
Isolation and Properties of the Bacterial Strain Janthinobacterium sp. SLB01


Limnological Institute, Siberian Branch, Russian Academy of Sciences, 664033 Irkutsk, Russia
*
Authors to whom correspondence should be addressed.
Microorganisms 2022, 10(5), 1071; https://doi.org/10.3390/microorganisms10051071
Submission received: 31 March 2022
/
Revised: 17 May 2022
/
Accepted: 19 May 2022
/
Published: 23 May 2022
(This article belongs to the Special Issue Blue Microbiology—Aquatic Microbial Resources for Sustainable Life on Earth)
Abstract
:Bacteria of the genus Janthinobacterium are widespread in soils and freshwater ecosystems and belong to the phylum Proteobacteria. The Janthinobacterium sp. SLB01 strain was isolated from diseased freshwater Lubomirskia baicalensis (Pallas, 1776) sponge, and the draft genome was published previously. However, the properties of the SLB01 strain are not known. The aim of the study is to describe some properties of the Janthinobacterium sp. SLB01 strain, isolated from L. baicalensis sponge. The identification of the SLB01 strain was established as Gram-negative, motile, rod-shaped, and psychrotolerant, with growth at 3 and 22 °C. We found that the SLB01 strain has proteolytic, lipolytic, and saccharolytic activity and can use citrates and reduce nitrates. The bacteria Janthinobacterium sp. SLB01 strain can grow, form biofilms, and produce the violet pigment violacein. We identified the pigments violacein and deoxyviolacein by chromatography and mass spectrometry. These metabolites may be of interest to biotechnology in the future. The studied characteristics of the Janthinobacterium sp. SLB01 strain are an important addition to previous studies of the genome of this strain. This study will help us to understand the relationship between the microbial communities of Lake Baikal and sponges.
1. Introduction
The bacteria of the genus Janthinobacterium are widespread and belong to the phylum Proteobacteria of the family Oxalobacteraceae [1,2,3]. Many authors have described the Janthinobacterium isolated from different environments, most commonly from soil and water ecosystems in cold temperatures, including freshwater ecosystems [4,5,6,7]. The Janthinobacterium are a Gram-negative, psychrotolerant, rod-shaped bacteria, motile by means of flagella [3,8]. It is known that Janthinobacterium can produce the purple pigment violacein, which has antibacterial, antiviral, antifungal, antitumor, antiparasitic, and antioxidant activity [9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25].
The Janthinobacterium can form biofilms on abiotic and biotic surfaces in aquatic ecosystems and can colonize fungi and fish skin [11,26,27,28,29,30]. Many researchers have shown that different bacterial species of the genus Janthinobacterium can be pathogens, e.g., Janthinobacterium lividum for the rainbow trout and Janthinobacterium agaricidamnosum, which can cause soft rot disease in the Agaricus bisporus mushroom [27,29,31].
For the first time, we isolated the Janthinobacterium sp. SLB01 strain from the diseased freshwater L. baicalensis sponges and sequenced its complete genome [32]. Since 2011, there has been a problem of the disease and death of freshwater endemic sponges in Lake Baikal [33,34]. Previously, we reported an increased abundance of microorganisms of the phyla Proteobacteria and the family Oxalobacteraceae in diseased L. baicalensis sponges and the infected cell cultures of primmorphs [34,35]. We carried out a genomic analysis comparing the Janthinobacterium sp. SLB01 strain to genomes of other related strains and revealed that it was closely related to the J. lividum MTR strain [20,36]. The aim of the study is to describe some properties of the Janthinobacterium sp. SLB01 strain, isolated from diseased L. baicalensis sponge. This study will expand our understanding of the relationship between the bacteria Janthinobacterium, with their properties, and the endemic freshwater sponges of Lake Baikal.
2. Materials and Methods
2.1. Isolation of the Janthinobacterium sp. SLB01 Strain
The Janthinobacterium sp. SLB01 strain was isolated earlier from a sample of diseased L. baicalensis sponge collected from Lake Baikal, Central Siberia, Russia. The strain was preserved as a 20% (v/v) glycerol suspension at −70 °C. We cultivated the Janthinobacterium sp. SLB01 strain on three different nutrient media for determining bacterial growth. The nutrient media had the following composition: plate count agar (PCA) with 0.5% tryptone, 0.25% yeast extract, 0.1% glucose, 1.5% agar, and final pH 7.2 (HiMedia, Mumbai, India); R 2A agar (R 2A) with 0.05% yeast extract, 0.05% protease peptone, 0.05% tryptone, 0.05% glucose, 0.05% soluble starch, 0.03% sodium pyruvate, 0.03% K2HPO4, 0.005% MgSO4, 1.5 agar, and final pH 7.2 (HiMedia, Mumbai, India), and medium Luria–Bertani (LB) with 1.5% agar, 1% tryptone, 0.05% yeast extract, 0.05% NaCl, and final pH 7.2 (HiMedia, Mumbai, India). The dishes were inoculated in three repetitions and cultivated at a temperature of 22 °C for 5 days, observing the growth of strains daily.
Additionally, the bacteria were cultivated in LB broth at different temperatures of 3, 22, and 30 °C for 10 days, measuring growth daily (OD600) with a spectrophotometer (GBC Scientific Equipment Ltd.—Cintra 20, Melbourne, Australia). Cell morphology was determined by light microscopy (Olympus IX73SC, Olympus Corp., Tokyo, Japan). Gram staining was carried out using the Gram staining kit HiAssortedTM K001-1KT (HiMedia, Mumbai, India).
2.2. Pigment Extraction
The 300 µL liquid culture medium containing a bacterial biomass of the Janthinobacterium sp. SLB01 strain with a pigment was transferred to a 1.5 mL centrifuge tube; 200 µL n-butanol was added, mixed thoroughly, and kept for 15 min in an ultrasonic bath (Sonorex super, Bandelin GmbH & Co., Berlin, Germany). Each sample was centrifuged with the Eppendorf Microcentrifuge 5415R (Merck, Darmstadt, Germany) at 7000 rpm for 20 min. The violacein containing an n-butanol layer was transferred to another tube. The bottom portion containing cell debris and traces of pigment was separated and re-extracted again with 100 µL n-butanol to recover the trace of pigment. The n-butanol layers were pooled together and concentrated through drying in a vacuum concentrator (Concentrator plus, Eppendorf, Hamburg, Germany) at 30 °C. The concentrated n-butanol extract containing violacein and other soluble impurities was extracted in 400 µL chloroform and was passed through a 0.45 μm pore-sized filter. The pigment precipitate was removed from the filter in two portions of 100 μL of methanol for LC analysis and MS spectrometry.
Chromatographic separations were obtained using a microcolumn chromatographic system (Milichrom-A02, Ltd Institute of Chromatography «EcoNova», Novosibirsk, Russia) with a Nucleosil 5-C18 column (75 × 2 mm, 1.7 μm). Gradient elution was carried out using the following solvent systems: phase A (Milli-Q water + 0.1% formic acid) and phase B (acetonitrile + 0.1% formic acid). The solvent gradient programming was 5% B in 8 min, and then from 5% to 100% B at a flow rate of 100 μL min−1, returning to the initial conditions in 2 min. The run time was 23 min per sample. The violacein methanol solution was characterized using UV–vis and electron spray ionization coupled with mass spectrometry (Ultraflex, Bruker Daltonik GmbH, Bremen, Germany).
2.3. Violacein Production
The Janthinobacterium sp. SLB01 strain was grown in 12-well plates (TPP Techno Plastic Products AG, Trasadingen, Switzerland) with LB broth at temperatures of 3, 22, and 30 °C to assess the accumulation of violacein. Violacein was extracted according to the previously described technique [11]. Bacterial cells from a single well were harvested by centrifugation at 16,000× g for 15 min every 24 h. The cells were lysed with 10% sodium dodecyl sulfate (v/v) (AppliChem, Darmstadt, Germany), and incubated at room temperature for 5 min. Then, water-saturated butanol (1:2) was added and shaken. The upper phase, containing violacein, was separated from the aqueous phase by centrifugation at 16,000× g for 10 min. The extracted violacein was quantified using a spectrophotometer (OD585) (GBC Scientific Equipment Ltd.—Cintra 20, Melbourne, Australia).
2.4. Biofilm Detection
We estimated the biofilm formation of the Janthinobacterium sp. SLB01 strain grown in 12-well plates (TPP Techno Plastic Products AG, Trasadingen, Switzerland) in LB broth at temperatures of 3, 22, and 30 °C. The biofilm amount was determined at regular time intervals according to a previously described method [11]. The wells of the plate were carefully emptied and washed twice with sterile 0.01 M phosphate-buffered saline (PBS) (Sigma-Aldrich, St. Louis, MO, USA) every 24 h. Then, 5 mL of an aqueous solution of 0.1% safranin (Sigma-Aldrich, St. Louis, MO, USA) was added to each well and incubated at room temperature for 20 min. The dye was removed with 0.01 M PBS buffer. Then, the plates were inverted onto filter paper (Macherey-Nagel, Düren, Germany) and dried for 3 h. Safranin was extracted with a mixture of ethanol 96% and acetone (80:20 v/v). The biofilm formation was quantified by measuring the optical density (OD484) of each sample using a spectrophotometer (GBC Scientific Equipment Ltd.—Cintra 20, Melbourne, Australia).
2.5. Biochemical Characterization of the Strain
The Janthinobacterium sp. SLB01 strain was analyzed using Hi Assorted Biochemical test kits for Gram-negative bacteria (HiMedia, Mumbai, India). We conducted biochemical tests for the determination of citrate utilization, L- lysine, L-ornithine, L-arginine, urease, phenylalanine deamination, nitrate reduction, hydrogen sulfide (H2S) production, glucose, lactose, arabinose, mannose, sucrose, adonitol, D-sorbitol, D-mannitol, and inositol. In addition, the SLB01 strain was tested using spirit blue agar, gelatin agar, and motility test medium (HiMedia, Mumbai, India). The test Janthinobacterium sp. SLB01 strain was grown in LB broth at 22 °C for 48 h and resuspended in the appropriate test media according to the manufacturer’s instructions, and 50 μL of the prepared inoculums was added to each well. The readings were performed after 48 h of incubation at 22 °C. The results were interpreted as per the standards given in the result chart.
2.6. Antibiotics Sensitivity Tests of the Strain
The Janthinobacterium sp. SLB01 strain was cultured on Mueller–Hinton agar (HiMedia, Mumbai, India) at 22 °C for 48 h and was tested with antimicrobial sensitivity discs of 20 antibiotics (HiMedia, Mumbai, India). The medium plates were prepared as per the manufacturer’s instructions. The following antibiotics were used in the study: ampicillin, ceftriaxone, imipenem, nalidixic acid, levofloxacin, lomefloxacin, norfloxacin, ofloxacin, ciprofloxacin, amikacin, gentalyn, gentamicin, kanamycin, netilmycin, tobramycin, doxycycline, tetracycline, chloramphenicol, erythromycin, and rifampicin. The results were analyzed as described in the manufacturer’s instructions.
2.7. Statistical Analysis
All the experiments were performed at least three times. The data were reported as means ± standard deviation (SD). The statistical analysis was carried out (single-factor (ANOVA) followed by Tukey’s multiple range test) using SPSS.16 software. Differences in mean values were considered significant at p < 0.05. All charts were built using Microsoft Office Professional Plus 2016 (Microsoft Corporation, Redmond, WA, USA).
3. Results
3.1. Isolation of the Bacterial Strains and Culture Conditions
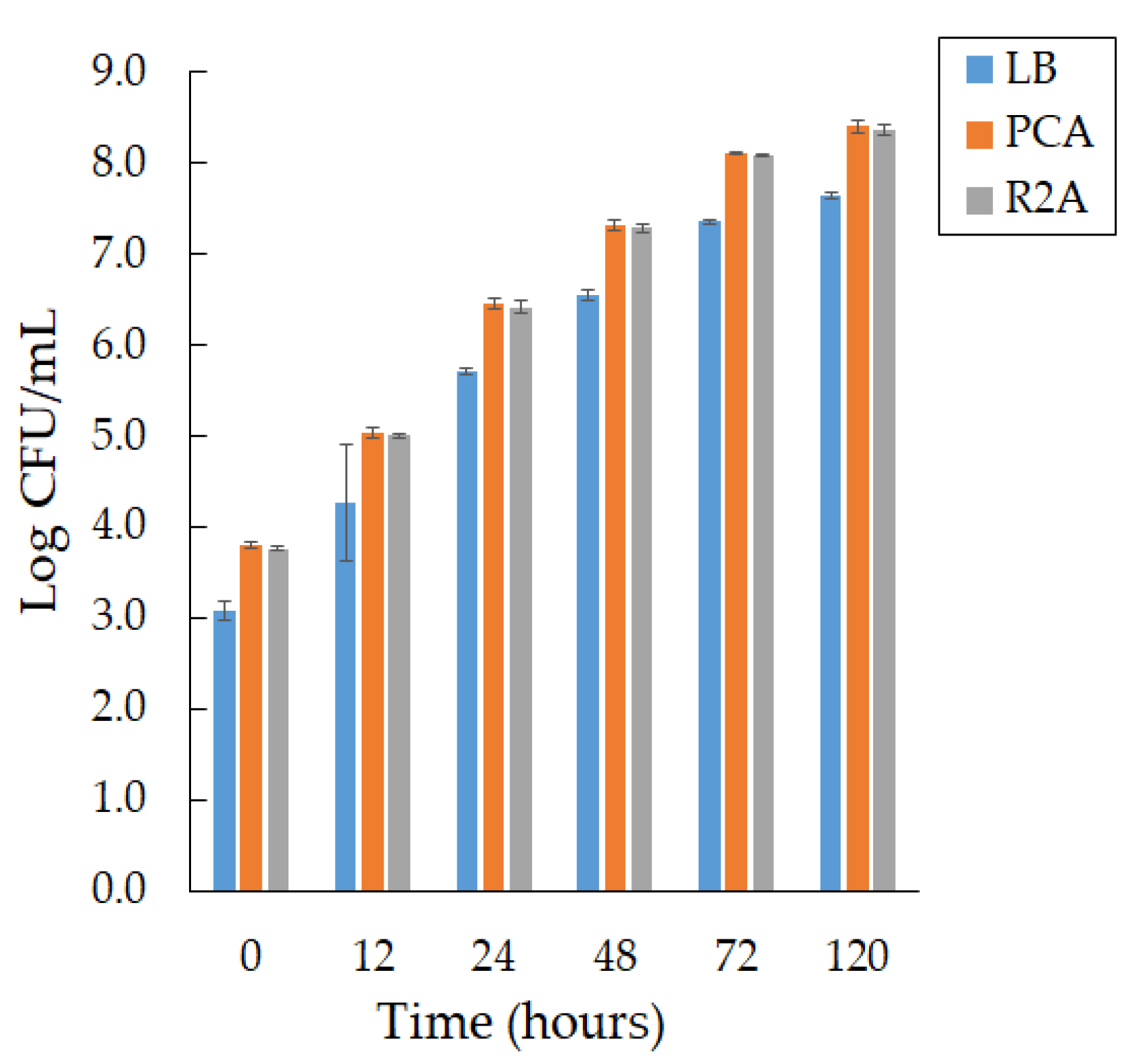
We observed active growth during the formation of smooth, convex colonies with a purple pigment on days 2 and 3 of cultivation at a temperature of 22 °C on different media, PCA and R 2A, and a slight difference in bacterial growth in LB agar (Figure 1).
We found that the growth rate of bacteria of the Janthinobacterium sp. SLB01 strain does not depend on the composition of the medium; however, PCA medium is preferred.
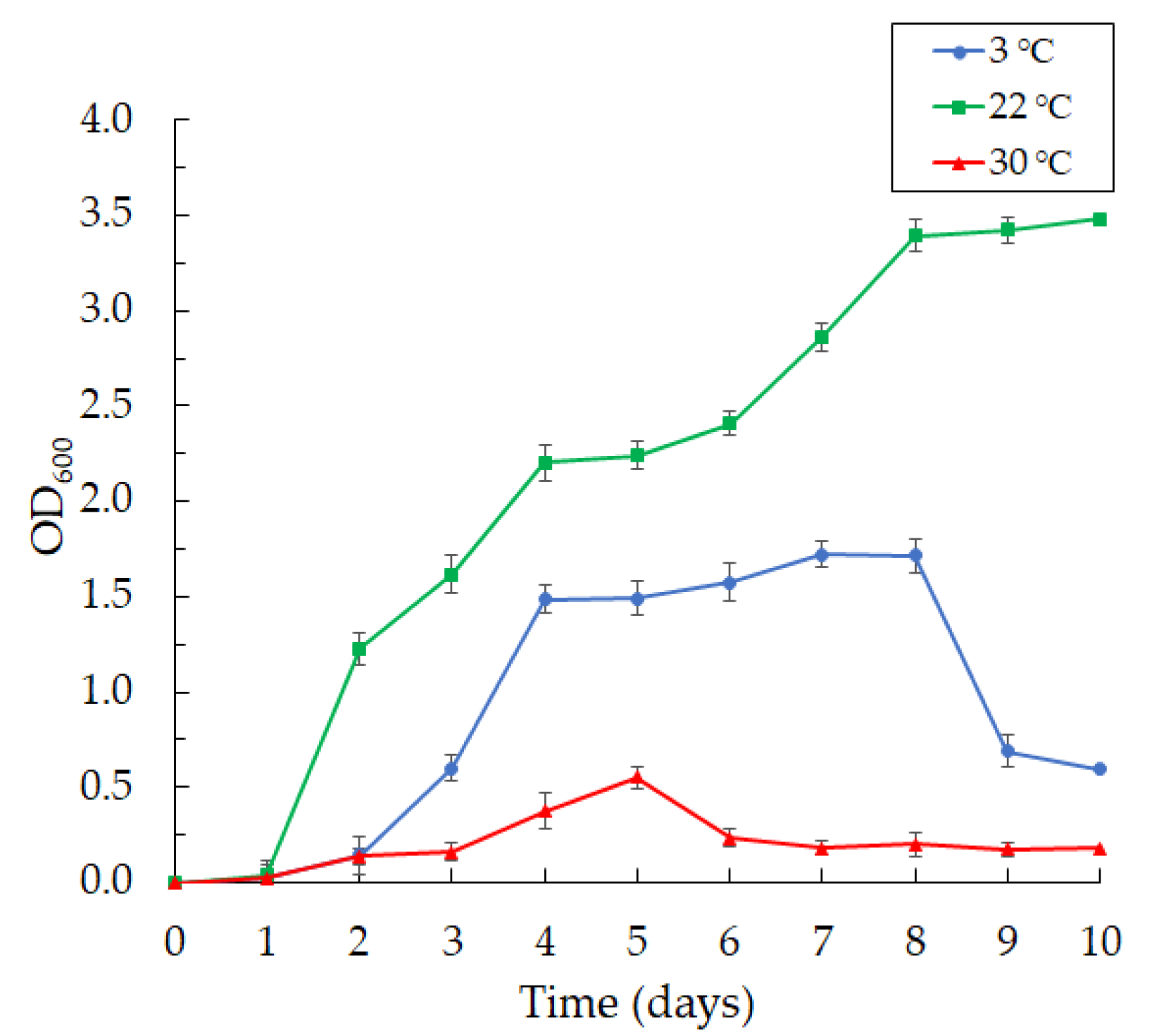
The bacteria were aerobic and psychrotolerant; they grew at 3 and 22 °C and did not grow well at 30 °C (Figure 2).
We found that the bacteria grew intensively on day 2 at 3 °C, reaching maximum growth by day 8 of cultivation, with a subsequent fall in growth.
The bacteria grew rapidly and produced violacein at a temperature of 22 °C. The bacteria of the Janthinobacterium sp. SLB01 strain were Gram-negative and did not form spores. They were rod-shaped with rounded ends and had a length of 1.0 to 5.0 µm and a width of 0.3 to 1.0 µm, with twitching motility. Moreover, we observed a change in the morphology of the bacteria during cultivation at different temperatures and media (Figure 3).
The bacterial cultures grew well at 22 °C in all the used nutrient media. However, we observed the formation of long cells in the Janthinobacterium sp. SLB01 strain at 30 °C.
3.2. Characterization of Violacein
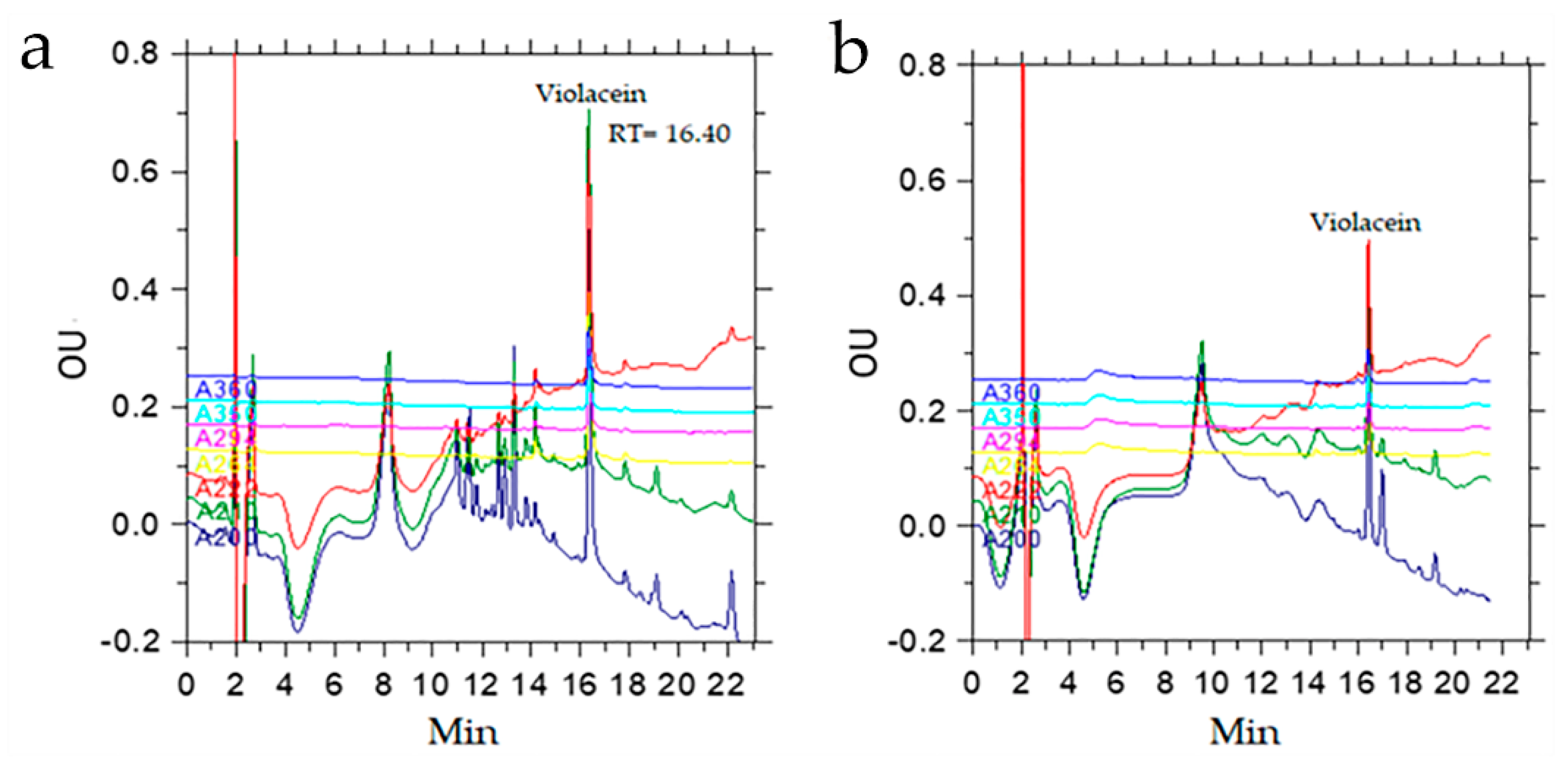
An extraction of 500 µL of the culture medium containing the biomass of the SLB01 strain with 300 µL of n-butanol allowed us to isolate more than 95% of the pigment. With repeated extraction, an additional 4% of the pigment can be isolated, as calculated from the ratio of the chromatographic peak areas. HPLC analysis data indicated that the butanol extract contained a peak with a retention time of 16.40 min, which was the bulk of the extract (53%), and the extract contained other substances with a lower retention time (Figure 4).
The presence of violacein and deoxyviolacein in the crude extract was confirmed by MALDY TOF MS analysis. The extract contained peaks of pseudomolecular ions with m/z 344 [M + H]+ and 328 [M + H]+, which belonged to violacein and deoxyviolacein, respectively. In addition, there was an ion with m/z 360 [M + H]+, corresponding to oxyviolacein [37].
3.3. Production of Violacein
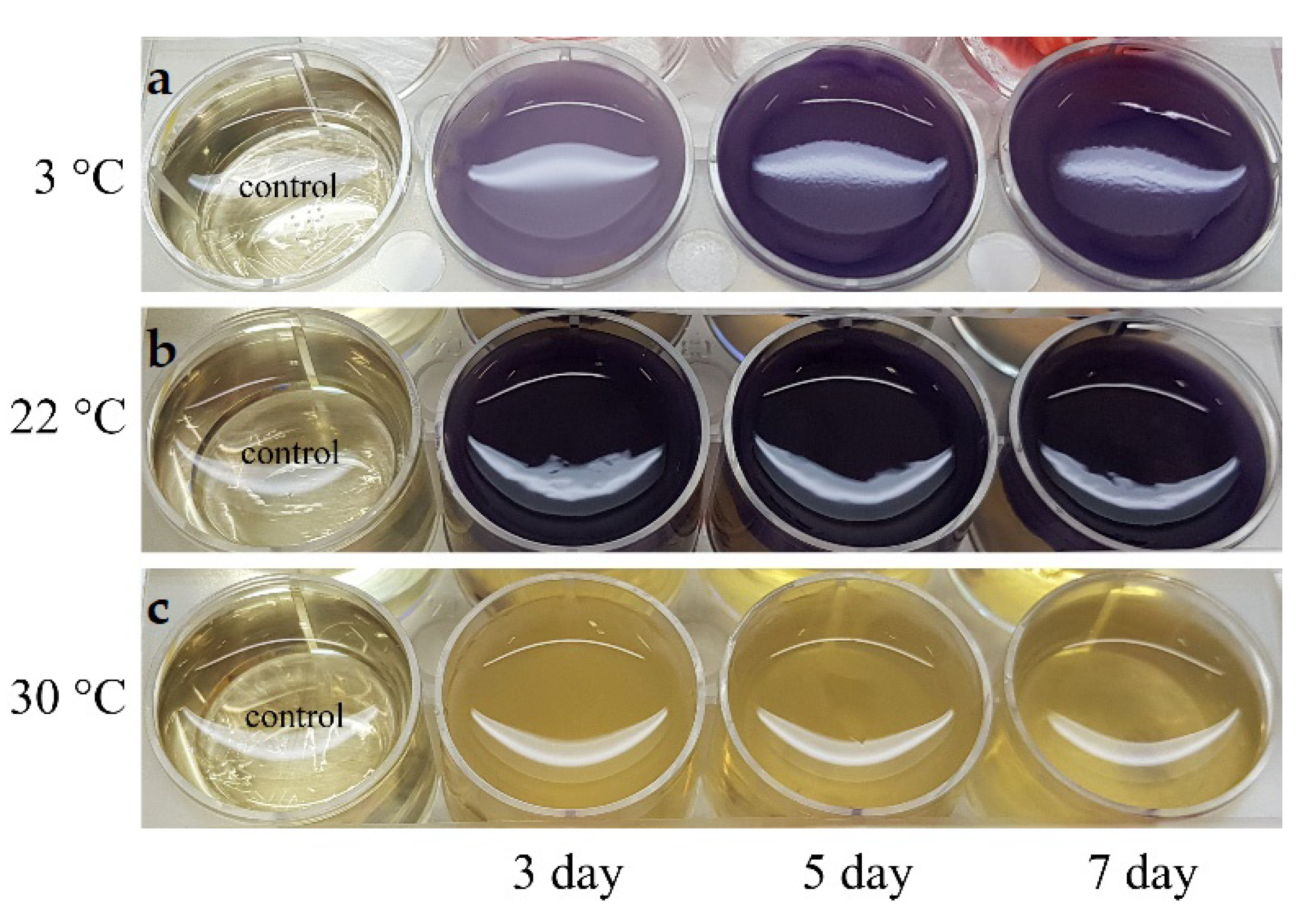
The cultivation of the Janthinobacterium sp. SLB01 strain in LB broth revealed the effect of different temperatures on the production of violacein (Figure 5).
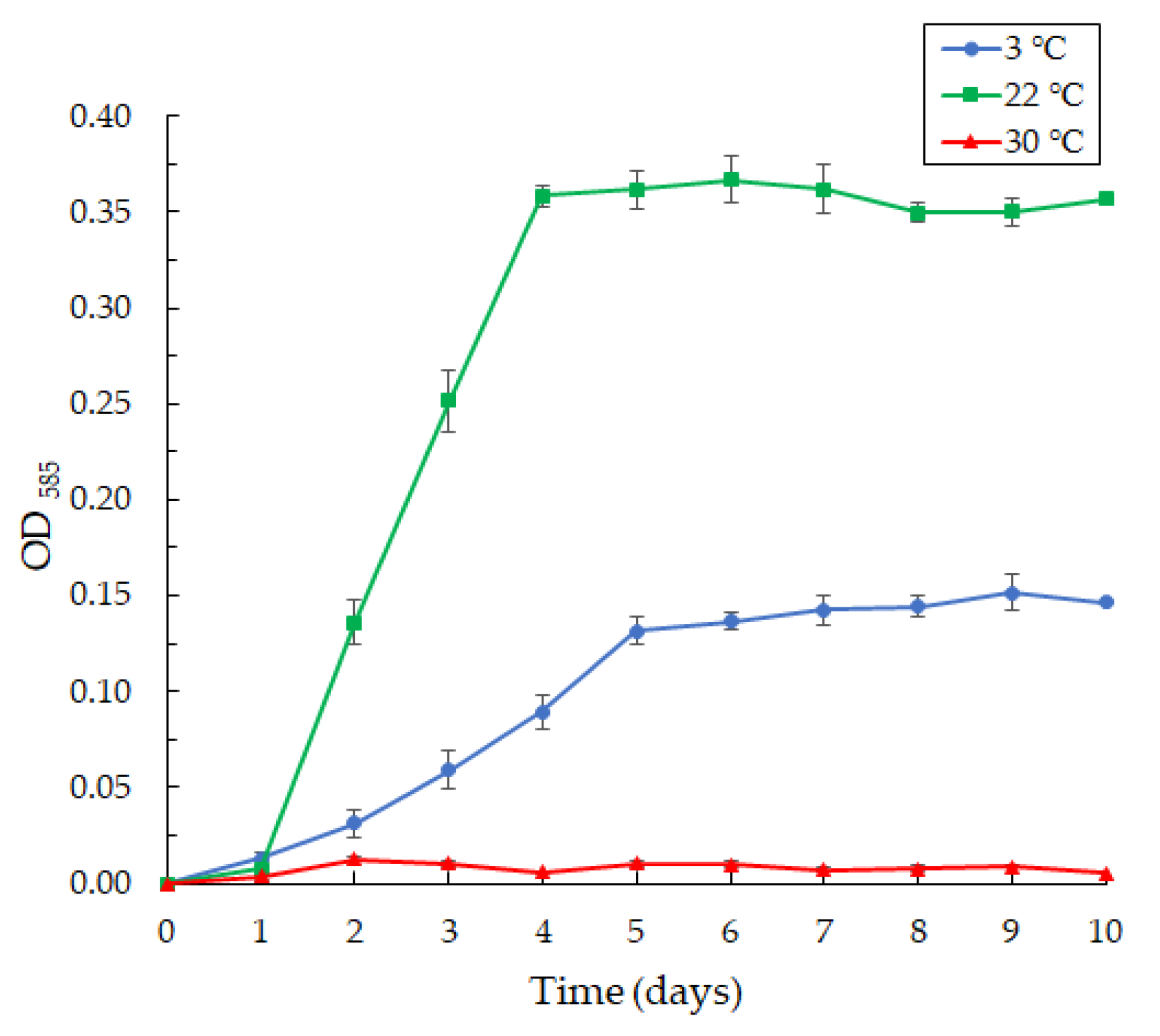
The production of violacein was noted on the second day of cultivation, with intensive formation on the fifth day (Figure 6).
The Janthinobacterium sp. SLB01 strain was isolated from a diseased Baikal sponge collected earlier at a depth of 9–10 m and water temperature of 3–4 °C [32]. The SLB01 strain grew and produced violacein at 3 and 22 °C (Figure 6). However, the growth of the strain and violacein production was suppressed at 30 °C. We found that the SLB01 strain grew rapidly during the first five to ten days, with the simultaneous formation of violacein at 22 °C, reaching a maximum density (OD585) of 3.682; at a temperature of 3 °C, the concentration of violacein (OD585) reached 1.517.
3.4. Biofilm Formation
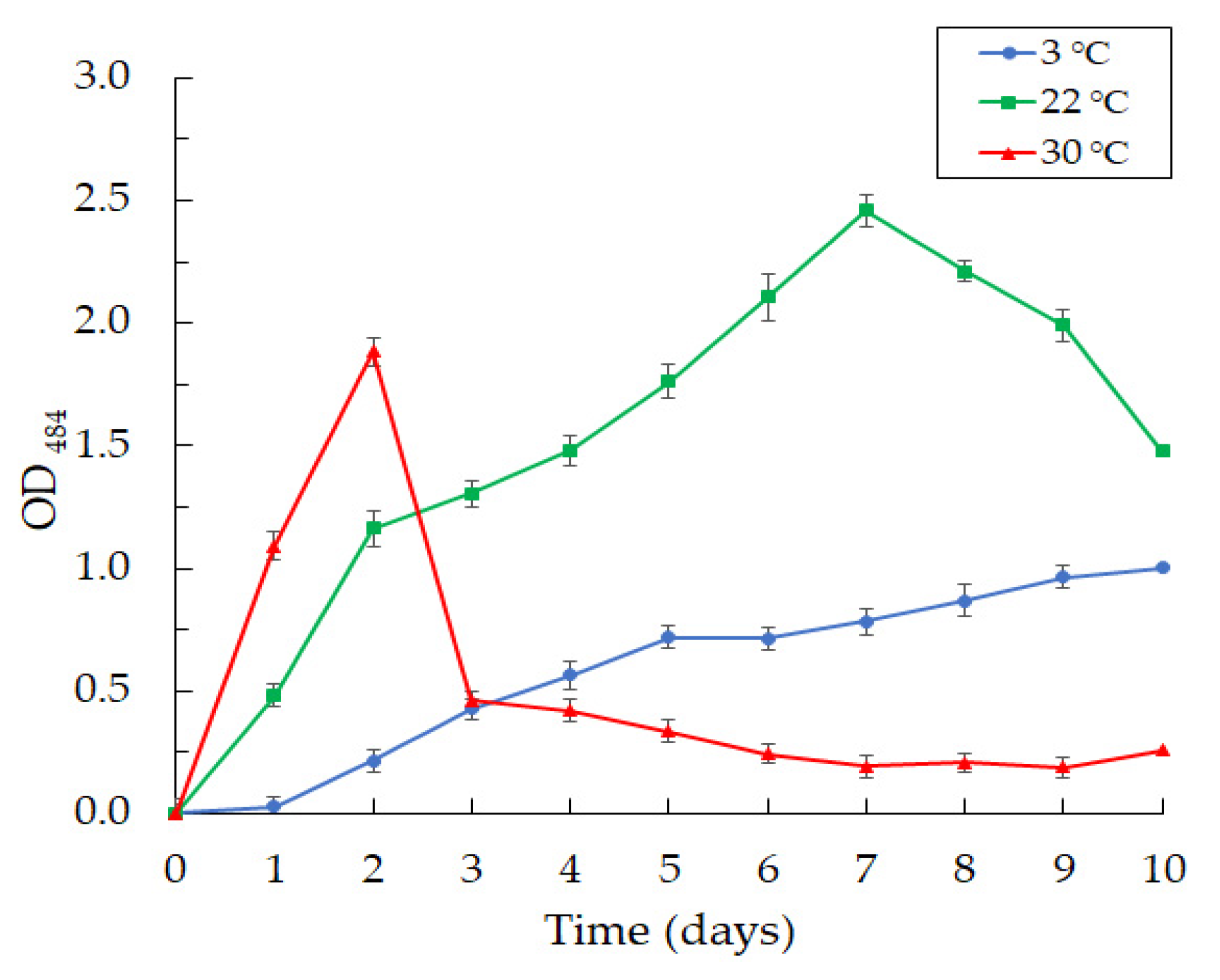
Along with violacein production, we observed an intensive formation of biofilms at the cultivation of the Janthinobacterium sp. SLB01 strain at 3, 22, and 30 °C in LB broth medium (Figure 7).
We observed the intensive formation of dense biofilms in the first 24 h of cultivation at 22 and 30 °C. The highest OD484 index reached 1.303 at 22 °C and 1.937 at 30 °C for two days of cultivation. Then, the achieved OD484 index decreased sharply to 0.334 on the fifth day and to 0.256 on the tenth day of SLB01 cultivation at the temperature of 30 °C. The structure of the biofilms became loose and poorly attached to the walls of the plates. Interestingly, the indicators grew, and the OD484 index reached 2.560 at 22 °C for seven days and remained stable; the biofilms were dense. A completely different picture was observed at 3 °C. Biofilm formation gradually increased and reached the highest OD484 of 1.051 at 3 °C after 10 days of cultivation.
3.5. Biochemical Analysis
The biochemical characteristics of the Janthinobacterium sp. SLB01 strain are shown in Table 1.
Positive tests for the Janthinobacterium sp. SLB01 strain were as follows: citrate utilization, L-lysine, L-ornithine, L-arginine, nitrate reduction, lipolytic activity, glucose, sucrose, mannose, lactose, inositol, casein, and gelatin. Additionally, the test motility showed a positive reaction. Tests negative for the Janthinobacterium sp. SLB01 strain were urease production, phenylalanine deamination, H2S production, adonitol, arabinose, D-sorbitol, and D-mannitol.
3.6. Antibiotic Sensitivity Tests of the Strain
The antibiotic susceptibility test for studying the Janthinobacterium sp. SLB01 strain showed high resistance to multiple tested antibiotics (Table 2).
The SLB01 strain was resistant to ampicillin, imipenem, nalidixic acid, levofloxacin, norfloxacin, gentalyn, netilmycin, chloramphenicol, and erythromycin but susceptible to gentamicin, kanamycin, tobramycin, doxycycline, tetracycline, and rifampicin.
4. Discussion
Lake Baikal, located in the rift zone in the center of the Asian continent, is a cold-water lake [38]. The temperature of the water in the habitats of the Baikal sponges is within 3–4 °C. The mass diseases and death of freshwater sponges in Lake Baikal were observed in 2011 [33]. Subsequently, the disease of the Baikal sponges spread to the entire littoral of the lake. The study of the microbiomes confirmed the shift in the compositions of symbiotic bacteria in freshwater sponges [34,35]. Earlier, we showed that bacteria of the families Oxalobacteraceae and Flavobacteraceae increased in relative abundance in experiments on the infection of the cell culture of primmorphs [35].
The diseases and mass death of sea sponges are also observed in the seas and oceans and are accompanied by a shift in the composition of symbiotic microbial communities [39,40,41]. Many researchers associate sea sponge diseases with changes in the composition of symbionts and the appearance of opportunistic infection resulting from changes in water temperature [42,43].
In this study, we have described the properties of the Janthinobacterium sp. SLB01 strain (Oxalobacteraceae) isolated from diseased L. baicalensis sponges. We have found that the SLB01 strain has proteolytic, lipolytic, and saccharolytic activities and can use citrate, lysine, ornithine, and arginine and reduce nitrates. The strain can use polysaccharides such as starch as a source of carbon; this was evidenced by the presence of amylase in the SLB01 genome [36]. The test for nitrate reductase was positive. Previously, it was shown that an increase in NO3+ can lead to a significant decrease in biofilm biomass in a nutrient-poor environment [44]. Moreover, the urease test in the SLB01 strain was negative, although its genome encodes a set of genes required for urea degradation [32]. Interestingly, the genomes of many Janthinobacterium spp. contain urease synthesis operons, but urease activity is not always detected. For example, in Janthinobacterium lividum ERGS5:01 and Janthinobacterium violaceinigrum, the test results were positive for urease [45,46], but they were negative for Janthinobacterium tructae and Janthinobacterium psychrotolerans [7,31].
We found that temperature affects violacein production and biofilm formation in the SLB01 strain. The concentration of violacein increases linearly on the fifth day at the temperature of 22 °C and remains constant, while the biofilm concentration continues to increase until the seventh day, followed by a gradual decrease. However, we revealed a sharp increase in biofilm formation at the temperature of 30 °C on the second day, with a subsequent fall. The reason for this phenomenon is not clear. The temperature of cultivation at 30 °C is critical for the survival of the strain, so neither bacterial biomass nor violacein increased during the cultivation. Bacteria probably begin to form a large amount of biofilm at this temperature to protect against aggressive environmental conditions [11].
At the same time, the production of violacein and biofilm constantly increases from the second to the tenth day at a temperature of 3 °C as the isolated strain is psychrophilic; the microorganisms and sponges live in the low temperatures of the Baikal water at 3–4 °C [35,38,47]. Moreover, violacein, as well as biofilms, represents a response to environmental stresses and is a key factor in the survival of these microorganisms [11]. It is known that biofilm development and violacein production are regulated by quorum-sensing systems [48,49]. We found that at 30 °C of cultivation, biofilm formation occurs without the production of violacein. The formation of biofilms in the absence of violacein production may indicate the complex nature of the induction.
In addition, the initial HPLC analysis allowed us to identify violacein and deoxyviolacein in the n-butanol extract of the Janthinobacterium SLB01 biomass, which constituted the bulk of the extract (53%), calculated from the area ratio of the peaks at 210 nm; the amount of deoxyviolacein was about 3.5% relative to the amount of violacein. MALDY TOF MS analysis confirmed the presence of violacein and deoxyviolacein in the crude extract. The extract contained peaks of pseudomolecular ions with m/z 344 [M + H]+ and 328 [M + H]+, which belong to violacein and deoxyviolacein, respectively.
Extraction of the concentrated butanol extract with chloroform allowed us to remove most of the unidentified compounds and substantially purify the desired violacein, but further experiments are needed to optimize the method, including optimization of the n-butanol/chloroform ratio and extraction temperature and extinction coefficient determination, as well as to accurately determine the compounds extractable with chloroform. Violacein can be easily isolated directly from the culture medium without the stages of destruction of bacterial cells and centrifugation, and it can be purified from the main impurities by extraction with chloroform. Among the substances that have a shorter retention time during chromatography, there may be metabolites of interest for biotechnology, for example, the tropodithietic acid antibiotic, the synthesis operon of which was identified in the analysis of the complete genome of the Janthinobacterium sp. SLB01 strain [36].
The results demonstrated that the SLB01 strain was resistant to multiple antibiotics. At the same time, there was sensitivity to various antibiotics such as gentamicin, kanamycin, tobramycin, doxycycline, tetracycline, and rifampicin. The analyzed strain is important in the realization of biological properties and adaptation to the environment.
Thus, in the present study, we have characterized in more detail the bacterial strain of Janthinobacterium sp. SLB01 from L. baicalensis sponge. Characteristics of the SLB01 strain make its properties an interesting addition to previous studies of the genome. We found a combination of twitching motility and biofilm formation that can be seen as factors facilitating Baikal sponge surface colonization. Moreover, violacein production can lead to a shift in the bacterial community of sponges and deterioration in their health. We expect that this study will contribute to the understanding of the relationships in microbial communities of sponges that are an important factor of the ecological situation of Lake Baikal.
Author Contributions
Conceptualization, L.C. and S.B.; methodology L.C., K.B. and A.B.; formal analysis, S.B. and L.C.; investigation, L.C., K.B. and A.B.; data curation, K.B. and L.C.; writing—original draft preparation, L.C. and K.B.; writing—review and editing, L.C., S.B. and K.B.; supervision, S.B.; funding acquisition, S.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the State Assignments of the Limnological Institute Siberian Branch of the Russian Academy of Sciences (0279-2021-0011).
Institutional Review Board Statement
Not applicable because humans or animals are not involved in the study.
Informed Consent Statement
Not applicable since studies not involving humans.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The authors appreciate the help and technical support of the National Scientific Center of Marine Biology FEB RAS (the former A.V. Zhirmunsky Institute of Marine Biology).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Garrity, G.M.; Bell, J.A.; Lilburn, T.E. Family II Oxalobacteraceae fam. nov. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Garrity, G.M., Brenner, D.J., Krieg, N.R., Staley, J., Eds.; Springer: New York, NY, USA, 2005; Volume 2, p. 623. [Google Scholar] [CrossRef]
- Coenye, T. The family Burkholderiaceae. In The Prokaryotes: Alphaproteobacteria and Betaproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: New York, NY, USA, 2014; pp. 759–776. [Google Scholar] [CrossRef]
- Gillis, M.; Logan, N.A. Janthinobacterium. In Bergey’s Manual of Systematics of Archaea and Bacteria; Trujillo, M.E., Dedysh, S., DeVos, P., Hedlund, B., Kämpfer, P., Rainey, F.A., Whitman, W.B., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA; Bergey’s Manual Trust: Athen, GA, USA, 2015; pp. 1–12. [Google Scholar] [CrossRef]
- De Ley, J.; Segers, P.; Gillis, M. Intra- and intergeneric similarities of Chromobacterium and Janthinobacterium ribosomal ribonucleic acid cistrons. Int. J. Syst. Evol. Microbiol. 1978, 28, 154–168. [Google Scholar] [CrossRef] [Green Version]
- Sneath, P.H.A. Genus Janthinobacterium. In Bergey’s Manual of Systematic Bacteriology; Krieg, N.R., Holt, J.C., Eds.; The Williams & Wilkins Co.: Baltimore, MD, USA, 1984; pp. 376–377. [Google Scholar]
- Schloss, P.D.; Allen, H.K.; Klimowicz, A.K.; Mlot, C.; Gross, J.A.; Savengsuksa, S.; McEllin, J.; Clardy, J.; Ruess, R.W.; Handelsman, J. Psychrotrophic strain of Janthinobacterium lividum from a cold Alaskan soil produces prodigiosin. DNA Cell Biol. 2010, 29, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Skrivergaard, S.; Korsgaard, B.S.; Schreiber, L.; Marshall, I.P.; Finster, K.; Schramm, A. High quality draft genome sequence of Janthinobacterium psychrotolerans sp. nov., isolated from a frozen freshwater pond. Stand. Genom. Sci. 2017, 12, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillis, M.; De Ley, J. The Genera Chromobacterium and Janthinobacterium. In The Prokaryotes: Handbook on the Biology of Bacteria, 2nd ed.; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 7, pp. 737–746. [Google Scholar] [CrossRef]
- Leon, L.L.; Miranda, C.C.; De Souza, A.O.; Durán, N. Antileishmanial activity of the violacein extracted from Chromobacterium violaceum. J. Antimicrob. Chemother. 2001, 48, 449–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrighetti-Fröhner, C.R.; Antonio, R.V.; Creczynski-Pasa, T.B.; Barardi, C.R.; Simões, C.M. Cytotoxicity and potential antiviral evaluation of violacein produced by Chromobacterium violaceum. Mem. Inst. Oswaldo Cruz 2003, 98, 843–848. [Google Scholar] [CrossRef]
- Pantanella, F.; Berlutti, F.; Passariello, C.; Sarli, S.; Morea, C.; Schippa, S. Violacein and biofilm production in Janthinobacterium lividum. J. Appl. Microbiol. 2006, 102, 992–999. [Google Scholar] [CrossRef]
- Konzen, M.; De Marco, D.; Cordova, C.A.; Vieira, T.O.; Antônio, R.V.; Creczynski-Pasa, T.B. Antioxidant properties of violacein: Possible relation on its biological function. Bioorg. Med. Chem. 2006, 14, 8307–8313. [Google Scholar] [CrossRef]
- Durán, N.; Justo, G.Z.; Ferreira, C.V.; Melo, P.S.; Cordi, L.; Martins, D. Violacein: Properties and biological activities. Biotechnol. Appl. Biochem. 2007, 48, 127–133. [Google Scholar] [CrossRef]
- Lopes, S.C.; Blanco, Y.C.; Justo, G.Z.; Nogueira, P.A.; Rodrigues, F.L.; Goelnitz, U.; Wunderlich, G.; Facchini, G.; Brocchi, M.; Duran, N.; et al. Violacein extracted from Chromobacterium violaceum inhibits Plasmodium growth in vitro and in vivo. Antimicrob. Agents Chemother. 2009, 53, 2149–2152. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, A.L.; Gocke, Y.; Bolten, C.; Brock, N.L.; Dickschat, J.S.; Wittmann, C. Microbial production of the drugs violacein and deoxyviolacein: Analytical development and strain comparison. Biotechnol. Lett. 2012, 34, 717–720. [Google Scholar] [CrossRef]
- Asencio, G.; Lavin, P.; Alegría, K.; Domínguez, M.; Bello, H.; González-Rocha, G.; González-Aravena, M. Antibacterial activity of the Antarctic bacterium Janthinobacterium sp. SMN 33.6 against multi-resistant gram-negative bacteria. Electron. J. Biotechnol. 2014, 17, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Ballestriero, F.; Daim, M.; Penesyan, A.; Nappi, J.; Schleheck, D.; Bazzicalupo, P.; Di Schiavi, E.; Egan, S. Antinematode activity of Violacein and the role of the insulin/IGF-1 pathway in controlling violacein sensitivity in Caenorhabditis elegans. PLoS ONE 2014, 9, e109201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimi, S.M.; Xu, T.; Wei, M.Q. Violacein anticancer activity is enhanced under hypoxia. Oncol. Rep. 2015, 33, 1731–1736. [Google Scholar] [CrossRef] [PubMed]
- Sasidharan, A.; Sasidharan, N.K.; Amma, D.B.; Vasu, R.K.; Nataraja, A.V.; Bhaskaran, K. Antifungal activity of violacein purified from a novel strain of Chromobacterium sp. NIIST (MTCC 5522). J. Microbiol. 2015, 53, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Valdes, N.; Soto, P.; Cottet, L.; Alarcon, P.; Gonzalez, A.; Castillo, A.; Corsini, G.; Tello, M. Draft genome sequence of Janthinobacterium lividum strain MTR reveals its mechanism of capnophilic behavior. Stand. Genom. Sci. 2015, 10, 110. [Google Scholar] [CrossRef] [Green Version]
- Baricz, A.; Teban, A.; Chiriac, C.M.; Szekeres, E.; Farkas, A.; Nica, M.; Dascălu, A.; Oprișan, C.; Lavin, P.; Coman, C. Investigating the potential use of an Antarctic variant of Janthinobacterium lividum for tackling antimicrobial resistance in a One Health approach. Sci. Rep. 2018, 8, 15272. [Google Scholar] [CrossRef] [Green Version]
- Dike-Ndudim, J.N.; Ugenyi, L.C.; Ndubueze, C.W. Assessment of antifungal potentials of violacein extract from Chromobacterium violaceum isolated from domestic and recreational water sources in Owerri, Imo State, Nigeria. World J. Adv. Res. Rev. 2021, 10, 168–172. [Google Scholar] [CrossRef]
- Kim, Y.J.; Yuk, N.; Shin, H.J.; Jung, H.J. The Natural Pigment Violacein Potentially Suppresses the Proliferation and Stemness of Hepatocellular Carcinoma Cells In Vitro. Int. J. Mol. Sci. 2021, 22, 10731. [Google Scholar] [CrossRef]
- Matz, C.; Deines, P.; Boenigk, J.; Arndt, H.; Eberl, L.; Kjelleberg, S.; Jürgens, K. Impact of violacein-producing bacteria on survival and feeding of bacterivorous nanoflagellates. Appl. Environ. Microbiol. 2004, 70, 1593–1599. [Google Scholar] [CrossRef] [Green Version]
- Harris, R.N.; Brucker, R.M.; Walke, J.B.; Becker, M.H.; Schwantes, C.R.; Flaherty, D.C.; Lam, B.A.; Woodhams, D.C.; Briggs, C.J.; Vredenburg, V.T.; et al. Skin microbes on frogs prevent morbidity and mortality caused by a lethal skin fungus. ISME J. 2009, 3, 818–824. [Google Scholar] [CrossRef]
- Jensen, S.; Ovreas, L.; Bergh, O.; Torsvik, V. Phylogenetic analysis of bacterial communities associated with larvae of the Atlantic halibut proposesuccession from a uniform normal flora. Syst. Appl. Microbiol. 2004, 27, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Oh, W.T.; Giri, S.S.; Yun, S.; Kim, H.J.; Kim, S.G.; Kim, S.W.; Kang, J.W.; Han, S.J.; Kwon, J.; Jun, J.W.; et al. Janthinobacterium lividum as An Emerging Pathogenic Bacterium Affecting Rainbow Trout (Oncorhynchus mykiss) Fisheries in Korea. Pathogens 2019, 8, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, B.; Gonzalez, C.J.; Stobie, M.; Curry, J.I.; McLoughlin, M.F. Recovery of Janthinobacterium lividum from diseased rainbow trout, Oncorhynchus mykiss (Walbaum), in Northern Ireland and Scotland. J. Fish Dis. 1992, 15, 357–359. [Google Scholar] [CrossRef]
- Lincoln, S.P.; Fermor, T.R.; Tindall, B.J. Janthinobacterium agaricidamnosum sp. nov., a soft rot pathogen of Agaricusbisporus. Int. J. Syst. Evol. Microbiol. 1999, 49, 1577–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haack, F.S.; Poehlein, A.; Kröger, C.; Voigt, C.A.; Piepenbring, M.; Bode, H.B.; Daniel, R.; Schäfer, W.; Streit, W.R. Molecular keys to the Janthinobacterium and Duganella spp. interaction with the plant pathogen Fusarium graminearum. Front. Microbiol. 2016, 7, 1668. [Google Scholar] [CrossRef] [Green Version]
- Jung, W.J.; Kim, S.W.; Giri, S.S.; Kim, H.J.; Kim, S.G.; Kang, J.W.; Kwon, J.; Lee, S.B.; Oh, W.T.; Jun, J.W.; et al. Janthinobacterium tructae sp. nov., Isolated from Kidney of Rainbow Trout (Oncorhynchus mykiss). Pathogens 2021, 10, 229. [Google Scholar] [CrossRef]
- Petrushin, I.S.; Belikov, S.I.; Chernogor, L.I. Draft Genome Sequence of Janthinobacterium sp. Strain SLB01, Isolated from the Diseased Sponge Lubomirskia baicalensis. Microbiol. Resour. Announc. 2019, 8, e01108–e01119. [Google Scholar] [CrossRef] [Green Version]
- Khanaev, I.V.; Kravtsova, L.S.; Maikova, O.O.; Bukshuk, N.A.; Sakirko, M.V.; Kulakova, N.V.; Butina, T.V.; Nebesnykh, I.A.; Belikov, S.I. Current state of the sponge fauna (Porifera: Lubomirskiidae) of Lake Baikal: Sponge disease and the problem of conservation of diversity. J. Great Lakes Res. 2018, 44, 77–85. [Google Scholar] [CrossRef]
- Belikov, S.; Belkova, N.; Butina, T.; Chernogor, L.; Van Kley, A.M.; Nalian, A.; Rorex, C.; Khanaev, I.; Maikova, O.; Feranchuk, S. Diversity and shifts of the bacterial community associated with Baikal sponge mass mortalities. PLoS ONE 2019, 14, e0213926. [Google Scholar] [CrossRef]
- Chernogor, L.; Klimenko, E.; Khanaev, I.; Belikov, S. Microbiome analysis of healthy and diseased sponges Lubomirskia baicalensis by using cell cultures of primmorphs. PeerJ 2020, 8, e9080. [Google Scholar] [CrossRef]
- Belikov, S.I.; Petrushin, I.S.; Chernogor, L.I. Genome Analysis of the Janthinobacterium sp. Strain SLB01 from the Diseased Sponge of the Lubomirskia baicalensis. Curr. Issues Mol. Biol. 2021, 43, 2220–2237. [Google Scholar] [CrossRef] [PubMed]
- Dantas, C.; Tauler, R.; Ferreira, M.M.C. Exploring in vivo violacein biosynthesis by application of multivariate curve resolution on fused UV–VIS absorption, fluorescence, and liquid chromatography–mass spectrometry data. Anal. Bioanal. Chem. 2012, 405, 1293–1302. [Google Scholar] [CrossRef] [PubMed]
- Shimaraev, M.N.; Troitskaya, E.S.; Gnatovskii, R.Y. Modern Climate Changes and Deep Water Temperature of Lake Baikal. Dokl. Earth Sci. 2009, 427, 804–808. [Google Scholar] [CrossRef]
- Vacelet, J.; Vacelet, E.; Gaino, E.; Gallissian, M.-F. Bacterial attack of spongin skeleton during the 1986–1990 Mediterranean sponge disease. In Sponges in Time and Space; van Kempen, T.M.G., Braekman, J.C., van Soest, R.W.M., Eds.; A.A. Balkema: Rotterdam, The Netherlands, 1994; pp. 355–362. [Google Scholar]
- Webster, N.S. Sponge disease: A global threat? Environ. Microbiol. 2007, 9, 1363–1375. [Google Scholar] [CrossRef]
- Pita, L.; Rix, L.; Slaby, B.M.; Franke, A.; Hentschel, U. The sponge holobiont in a changing ocean: From microbes to ecosystems. Microbiome 2018, 6, 46. [Google Scholar] [CrossRef]
- Erwin, P.M.; Pita, L.; López-Legentil, S.; Turon, X. Stability of sponge-associated bacteria over large seasonal shifts in temperature and irradiance. Appl. Environ. Microbiol. 2012, 78, 7358–7368. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.; Liu, M.; Simister, R.; Webster, N.S.; Thomas, T. Marine microbial symbiosis heats up: The phylogenetic and functional response of a sponge holobiont to thermal stress. ISME J. 2013, 7, 991–1002. [Google Scholar] [CrossRef]
- Vizza, C.; Pechal, J.L.; Benbow, M.E.; Lang, J.M.; Chaloner, D.T.; Jones, S.E.; Lamberti, G.A. Nitrate amendment reduces biofilm biomass and shifts microbial communities in remote, oligotrophic ponds. Freshw. Sci. 2018, 37, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Acharya, V.; Singh, D.; Kumar, S. Strategies for high-altitude adaptation revealed from high-quality draft genome of non-violacein producing Janthinobacterium lividum ERGS5:01. Stand. Genom. Sci. 2018, 13, 11. [Google Scholar] [CrossRef]
- Lu, H.; Deng, T.; Cai, Z.; Liu, F.; Yang, X.; Wang, Y.; Xu, M. Janthinobacterium violaceinigrum sp. nov., Janthinobacterium aquaticum sp. nov. and Janthinobacterium rivuli sp. nov., isolated from a subtropical stream in China. Int. J. Syst. Evol. Microbiol. 2020, 70, 2719–2725. [Google Scholar] [CrossRef]
- Aslamov, I.A.; Kozlov, V.V.; Kirillin, G.B.; Mizandrontsev, I.B.; Kucher, K.M.; Makarov, M.M.; Gornov, A.Y.; Granin, N.G. Ice–water heat exchange during ice growth in Lake Baikal. J. Great Lakes Res. 2014, 40, 599–607. [Google Scholar] [CrossRef]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Camara, M.; Daykin, M.; Lamb, J.H.; Swift, S.; Bycroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 1997, 143, 3703–3711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stauff, D.L.; Bassler, B.L. Quorum sensing in Chromobacterium violaceum: DNA recognition and gene regulation by the CviR receptor. J. Bacteriol. 2011, 193, 3871–3878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
The growth of the Janthinobacterium sp. SLB01 strain at 22 °C on the different media (LB agar, PCA, and R 2A). Results are reported as mean values of three independent experiments performed.
Figure 1.
The growth of the Janthinobacterium sp. SLB01 strain at 22 °C on the different media (LB agar, PCA, and R 2A). Results are reported as mean values of three independent experiments performed.

Figure 2.
The curves of the Janthinobacterium sp. SLB01 strain at different temperatures of cultivation in LB broth (at 3, 22 and 30 °C). Results are shown as mean values of three independent experiments performed.
Figure 2.
The curves of the Janthinobacterium sp. SLB01 strain at different temperatures of cultivation in LB broth (at 3, 22 and 30 °C). Results are shown as mean values of three independent experiments performed.

Figure 3.
Light microscopy of the growth of the Janthinobacterium sp. SLB01 strain at different temperatures and in different media: (a) cultivation at 3 °C in PCA; (b) cultivation at 3 °C in R 2A; (c) cultivation at 3 °C in LB agar; (d) cultivation at 22 °C in PCA; (e) cultivation at 22 °C in R 2A; (f) cultivation at 22 °C in LB agar; (g) cultivation at 30 °C in PCA; (h) cultivation at 30 °C in R 2A; (i) cultivation at 30 °C in LB agar. Scale bar 10 µm.
Figure 3.
Light microscopy of the growth of the Janthinobacterium sp. SLB01 strain at different temperatures and in different media: (a) cultivation at 3 °C in PCA; (b) cultivation at 3 °C in R 2A; (c) cultivation at 3 °C in LB agar; (d) cultivation at 22 °C in PCA; (e) cultivation at 22 °C in R 2A; (f) cultivation at 22 °C in LB agar; (g) cultivation at 30 °C in PCA; (h) cultivation at 30 °C in R 2A; (i) cultivation at 30 °C in LB agar. Scale bar 10 µm.

Figure 4.
Characterization of the violacein of the Janthinobacterium sp. SLB01 strain extracted by different methods: (a) chromatogram of violacein using extraction with n-butanol; (b) chromatogram of violacein using extraction with chloroform. OU—optical density in optical units.
Figure 4.
Characterization of the violacein of the Janthinobacterium sp. SLB01 strain extracted by different methods: (a) chromatogram of violacein using extraction with n-butanol; (b) chromatogram of violacein using extraction with chloroform. OU—optical density in optical units.

Figure 5.
Production of violacein of the Janthinobacterium sp. SLB01 strain at different temperatures for five days. (a) Cultivation at 3 °C; (b) cultivation at 22 °C; (c) cultivation at 30 °C. The LB broth was taken as a control.
Figure 5.
Production of violacein of the Janthinobacterium sp. SLB01 strain at different temperatures for five days. (a) Cultivation at 3 °C; (b) cultivation at 22 °C; (c) cultivation at 30 °C. The LB broth was taken as a control.

Figure 6.
The curves of violacein production by the Janthinobacterium sp. SLB01 strain at 3, 22, and 30 °C in LB broth, OD585. Results are shown as mean values of three independent experiments performed.
Figure 6.
The curves of violacein production by the Janthinobacterium sp. SLB01 strain at 3, 22, and 30 °C in LB broth, OD585. Results are shown as mean values of three independent experiments performed.

Figure 7.
The curves of biofilm formation by Janthinobacterium sp. SLB01 strain at 3, 22, and 30 °C. Results are shown as mean values of three independent experiments performed.
Figure 7.
The curves of biofilm formation by Janthinobacterium sp. SLB01 strain at 3, 22, and 30 °C. Results are shown as mean values of three independent experiments performed.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characteristics strain of the Janthinobacterium sp. SLB01 strain.
| Characteristics | Janthinobacterium sp. Strain SLB01 |
|---|---|
| Gram’s reaction | − |
| Motility | + |
| Nitrate reduction | + |
| H2S production | + |
| Urease production | − |
| Phenylalanine deamination | − |
| Lipolytic activity | + |
| Hydrolysis of gelatine | + |
| Starch | + |
| Casein | + |
| Arabinose | − |
| Glucose | + |
| Lactose | + |
| Mannose | + |
| Sucrose | + |
| Citrate | + |
| Adonitol | − |
| Inositol | + |
| D-Mannitol | − |
| D-Sorbitol | − |
| L-Arginine | + |
| L-Lysine | + |
| L-Ornithine | + |
+ = Positive, − = negative.
Table 2.
Antibiotic susceptibility test results of the Janthinobacterium sp. SLB01 strain by the disk diffusion method.
Table 2.
Antibiotic susceptibility test results of the Janthinobacterium sp. SLB01 strain by the disk diffusion method.
| Antibiotic Tested | Symbol | Dose (µg) | Inhibition Zone Diameter (mm) | Interpretation |
|---|---|---|---|---|
| Ampicillin | AMP | 10 | 5 | R |
| Ceftriaxone | CTR | 30 | 19 | I |
| Imipenem | IPM | 10 | 6 | R |
| Nalidixic acid | NA | 30 | 13 | R |
| Levofloxacin | LE | 5 | 15 | R |
| Lomefloxacin | LOM | 10 | 21 | I |
| Norfloxacin | NX | 10 | 5 | R |
| Ofloxacin | OF | 5 | 14 | I |
| Ciprofloxacin | CIP | 5 | 22 | I |
| Amikacin | AK | 30 | 22 | S |
| Gentalyn | GEN | 10 | 8 | R |
| Gentamicin | HLG | 120 | 16 | S |
| Kanamycin | K | 30 | 18 | S |
| Netilmycin | NET | 30 | 9 | R |
| Tobramycin | TOB | 10 | 15 | S |
| Doxycycline | DO | 30 | 30 | S |
| Tetracycline | TE | 30 | 25 | S |
| Chloramphenicol | C | 30 | 5 | R |
| Erythromycin | E | 15 | 5 | R |
| Rifampicin | RIF | 5 | 20 | S |
S—susceptible, I—intermediate, R—resistant.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chernogor, L.; Bakhvalova, K.; Belikova, A.; Belikov, S. Isolation and Properties of the Bacterial Strain Janthinobacterium sp. SLB01. Microorganisms 2022, 10, 1071. https://doi.org/10.3390/microorganisms10051071
AMA Style
Chernogor L, Bakhvalova K, Belikova A, Belikov S. Isolation and Properties of the Bacterial Strain Janthinobacterium sp. SLB01. Microorganisms. 2022; 10(5):1071. https://doi.org/10.3390/microorganisms10051071
Chicago/Turabian StyleChernogor, Lubov, Kseniya Bakhvalova, Alina Belikova, and Sergei Belikov. 2022. "Isolation and Properties of the Bacterial Strain Janthinobacterium sp. SLB01" Microorganisms 10, no. 5: 1071. https://doi.org/10.3390/microorganisms10051071
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.