
A Method to Determine the Efficacy of a Commercial Phage Preparation against Uropathogens in Urine and Artificial Urine Determined by Isothermal Microcalorimetry

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Urine and Artificial Urine Preparation
2.2. Microorganisms and Culture Conditions
2.3. Phage Cocktail Description
2.4. Comparison of Isothermal Microcalorimetry with CFU, PFU and OD Data
2.5. Phage Cocktail Efficacy
2.6. Combination with Trimethoprim/Sulfamethoxazole
2.7. Data Analysis
3. Results
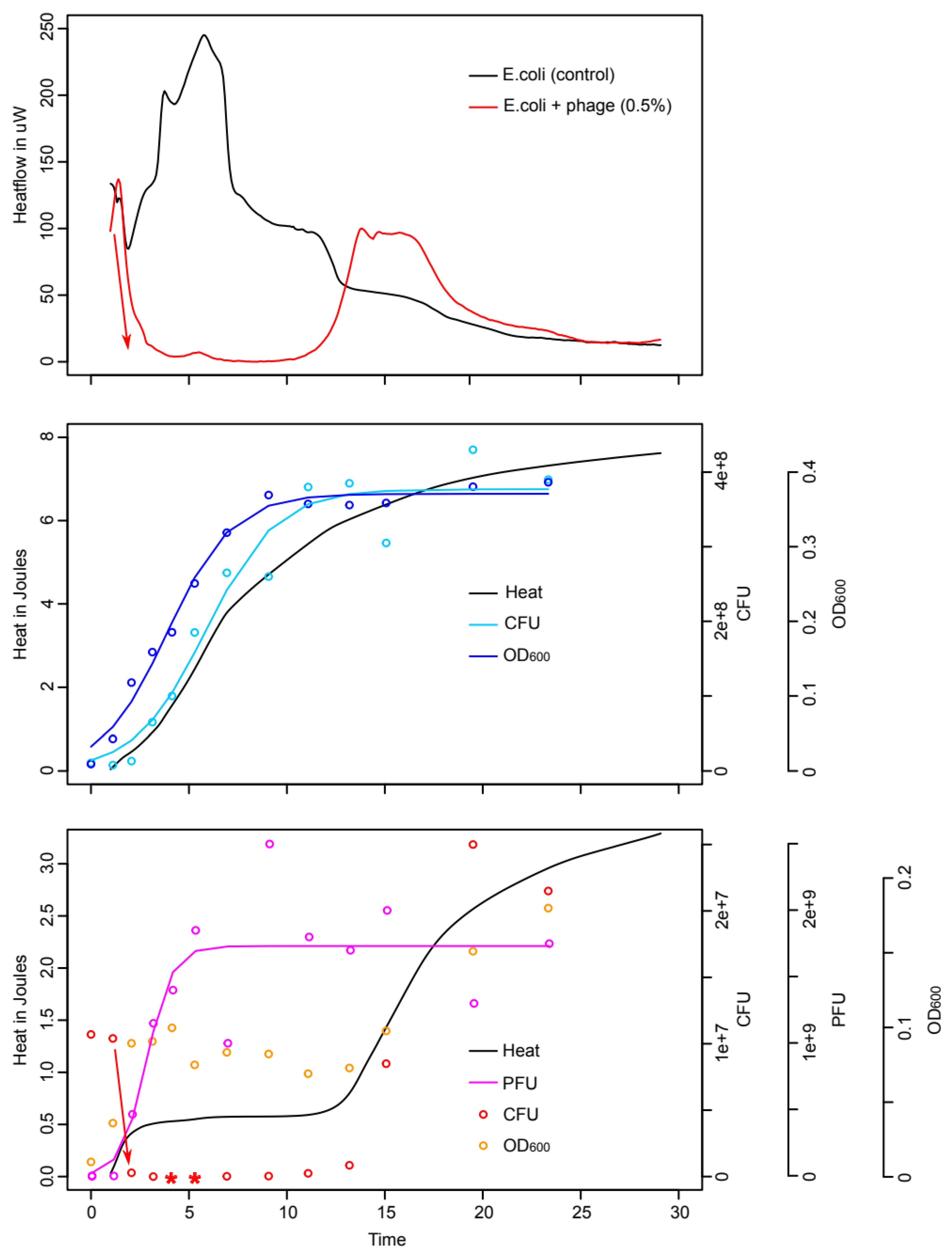
3.1. Comparison of Isothermal Microcalorimetry and Conventional Microbiology
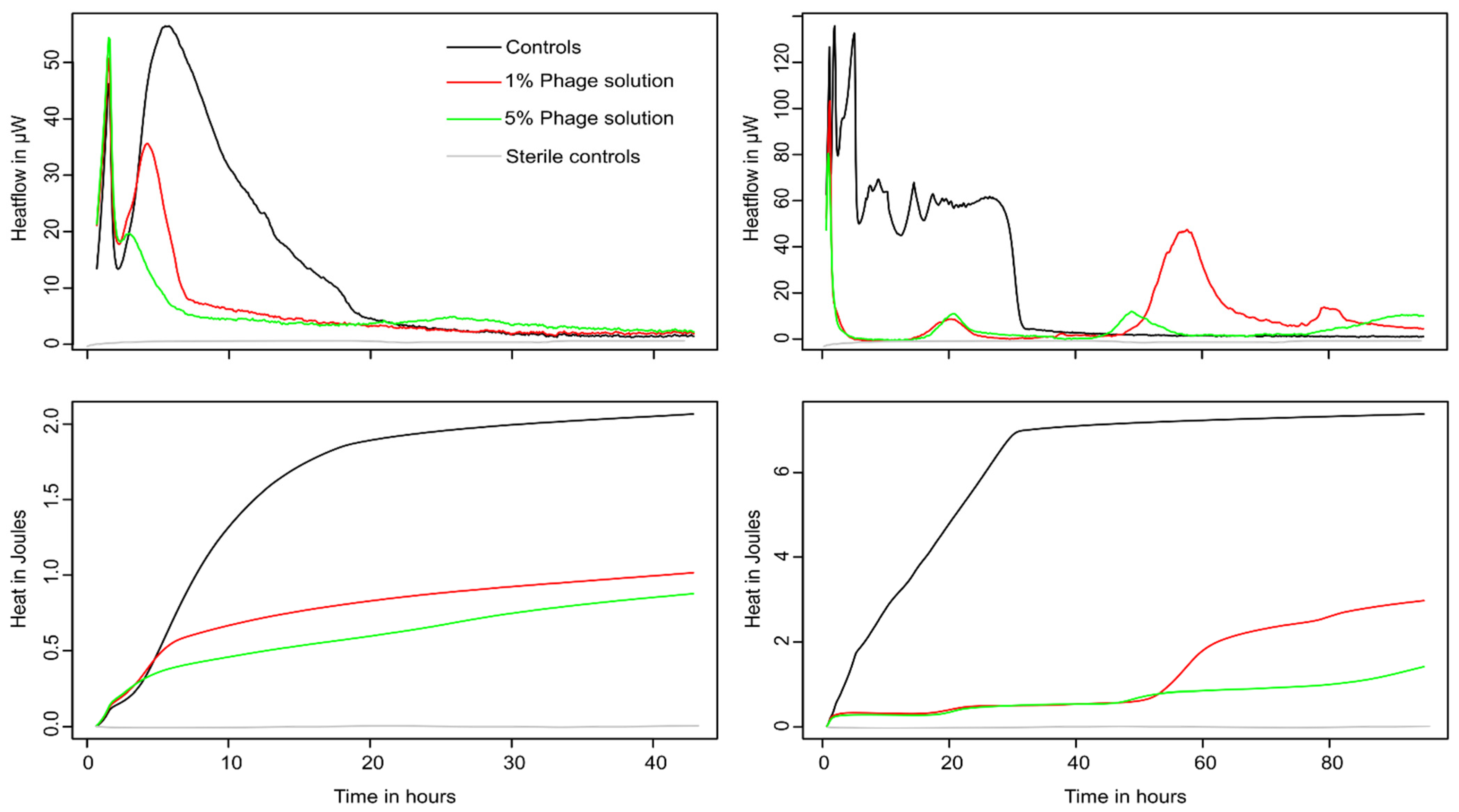
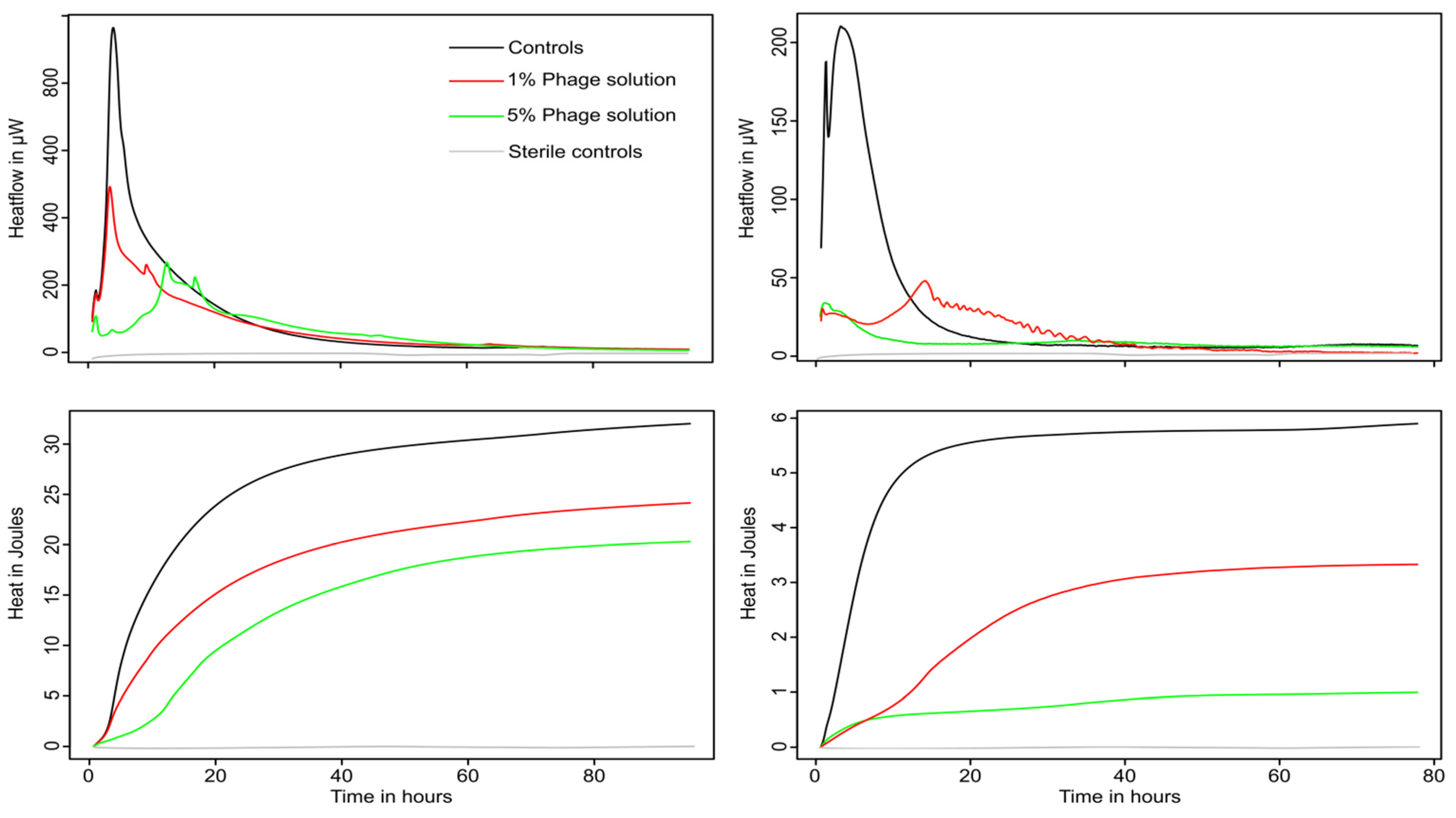
3.2. Monitoring of Phage Cocktail Antibacterial Activity
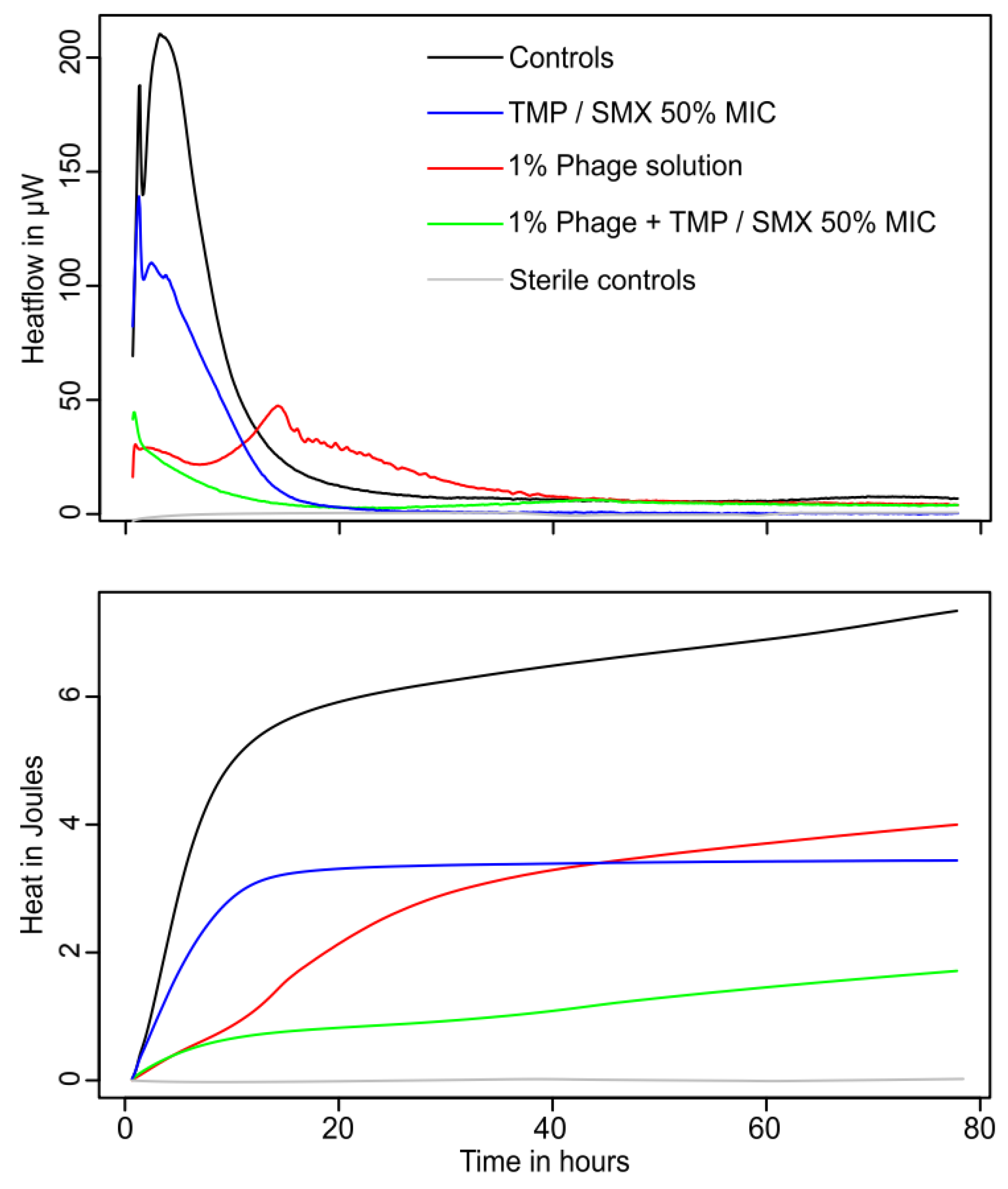
3.3. Combination of Phages and Trimethoprim/Sulfamethoxazole
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonkat, G.; Braissant, O.; Widmer, A.F.; Frei, R.; Rieken, M.; Wyler, S.; Gasser, T.C.; Wirz, D.; Daniels, A.U.; Bachmann, A. Rapid detection of urinary tract pathogens using microcalorimetry: Principle, technique and first results. BJU Int. 2012, 110, 892–897. [Google Scholar] [CrossRef] [PubMed]
- McLellan, L.K.; Hunstad, D.A. Urinary Tract Infection: Pathogenesis and Outlook. Trends Mol. Med. 2016, 22, 946–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolle, L.E. Urinary Tract Infection. Crit. Care Clin. 2013, 29, 699–715. [Google Scholar] [CrossRef] [PubMed]
- Mathai, D.; Jones, R.N.; Pfaller, M.A. Epidemiology and frequency of resistance among pathogens causing urinary tract infections in 1510 hospitalized patients: A report from the SENTRY Antimicrobial Surveillance Program (North America). Diagn. Microbiol. Infect. Dis. 2001, 40, 129–136. [Google Scholar] [CrossRef]
- Grabe, M.; Bjerklund-Johansen, T.E.; Botto, H.; European Association of Urology. Guidelines on Urological Infections 2013. Eur. Assoc. Urol. 2013. Available online: https://uroweb.org/guidelines/urological-infections (accessed on 3 April 2022).
- Bolocan, A.S.; Callanan, J.; Forde, A.; Ross, P.; Hill, C. Phage therapy targeting Escherichia coli-a story with no end? FEMS Microbiol. Lett. 2016, 363, fnw256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabal, A.; García-Castillo, M.; Cantón, R.; Gortázar, C.; Domínguez, L.; Álvarez, J. Prevalence of Escherichia coli virulence genes in patients with diarrhea and a subpopulation of healthy volunteers in Madrid, Spain. Front. Microbiol. 2016, 7, 641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaffer, J.N.; Pearson, M.M. Proteus mirabilis and Urinary Tract Infections. Microbiol. Spectr. 2015, 3, 383–433. [Google Scholar] [CrossRef] [Green Version]
- Gomaa, S.; Serry, F.; Abdellatif, H.; Abbas, H. Elimination of multidrug-resistant Proteus mirabilis biofilms using bacteriophages. Arch. Virol. 2019, 164, 2265–2275. [Google Scholar] [CrossRef]
- Pormohammad, A.; Nasiri, M.J.; Azimi, T. Prevalence of antibiotic resistance in escherichia coli strains simultaneously isolated from humans, animals, food, and the environment: A systematic review and meta-analysis. Infect. Drug Resist. 2019, 12, 1181–1197. [Google Scholar] [CrossRef] [Green Version]
- Fasugba, O.; Gardner, A.; Mitchell, B.G.; Mnatzaganian, G. Ciprofloxacin resistance in community- and hospital-acquired Escherichia coli urinary tract infections: A systematic review and meta-analysis of observational studies. BMC Infect. Dis. 2015, 15, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Kot, B.; Grużewska, A.; Szweda, P.; Wicha, J.; Parulska, U. Antibiotic resistance of uropathogens isolated from patients hospitalized in district hospital in central Poland in 2020. Antibiotics 2021, 10, 447. [Google Scholar] [CrossRef] [PubMed]
- Adamus-Bialek, W.; Zajac, E.; Parniewski, P.; Kaca, W. Comparison of antibiotic resistance patterns in collections of Escherichia coli and Proteus mirabilis uropathogenic strains. Mol. Biol. Rep. 2013, 40, 3429–3435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajdács, M.; Bátori, Z.; Burián, K. Interplay between phenotypic resistance to relevant antibiotics in gram-negative urinary pathogens: A data-driven analysis of 10 years’ worth of antibiogram data. Life 2021, 11, 1059. [Google Scholar] [CrossRef] [PubMed]
- Gajdács, M.; Urbán, E. Comparative epidemiology and resistance trends of Proteae in urinary tract infections of inpatients and outpatients: A 10-year retrospective study. Antibiotics 2019, 8, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Summers, W.C. Bacteriophage therapy. Annu. Rev. Microbiol. 2001, 55, 437–451. [Google Scholar] [CrossRef] [Green Version]
- Deresinski, S. Bacteriophage therapy: Exploiting smaller fleas. Clin. Infect. Dis. 2009, 48, 1096–1101. [Google Scholar] [CrossRef] [Green Version]
- Monk, A.B.; Rees, C.D.; Barrow, P.; Hagens, S.; Harper, D.R. Bacteriophage applications: Where are we now? Lett. Appl. Microbiol. 2010, 51, 363–369. [Google Scholar] [CrossRef]
- Gill, J.; Hyman, P. Phage Choice, Isolation, and Preparation for Phage Therapy. Curr. Pharm. Biotechnol. 2010, 11, 2–14. [Google Scholar] [CrossRef]
- Kutter, E.; De Vos, D.; Gvasalia, G.; Alavidze, Z.; Gogokhia, L.; Kuhl, S.; Abedon, S. Phage Therapy in Clinical Practice: Treatment of Human Infections. Curr. Pharm. Biotechnol. 2010, 11, 69–86. [Google Scholar] [CrossRef]
- Leitner, L.; Ujmajuridze, A.; Chanishvili, N.; Goderdzishvili, M.; Chkonia, I.; Rigvava, S.; Chkhotua, A.; Changashvili, G.; McCallin, S.; Schneider, M.P.; et al. Intravesical Bacteriophages for Treating Urinary Tract Infections in Patients Undergoing Transurethral Resection of the Prostate: A Randomised, Placebo-Controlled, Double-Blind Clinical Trial. Lancet Infect. Dis. 2021, 21, 427–436. [Google Scholar] [CrossRef]
- Leitner, L.; Kessler, T.M.; Klumpp, J. Bacteriophages: A Panacea in Neuro-Urology? Eur. Urol. Focus 2020, 6, 518–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirnay, J.P.; Kutter, E. Bacteriophages: It’s a medicine, Jim, but not as we know it. Lancet Infect. Dis. 2021, 21, 309–311. [Google Scholar] [CrossRef]
- Buckling, A.; Rainey, P.B. Antagonistic coevolution between a bacterium and a bacteriophage. Proc. R. Soc. B Biol. Sci. 2002, 269, 931–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clokie, M.R.J.; Kropinski, A.M. Bacteriophages: Methods and Protocols; Humana: Louisville, KY, USA, 2009. [Google Scholar]
- Braissant, O.; Wirz, D.; Göpfert, B.; Daniels, A.U. Use of isothermal microcalorimetry to monitor microbial activities. FEMS Microbiol. Lett. 2010, 303, 1–8. [Google Scholar] [CrossRef]
- Wadsö, I. Isothermal microcalorimetry in applied biology. Thermochim. Acta 2002, 394, 305–311. [Google Scholar] [CrossRef]
- Maskow, T.; Harms, H. Real time insights into bioprocesses using calorimetry: State of the art and potential. Eng. Life Sci. 2006, 6, 266–277. [Google Scholar] [CrossRef]
- Tkhilaishvili, T.; Wang, L.; Tavanti, A.; Trampuz, A.; Di Luca, M. Antibacterial efficacy of two commercially available bacteriophage formulations, staphylococcal bacteriophage and PYO bacteriophage, against methicillin-resistant Staphylococcus aureus: Prevention and eradication of biofilm formation and control of a systemic infection of Galleria mellonella larvae. Front. Microbiol. 2020, 11, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tkhilaishvili, T.; Di Luca, M.; Abbandonato, G.; Maiolo, E.M.; Klatt, A.B.; Reuter, M.; Möncke-Buchner, E.; Trampuz, A. Real-time assessment of bacteriophage T3-derived antimicrobial activity against planktonic and biofilm-embedded Escherichia coli by isothermal microcalorimetry. Res. Microbiol. 2018, 169, 515–521. [Google Scholar] [CrossRef]
- Guosheng, L.; Yi, L.; Xiangdong, C.; Peng, L.; Ping, S.; Songsheng, Q. Study on interaction between T4 phage and Escherichia coli B by microcalorimetric method. J. Virol. Methods 2003, 112, 137–143. [Google Scholar] [CrossRef]
- Morais, F.M.; Buchholz, F.; Hartmann, T.; Lerchner, J.; Neu, T.R.; Kiesel, B.; Harms, H.; Maskow, T. Chip-calorimetric monitoring of biofilm eradication with bacteriophages reveals an unexpected infection-related heat profile. J. Therm. Anal. Calorim. 2014, 115, 2203–2210. [Google Scholar] [CrossRef]
- Tan, A.M.; Lu, J.H. Microcalorimetric study of antiviral effect of drug. J. Biochem. Biophys. Methods 1999, 38, 225–228. [Google Scholar] [CrossRef]
- McCallin, S.; Sarker, S.A.; Sultana, S.; Oechslin, F.; Brüssow, H. Metagenome analysis of Russian and Georgian Pyophage cocktails and a placebo-controlled safety trial of single phage versus phage cocktail in healthy Staphylococcus aureus carriers. Environ. Microbiol. 2018, 20, 3278–3293. [Google Scholar] [CrossRef] [PubMed]
- Sarker, S.A.; Sultana, S.; Reuteler, G.; Moine, D.; Descombes, P.; Charton, F.; Bourdin, G.; McCallin, S.; Ngom-Bru, C.; Neville, T.; et al. Oral Phage Therapy of Acute Bacterial Diarrhea with Two Coliphage Preparations: A Randomized Trial in Children From Bangladesh. EBioMedicine 2016, 4, 124–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; ISBN 3-900051-07-0. [Google Scholar] [CrossRef] [Green Version]
- Kahm, M.; Hasenbrink, G.; Lichtenberg-Fraté, H.; Ludwig, J.; Kschischo, M. Grofit: Fitting Biological Growth Curves with R. J. Stat. Softw. 2010, 33, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Cosmidis, J.; Benzerara, K.; Guyot, F.; Skouri-Panet, F.; Duprat, E.; Férard, C.; Guigner, J.M.; Babonneau, F.; Coelho, C. Calcium-phosphate biomineralization induced by alkaline phosphatase activity in Escherichia coli: Localization, kinetics, and potential signatures in the fossil record. Front. Earth Sci. 2015, 3, 84. [Google Scholar] [CrossRef]
- Cohen, M.S.; Davis, C.P.; Czerwinski, E.W.; Warren, M.M. Calcium phosphate crystal formation in Escherichia coli from human urine: An in vitro study. J. Urol. 1982, 127, 184–185. [Google Scholar] [CrossRef]
- Edin-Liljegren, A.; Hedelin, H.H.; Grenabo, L.; Pettersson, S.; McLean, R.J.C. Impact of Escherichia coli on urine citrate and urease-induced crystallization. Scanning Microsc. 1995, 9, 24. [Google Scholar]
- Rivadeneyra, M.A.; Gutierrez-Calderón, A.; Rivadeneyra, A.M.; Ramos-Cormenzana, A. A study of struvite precipitation and urease activity in bacteria isolated from patients with urinary infections and their possible involvement in the formation of renal calculi. Urol. Int. 1999, 63, 188–192. [Google Scholar] [CrossRef]
- Gelman, D.; Yerushalmy, O.; Alkalay-Oren, S.; Rakov, C.; Ben-Porat, S.; Khalifa, L.; Adler, K.; Abdalrhman, M.; Coppenhagen-Glazer, S.; Aslam, S.; et al. Clinical Phage Microbiology: A suggested framework and recommendations for the in-vitro matching steps of phage therapy. Lancet Microbe 2021, 2, e555–e563. [Google Scholar] [CrossRef]
- McCallin, S.; Brüssow, H. Phage therapy: An alternative or adjunct to antibiotics? Emerg. Top. Life Sci. 2017, 1, 105–116. [Google Scholar]
- Chadha, P.; Katare, O.P.; Chhibber, S. In vivo efficacy of single phage versus phage cocktail in resolving burn wound infection in BALB/c mice. Microb. Pathog. 2016, 99, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Friman, V.P.; Soanes-Brown, D.; Sierocinski, P.; Molin, S.; Johansen, H.K.; Merabishvili, M.; Pirnay, J.P.; De Vos, D.; Buckling, A. Pre-adapting parasitic phages to a pathogen leads to increased pathogen clearance and lowered resistance evolution with Pseudomonas aeruginosa cystic fibrosis bacterial isolates. J. Evol. Biol. 2016, 29, 188–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braissant, O.; Müller, G.; Egli, A.; Widmer, A.; Frei, R.; Halla, A.; Wirz, D.; Gasser, T.C.; Bachmann, A.; Wagenlehner, F.; et al. Seven hours to adequate antimicrobial therapy in urosepsis using isothermal microcalorimetry. J. Clin. Microbiol. 2014, 52, 624–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Górski, A.; Borysowski, J.; Międzybrodzki, R. Phage therapy: Towards a successful clinical trial. Antibiotics 2020, 9, 827. [Google Scholar] [CrossRef] [PubMed]
- Jeon, G.; Ahn, J. Assessment of phage-mediated inhibition of Salmonella Typhimurium treated with sublethal concentrations of ceftriaxone and ciprofloxacin. FEMS Microbiol. Lett. 2020, 367, fnaa159. [Google Scholar] [CrossRef]
- Luscher, A.; Simonin, J.; Falconnet, L.; Valot, B.; Hocquet, D.; Chanson, M.; Resch, G.; Köhler, T.; van Delden, C. Combined Bacteriophage and Antibiotic Treatment Prevents Pseudomonas aeruginosa Infection of Wild Type and cftr- Epithelial Cells. Front. Microbiol. 2020, 11, 1947. [Google Scholar] [CrossRef] [PubMed]
- Bonkat, G.; Braissant, O.; Rieken, M.; Solokhina, A.; Widmer, A.F.; Frei, R.; van der Merwe, A.; Wyler, S.; Gasser, T.C.; Bachmann, A. Standardization of isothermal microcalorimetry in urinary tract infection detection by using artificial urine. World J. Urol. 2013, 31, 553–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, L.; Sánchez-Juanes, F.; González-Ávila, M.; Cembrero-Fuciños, D.; Herrero-Hernández, A.; González-Buitrago, J.M.; Muñoz-Bellido, J.L. Direct identification of urinary tract pathogens from urine samples by matrix-assisted laser desorption ionization-time of flight mass spectrometry. J. Clin. Microbiol. 2010, 48, 2110–2115. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, K.L.; Dynesen, P.; Larsen, P.; Jakobsen, L.; Andersen, P.S.; Frimodt-Møller, N. Role of urinary cathelicidin LL-37 and human β-defensin 1 in uncomplicated escherichia coli urinary tract infections. Infect. Immun. 2014, 82, 1572–1578. [Google Scholar] [CrossRef] [Green Version]
- Stern, A.; Sorek, R. The phage-host arms race: Shaping the evolution of microbes. BioEssays 2011, 33, 43–51. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| E. coli | ||||
|---|---|---|---|---|
| Sample | μ (h−1) | λ (h) | Q (J) | TTP (h) |
| Control | 0.18 ± 0.01 | 2.08 ± 0.18 | 1.93 ± 0.11 | 5.67 ± 0.12 |
| 1% | 0.08 ± 0.01 | 0.00 ± 0.16 | 0.89 ± 0.16 | 1.50 ± 0.00 |
| 5% | 0.04 ± 0.01 | 0.00 ± 2.69 | 0.57 ± 0.02 | 1.52 ± 0.04 |
| P. mirabilis | ||||
| Sample | μ (h−1) | λ (h) | Q (J) | TTP (h) |
| Control | 1.28 ± 0.05 | 0.0 ± 0.3 | 28.89 ± 0.3 | 4.00 ± 0.3 |
| 1% | 0.66 ± 0.00 | 0.0 ± 0.0 | 21.09 ± 0.1 | 3.41 ± 0.0 |
| 5% | 0.59 ± 0.08 | 4.6 ± 0.4 | 19.6 ± 1.6 | 12.5 ± 0.2 |
| E. coli | First Peak | Second Peak | Third Peak | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Sample | μ (h−1) | λ (h) | Q (J) | μ (h−1) | λ (h) | Q (J) | μ (h−1) | λ (h) | Q (J) |
| Control | 0.29 ± 0.01 | 0.9 ± 0.2 | 7.0 ± 0.2 | ||||||
| 1% | 0.04 ± 0.02 | 17.8 ± 3.6 | 0.7 ± 0.5 | 0.09 ± 0.04 | 63.4 ± 12.1 | 1.6 ± 1.0 | |||
| 5% | 0.02 ± 0.01 | 18.3 ± 2.6 | 0.2 ± 0.1 | 0.03 ± 0.03 | 48.3 ± 32.1 | 0.5 ± 0.3 | 0.03 | 60.5 | 0.6 |
| P. mirabilis | |||||||||
| Sample | μ (h−1) | λ (h) | Q (J) | μ (h−1) | λ (h) | Q (J) | μ (h−1) | λ (h) | Q (J) |
| Control | 0.62 ± 0.04 | 1.1 ± 0.5 | 5.3 ± 0.5 | ||||||
| 1% | 0.11 ± 0.01 | 4.0 ± 1.1 | 2.9 ± 0.2 | ||||||
| 5% | 0.04 ± 0.02 | 0.0 ± 7.1 | 1.3 ± 0.0 | ||||||
| P. mirabilis + TMP/SMX | ||||
|---|---|---|---|---|
| Sample | μ (h−1) | λ (h) | Q (J) | TTP (h) |
| Control | 0.54 ± 0.01 | 0.0 ± 0.0 | 5.2 ± 0.1 | 3.2 ± 0.1 |
| 1% phage | 0.11 ± 0.01 | 4.0 ± 1.1 | 2.9 ± 0.2 | 14.2 ± 0.1 |
| 1% phage + 50% MIC | 0.01 ± 0.01 | 0.0 ± 13.6 | 1.0 ± 0.7 | 0.6 ± 0.4 |
| 1% phage +25% MIC | 0.01 ± 0.01 | 0.0 ± 11.9 | 0.8 ± 0.7 | 0.7 ± 0.5 |
| TMP/SMX: 50% MIC | 0.31 ± 0.01 | 0.0 ± 0.2 | 3.6 ± 0.1 | 1.3 ± 0.0 |
| Blanks | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sigg, A.P.; Mariotti, M.; Grütter, A.E.; Lafranca, T.; Leitner, L.; Bonkat, G.; Braissant, O. A Method to Determine the Efficacy of a Commercial Phage Preparation against Uropathogens in Urine and Artificial Urine Determined by Isothermal Microcalorimetry. Microorganisms 2022, 10, 845. https://doi.org/10.3390/microorganisms10050845
Sigg AP, Mariotti M, Grütter AE, Lafranca T, Leitner L, Bonkat G, Braissant O. A Method to Determine the Efficacy of a Commercial Phage Preparation against Uropathogens in Urine and Artificial Urine Determined by Isothermal Microcalorimetry. Microorganisms. 2022; 10(5):845. https://doi.org/10.3390/microorganisms10050845
Chicago/Turabian StyleSigg, Aurelia Pahnita, Max Mariotti, Anabel E. Grütter, Tecla Lafranca, Lorenz Leitner, Gernot Bonkat, and Olivier Braissant. 2022. "A Method to Determine the Efficacy of a Commercial Phage Preparation against Uropathogens in Urine and Artificial Urine Determined by Isothermal Microcalorimetry" Microorganisms 10, no. 5: 845. https://doi.org/10.3390/microorganisms10050845