
The Scenario of Ticks and Tick-Borne Pathogens of Sheep on a Mediterranean Island

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Sheep Farming
2.2. Study Design
2.3. Questionnaire
2.4. Blood Sampling and Tick Collection
2.5. DNA Extraction from Blood and Ticks
2.6. PCR Amplification
2.7. Reverse Line Blot Hybridization (RLB) Assay
2.8. Sequencing
2.9. Statistics
2.10. Ethical Considerations
3. Results
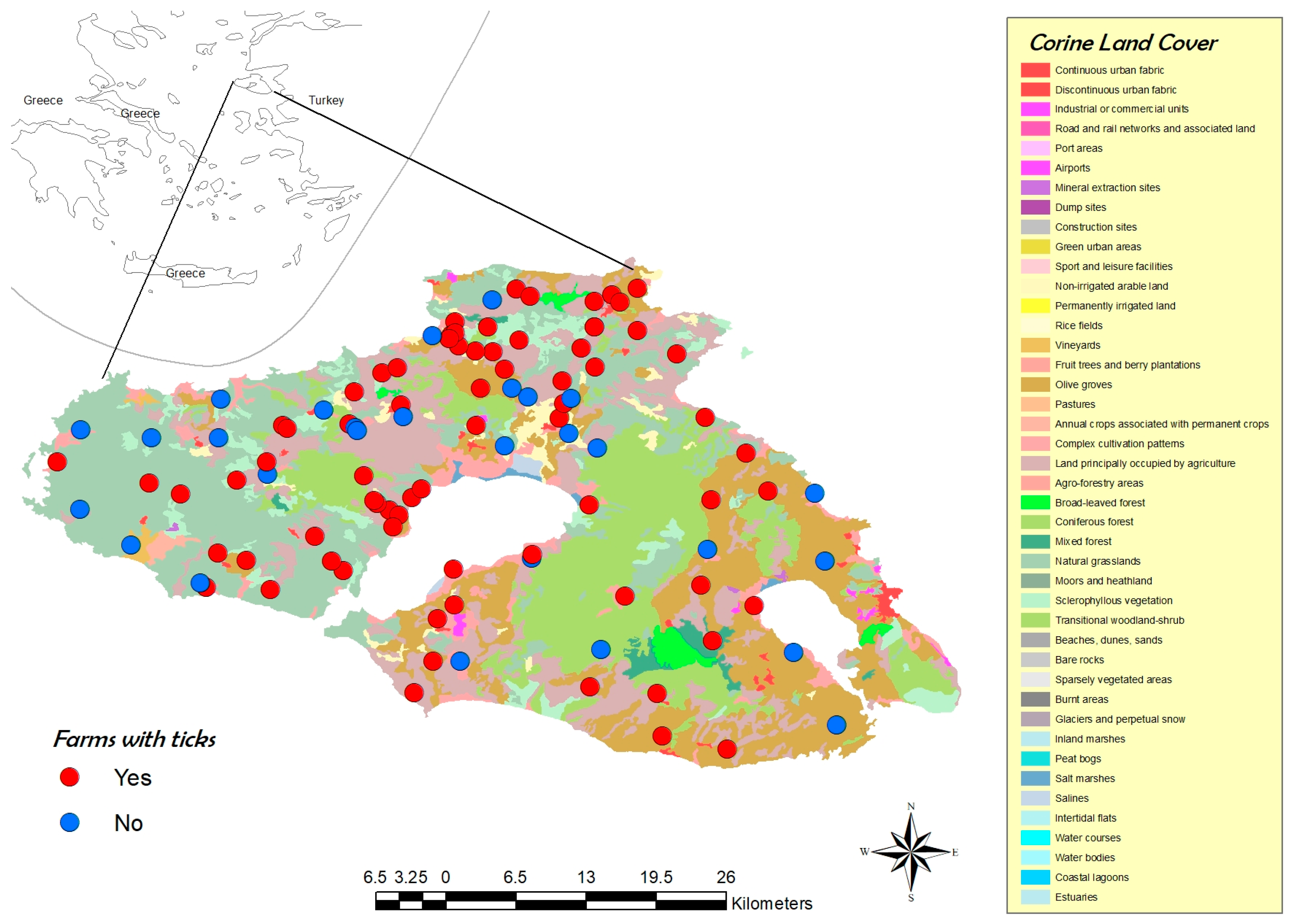
3.1. Farm Description (Questionnaire Data)
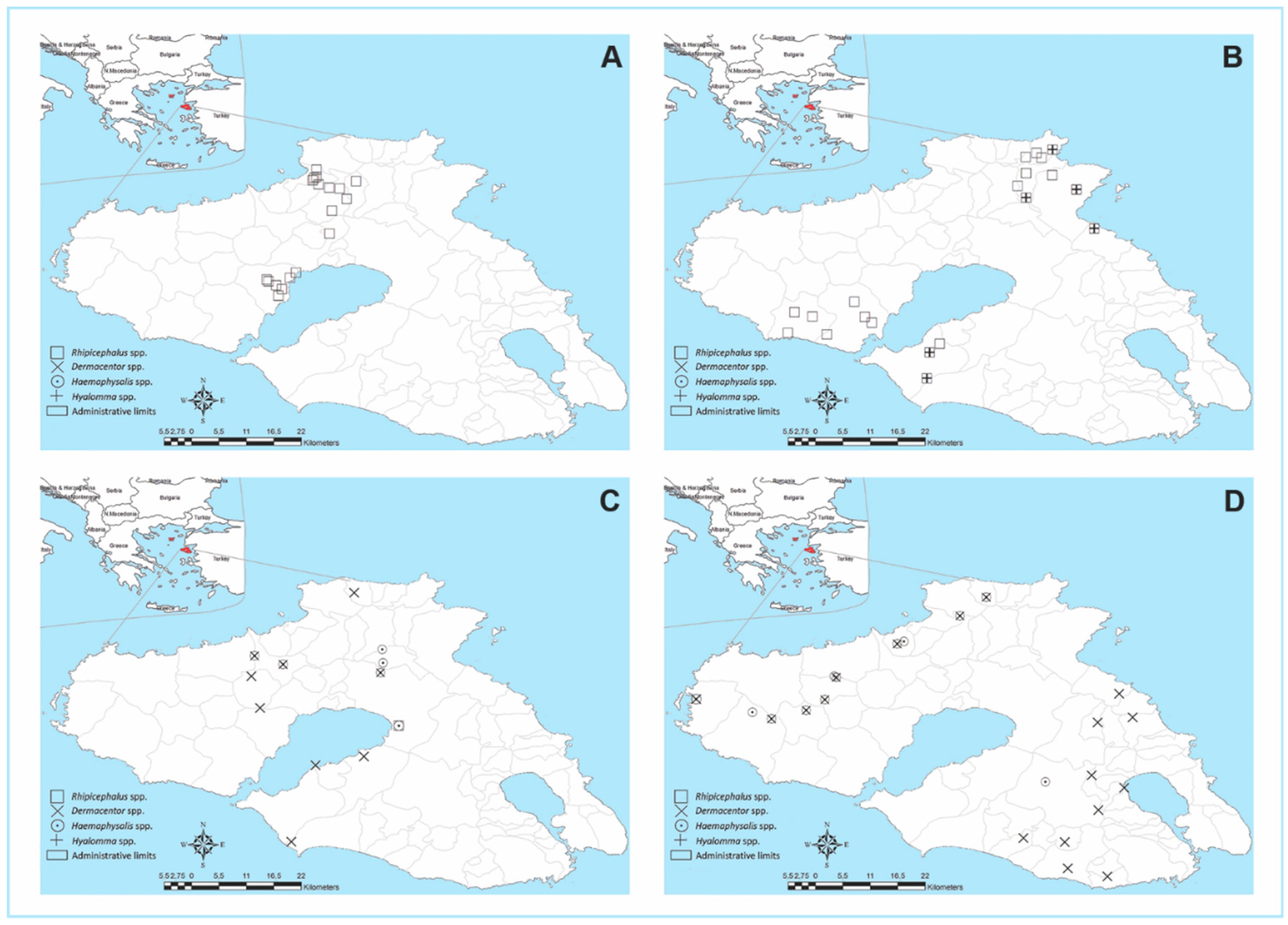
3.2. Tick Collection
3.3. Prevalence of Tick-Borne Pathogens in Sheep
3.4. Detection of TBPs in R. turanicus Ticks by PCR–RLB
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Jongejan, F.; Uilenberg, G. The Global Importance of Ticks. Parasitology 2004, 129, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Defaye, B.; Moutailler, S.; Pasqualini, V.; Quilichini, Y. Distribution of Tick-Borne Pathogens in Domestic Animals and Their Ticks in the Countries of the Mediterranean Basin between 2000 and 2021: ASystematic Review. Microorganisms 2022, 10, 1236. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F. Climate Change, Biodiversity, Ticks and Tick-Borne Diseases: The Butterfly Effect. Int. J. Parasitol. Parasites Wildl. 2015, 4, 452–461. [Google Scholar] [CrossRef] [Green Version]
- Parola, P.; Socolovschi, C.; Jeanjean, L.; Bitam, I.; Fournier, P.E.; Sotto, A.; Labauge, P.; Raoult, D. Warmer Weather Linked to Tick Attack and Emergence of Severe Rickettsioses. PLoS Negl. Trop. Dis. 2008, 2, e338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada-peña, A.A.A.; Venzal, J.M. Climate Niches of Tick Species in the Mediterranean Region: Modeling of Occurrence Data, Distributional Constraints, and Impact of Climate Change. J. Med. Entomol. 2007, 44, 1130–1138. [Google Scholar] [CrossRef]
- Stuen, S. Haemoparasites in Small Ruminants in European Countries: Challenges and Clinical Relevance. Small Rumin. Res. 2016, 142, 22–27. [Google Scholar] [CrossRef]
- Chochlakis, D.; Ioannou, I.; Sharif, L.; Kokkini, S.; Hristophi, N.; Dimitriou, T.; Tselentis, Y.; Psaroulaki, A. Prevalence of Anaplasma Sp. in Goats and Sheep in Cyprus. Vector Borne Zoonotic Dis. 2009, 9, 457–463. [Google Scholar] [CrossRef]
- de la Fuente, J.; Atkinson, M.W.; Naranjo, V.; Fernández de Mera, I.G.; Mangold, A.J.; Keating, K.A.; Kocan, K.M. Sequence Analysis of the Msp4 Gene of Anaplasma Ovis Strains. Vet. Microbiol. 2007, 119, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Torina, A.; Vicente, J.; Alongi, A.; Scimeca, S.; Turlá, R.; Nicosia, S.; Di Marco, V.; Caracappa, S.; de la Fuente, J. Observed Prevalence of Tick-Borne Pathogens in Domestic Animals in Sicily, Italy during 2003–2005. Zoonoses Public Health 2007, 54, 8–15. [Google Scholar] [CrossRef]
- M’ghirbi, Y.; Oporto, B.; Hurtado, A.; Bouattour, A. First Molecular Evidence for the Presence of Anaplasma phagocytophilum in Naturally Infected Small Ruminants in Tunisia, and Confirmation of Anaplasma ovis Endemicity. Pathogens 2022, 11, 315. [Google Scholar] [CrossRef]
- Jouglin, M.; Rispe, C.; Grech-Angelini, S.; Gallois, M.; Malandrin, L. Anaplasma capra in sheep and goats on Corsica Island, France: A European lineage within A. capra clade II? Ticks Tick Borne Dis. 2022, 13, 101934. [Google Scholar] [CrossRef] [PubMed]
- Altay, K.; Erol, U.; Sahin, O.F. The first molecular detection of Anaplasma capra in domestic ruminants in the central part of Turkey, with genetic diversity and genotyping of Anaplasma capra. Trop. Anim. Health Prod. 2022, 54, 129. [Google Scholar] [CrossRef] [PubMed]
- Diakou, A.; Norte, A.C.; Lopes de Carvalho, I.; Núncio, S.; Nováková, M.; Kautman, M.; Alivizatos, H.; Kazantzidis, S.; Sychra, O.; Literák, I. Ticks and Tick-Borne Pathogens in Wild Birds in Greece. Parasitol. Res. 2016, 115, 2011–2016. [Google Scholar] [CrossRef] [PubMed]
- Papa, A.; Chaligiannis, I.; Xanthopoulou, K.; Papaioakim, M.; Papanastasiou, S.; Sotiraki, S. Ticks Parasitizing Humans in Greece. Vector Borne Zoonotic Dis. 2011, 11, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Chaligiannis, I.; Musella, V.; Rinaldi, L.; Cringoli, G.; de la Fuente, J.; Papa, A.; Sotiraki, S. Species Diversity and Spatial Distribution of Ixodid Ticks on Small Ruminants in Greece. Parasitol. Res. 2016, 115, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Papazahariadou, M.G.; Saridomichelakis, M.N.; Koutinas, A.F.; Papadopoulos, E.G.; Leontides, L. Tick Infestation of Dogs in Thessaloniki, Northern Greece. Med. Vet. Entomol. 2003, 17, 110–113. [Google Scholar] [CrossRef] [Green Version]
- Papazahaiadou, M.; Papadopoulos, E.; Himonas, C. Seasonal Activity of Ixodid Ticks on Goats in Northern Greece. Vet. Rec. 1995, 136, 585–588. [Google Scholar] [CrossRef]
- Pavlidou, V.; Gerou, S.; Kahrimanidou, M.; Papa, A. Ticks Infesting Domestic Animals in Northern Greece. Exp. Appl. Acarol. 2008, 45, 195–198. [Google Scholar] [CrossRef]
- Papadopoulos, B.; Morel, P.C.; Aeschlimann, A. Ticks of Domestic Animals in the Macedonia Region of Greece. Vet. Parasitol. 1996, 63, 25–40. [Google Scholar] [CrossRef]
- Hagman, K.; Barboutis, C.; Ehrenborg, C.; Fransson, T.; Jaenson, T.G.T.; Lindgren, P.-E.; Lundkvist, A.; Nyström, F.; Waldenström, J.; Salaneck, E. On the Potential Roles of Ticks and Migrating Birds in the Ecology of West Nile Virus. Infect. Ecol. Epidemiol. 2014, 4, 37. [Google Scholar] [CrossRef] [Green Version]
- Latrofa, M.S.; Angelou, A.; Giannelli, A.; Annoscia, G.; Ravagnan, S.; Dantas-Torres, F.; Capelli, G.; Halos, L.; Beugnet, F.; Papadopoulos, E.; et al. Ticks and Associated Pathogens in Dogs from Greece. Parasites Vectors 2017, 10, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadopoulos, B.; Brossard, M.; Perié, N.M. Piroplasms of Domestic Animals in the Macedonia Region of Greece. 3. Piroplasms of Small Ruminants. Vet. Parasitol. 1996, 63, 67–74. [Google Scholar] [CrossRef]
- Theodoropoulos, G.; Gazouli, M.; Ikonomopoulos, J.A.; Kantzoura, V.; Kominakis, A. Determination of Prevalence and Risk Factors of Infection with Babesia in Small Ruminants from Greece by Polymerase Chain Reaction Amplification. Vet. Parasitol. 2006, 135, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Giadinis, N.D.; Chochlakis, D.; Ioannou, I.; Kritsepi-Konstantinou, M.; Papadopoulos, E.; Psaroulaki, A.; Karatzias, H. Haemorrhagic Diathesis in a Ram with Anaplasma Phagocytophilum Infection. J. Comp. Pathol. 2011, 144, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Giadinis, N.D.; Chochlakis, D.; Kritsepi-Konstantinou, M.; Makridaki, E.; Tselentis, Y.; Kostopoulou, D.; Karatzias, H.; Psaroulaki, A. Haemolytic Disease in Sheep Attributed to a Babesia Lengau-like Organism. Vet. Rec. 2012, 170, 155. [Google Scholar] [CrossRef]
- Efstratiou, A.; Karanis, G.; Karanis, P. Tick-Borne Pathogens and Diseases in Greece. Microorganisms 2021, 9, 1732. [Google Scholar] [CrossRef] [PubMed]
- Baneth, G. Tick-Borne Infections of Animals and Humans: A Common Ground. Int. J. Parasitol. 2014, 44, 591–596. [Google Scholar] [CrossRef]
- Moutailler, S.; Valiente Moro, C.; Vaumourin, E.; Michelet, L.; Tran, F.H.; Devillers, E.; Cosson, J.F.; Gasqui, P.; Van, V.T.; Mavingui, P.; et al. Co-Infection of Ticks: The Rule Rather Than the Exception. PLoS Negl. Trop. Dis. 2016, 10, e0004539. [Google Scholar] [CrossRef] [Green Version]
- Lesvos Biodiversity Observatory—Greece|DEIMS-SDR. Available online: https://deims.org/5a29e045-e8e0-49bc-b60c-fe640389aa8f (accessed on 23 July 2020).
- Vagliano, C. The Continental and Island Migration Route of the Southeast Mediterranean: Problems and Propositions. ICBP Tech. Publ. 1985, 5, 263–269. [Google Scholar]
- Mastranestasis, I.; Kominakis, A.; Hager-Theodorides, A.L.; Ekateriniadou, L.V.; Ligda, C.; Theodorou, K. Associations between Genetic Polymorphisms and Phenotypic Traits in the Lesvos Dairy Sheep. Small Rumin. Res. 2016, 144, 205–210. [Google Scholar] [CrossRef]
- Mastranestasis, I.; Ekateriniadou, L.V.; Ligda, C.; Theodorou, K. Genetic Diversity and Structure of the Lesvos Sheep Breed. Small Rumin. Res. 2015, 130, 54–59. [Google Scholar] [CrossRef]
- Sossidou, E.; Ligda, C.; Mastranestasis, I.; Tsiokos, D. Sheep and Goat Farming in Greece: Implications and Challenges for the Sustainable Development of Less Favoured Areas. Anim. Sci. Biotechnol. 2013, 46, 446–449. [Google Scholar]
- Estrada-Pena, A.; Bouattour, A.; Camica, J.W.A. Ticks of Domestic Animals in the Mediterranean Region: A Guide to Identification of Species; University of Zaragosa: Zaragosa, Spain, 2004; ISBN 84-96214-18-4. [Google Scholar]
- Walker, J.B.; Keirans, J.E.H.I. Genus Rhipicephalus (Acari, Ixodidae). In A Guide to the Brown Ticks of the World; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Giangaspero, A.; Marangi, M.; Papini, R.; Paoletti, B.; Wijnveld, M.; Jongejan, F. Theileria Sp. OT3 and Other Tick-Borne Pathogens in Sheep and Ticks in Italy: Molecular Characterization and Phylogeny. Ticks Tick Borne Dis. 2015, 6, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Bekker, C.P.; de Vos, S.; Taoufik, A.; Sparagano, O.A.; Jongejan, F. Simultaneous Detection of Anaplasma and Ehrlichia Species in Ruminants and Detection of Ehrlichia Ruminantium in Amblyomma Variegatum Ticks by Reverse Line Blot Hybridization. Vet. Microbiol. 2002, 89, 223–238. [Google Scholar] [CrossRef]
- Nijhof, A.M.; Penzhorn, B.L.; Lynen, G.; Mollel, J.O.; Morkel, P.; Bekker, C.P.J.; Jongejan, F. Babesia Bicornis Sp. Nov. and Theileria Bicornis Sp. Nov.: Tick-Borne Parasites Associated with Mortality in the Black Rhinoceros (Diceros Bicornis). J. Clin. Microbiol. 2003, 41, 2249–2254. [Google Scholar] [CrossRef] [Green Version]
- Matjila, P.T.; Penzhorn, B.L.; Bekker, C.P.J.; Nijhof, A.M.; Jongejan, F. Confirmation of Occurrence of Babesia Canis Vogeli in Domestic Dogs in South Africa. Vet. Parasitol. 2004, 122, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Gubbels, J.M.; De Vos, A.P.; Van Der Weide, M.; Viseras, J.; Schouls, L.M.; De Vries, E.; Jongejan, F. Simultaneous Detection of Bovine Theileria and Babesia Species by Reverse Line Blot Hybridization. J. Clin. Microbiol. 1999, 37, 1782–1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zygogiannis, D. Sheep Husbandry, 2nd ed.; Contemporary Education: Thessaloniki, Greece, 2006. [Google Scholar]
- Sonenshine, D.E. Biology of Ticks; Oxford University Press: Oxford, UK, 1991; Volume 2. [Google Scholar]
- Wang, H.H.; Grant, W.E.; Teel, P.D. Simulation of climate-host-parasite-landscape interactions: A spatially explicit model for ticks (Acari: Ixodidae). Ecol. Model. 2012, 243, 42–62. [Google Scholar] [CrossRef]
- Nasirian, H. Detailed new insights about tick infestations in domestic ruminant groups: A global systematic review and meta-analysis. J. Parasit Dis. 2022, 46, 526–601. [Google Scholar] [CrossRef] [PubMed]
- Pegram, R.G.; Wilson, D.D.; Hansen, J.W. Past and present national tick control programs: Why they succeed or fail. Ann. N. Y. Acad. Sci. 2000, 916, 546–554. [Google Scholar] [CrossRef]
- Psaroulaki, A.; Ragiadakou, D.; Kouris, G.; Papadopoulos, B.; Chaniotis, B.; Tselentis, Y. Ticks, Tick-Borne Rickettsiae, and Coxiella Burnetii in the Greek Island of Cephalonia. Ann. N. Y. Acad. Sci. 2006, 1078, 389–399. [Google Scholar] [CrossRef]
- Mumcuoglu, K.Y.; Frish, K.; Sarov, B.; Manor, E.; Gross, E.; Gat, Z.; Galun, R. Ecological Studies on the Brown Dog Tick Rhipicephalus Sanguineus (Acari: Ixodidae) in Southern Israel and Its Relationship to Spotted Fever Group Rickettsiae. J. Med. Entomol. 1993, 30, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Chochlakis, D.; Ioannou, I.; Papadopoulos, B.; Tselentis, Y.; Psaroulaki, A. Rhipicephalus Turanicus: From Low Numbers to Complete Establishment in Cyprus. Its Possible Role as a Bridge-Vector. Parasites Vectors 2014, 7, P11. [Google Scholar] [CrossRef] [Green Version]
- Yesilbag, K.; Aydin, L.; Dincer, E.; Alpay, G.; Girisgin, A.O.; Tuncer, P.; Ozkul, A. Tick Survey and Detection of Crimean-Congo Hemorrhagic Fever Virus in Tick Species from a Non-Endemic Area, South Marmara Region, Turkey. Exp. Appl. Acarol. 2013, 60, 253–261. [Google Scholar] [CrossRef]
- Di Luca, M.; Toma, L.; Bianchi, R.; Quarchioni, E.; Marini, L.; Mancini, F.; Ciervo, A.; Khoury, C. Seasonal Dynamics of Tick Species in an Urban Park of Rome. Ticks Tick Borne Dis. 2013, 4, 513–517. [Google Scholar] [CrossRef] [PubMed]
- Tsatsaris, A.; Chochlakis, D.; Papadopoulos, B.; Petsa, A.; Georgalis, L.; Angelakis, E.; Ioannou, I.; Tselentis, Y.; Psaroulaki, A. Species Composition, Distribution, Ecological Preference and Host Association of Ticks in Cyprus. Exp. Appl. Acarol. 2016, 70, 523–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.-Y.; Zhao, S.-S.; Hornok, S.; Farkas, R.; Guo, L.-P.; Chen, C.-F.; Shao, R.-F.; Lv, J.-Z.; Wang, Y.-Z. Morphological and Molecular Divergence of Rhipicephalus Turanicus Tick from Albania and China. Exp. Appl. Acarol. 2017, 73, 493–499. [Google Scholar] [CrossRef] [Green Version]
- Wei, Q.-Q.; Guo, L.-P.; Wang, A.-D.; Mu, L.-M.; Zhang, K.; Chen, C.-F.; Zhang, W.-J.; Wang, Y.-Z. The First Detection of Rickettsia Aeschlimannii and Rickettsia Massiliae in Rhipicephalus Turanicus Ticks, in Northwest China. Parasites Vectors 2015, 8, 631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallivan, G.J.; Spickett, A.; Heyne, H.; Spickett, A.M.; Horak, I.G. The Dynamics of Questing Ticks Collected for 164 Consecutive Months off the Vegetation of Two Landscape Zones in the Kruger National Park (1988–2002). Part III. The Less Commonly Collected Species. Onderstepoort J. Vet. Res. 2011, 78, 9. [Google Scholar] [CrossRef] [PubMed]
- Orkun, Ö.; Karaer, Z.; Çakmak, A.; Nalbantoğlu, S. Identification of Tick-Borne Pathogens in Ticks Feeding on Humans in Turkey. PLoS Negl. Trop. Dis. 2014, 8, e3067. [Google Scholar] [CrossRef] [PubMed]
- Bakkes, D.K.; Chitimia-Dobler, L.; Matloa, D.; Oosthuysen, M.; Mumcuoglu, K.Y.; Mans, B.J.; Matthee, C.A. Integrative Taxonomy and Species Delimitation of Rhipicephalus Turanicus (Acari: Ixodida: Ixodidae). Int. J. Parasitol. 2020, 50, 577–594. [Google Scholar] [CrossRef] [PubMed]
- Feldman-Muhsam, B.; Borut, S. Some Ecological Observations on Two East-Mediterranean Species of Haemaphysalis Ticks Parasitizing Stock. Vet. Parasitol. 1983, 13, 171–181. [Google Scholar] [CrossRef]
- Guglielmone, A.A.; Robbins, R.G.; Apanaskevich, D.A.; Petney, T.N.; Estrada-Peña, A.; Horak, I.G. The Hard Ticks of the World; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; New York, NY, USA; London, UK, 2014; ISBN 978-94-007-7496-4. [Google Scholar]
- Hosseini-Chegeni, A.; Telmadarraiy, Z.; Salimi, M.; Arzamani, K.; Banafshi, O. A Record of Haemaphysalis Erinacei (Acari: Ixodidae) Collected from Hedgehog and an Identification Key for the Species of Haemaphysalis Occurring in Iran. Persian J. Acarol. 2014, 3, 203–215. [Google Scholar]
- Gherman, C.M.; Mihalca, A.D. A Synoptic Overview of Golden Jackal Parasites Reveals High Diversity of Species. Parasites Vectors 2017, 10, 419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahbari, S.; Nabian, S.; Shayan, P.; Haddadzadeh, H.R. Status of Haemaphysalis Tick Infestation in Domestic Ruminants in Iran. Korean J. Parasitol. 2007, 45, 129–132. [Google Scholar] [CrossRef] [Green Version]
- Mihalca, A.D.; Gherman, C.M.; Magdaş, C.; Dumitrache, M.O.; Györke, A.; Sándor, A.D.; Domşa, C.; Oltean, M.; Mircean, V.; Mărcuţan, D.I.; et al. Ixodes Ricinus Is the Dominant Questing Tick in Forest Habitats in Romania: The Results from a Countrywide Dragging Campaign. Exp. Appl. Acarol. 2012, 58, 175–182. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; Otranto, D. Species Diversity and Abundance of Ticks in Three Habitats in Southern Italy. Ticks Tick Borne Dis. 2013, 4, 251–255. [Google Scholar] [CrossRef]
- Krčmar, S. Hard Ticks (Acari, Ixodidae) of Croatia. Zookeys 2012, 234, 19–57. [Google Scholar] [CrossRef] [Green Version]
- Jiménez, C.; Benito, A.; Arnal, J.L.; Ortín, A.; Gómez, M.; López, A.; Villanueva-Saz, S.; Lacasta, D. Anaplasma ovis in sheep: Experimental infection, vertical transmission and colostral immunity. Small Rumin. Res. 2019, 178, 7–14. [Google Scholar] [CrossRef]
- Renneker, S.; Abdo, J.; Salih, D.E.; Karagenç, T.; Bilgiç, H.; Torina, A.; Oliva, A.G.; Campos, J.; Kullmann, B.; Ahmed, J.; et al. Can Anaplasma ovis in small ruminants be neglected any longer? Transbound. Emerg. Dis. 2013, 60, 105–112. [Google Scholar] [CrossRef]
- Bilgic, H.B.; Bakırcı, S.; Kose, O.; Unlu, A.H.; Hacılarlıoglu, S.; Eren, H.; Weir, W.; Karagenc, T. Prevalence of Tick-Borne Haemoparasites in Small Ruminants in Turkey and Diagnostic Sensitivity of Single-PCR and RLB. Parasites Vectors 2017, 10, 211. [Google Scholar] [CrossRef] [PubMed]
- Aktas, M.; Altay, K.; Dumanli, N. Molecular Detection and Identification of Anaplasma and Ehrlichia Species in Cattle from Turkey. Ticks Tick Borne Dis. 2011, 2, 62–65. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Cao, S.; Sevinc, F.; Sevinc, M.; Ceylan, O.; Ekici, S.; Jirapattharasate, C.; Moumouni, P.F.A.; Liu, M.; Wang, G.; et al. Molecular Detection and Genetic Characterization of Babesia, Theileria and Anaplasma amongst Apparently Healthy Sheep and Goats in the Central Region of Turkey. Ticks Tick Borne Dis. 2017, 8, 246–252. [Google Scholar] [CrossRef]
- Torina, A.; Alongi, A.; Naranjo, V.; Estrada-Peña, A.; Vicente, J.; Scimeca, S.; Marino, A.M.F.; Salina, F.; Caracappa, S.; De La Fuente, J. Prevalence and Genotypes of Anaplasma Species and Habitat Suitability for Ticks in a Mediterranean Ecosystem. Appl. Environ. Microbiol. 2008, 74, 7578–7584. [Google Scholar] [CrossRef] [Green Version]
- Belkahia, H.; Ben Said, M.; El Mabrouk, N.; Saidani, M.; Cherni, C.; Ben Hassen, M.; Bouattour, A.; Messadi, L. Seasonal Dynamics, Spatial Distribution and Genetic Analysis of Anaplasma Species Infecting Small Ruminants from Northern Tunisia. Infect. Genet. Evol. 2017, 54, 66–73. [Google Scholar] [CrossRef]
- Psaroulaki, A.; Chochlakis, D.; Sandalakis, V.; Vranakis, I.; Ioannou, I.; Tselentis, Y. Phylogentic Analysis of Anaplasma Ovis Strains Isolated from Sheep and Goats Using GroEL and Mps4 Genes. Vet. Microbiol. 2009, 138, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Lacasta, D.; Lorenzo, M.; González, J.M.; Ruiz de Arcaute, M.; Benito, A.; Baselga, C.; Milian, M.E.; Lorenzo, N.; Jiménez, C.; Villanueva-Saz, S.; et al. Epidemiological study related to the first outbreak of ovine anaplasmosis in Spain. Animals 2021, 11, 2036. [Google Scholar] [CrossRef] [PubMed]
- Hornok, S.; Elek, V.; de la Fuente, J.; Naranjo, V.; Farkas, R.; Majoros, G.; Földvári, G. First serological and molecular evidence on the endemicity of Anaplasma ovis and A. marginale in Hungary. Vet. Microbiol. 2007, 122, 316–322. [Google Scholar] [CrossRef]
- Torina, A.; Galindo, R.C.; Vicente, J.; Di Marco, V.; Russo, M.; Aronica, V.; Fiasconaro, M.; Scimeca, S.; Alongi, A.; Caracappa, S.; et al. Characterization of Anaplasma phagocytophilum and A. ovis infection in a naturally infected sheep flock with poor health condition. Trop. Anim. Health Prod. 2010, 42, 1327–1331. [Google Scholar] [CrossRef] [Green Version]
- Cabezas-Cruz, A.; Gallois, M.; Fontugne, M.; Allain, E.; Denoual, M.; Moutailler, S.; Devillers, E.; Zientara, S.; Memmi, M.; Chauvin, A. Epidemiology and genetic diversity of Anaplasma ovis in goats in Corsica, France. Parasites Vectors 2019, 12, 3. [Google Scholar] [CrossRef] [Green Version]
- Derdáková, M.; Štefančíková, A.; Špitalská, E.; Tarageľová, V.; Košťálová, T.; Hrkľová, G.; Kybicová, K.; Schánilec, P.; Majláthová, V.; Várady, M.; et al. Emergence and genetic variability of Anaplasma species in small ruminants and ticks from Central Europe. Vet. Microbiol. 2011, 153, 293–298. [Google Scholar] [CrossRef]
- Giadinis, N.; Katsoulos, P.; Chochlakis, D.; Tselentis, Y.; Ntais, P.; Lafi, S.; Karatzias, H.; Psaroulaki, A. Serological Investigation for West Nile Virus, Anaplasma Ovis and Leishmania Infantum in Greek Cattle. Vet. Ital. 2015, 51, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Giadinis, N.D.; Chochlakis, D.; Tselentis, Y.; Petridou, E.; Karatzias, H.; Psaroulaki, A. Hypogalactia in Dairy Sheep Associated with Anaplasma Ovis. In Proceedings of the 2nd Greek Veterinary Congress, Thessaloniki, Greece, 18–20 March 2011; pp. 134–135. [Google Scholar]
- Ciani, E.; Alloggio, I.; Petazzi, F.; Pieragostini, E. Looking for Prognosticators in Ovine Anaplasmosis: Discriminant Analysis of Clinical and Haematological Parameters in Lambs Belonging to Differently Susceptible Breeds Experimentally Infected with Anaplasma Ovis. Acta Vet. Scand. 2013, 55, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, R.; Liu, H.-B.; Jongejan, F.; Jiang, B.-G.; Chang, Q.-C.; Fu, X.; Jiang, J.-F.; Jia, N.; Cao, W.-C. Cultivation of Anaplasma Ovis in the HL-60 Human Promyelocytic Leukemia Cell Line. Emerg. Microbes Infect. 2017, 6, e83. [Google Scholar] [CrossRef] [PubMed]
- Chochlakis, D.; Ioannou, I.; Tselentis, Y.; Psaroulaki, A. Human Anaplasmosis and Anaplasma Ovis Variant. Emerg. Infect. Dis. 2010, 16, 1031–1032. [Google Scholar] [CrossRef]
- Hosseini-Vasoukolaei, N.; Ali Oshaghi, M.; Shayan, P.; Vatandoost, H.; Babamahmoudi, F.; Yaghoobi-Ershadi, M.R.; Telmadarraiy, Z.; Mohtarami, F. Anaplasma Infection in Ticks, Livestock and Human in Ghaemshahr, Mazandaran Province, Iran. J. Arthropod. Borne. Dis. 2014, 8, 204–211. [Google Scholar]
- Matei, I.A.; Estrada-Peña, A.; Cutler, S.J.; Vayssier-Taussat, M.; Varela-Castro, L.; Potkonjak, A.; Zeller, H.; Mihalca, A.D. A review on the eco-epidemiology and clinical management of human granulocytic anaplasmosis and its agent in Europe. Parasites Vectors 2019, 12, 599. [Google Scholar] [CrossRef]
- Stuen, S.; Granquist, E.G.; Silaghi, C. Anaplasma phagocytophilum: A widespread multi-host pathogen with highly adaptive strategies. Front. Cell. Infect. Microbiol. 2013, 3, 31. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.S.; Garyu, J.; Park, J.; Dumler, J.S. Diminished adhesion of Anaplasma phagocytophilum-infected neutrophils to endothelial cells is associated with reduced expression of leukocyte surface selectin. Infect. Immun. 2003, 71, 4586–4594. [Google Scholar] [CrossRef] [Green Version]
- Chochlakis, D.; Psaroulaki, A.; Kokkini, S.; Kostanatis, S.; Arkalati, E.; Karagrannaki, E.; Tsiatis, K.; Tselentis, Y.; Gikas, A. First Evidence of Anaplasma Infection in Crete, Greece. Report of Six Human Cases. Clin. Microbiol. Infect. 2009, 15, 8–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsiodras, S.; Spanakis, N.; Spanakos, G.; Pervanidou, D.; Georgakopoulou, T.; Campos, E.; Petra, T.; Kanellopoulos, P.; Georgiadis, G.; Antalis, E.; et al. Fatal Human Anaplasmosis Associated with Macrophage Activation Syndrome in Greece and the Public Health Response. J. Infect. Public Health 2017, 10, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Papa, A.; Tsioka, K.; Kontana, A.; Papadopoulos, C.; Giadinis, N. Bacterial Pathogens and Endosymbionts in Ticks. Ticks Tick Borne Dis. 2017, 8, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Dugat, T.; Lagrée, A.-C.; Maillard, R.; Boulouis, H.-J.; Haddad, N. Opening the Black Box of Anaplasma Phagocytophilum Diversity: Current Situation and Future Perspectives. Front. Cell. Infect. Microbiol. 2015, 5, 61. [Google Scholar] [CrossRef]
- Li, H.; Zheng, Y.C.; Ma, L.; Jia, N.; Jiang, B.G.; Jiang, R.R.; Huo, Q.B.; Wang, Y.W.; Liu, H.B.; Chu, Y.L.; et al. Human infection with a novel tick-borne Anaplasma species in China: A surveillance study. Lancet Infect. Dis. 2015, 15, 663–670. [Google Scholar] [CrossRef]
- Jouglin, M.; Blanc, B.; de la Cotte, N.; Bastian, S.; Ortiz, K.; Malandrin, L. First detection and molecular identifica- tion of the zoonotic Anaplasma capra in deer in France. PLoS ONE 2019, 14, e0219184. [Google Scholar]
- Remesar, S.; Prieto, A.; García-Dios, D.; López-Lorenzo, G.; Martínez-Calabuig, N.; Díaz-Cao, J.M.; Panadera, R.; López, C.M.; Fernández, G.; Díez-Baños, P.; et al. Diversity of Anaplasma species and importance of mixed infections in roe deer from Spain. Transbound. Emerg. Dis. 2022, 69, e374–e385. [Google Scholar] [CrossRef]
- Ge, Y.; Yin, H.; Rikihisa, Y.; Pan, W.; Yin, H. Molecular Detection of Tick-Borne Rickettsiales in Goats and Sheep from Southeastern China. Vector-Borne Zoonotic Dis. 2016, 16, 309–316. [Google Scholar] [CrossRef]
- Dahmani, M.; Davoust, B.; Rousseau, F.; Raoult, D.; Fenollar, F.; Mediannikov, O. Natural Anaplasmataceae Infection in Rhipicephalus Bursa Ticks Collected from Sheep in the French Basque Country. Ticks Tick Borne Dis. 2017, 8, 18–24. [Google Scholar] [CrossRef]
- Friedhoff, K.T. Tick-Borne Diseases of Sheep and Goats Caused by Babesia, Theileria or Anaplasma spp. Parassitologia 1997, 39, 99–109. [Google Scholar]
- Ben Said, M.; Belkahia, H.; Messadi, L. Anaplasma Spp. in North Africa: A Review on Molecular Epidemiology, Associated Risk Factors and Genetic Characteristics. Ticks Tick Borne Dis. 2018, 9, 543–555. [Google Scholar] [CrossRef]
- Palmer, G.H.; Abbott, J.R.; French, D.M.; McElwain, T.F. Persistence of Anaplasma Ovis Infection and Conservation of the Msp-2 and Msp-3 Multigene Families within the Genus Anaplasma. Infect. Immun. 1998, 66, 6035–6039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocan, K.M.; de la Fuente, J.; Blouin, E.F. Advances toward Understanding the Molecular Biology of the Anaplasma-Tick Interface. Front. Biosci. 2008, 13, 7032–7045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocan, K.M.; Stiller, D.; Goff, W.L.; Claypool, P.L.; Edwards, W.; Ewing, S.A.; McGuire, T.C.; Hair, J.A.; Barron, S.J. Development of Anaplasma Marginale in Male Dermacentor Andersoni Transferred from Parasitemic to Susceptible Cattle. Am. J. Vet. Res. 1992, 53, 499–507. [Google Scholar] [PubMed]
- Yeruham, I.; Hadani, A.; Galker, F.; Rosen, S. The Seasonal Occurrence of Ticks (Acari: Ixodidae) on Sheep and in the Field in the Judean Area of Israel. Exp. Appl. Acarol. 1996, 20, 47–56. [Google Scholar] [CrossRef]
- Yeruham, I.; Hadani, A.; Galker, F. Some Epizootiological and Clinical Aspects of Ovine Babesiosis Caused by Babesia Ovis—A Review. Vet. Parasitol. 1998, 74, 153–163. [Google Scholar] [CrossRef]
- Savini, G.; Conte, A.; Semproni, G.; Scaramozzino, P. Tick-Borne Diseases in Ruminants of Central and Southern Italy: Epidemiology and Case Reports. Parassitologia 1999, 41 (Suppl. S1), 95–100. [Google Scholar]
- Hurtado, A.; Barandika, J.F.; Oporto, B.; Minguijón, E.; Povedano, I.; García-Pérez, A.L. Risks of Suffering Tick-Borne Diseases in Sheep Translocated to a Tick Infested Area: A Laboratory Approach for the Investigation of an Outbreak. Ticks Tick Borne Dis. 2015, 6, 31–37. [Google Scholar] [CrossRef]
- Sevinc, F.; Zhou, M.; Cao, S.; Ceylan, O.; Aydin, M.F.; Sevinc, M.; Xuan, X. Haemoparasitic Agents Associated with Ovine Babesiosis: A Possible Negative Interaction between Babesia Ovis and Theileria Ovis. Vet. Parasitol. 2018, 252, 143–147. [Google Scholar] [CrossRef]
- Aydin, M.F.; Aktas, M.; Dumanli, N. Molecular Identification of Theileria and Babesia in Sheep and Goats in the Black Sea Region in Turkey. Parasitol. Res. 2013, 112, 2817–2824. [Google Scholar] [CrossRef]
- Büscher, G.; Friedhoff, K.T.; El-Allawy, T.A.A. Quantitative Description of the Development of Babesia ovis in Rhipicephalus bursa (Hemolymph, Ovary, Eggs). Parasitol. Res. 1988, 74, 331–339. [Google Scholar] [CrossRef]
- Erster, O.; Roth, A.; Wolkomirsky, R.; Leibovich, B.; Savitzky, I.; Shkap, V. Transmission of Babesia Ovis by Different Rhipicephalus Bursa Developmental Stages and Infected Blood Injection. Ticks Tick Borne Dis. 2016, 7, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Altay, K.; Aktas, M.; Dumanli, N. Detection of Babesia Ovis by PCR in Rhipicephalus Bursa Collected from Naturally Infested Sheep and Goats. Res. Vet. Sci. 2008, 85, 116–119. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| (a) | |||||
|---|---|---|---|---|---|
| Species/Month | May | June | September | October | Whole Period |
| Spring/Summer | Autumn | ||||
| Rhipicephalus turanicus | 95.2% | 84.5% | - | - | 61.3% |
| (n = 589) | (n = 457) | (n = 1046) | |||
| Rhipicephalus bursa | 0.8% | 10.0% | 4.4% | - | 3.6% |
| (n = 5) | (n = 54) | (n = 2) | (n = 61) | ||
| Rhipicephalus sanguineus sensu lato | 4.0% | 0.2% | - | - | 1.5% |
| (n = 25) | (n = 1) | (n = 26) | |||
| Hyalomma marginatum | - | 3.9% | - | - | 1.2% |
| (n = 21) | (n = 21) | ||||
| Hyalomma excavatum | - | 1.5% | - | - | 0.5% |
| (n = 8) | (n = 8) | ||||
| Dermacentor marginatus | - | - | 57.8% | 48.0% | 15.6% |
| (n = 26) | (n = 241) | (n = 267) | |||
| Haemaphysalis parva | - | - | 37.8% | 44.2% | 14.0% |
| (n = 17) | (n = 222) | (n = 239) | |||
| Haemaphysalis punctata | - | - | - | 7.6% | 2.2% |
| (n = 38) | (n = 38) | ||||
| Haemaphysalis sulcata | - | - | - | 0.2% | 0.1% |
| (n = 1) | (n = 1) | ||||
| Total | n = 619 | n = 541 | n = 45 | n = 502 | n = 1707 |
| (b) | |||||
| Infestation rate (sheep) | 96.7% | 89.5% | 16.3% | 48.7% | 55.4% |
| (n = 90) | (n = 105) | (n = 160) | (n = 150) | (n = 505) | |
| 92.8% | 31.9% | ||||
| (n = 195) | (n = 310) | ||||
| Infestation rate (farms) | 100% | 100% | 37.5% | 73.3% | 72.3% |
| (n = 18) | (n = 21) | (n = 32) | (n = 30) | (n = 101) | |
| 100% | 54.8% | ||||
| (n = 39) | (n = 62) | ||||
| Spring/Early Summer | Autumn | Whole Period | |
|---|---|---|---|
| Tick-Borne Pathogens | Number of Positive Samples (Exposure Rate; 95% CI) | ||
| Anaplasma ovis | 183 a (93.8%) | 207 b (66.7%) | 390 (77.2%; 73.4–80.8.) |
| Theileria ovis | 125 a (64.1%) | 186 a (60.0%) | 311 (61.6%; 57.2–65.7) |
| Babesia ovis | 2 a (1.0%) | 0 a (0%) | 2 (0.4%; <0.01–1.5) |
| Anaplasma bovis | 0 a (0%) | 5 a (1.6%) | 5 (1.0%; 0.4–2.4) |
| Anaplasma/Ehrlichia spp. unknown | 2 a (1.0%) | 8 a (2.6%) | 10 (2.0%; 1.0–3.7) |
| Babesia/Theileria spp. unknown | 6 (3.1%) | 3 (1.0%) | 9 (1.8%; 0.9–3.4) |
| Number of samples tested | n = 195 | n = 310 | n = 505 |
| Number of Cases | Prevalence (95% CI) | |
|---|---|---|
| No infection | 76 | 15.1% (12.2–18.4) |
| Single infections | 135 | 26.7% (23.1–30.8) |
| Anaplasma ovis | 108 | 21.4% (18.0–25.2) |
| Theileria ovis | 26 | 5.1% (3.5–7.5) |
| Unknown Babesia/Theileria spp. | 1 | 0.2% (<0.01–1.2) |
| Double infections | 290 | 57.4% (53.3–61.9) |
| Anaplasma ovis + Theileria ovis | 270 | 53.4% (51.1–59.7) |
| Unknown Anaplasma/Ehrlichia spp./Theileria ovis | 10 | 2.0% (1.0–3.7) |
| Anaplasma ovis + Babesia ovis | 2 | 0.4% (0.01–1.5) |
| Anaplasma ovis + unknown Babesia/Theileria | 6 | 1.2% (0.5–2.6) |
| Anaplasma bovis + Theileria ovis | 1 | 0.2% (<0.01–1.2) |
| Anaplasma bovis + unknown Babesia spp. | 1 | 0.2% (<0.01–1.2) |
| Triple infections | 4 | 0.8% (0.2–2.1) |
| Anaplasma ovis + Anaplasma bovis + Theileria ovis | 3 | 0.6% (0.1–18) |
| Anaplasma ovis + Theileria ovis + unknown Babesia spp. | 1 | 0.2% (<0.01–1.2) |
| Total | 505 | 100.0 |
| R. turanicus | |
|---|---|
| Number of ticks tested (females/males) | n = 169 (169/0) |
| Tick-borne pathogens | Number of positive samples (exposure rate; 95% CI) |
| Anaplasma ovis | 24 (14.2%; 9.7–20.0) |
| Anaplasma phagocytophilum | 31 (18.3%; 13.2–24.9) |
| Anaplasma marginale | 42 (24.9%; 18.9–31.9) |
| Ehrlichia chaffensis | 5 (3.0%; 1.1–6.9) |
| Ehrlichia canis | 1 (0.6%; <0.01–3.6) |
| Anaplasma/Ehrlichia spp. unknown | 33 (19.5%; 13.2–24.9) |
| Rickettsia spp. unknown | 68 (40.2%; 33.1–47.8) |
| Babesia major | 4 (2.4%; 0.7–6.1) |
| Theileria ovis | 24 (14.2%; 9.7–20.3) |
| Theileria annulata | 3 (1.8%; 0.4–5.3) |
| Babesia/Theileria spp. unknown | - |
| Babesia spp. unknown | 20 (11.8%; 7.7–17.6) |
| Theileria spp. unknown | 5 (3.0%; 1.1–6.9) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saratsis, A.; Ligda, P.; Aal, F.; Jelicic, M.; Polgar, J.; de Vries, M.; Mastranestasis, I.; Musella, V.; Rinaldi, L.; Jongejan, F.; et al. The Scenario of Ticks and Tick-Borne Pathogens of Sheep on a Mediterranean Island. Microorganisms 2022, 10, 1551. https://doi.org/10.3390/microorganisms10081551
Saratsis A, Ligda P, Aal F, Jelicic M, Polgar J, de Vries M, Mastranestasis I, Musella V, Rinaldi L, Jongejan F, et al. The Scenario of Ticks and Tick-Borne Pathogens of Sheep on a Mediterranean Island. Microorganisms. 2022; 10(8):1551. https://doi.org/10.3390/microorganisms10081551
Chicago/Turabian StyleSaratsis, Anastasios, Panagiota Ligda, Fredie Aal, Mandy Jelicic, Juliette Polgar, Myrthe de Vries, Ioannis Mastranestasis, Vincenzo Musella, Laura Rinaldi, Frans Jongejan, and et al. 2022. "The Scenario of Ticks and Tick-Borne Pathogens of Sheep on a Mediterranean Island" Microorganisms 10, no. 8: 1551. https://doi.org/10.3390/microorganisms10081551