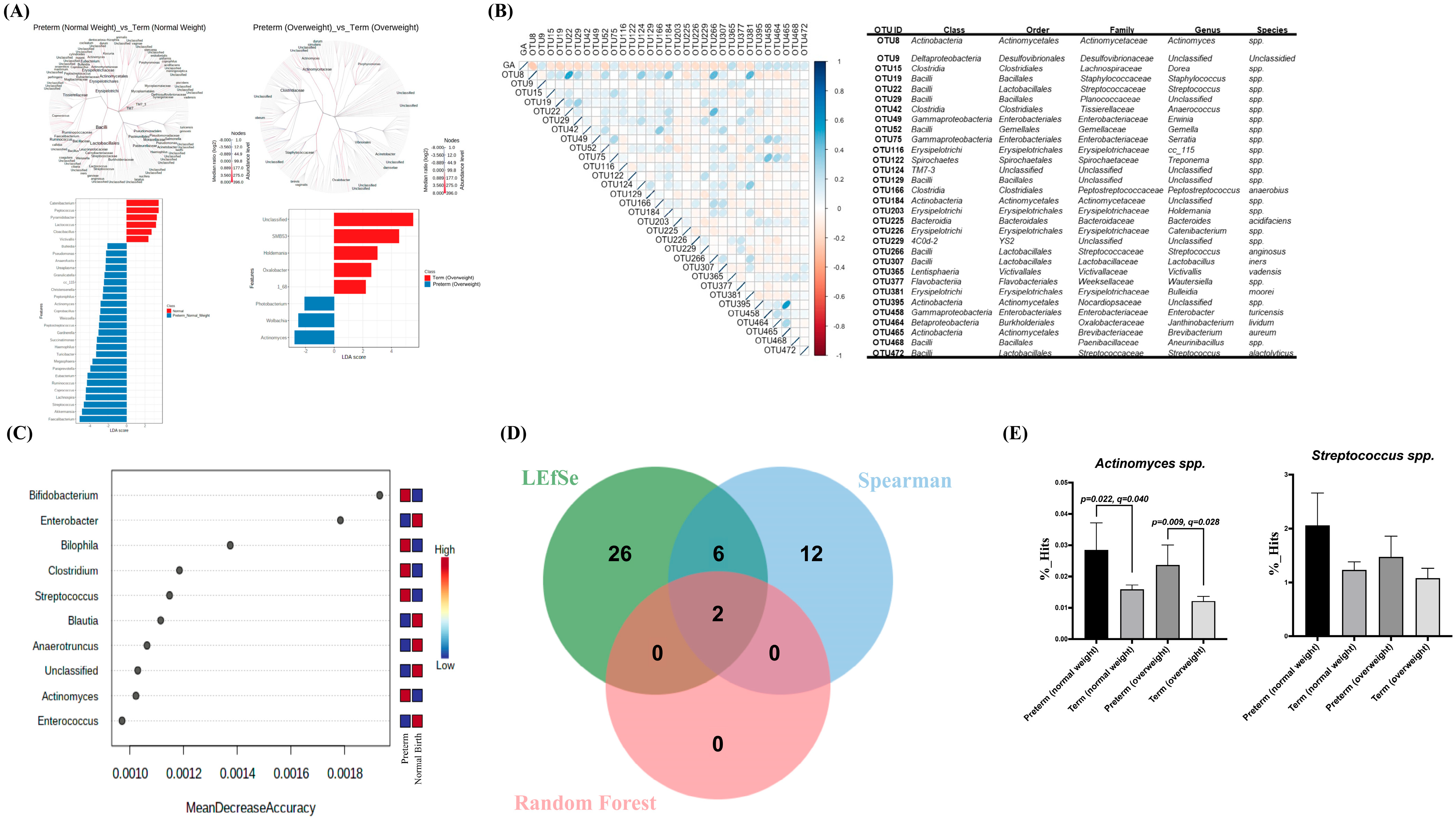
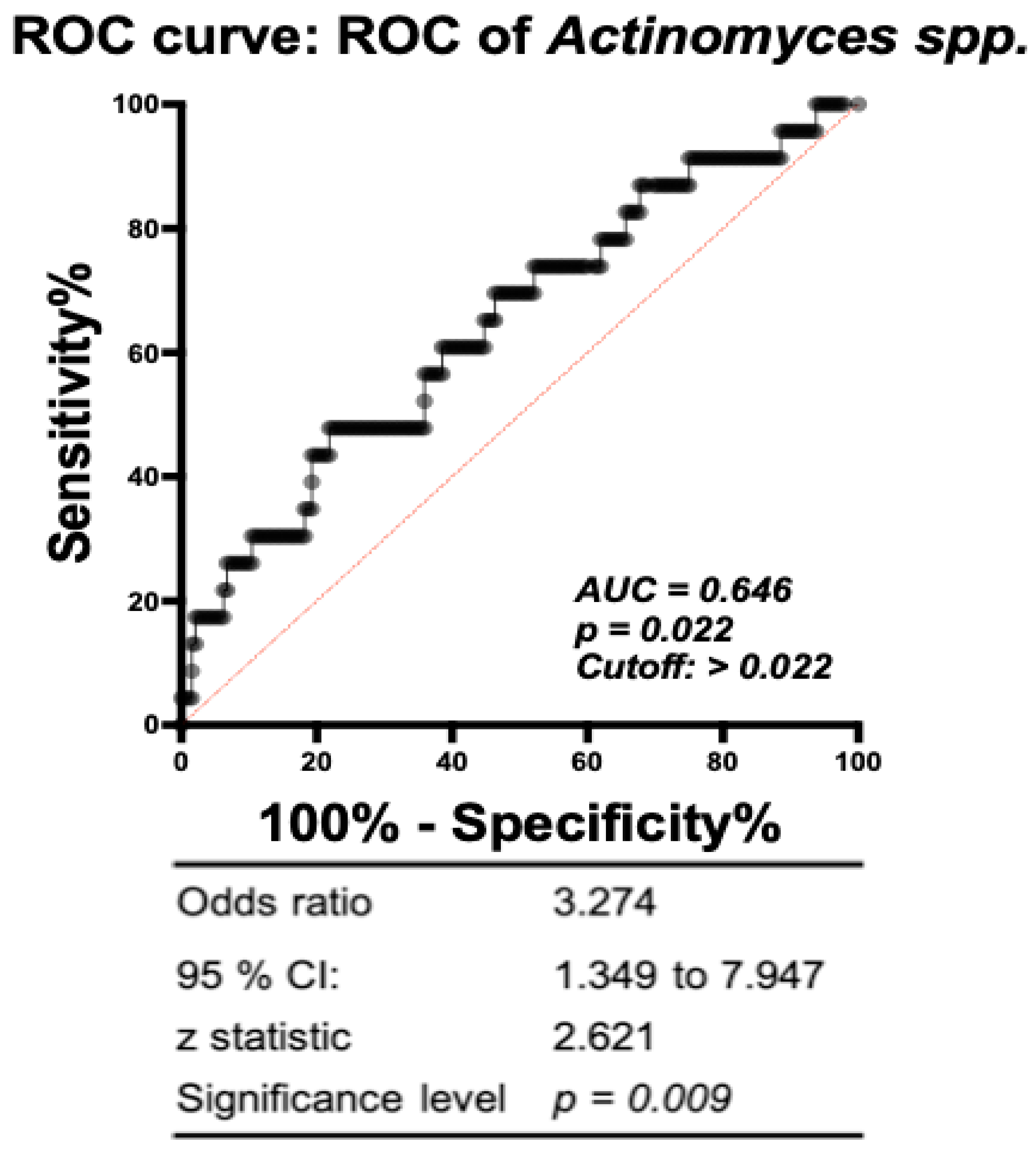
3.4. Association of Specific Microbial Biomarkers with sPTB Risk
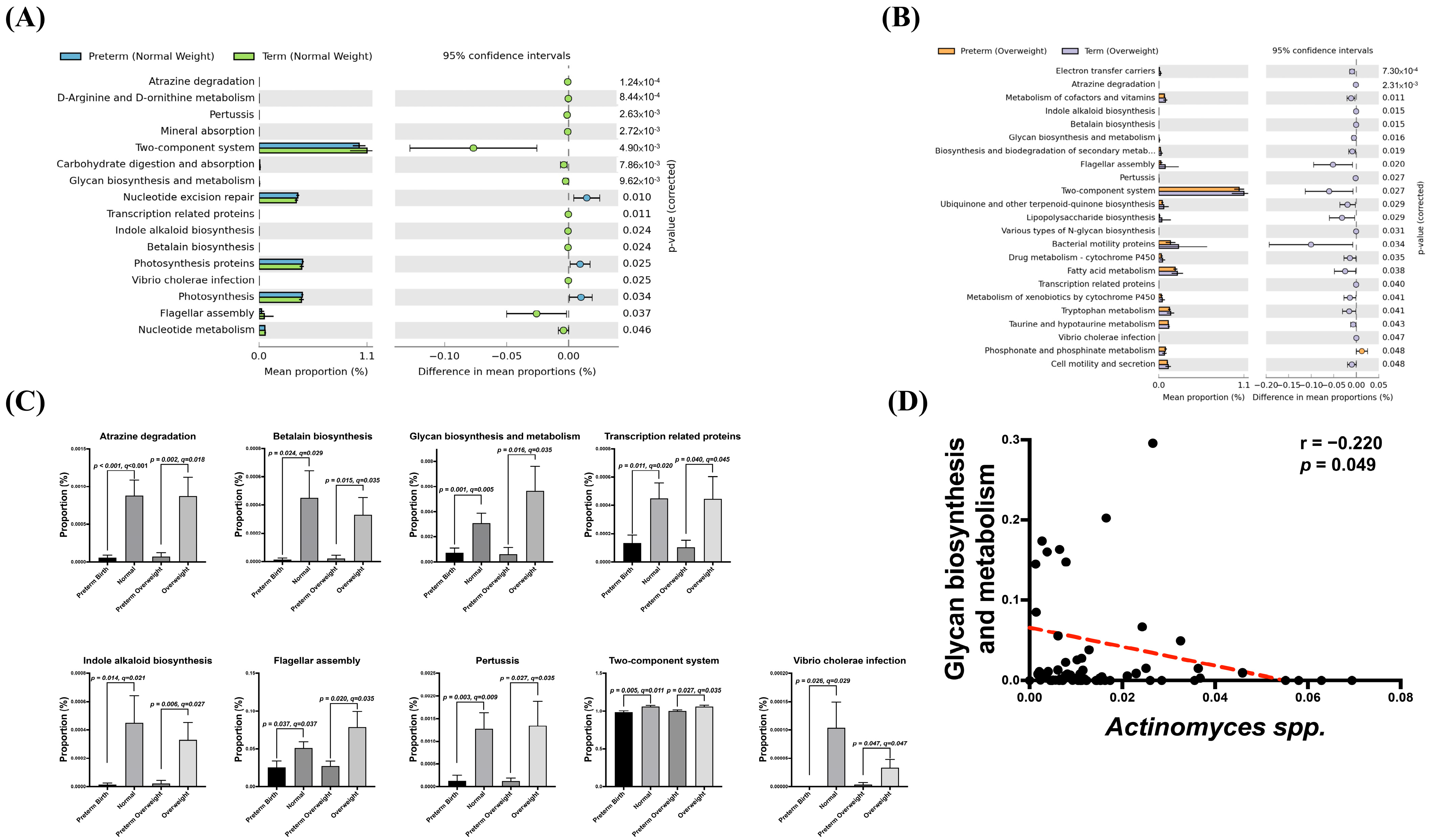
3.5. Association of Potential Gut Dysbiosis-Associated Function Pathways with sPTB Risk
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, L.; Oza, S.; Hogan, D.; Chu, Y.; Perin, J.; Zhu, J.; Lawn, J.E.; Cousens, S.; Mathers, C.; Black, R.E. Global, regional, and national causes of under-5 mortality in 2000–15: An updated systematic analysis with implications for the Sustainable Development Goals. Lancet 2016, 388, 3027–3035. [Google Scholar] [CrossRef] [PubMed]
- Lawn, J.E.; Cousens, S.; Zupan, J. 4 million neonatal deaths: When? Where? Why? Lancet 2005, 365, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Platt, M.J. Outcomes in preterm infants. Public Health 2014, 128, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Menon, R. Spontaneous preterm birth, a clinical dilemma: Etiologic, pathophysiologic and genetic heterogeneities and racial disparity. Acta Obstet. Gynecol. Scand. 2008, 87, 590–600. [Google Scholar] [CrossRef]
- Goldenberg, R.L.; Hauth, J.C.; Andrews, W.W. Intrauterine Infection and Preterm Delivery. N. Engl. J. Med. 2000, 342, 1500–1507. [Google Scholar] [CrossRef]
- Romero, R.; Espinoza, J.; Kusanovic, J.P.; Gotsch, F.; Hassan, S.; Erez, O.; Chaiworapongsa, T.; Mazor, M. The preterm parturition syndrome. BJOG Int. J. Obstet. Gynaecol. 2006, 113 (Suppl. 3), 17–42. [Google Scholar] [CrossRef] [PubMed]
- Gravett, M.G.; Novy, M.J.; Rosenfeld, R.G.; Reddy, A.P.; Jacob, T.; Turner, M.; McCormack, A.; Lapidus, J.A.; Hitti, J.; Eschenbach, D.A.; et al. Diagnosis of Intra-amniotic Infection by Proteomic Profiling and Identification of Novel Biomarkers. JAMA 2004, 292, 462–469. [Google Scholar] [CrossRef]
- Sweeney, E.L.; Kallapur, S.G.; Gisslen, T.; Lambers, D.S.; Chougnet, C.A.; Stephenson, S.-A.; Jobe, A.H.; Knox, C.L. Placental Infection With Ureaplasma species Is Associated With Histologic Chorioamnionitis and Adverse Outcomes in Moderately Preterm and Late-Preterm Infants. J. Infect. Dis. 2015, 213, 1340–1347. [Google Scholar] [CrossRef] [PubMed]
- Andrews, W.W.; Goldenberg, R.L.; Mercer, B.; Iams, J.; Meis, P.; Moawad, A.; Das, A.; VanDorsten, J.P.; Caritis, S.N.; Thurnau, G.; et al. The Preterm Prediction Study: Association of second-trimester genitourinary chlamydia infection with subsequent spontaneous preterm birth. Am. J. Obstet. Gynecol. 2000, 183, 662–668. [Google Scholar] [CrossRef]
- Kahwati, L.C.; Clark, R.; Berkman, N.; Urrutia, R.; Patel, S.V.; Zeng, J.; Viswanathan, M. Screening for Bacterial Vaginosis in Pregnant Adolescents and Women to Prevent Preterm Delivery: Updated Evidence Report and Systematic Review for the US Preventive Services Task Force. JAMA 2020, 323, 1293–1309. [Google Scholar] [CrossRef]
- Pararas, M.V.; Skevaki, C.L.; Kafetzis, D.A. Preterm birth due to maternal infection: Causative pathogens and modes of prevention. Eur. J. Clin. Microbiol. Infect. Dis. 2006, 25, 562–569. [Google Scholar] [CrossRef]
- Andrews, W.W.; Klebanoff, M.A.; Thom, E.A.; Hauth, J.C.; Carey, J.C.; Meis, P.J.; Caritis, S.N.; Leveno, K.J.; Wapner, R.J.; Varner, M.W.; et al. Midpregnancy genitourinary tract infection with Chlamydia trachomatis: Association with subsequent preterm delivery in women with bacterial vaginosis and Trichomonas vaginalis. Am. J. Obstet. Gynecol. 2006, 194, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, R.L.; Culhane, J.F.; Iams, J.D.; Romero, R. Epidemiology and causes of preterm birth. Lancet 2008, 371, 75–84. [Google Scholar] [CrossRef]
- Bayar, E.; Bennett, P.R.; Chan, D.; Sykes, L.; MacIntyre, D.A. The pregnancy microbiome and preterm birth. Semin. Immunopathol. 2020, 42, 487–499. [Google Scholar] [CrossRef]
- Chu, D.M.; Seferovic, M.; Pace, R.M.; Aagaard, K.M. The microbiome in preterm birth. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 52, 103–113. [Google Scholar] [CrossRef]
- Mueller-Heubach, E.; Rubinstein, D.N.; Schwarz, S.S. Histologic chorioamnionitis and preterm delivery in different patient populations. Obstet. Gynecol. 1990, 75, 622–626. [Google Scholar] [PubMed]
- Romero, R.; Avila, C.; Santhanam, U.; Sehgal, P.B. Amniotic fluid interleukin 6 in preterm labor. Association with infection. J. Clin. Investig. 1990, 85, 1392–1400. [Google Scholar] [CrossRef] [PubMed]
- O’hanlon, D.E.; Moench, T.R.; Cone, R.A. Vaginal pH and Microbicidal Lactic Acid When Lactobacilli Dominate the Microbiota. PLoS ONE 2013, 8, e80074. [Google Scholar] [CrossRef] [PubMed]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.K.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4680–4687. [Google Scholar] [CrossRef] [PubMed]
- Tortelli, B.A.; Lewis, W.G.; Allsworth, J.E.; Member-Meneh, N.; Foster, L.R.; Reno, H.E.; Peipert, J.F.; Fay, J.C.; Lewis, A.L. Associations between the vaginal microbiome and Candida colonization in women of reproductive age. Am. J. Obstet. Gynecol. 2020, 222, 471-e1. [Google Scholar] [CrossRef] [PubMed]
- Fettweis, J.M.; Brooks, J.P.; Serrano, M.G.; Sheth, N.U.; Girerd, P.H.; Edwards, D.J.; Strauss, J.F.; Jefferson, K.K.; Buck, G.A. The Vaginal Microbiome Consortium Differences in vaginal microbiome in African American women versus women of European ancestry. Microbiology 2014, 160, 2272–2282. [Google Scholar] [CrossRef]
- Fettweis, J.M.; Serrano, M.G.; Brooks, J.P.; Edwards, D.J.; Girerd, P.H.; Parikh, H.I.; Huang, B.; Arodz, T.J.; Edupuganti, L.; Glascock, A.L.; et al. The vaginal microbiome and preterm birth. Nat. Med. 2019, 25, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; DiGiulio, D.B.; Goltsman, D.S.A.; Sun, C.L.; Costello, E.K.; Jeganathan, P.; Biggio, J.R.; Wong, R.J.; Druzin, M.L.; Shaw, G.M.; et al. Replication and refinement of a vaginal microbial signature of preterm birth in two racially distinct cohorts of US women. Proc. Natl. Acad. Sci. 2017, 114, 9966–9971. [Google Scholar] [CrossRef] [PubMed]
- Kindinger, L.M.; Bennett, P.R.; Lee, Y.S.; Marchesi, J.R.; Smith, A.; Cacciatore, S.; Holmes, E.; Nicholson, J.K.; Teoh, T.G.; MacIntyre, D.A. The interaction between vaginal microbiota, cervical length, and vaginal progesterone treatment for preterm birth risk. Microbiome 2017, 5, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef]
- Dunlop, A.L.; Knight, A.K.; Satten, G.A.; Cutler, A.J.; Wright, M.L.; Mitchell, R.M.; Read, T.D.; Mulle, J.; Hertzberg, V.S.; Hill, C.C.; et al. Stability of the vaginal, oral, and gut microbiota across pregnancy among African American women: The effect of socioeconomic status and antibiotic exposure. PeerJ 2019, 7, e8004. [Google Scholar] [CrossRef]
- Koren, O.; Goodrich, J.K.; Cullender, T.C.; Spor, A.; Laitinen, K.; Bäckhed, H.K.; Gonzalez, A.; Werner, J.J.; Angenent, L.T.; Knight, R.; et al. Host Remodeling of the Gut Microbiome and Metabolic Changes during Pregnancy. Cell 2012, 150, 470–480. [Google Scholar] [CrossRef]
- Dahl, C.; Stanislawski, M.; Iszatt, N.; Mandal, S.; Lozupone, C.; Clemente, J.C.; Knight, R.; Stigum, H.; Eggesbø, M. Gut microbiome of mothers delivering prematurely shows reduced diversity and lower relative abundance of Bifidobacterium and Streptococcus. PLoS ONE 2017, 12, e0184336. [Google Scholar] [CrossRef]
- Shiozaki, A.; Yoneda, S.; Yoneda, N.; Yonezawa, R.; Matsubayashi, T.; Seo, G.; Saito, S. Intestinal Microbiota is Different in Women with Preterm Birth: Results from Terminal Restriction Fragment Length Polymorphism Analysis. PLoS ONE 2014, 9, e111374. [Google Scholar] [CrossRef]
- Shih, C.-T.; Yeh, Y.-T.; Lin, C.-C.; Yang, L.-Y.; Chiang, C.-P. Akkermansia muciniphila is Negatively Correlated with Hemoglobin A1c in Refractory Diabetes. Microorganisms 2020, 8, 1360. [Google Scholar] [CrossRef]
- Fu, M.-L.; Cheng, L.; Tu, S.-H.; Pan, W.-H. Association between Unhealthful Eating Patterns and Unfavorable Overall School Performance in Children. J. Am. Diet. Assoc. 2007, 107, 1935–1943. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef]
- Grigor’eva, I.N. Gallstone Disease, Obesity and the Firmicutes/Bacteroidetes Ratio as a Possible Biomarker of Gut Dysbiosis. J. Pers. Med. 2020, 11, 13. [Google Scholar] [CrossRef] [PubMed]
- Lima, S.A.M.; El Dib, R.P.; Rodrigues, M.R.K.; Ferraz, G.A.R.; Molina, A.C.; Neto, C.A.P.; De Lima, M.A.F.; Rudge, M.V.C. Is the risk of low birth weight or preterm labor greater when maternal stress is experienced during pregnancy? A systematic review and meta-analysis of cohort studies. PLoS ONE 2018, 13, e0200594. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.; Holzman, C.; Senagore, P.; Wang, J.; Tian, Y.; Rahbar, M.H.; Chung, H. Placental Vascular Pathology Findings and Pathways to Preterm Delivery. Am. J. Epidemiol. 2009, 170, 148–158. [Google Scholar] [CrossRef]
- Donders, G.G.; Van Calsteren, K.; Bellen, G.; Reybrouck, R.; Van den Bosch, T.; Riphagen, I.; Van Lierde, S. Predictive value for preterm birth of abnormal vaginal flora, bacterial vaginosis and aerobic vaginitis during the first trimester of pregnancy. BJOG Int. J. Obstet. Gynaecol. 2009, 116, 1315–1324. [Google Scholar] [CrossRef]
- Sheiner, E.; Mazor-Drey, E.; Levy, A. Asymptomatic bacteriuria during pregnancy. J. Matern. Neonatal Med. 2009, 22, 423–427. [Google Scholar] [CrossRef]
- Smaill, F.M.; Vazquez, J.C. Antibiotics for asymptomatic bacteriuria in pregnancy. Cochrane Database Syst. Rev. 2019, 11, CD000490. [Google Scholar] [CrossRef]
- Elovitz, M.A.; Gajer, P.; Riis, V.; Brown, A.G.; Humphrys, M.S.; Holm, J.B.; Ravel, J. Cervicovaginal microbiota and local immune response modulate the risk of spontaneous preterm delivery. Nat. Commun. 2019, 10, 1305. [Google Scholar] [CrossRef]
- Kumar, M.; Murugesan, S.; Singh, P.; Saadaoui, M.; Elhag, D.A.; Terranegra, A.; Kabeer, B.S.A.; Marr, A.K.; Kino, T.; Brummaier, T.; et al. Vaginal Microbiota and Cytokine Levels Predict Preterm Delivery in Asian Women. Front. Cell. Infect. Microbiol. 2021, 11, 639665. [Google Scholar] [CrossRef]
- Ansari, A.; Bose, S.; You, Y.; Park, S.; Kim, Y. Molecular Mechanism of Microbiota Metabolites in Preterm Birth: Pathological and Therapeutic Insights. Int. J. Mol. Sci. 2021, 22, 8145. [Google Scholar] [CrossRef] [PubMed]
- Thinkhamrop, J.; Hofmeyr, G.J.; Adetoro, O.; Lumbiganon, P.; Ota, E. Antibiotic prophylaxis during the second and third trimester to reduce adverse pregnancy outcomes and morbidity. Cochrane Database Syst. Rev. 2015, 6, CD002250. [Google Scholar] [CrossRef]
- Bröms, G.; Granath, F.; Linder, M.; Stephansson, O.; Elmberg, M.; Kieler, H. Birth Outcomes in Women with Inflammatory Bowel Disease: Effects of disease activity and drug exposure. Inflamm. Bowel Dis. 2014, 20, 1091–1098. [Google Scholar] [CrossRef]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Nageshwar Reddy, D. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787–8803. [Google Scholar] [CrossRef]
- Gershuni, V.; Li, Y.; Elovitz, M.; Li, H.; Wu, G.D.; Compher, C.W. Maternal gut microbiota reflecting poor diet quality is associated with spontaneous preterm birth in a prospective cohort study. Am. J. Clin. Nutr. 2021, 113, 602–611. [Google Scholar] [CrossRef]
- Hiltunen, H.; Collado, M.C.; Ollila, H.; Kolari, T.; Tölkkö, S.; Isolauri, E.; Salminen, S.; Rautava, S. Spontaneous preterm delivery is reflected in both early neonatal and maternal gut microbiota. Pediatr. Res. 2021, 91, 1804–1811. [Google Scholar] [CrossRef] [PubMed]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An Obesity-Associated Gut Microbiome with Increased Capacity for Energy Harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Hanning, I.; Diaz-Sanchez, S. The functionality of the gastrointestinal microbiome in non-human animals. Microbiome 2015, 3, 51. [Google Scholar] [CrossRef]
- Estrada, S.M.; Magann, E.F.; Napolitano, P.G. Actinomyces in Pregnancy: A Review of the Literature. Obstet. Gynecol. Surv. 2017, 72, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, C.M.; Durand, G.; Coutté, M.-B.; Chevrier, C.; Cordier, S. Atrazine in municipal drinking water and risk of low birth weight, preterm delivery, and small-for-gestational-age status. Occup. Environ. Med. 2005, 62, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Stayner, L.T.; Almberg, K.; Jones, R.; Graber, J.; Pedersen, M.; Turyk, M. Atrazine and nitrate in drinking water and the risk of preterm delivery and low birth weight in four Midwestern states. Environ. Res. 2017, 152, 294–303. [Google Scholar] [CrossRef]
- Rinsky, J.L.; Hopenhayn, C.; Golla, V.; Browning, S.; Bush, H.M. Atrazine Exposure in Public Drinking Water and Preterm Birth. Public Health Rep. 2012, 127, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Karahoda, R.; Robles, M.; Marushka, J.; Stranik, J.; Abad, C.; Horackova, H.; Tebbens, J.D.; Vaillancourt, C.; Kacerovsky, M.; Staud, F. Prenatal inflammation as a link between placental expression signature of tryptophan metabolism and preterm birth. Hum. Mol. Genet. 2021, 30, 2053–2067. [Google Scholar] [CrossRef]
- Woting, A.; Blaut, M. The Intestinal Microbiota in Metabolic Disease. Nutrients 2016, 8, 202. [Google Scholar] [CrossRef]
- Tersigni, C.; Neri, C.; D’Ippolito, S.; Garofalo, S.; Martino, C.; Lanzone, A.; Scambia, G.; Di Simone, N. Impact of maternal obesity on the risk of preterm delivery: Insights into pathogenic mechanisms. J. Matern. Fetal Neonatal Med. 2020, 35, 3216–3221. [Google Scholar] [CrossRef] [PubMed]
- Cnattingius, S.; Villamor, E.; Johansson, S.; Bonamy, A.-K.E.; Persson, M.; Wikström, A.-K.; Granath, F. Maternal Obesity and Risk of Preterm Delivery. JAMA 2013, 309, 2362–2370. [Google Scholar] [CrossRef]
- Liang, C.; Lee, P.-F.; Yeh, P.-C. Relationship between Regular Leisure-Time Physical Activity and Underweight and Overweight Status in Taiwanese Young Adults: A Cross-Sectional Study. Int. J. Environ. Res. Public Health 2022, 20, 284. [Google Scholar] [CrossRef]
- Huang, K.-C. Obesity and its related diseases in Taiwan. Obes. Rev. 2008, 9 (Suppl. 1), 32–34. [Google Scholar] [CrossRef]
- World Health Organization. Obesity: Preventing and managing the global epidemic. Report of a WHO consultation. World Health Organ. Tech. Rep. Ser. 2000, 894, 1–253. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Preterm | Term | p |
|---|---|---|---|
| Gestational Age | 34.5 ± 0.5 | 38.9 ± 0.1 | <0.001 |
| Maternal Age | 35.8 ± 0.7 | 35.3 ± 0.3 | 0.608 |
| Maternal Height (cm) | 160.4 ± 1.6 | 160.3 ± 0.4 | 0.925 |
| Maternal Weight (Kg) | 68.6 ± 3.2 | 59.7 ± 0.8 | 0.012 |
| Maternal BMI | 26.6 ± 1.2 | 23.2 ± 0.3 | 0.009 |
| Parity | 2.0 ± 0.2 | 1.9 ± 0.1 | 0.575 |
| Glucose AC | 136.7 ± 7.0 | 133.4 ± 2.3 | 0.655 |
| Gestational DM | 37% | 35% | 0.885 |
| WBC (1000/μL) | 12.4 ± 1.1 | 10.5 ± 0.3 | 0.040 |
| RBC (million/μL) | 4.2 ± 0.1 | 4.4 ± 0.2 | 0.745 |
| Hemoglobin (g/dL) | 11.6 ± 0.3 | 11.9 ± 0.1 | 0.475 |
| MCV (fL) | 86.3 ± 1.9 | 87.2 ± 0.6 | 0.622 |
| Platelets (1000/μL) | 239.5 ± 15.1 | 231.9 ± 4.1 | 0.562 |
| Segment (%) | 75.1 ± 2.1 | 76.5 ± 0.8 | 0.527 |
| Lymphocyte (%) | 17.7 ± 1.9 | 16.6 ± 0.6 | 0.542 |
| Monocyte (%) | 5.3 ± 0.4 | 5.3 ± 0.1 | 0.990 |
| Eosinophil (%) | 1.1 ± 0.2 | 0.8 ± 0.1 | 0.195 |
| Basophil (%) | 0.3 ± 0.1 | 0.3 ± 0.0 | 0.647 |
| Delivery mode (CS/NSD) | 10/12 | 57/132 | 0.280 |
| Neonates data | |||
| Neonatal Sex (M/F) | 10/12 | 96/93 | 0.604 |
| Neonatal Hight | 46.4 ± 0.8 | 49.6 ± 0.2 | 0.001 |
| Neonatal Weight | 2394.8 ± 136.3 | 3152.83 ± 27.9 | <0.001 |
| Hand Circumference | 31.2 ± 0.5 | 33.54 ± 0.1 | <0.001 |
| Chest Circumference | 28.7 ± 0.6 | 32.51 ± 0.1 | <0.001 |
| Abdominal Circumference | 26.5 ± 0.8 | 29.84 ± 0.1 | <0.001 |
| Preterm | Term | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| BMI < 24 | BMI ≥ 24 | BMI < 24 | BMI ≥ 24 | |||||||
| Group | 1 (N = 10) | 2 (N = 12) | 3 (N = 123) | 4 (N = 66) | 1 vs. 2 | 1 vs. 3 | 1 vs. 4 | 2 vs. 3 | 2 vs. 4 | 3 vs. 4 |
| Parameters | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | p-Value | |||||
| Maternal Height | 160.9 ± 3.2 | 159.9 ± 1.4 | 160.7 ± 0.5 | 159.4 ± 0.7 | 0.768 | 0.947 | 0.658 | 0.667 | 0.769 | 0.154 |
| Maternal Weight | 57.1 ± 3.3 | 78.1 ± 3.1 | 54.3 ± 0.6 | 69.7 ± 1.2 | <0.001 | 0.197 | <0.001 | <0.001 | 0.008 | <0.001 |
| Maternal Age | 36.3 ± 1.0 | 35.4 ± 1.0 | 35.0 ± 0.4 | 36.0 ± 0.5 | 0.547 | 0.374 | 0.798 | 0.747 | 0.667 | 0.148 |
| Maternal BMI | 21.9 ± 0.6 | 30.5 ± 1.8 | 21.0 ± 0.2 | 27.4 ± 0.4 | <0.001 | 0.135 | <0.001 | <0.001 | 0.003 | <0.001 |
| Parity | 1.8 ± 0.3 | 2.2 ± 0.4 | 1.8 ± 0.1 | 2.0 ± 0.1 | 0.455 | 0.992 | 0.463 | 0.257 | 0.697 | 0.125 |
| Glucose AC (mg/dl) | 124.0 ± 7.2 | 150.8 ± 10.8 | 128.6 ± 2.4 | 142.3 ± 4.5 | 0.051 | 0.582 | 0.129 | 0.015 | 0.508 | 0.009 |
| Gestational DM (N) | 2 | 7 | 34 | 32 | 0.121 | 0.591 | 0.075 | 0.083 | 0.694 | 0.008 |
| WBC (1000/μL) | 12.6 ± 1.9 | 12.2 ± 1.4 | 10.8 ± 0.4 | 10.0 ± 0.4 | 0.883 | 0.218 | 0.046 | 0.247 | 0.138 | 0.181 |
| RBC (million/μL) | 4.1 ± 0.2 | 4.2 ± 0.2 | 4.2 ± 0.1 | 4.8 ± 0.6 | 0.572 | 0.748 | 0.649 | 0.576 | 0.683 | 0.130 |
| Hemoglgbin (g/dL) | 11.3 ± 0.6 | 11.8 ± 0.3 | 11.9 ± 0.2 | 11.9 ± 0.2 | 0.433 | 0.318 | 0.321 | 0.962 | 0.923 | 0.921 |
| MCV (fL) | 86.4 ± 3.8 | 86.3 ± 2.0 | 87.7 ± 0.7 | 86.4 ± 1.0 | 0.972 | 0.650 | 0.989 | 0.548 | 0.966 | 0.273 |
| Platelets (1000/μL) | 238.8 ± 30.8 | 240.1 ± 14.4 | 225.6 ± 5.1 | 243.7 ± 6.6 | 0.967 | 0.518 | 0.880 | 0.390 | 0.829 | 0.034 |
| Segment (%) | 78.5 ± 3.6 | 72.3 ± 2.5 | 78.4 ± 0.7 | 73.1 ± 1.8 | 0.157 | 0.975 | 0.285 | 0.013 | 0.854 | 0.001 |
| Lymphocyte (%) | 15.0 ± 3.0 | 19.6 ± 2.3 | 15.4 ± 0.6 | 18.8 ± 1.2 | 0.232 | 0.915 | 0.249 | 0.028 | 0.784 | 0.003 |
| Monocyte (%) | 5.1 ± 0.4 | 5.4 ± 0.6 | 5.2 ± 0.2 | 5.4 ± 0.2 | 0.644 | 0.820 | 0.555 | 0.727 | 0.941 | 0.547 |
| Eosinophil (%) | 0.9 ± 0.3 | 1.2 ± 0.2 | 0.7 ± 0.1 | 1.0 ± 0.1 | 0.327 | 0.590 | 0.720 | 0.034 | 0.438 | 0.042 |
| Basophil (%) | 0.2 ± 0.1 | 0.3 ± 0.1 | 0.3 ± 0.0 | 0.3 ± 0.0 | 0.330 | 0.665 | 0.539 | 0.249 | 0.398 | 0.614 |
| Gestational Age | 34.6 ± 0.8 | 34.3 ± 0.5 | 38.8 ± 0.1 | 39.1 ± 0.1 | 0.776 | <0.001 | <0.001 | <0.001 | <0.001 | 0.206 |
| Neonatal Gender | 0.6 ± 0.2 | 0.3 ± 0.1 | 0.5 ± 0.1 | 0.5 ± 0.1 | 0.230 | 0.738 | 0.397 | 0.179 | 0.446 | 0.239 |
| Neonatal Height | 45.6 ± 1.2 | 47.0 ± 1.1 | 49.6 ± 0.2 | 49.7 ± 0.3 | 0.387 | <0.001 | <0.001 | 0.001 | 0.001 | 0.775 |
| Neonatal Weight | 2249.0 ± 154.9 | 2516.3 ± 213.7 | 3098.7 ± 32.9 | 3253.8 ± 49.3 | 0.341 | <0.001 | <0.001 | 0.020 | 0.006 | 0.008 |
| Neonatal HC | 30.5 ± 0.8 | 31.8 ± 0.6 | 33.3 ± 0.1 | 34.0 ± 0.2 | 0.196 | <0.001 | <0.001 | 0.001 | <0.001 | 0.002 * |
| Neonatal CC | 28.1 ± 0.8 | 29.3 ± 0.9 | 32.3 ± 0.2 | 32.8 ± 0.2 | 0.346 | <0.001 | <0.001 | 0.005 | 0.002 | 0.056 |
| Neonatal AC | 25.7 ± 1.1 | 27.2 ± 1.0 | 29.7 ± 0.2 | 30.2 ± 0.2 | 0.347 | 0.007 | 0.003 | 0.038 | 0.018 | 0.096 |
| APGAR (1 min) | 8.4 ± 0.3 | 7.8 ± 0.4 | 8.9 ± 0.1 | 8.7 ± 0.1 | 0.336 | 0.165 | 0.367 | 0.032 | 0.074 | 0.180 |
| APGAR (5 min) | 9.6 ± 0.2 | 9.3 ± 0.3 | 10.0 ± 0.0 | 9.9 ± 0.1 | 0.481 | 0.131 | 0.227 | 0.048 | 0.077 | 0.223 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, H.-R.; Tsai, C.-C.; Chan, J.Y.H.; Lee, W.-C.; Wu, K.L.H.; Tain, Y.-L.; Hsu, T.-Y.; Cheng, H.-H.; Huang, H.-C.; Huang, C.-H.; et al. A Higher Abundance of Actinomyces spp. in the Gut Is Associated with Spontaneous Preterm Birth. Microorganisms 2023, 11, 1171. https://doi.org/10.3390/microorganisms11051171
Yu H-R, Tsai C-C, Chan JYH, Lee W-C, Wu KLH, Tain Y-L, Hsu T-Y, Cheng H-H, Huang H-C, Huang C-H, et al. A Higher Abundance of Actinomyces spp. in the Gut Is Associated with Spontaneous Preterm Birth. Microorganisms. 2023; 11(5):1171. https://doi.org/10.3390/microorganisms11051171
Chicago/Turabian StyleYu, Hong-Ren, Ching-Chang Tsai, Julie Y. H. Chan, Wei-Chia Lee, Kay L. H. Wu, You-Lin Tain, Te-Yao Hsu, Hsin-Hsin Cheng, Hsin-Chun Huang, Cheng-Hsieh Huang, and et al. 2023. "A Higher Abundance of Actinomyces spp. in the Gut Is Associated with Spontaneous Preterm Birth" Microorganisms 11, no. 5: 1171. https://doi.org/10.3390/microorganisms11051171