
First Report of Lumpy Skin Disease in Myanmar and Molecular Analysis of the Field Virus Isolates

 , , ,
, , ,
Abstract
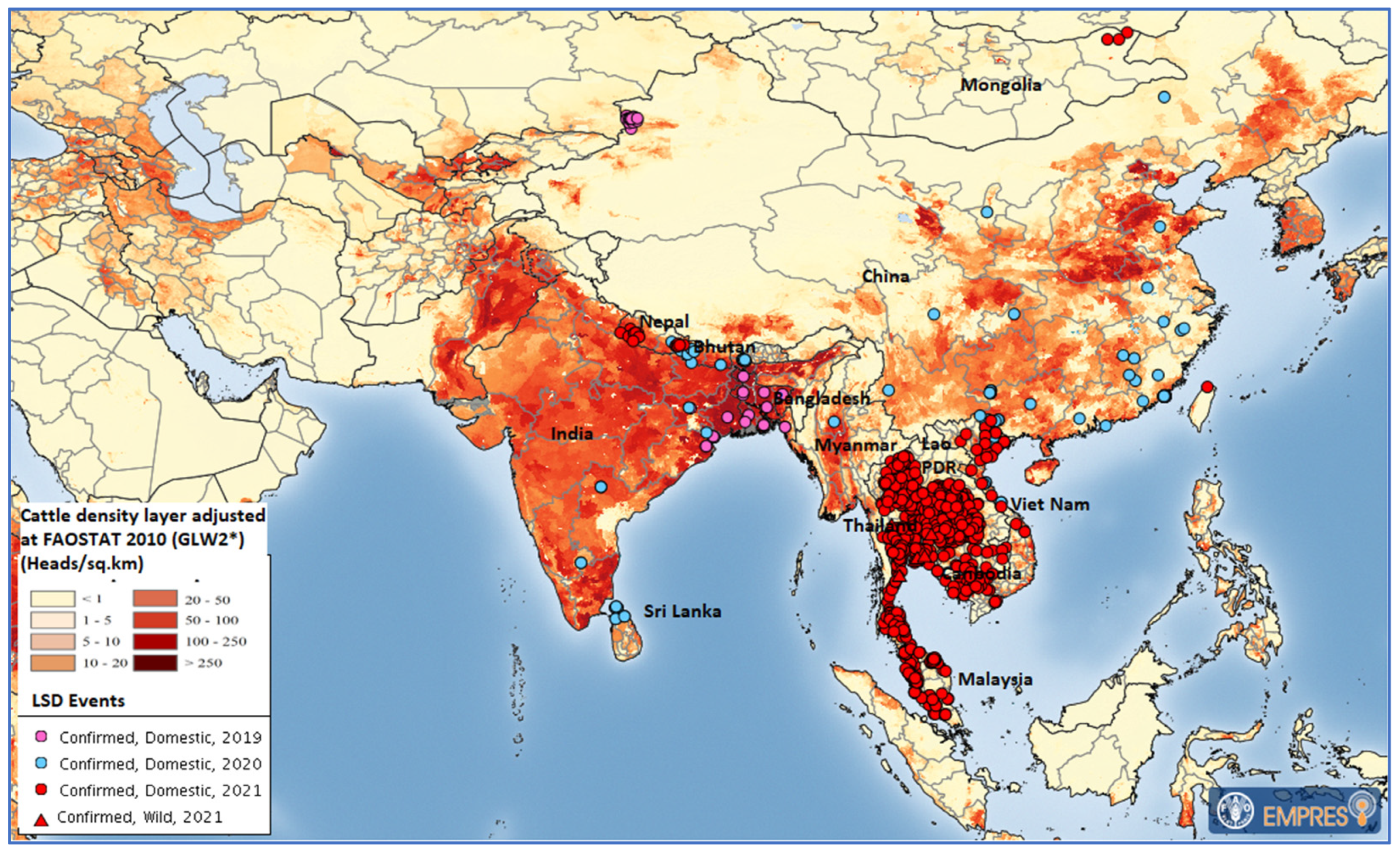
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Transport, Storage, and DNA Extraction
2.2. Detection of Capripoxvirus and Differential Diagnosis
2.3. Amplification and Sequencing of LSDV Target Genes
2.4. Sequence Analysis
3. Results
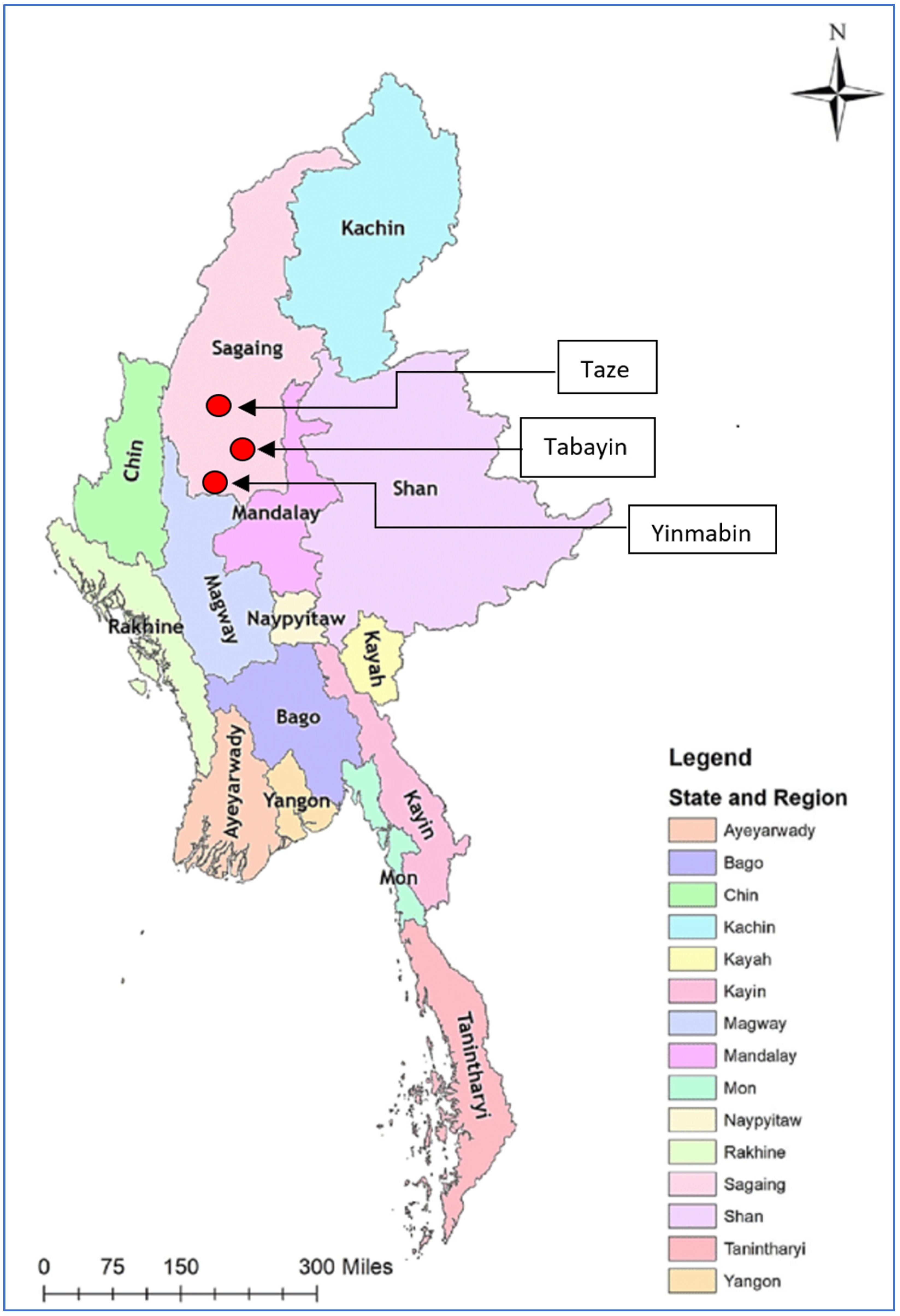
3.1. Clinical Findings
3.2. Molecular Diagnosis of CaPV
3.3. LSDV Gene Amplification and Sequencing
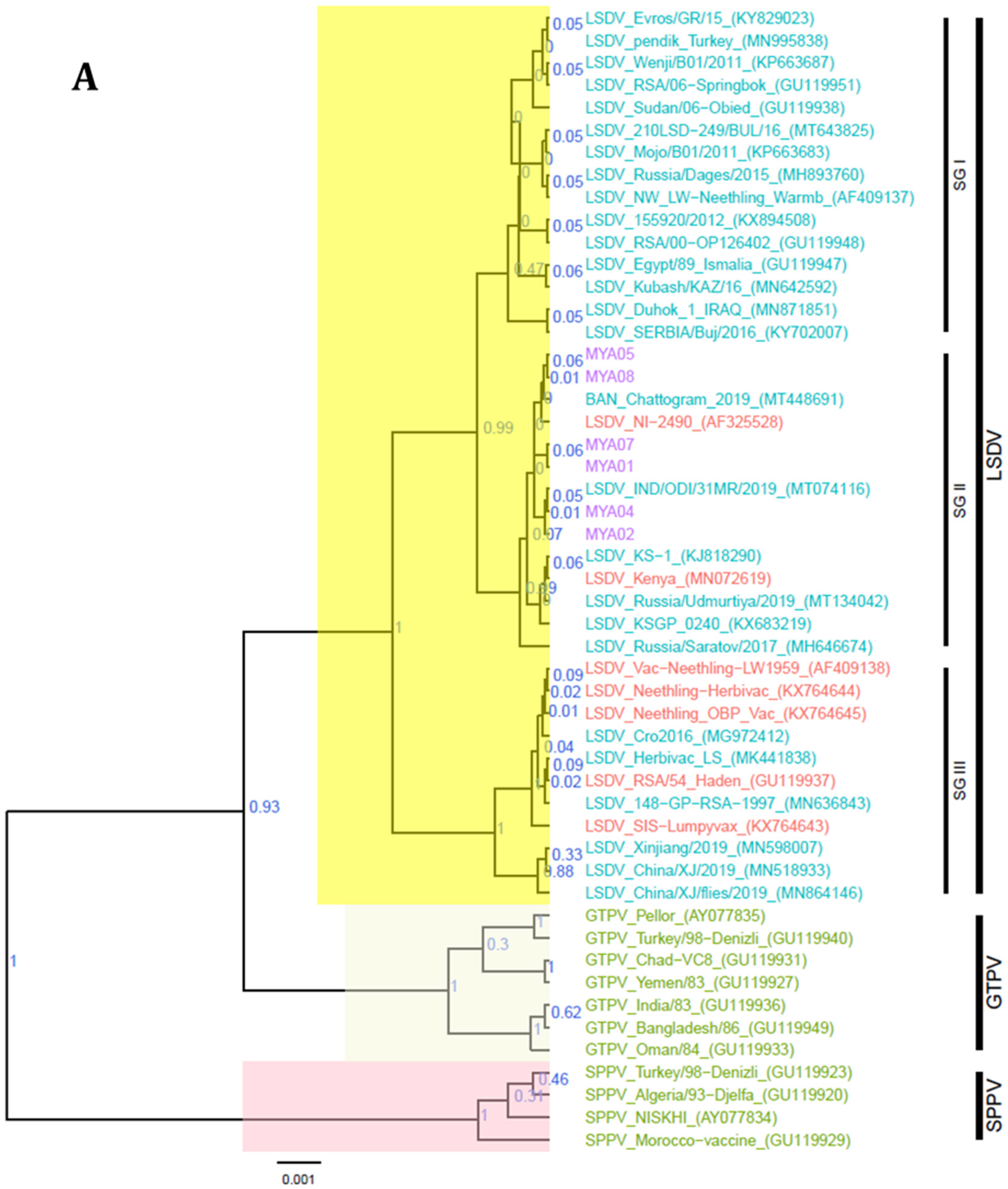
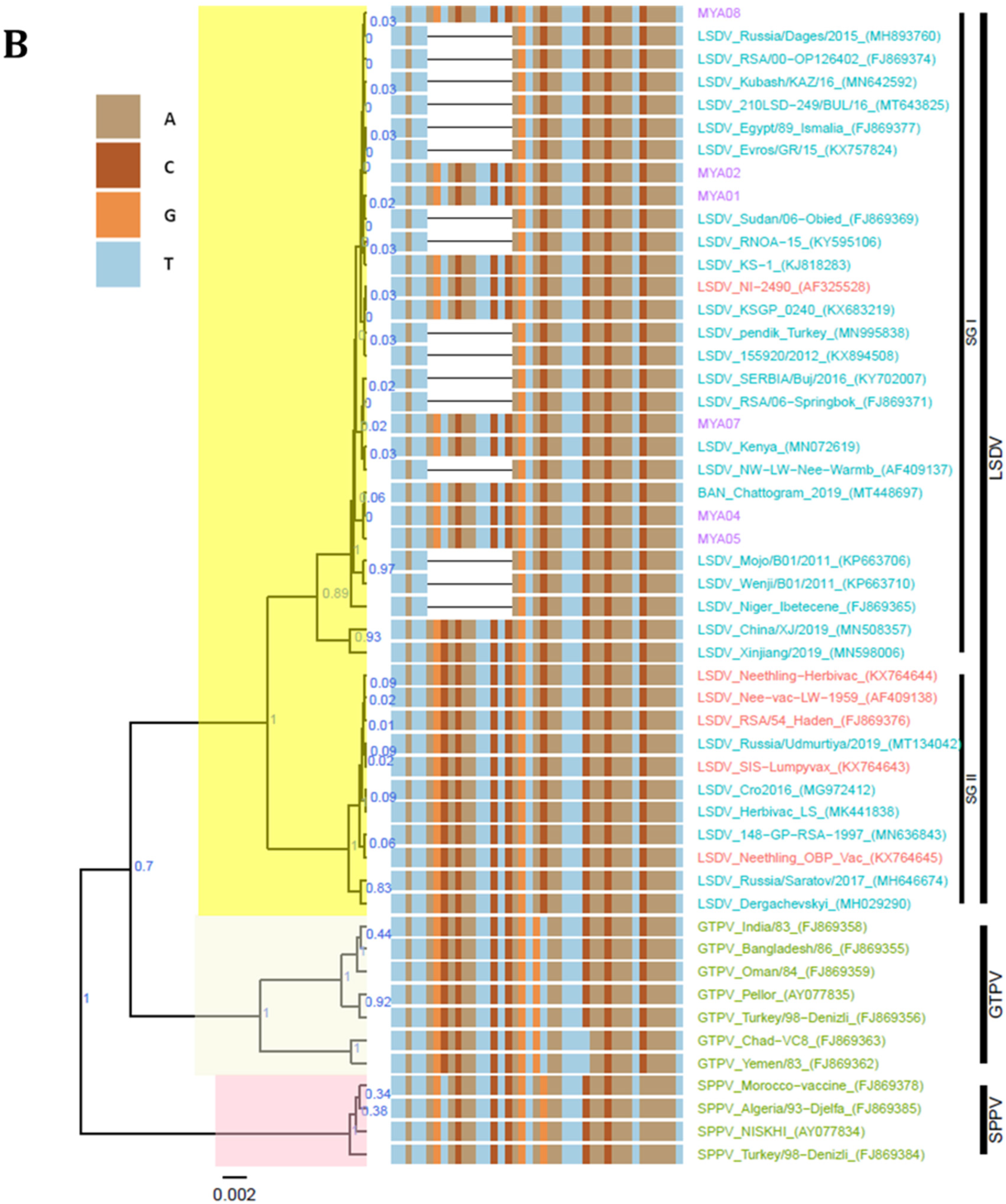
3.4. Sequence Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Buller, R.M.; Arif, B.M.; Black, D.N.; Dumbell, K.R.; Esposito, J.J.; Lefkowitz, E.J.; McFadden, G.; Moss, B.; Mercer, A.A.; Moyer, R.W. Family Poxviridae. In Virus Taxonomy: Classification and Nomenclature of Viruses. Eighth Report of the International Committee on Taxonomy of Viruses; Elsevier: Amsterdam, The Netherlands; Academic Press: San Diego, CA, USA, 2005; pp. 117–133. [Google Scholar]
- Baldacchino, F.; Muenworn, V.; Desquesnes, M.; Desoli, F.; Charoenviriyaphap, T.; Duvallet, G. Transmission of Pathogens by Stomoxys Flies (Diptera, Muscidae): A Review. Parasite 2013, 20, 26. [Google Scholar] [CrossRef] [Green Version]
- OIE. Lumpy Skin Disease. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; OIE: Paris, France, 2021. [Google Scholar]
- Babiuk, S.; Bowden, T.R.; Boyle, D.B.; Wallace, D.B.; Kitching, R.P. Capripoxviruses: An Emerging Worldwide Threat to Sheep, Goats and Cattle. Transbound. Emerg. Dis. 2008, 55, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Livestock Breeding and Veterinary Department. National Livestock Baseline Survey 2018 Report; Livestock Breeding and Veterinary Department: Yangon, Myanmar, 2019. [Google Scholar]
- Lumpy Skin Disease (LSD)—OIE—Asia. Available online: https://rr-asia.oie.int/en/projects/lumpy-skin-disease-lsd/ (accessed on 17 March 2022).
- Talukdar, F.; Fakir, A.; Hasan, Z.; Osmani, T.M.G.; Choudhury, G.A.; Budhay, S.; Sadekuzzaman, M.; Brum, E.; Bhowmic, H.R.; Noorjahan, B. Identification of Lumpy Skin Disease (LSD): First-Time in Bangladesh during the Investigation of Unknown Skin Disease of Cattle. Int. J. Infect. Dis. 2020, 101, 360–361. [Google Scholar] [CrossRef]
- Hasib, F.M.Y.; Islam, M.S.; Das, T.; Rana, E.A.; Uddin, M.H.; Bayzid, M.; Nath, C.; Hossain, M.A.; Masuduzzaman, M.; Das, S.; et al. Lumpy Skin Disease Outbreak in Cattle Population of Chattogram, Bangladesh. Vet. Med. Sci. 2021, 7, 1616–1624. [Google Scholar] [CrossRef] [PubMed]
- Sudhakar, S.B.; Mishra, N.; Kalaiyarasu, S.; Jhade, S.K.; Hemadri, D.; Sood, R.; Bal, G.C.; Nayak, M.K.; Pradhan, S.K.; Singh, V.P. Lumpy Skin Disease (LSD) Outbreaks in Cattle in Odisha State, India in August 2019: Epidemiological Features and Molecular Studies. Transbound. Emerg. Dis. 2020, 67, 2408–2422. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Xie, J.; Luo, J.; Shao, R.; Jia, K.; Li, S. Lumpy Skin Disease Outbreaks in China, since 3 August 2019. Transbound. Emerg. Dis. 2021, 68, 216–219. [Google Scholar] [CrossRef]
- Acharya, K.P.; Subedi, D. First Outbreak of Lumpy Skin Disease in Nepal. Transbound. Emerg. Dis. 2020, 67, 2280–2281. [Google Scholar] [CrossRef]
- Tran, H.T.T.; Truong, A.D.; Dang, A.K.; Ly, D.V.; Nguyen, C.T.; Chu, N.T.; Van Hoang, T.; Nguyen, H.T.; Nguyen, V.T.; Dang, H.V. Lumpy Skin Disease Outbreaks in Vietnam, 2020. Transbound. Emerg. Dis. 2021, 68, 977–980. [Google Scholar] [CrossRef]
- OIE-WAHIS. Available online: https://wahis.oie.int/#/report-info?reportId=28135 (accessed on 17 March 2022).
- Bowden, T.R.; Babiuk, S.L.; Parkyn, G.R.; Copps, J.S.; Boyle, D.B. Capripoxvirus Tissue Tropism and Shedding: A Quantitative Study in Experimentally Infected Sheep and Goats. Virology 2008, 371, 380–393. [Google Scholar] [CrossRef] [Green Version]
- Gelaye, E.; Mach, L.; Kolodziejek, J.; Grabherr, R.; Loitsch, A.; Achenbach, J.E.; Nowotny, N.; Diallo, A.; Lamien, C.E. A Novel HRM Assay for the Simultaneous Detection and Differentiation of Eight Poxviruses of Medical and Veterinary Importance. Sci. Rep. 2017, 7, srep42892. [Google Scholar] [CrossRef] [Green Version]
- Badhy, S.C.; Chowdhury, M.G.A.; Settypalli, T.B.K.; Cattoli, G.; Lamien, C.E.; Fakir, M.A.U.; Akter, S.; Osmani, M.G.; Talukdar, F.; Begum, N.; et al. Molecular Characterization of Lumpy Skin Disease Virus (LSDV) Emerged in Bangladesh Reveals Unique Genetic Features Compared to Contemporary Field Strains. BMC Vet. Res. 2021, 17, 61. [Google Scholar] [CrossRef] [PubMed]
- Gelaye, E.; Belay, A.; Ayelet, G.; Jenberie, S.; Yami, M.; Loitsch, A.; Tuppurainen, E.; Grabherr, R.; Diallo, A.; Lamien, C.E. Capripox Disease in Ethiopia: Genetic Differences between Field Isolates and Vaccine Strain, and Implications for Vaccination Failure. Antivir. Res. 2015, 119, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Koirala, P.; Meki, I.K.; Maharjan, M.; Settypalli, B.K.; Manandhar, S.; Yadav, S.K.; Cattoli, G.; Lamien, C.E. Molecular Characterization of the 2020 Outbreak of Lumpy Skin Disease in Nepal. Microorganisms 2022, 10, 539. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian Phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.Y. Ggtree: An r Package for Visualization and Annotation of Phylogenetic Trees with Their Covariates and Other Associated Data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.M.; Venter, E.H.; Shisler, J.L.; Gari, G.; Mekonnen, G.A.; Juleff, N.; Lyons, N.A.; De Clercq, K.; Upton, C.; Bowden, T.R.; et al. Review: Capripoxvirus Diseases: Current Status and Opportunities for Control. Transbound. Emerg. Dis. 2017, 64, 729–745. [Google Scholar] [CrossRef]
- Das, M.; Chowdhury, M.S.R.; Akter, S.; Mondal, A.K.; Uddin, M.J.; Rahman, M.M.; Rahman, M.M. An Updated Review on Lumpy Skin Disease: Perspective of Southeast Asian Countries. J. Adv. Biotechnol. Exp. Ther. 2021, 4, 322–333. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.M.; Stoltsz, W.H.; Troskie, M.; Wallace, D.B.; Oura, C.A.L.; Mellor, P.S.; Coetzer, J.A.W.; Venter, E.H. A Potential Role for Ixodid (Hard) Tick Vectors in the Transmission of Lumpy Skin Disease Virus in Cattle. Transbound. Emerg. Dis. 2011, 58, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Sprygin, A.; Pestova, Y.; Wallace, D.B.; Tuppurainen, E.; Kononov, A.V. Transmission of Lumpy Skin Disease Virus: A Short Review. Virus Res. 2019, 269, 197637. [Google Scholar] [CrossRef]
- Lamien, C.E.; Le Goff, C.; Silber, R.; Wallace, D.B.; Gulyaz, V.; Tuppurainen, E.; Madani, H.; Caufour, P.; Adam, T.; El Harrak, M.; et al. Use of the Capripoxvirus Homologue of Vaccinia Virus 30kDa RNA Polymerase Subunit (RPO30) Gene as a Novel Diagnostic and Genotyping Target: Development of a Classical PCR Method to Differentiate Goat Poxvirus from Sheep Poxvirus. Vet. Microbiol. 2011, 149, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Le Goff, C.; Lamien, C.E.; Fakhfakh, E.; Chadeyras, A.; Aba-Adulugba, E.; Libeau, G.; Tuppurainen, E.; Wallace, D.B.; Adam, T.; Silber, R.; et al. Capripoxvirus G-Protein-Coupled Chemokine Receptor: A Host-Range Gene Suitable for Virus Animal Origin Discrimination. J. Gen. Virol. 2009, 90, 1967–1977. [Google Scholar] [CrossRef] [PubMed]
- Menasherow, S.; Erster, O.; Rubinstein-Giuni, M.; Kovtunenko, A.; Eyngor, E.; Gelman, B.; Khinich, E.; Stram, Y. A High-Resolution Melting (HRM) Assay for the Differentiation between Israeli Field and Neethling Vaccine Lumpy Skin Disease Viruses. J. Virol. Methods 2016, 232, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Chibssa, T.R.; Settypalli, T.B.K.; Berguido, F.J.; Grabherr, R.; Loitsch, A.; Tuppurainen, E.; Nwankpa, N.; Tounkara, K.; Madani, H.; Omani, A.; et al. An HRM Assay to Differentiate Sheeppox Virus Vaccine Strains from Sheeppox Virus Field Isolates and Other Capripoxvirus Species. Sci. Rep. 2019, 9, 6646. [Google Scholar] [CrossRef] [Green Version]
- Bo, L.L.; Lwin, K.S.; Ungvanijban, S.; Knowles, N.J.; Wadsworth, J.; King, D.P.; Abila, R.; Qiu, Y. Foot-and-Mouth Disease Outbreaks due to an Exotic Serotype Asia 1 Virus in Myanmar in 2017. Transbound. Emerg. Dis. 2019, 66, 1067–1072. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Township | No. of Affected Animals | Species | Clinical Signs | No. of Samples | Samples Type | No. of Animals at Risk in Township | Morbidity % | No. of Dead Animals |
|---|---|---|---|---|---|---|---|---|
| Tabayin | 90 | cattle | Nodule, leg swollen, brisket swollen, nasal and ocular discharges | 7 | Exposed nodular lesion, blood | 5696 | 1.6% | - |
| Taze | 50 | cattle | Nodule, leg swollen | 3 | Exposed nodular lesion | 3571 | 1.4% | - |
| Yinmarbin | 40 | cattle | Nodule, leg swollen | 3 | Exposed nodular lesion | 4000 | 1% | - |
| Sr. | Host | Origin in Myanmar | Sample Type | Date of Collection | Vaccination History | CaPV RT-PCR | HRM Tm | Cq Value |
|---|---|---|---|---|---|---|---|---|
| 1 | Cattle | Taze township, Sagaing region | Nodule | Oct 2020 | Non-vaccinated | (+) | ||
| 2 | Cattle | Yinmabin township, Sagaing region | Serum | Oct 2020 | Non-vaccinated | (+) | ||
| 3 | Cattle | Tabayin township, Sagaing region | Nodule | Nov 2020 | Non-vaccinated | (+) | 77.6 | 19.08 |
| 4 | Cattle | Tabayin township, Sagaing region | Nodule | Nov 2020 | Non-vaccinated | (+) | 77.6 | 16.46 |
| 5 | Cattle | Tabayin township, Sagaing region | Whole Blood | Nov 2020 | Non-vaccinated | (+) | 77.4 | 32.14 |
| 6 | Cattle | Tabayin township, Sagaing region | Nodule | Nov 2020 | Non-vaccinated | (+) | 77.6 | 17.18 |
| 7 | Cattle | Tabayin township, Sagaing region | Nodule | Nov 2020 | Non-vaccinated | (+) | 77.6 | 14.91 |
| 8 | Cattle | Tabayin township, Sagaing region | Nodule | Nov 2020 | Non-vaccinated | (+) | 77.6 | 25.44 |
| 9 | Cattle | Tabayin township, Sagaing region | Nodule | Nov 2020 | Non-vaccinated | (+) | 77.6 | 26.01 |
| 10 | Cattle | Tabayin township, Sagaing region | Nodule | Nov 2020 | Non-vaccinated | (+) | 77.6 | 23.89 |
| 11 | Cattle | Tabayin township, Sagaing region | Nodule | Nov 2020 | Non-vaccinated | (+) | 77.8 | 23.87 |
| 12 | Cattle | Taze township, Sagaing region | Serum | Nov 2020 | Non-vaccinated | (+) | ||
| 13 | Cattle | Yinmabin township, Sagaing region | Nodule | Nov 2020 | Non-vaccinated | (+) | 77.6 | 33.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© FAO, 2022. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Com-mons Attribution (CC BY NC SA) license (https://creativecommons.org/licenses/by-nc-sa/3.0/igo/).
Share and Cite
Maw, M.T.; Khin, M.M.; Hadrill, D.; Meki, I.K.; Settypalli, T.B.K.; Kyin, M.M.; Myint, W.W.; Thein, W.Z.; Aye, O.; Palamara, E.; et al. First Report of Lumpy Skin Disease in Myanmar and Molecular Analysis of the Field Virus Isolates. Microorganisms 2022, 10, 897. https://doi.org/10.3390/microorganisms10050897
Maw MT, Khin MM, Hadrill D, Meki IK, Settypalli TBK, Kyin MM, Myint WW, Thein WZ, Aye O, Palamara E, et al. First Report of Lumpy Skin Disease in Myanmar and Molecular Analysis of the Field Virus Isolates. Microorganisms. 2022; 10(5):897. https://doi.org/10.3390/microorganisms10050897
Chicago/Turabian StyleMaw, Min Thein, Myint Myint Khin, David Hadrill, Irene Kasindi Meki, Tirumala Bharani Kumar Settypalli, Maung Maung Kyin, Win Win Myint, Wai Zin Thein, Ohnmar Aye, Elisa Palamara, and et al. 2022. "First Report of Lumpy Skin Disease in Myanmar and Molecular Analysis of the Field Virus Isolates" Microorganisms 10, no. 5: 897. https://doi.org/10.3390/microorganisms10050897