Roux-en-Y Gastric Bypass Surgery Induces Distinct but Frequently Transient Effects on Acylcarnitine, Bile Acid and Phospholipid Levels

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Cohort and Sample Collection
2.2. Biochemical Analyses
2.3. Metabolite Profiling
2.4. Statistical Analysis
2.5. Ethical Approval
3. Results
3.1. Clinical Parameters
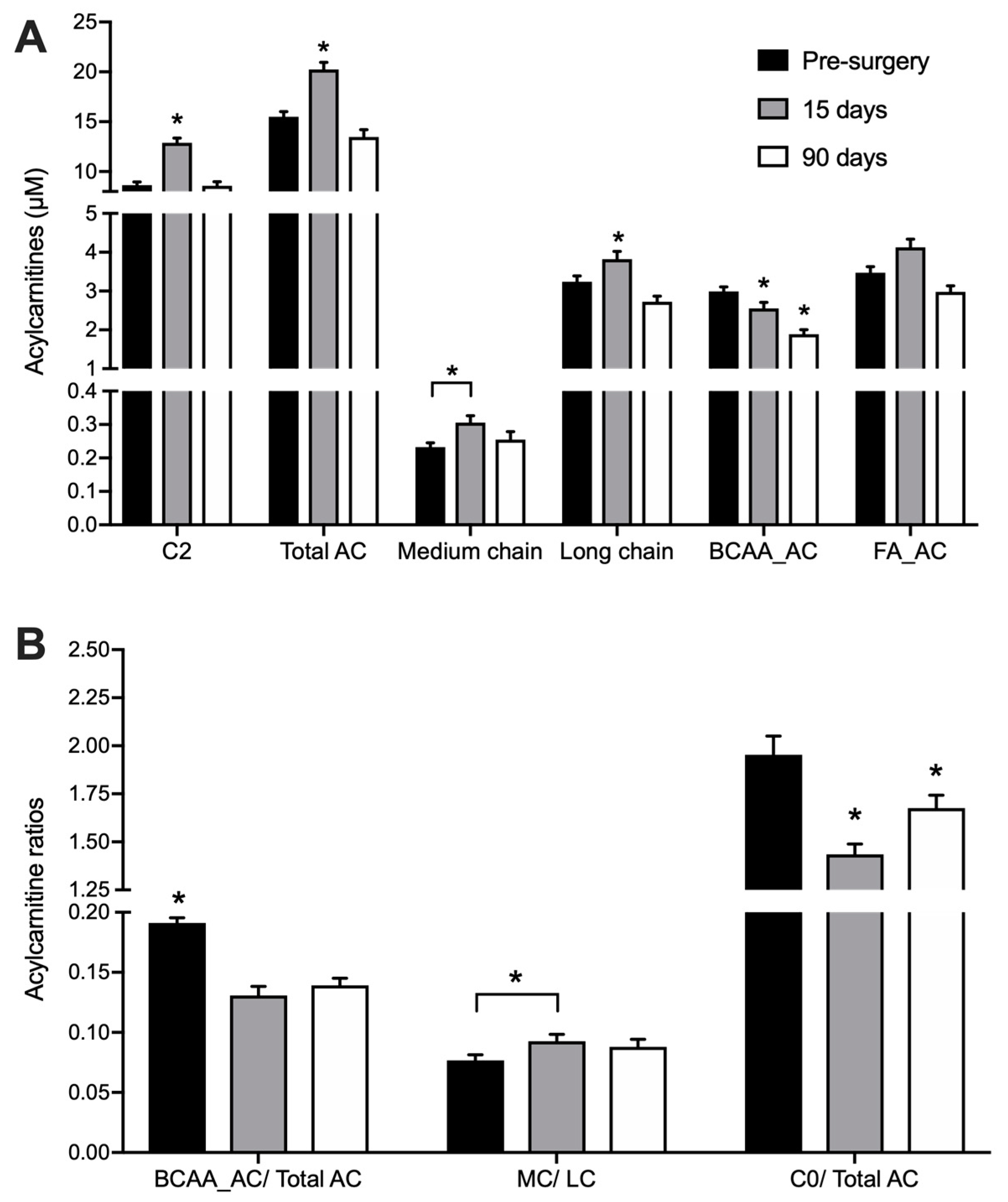
3.2. Acylcarnitine Profile
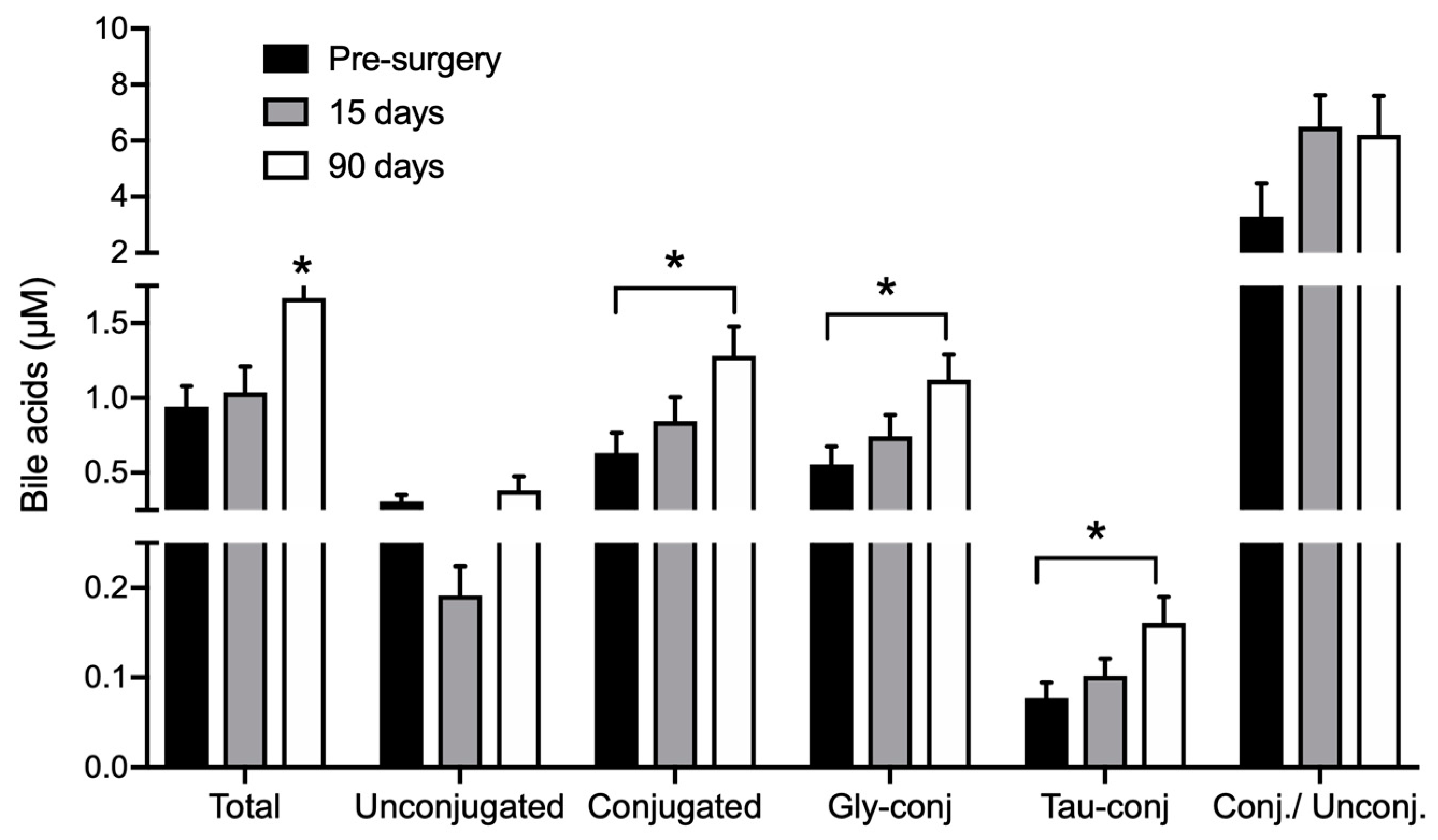
3.3. Bile Acids Profile
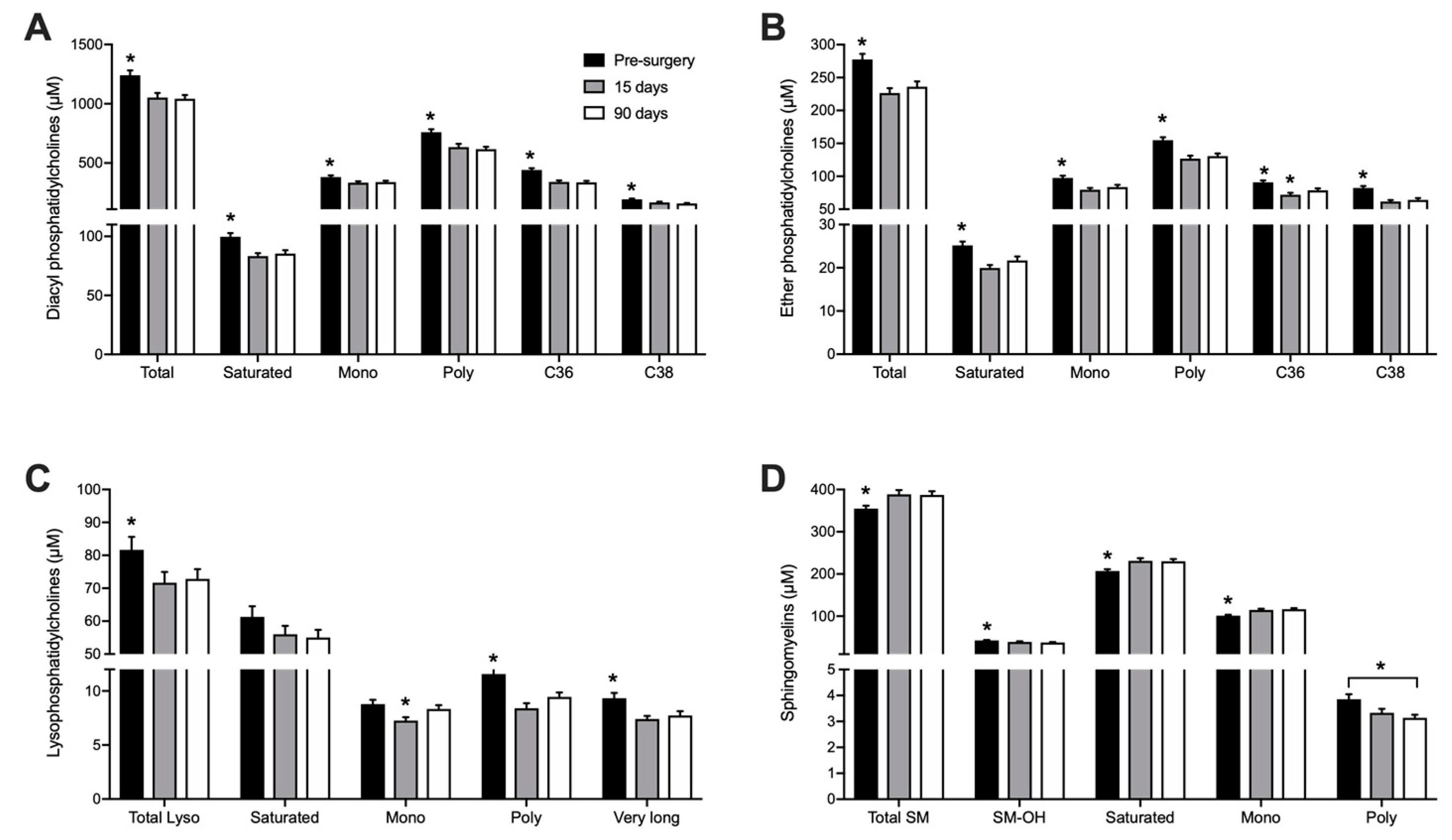
3.4. Phospholipid Profile
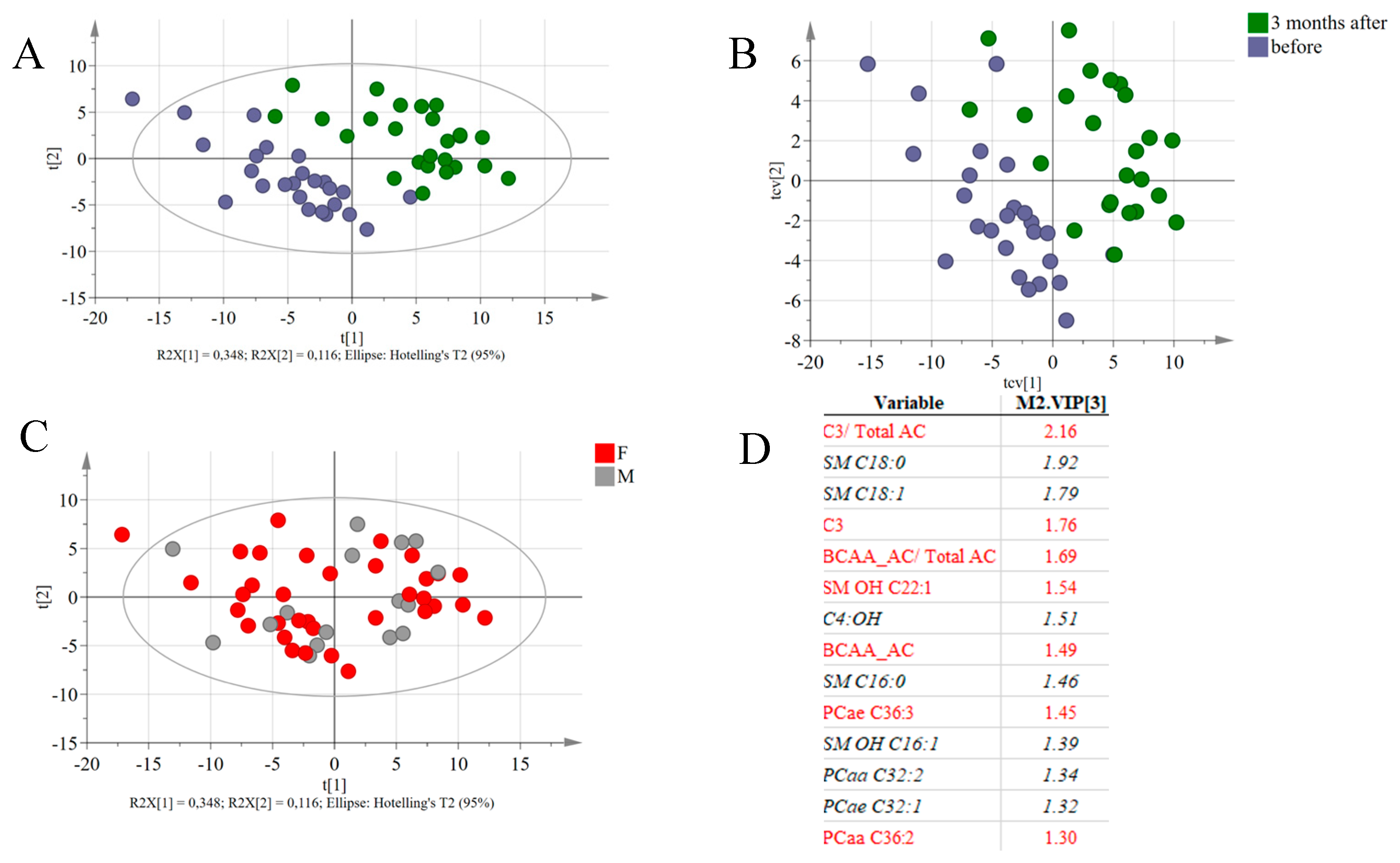
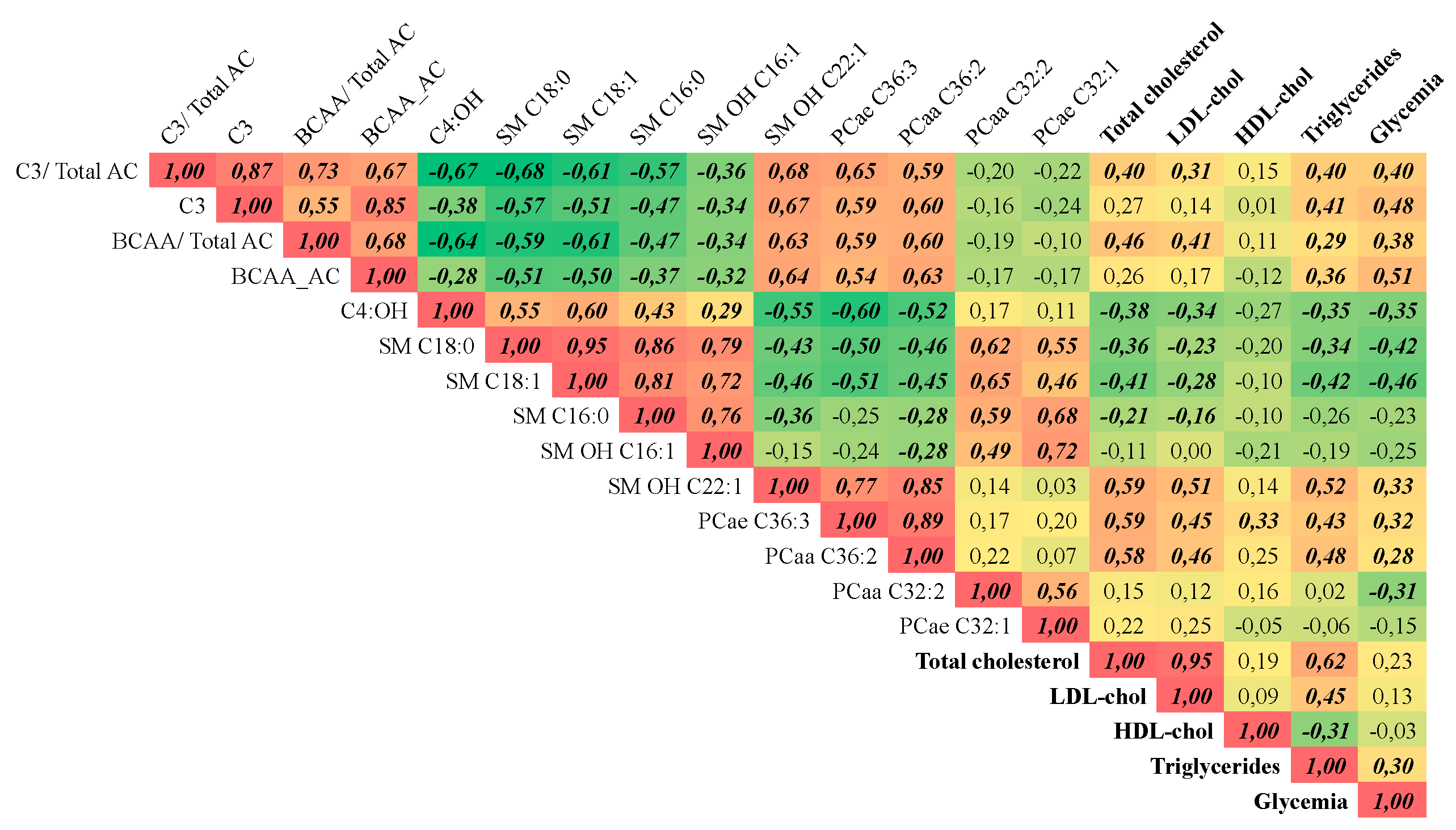
3.5. Global Metabolic Responses to RYGB
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | Pre-Surgery | 15 Days After | 90 Days After |
|---|---|---|---|
| L-carnitine | 30.034 ± 1.764 | 27.302 ± 1.583 | 23.611 ± 1.117 a,b |
| C4-OH | 0.027 ± 0.001 | 0.217 ± 0.022 a | 0.06 ± 0.004 a,b |
| C2 | 8.756 ± 0.369 | 11.956 ± 0.526 a | 8.81 ± 0.485 b |
| C5-OH | 0.222 ± 0.023 | 0.232 ± 0.021 | 0.164 ± 0.019 a,b |
| C3 | 1.617 ± 0.084 | 1.224 ± 0.119 a | 0.855 ± 0.051 a,b |
| 2M-C3 | 0.116 ± 0.009 | 0.183 ± 0.015 a | 0.098 ± 0.006 b |
| C4 | 0.058 ± 0.008 | 0.05 ± 0.006 a | 0.041 ± 0.004 a,b |
| 2M-C4 | 0.064 ± 0.004 | 0.07 ± 0.005 | 0.039 ± 0.002 a,b |
| 3M-C4 | 0.112 ± 0.006 | 0.119 ± 0.009 | 0.089 ± 0.006 a,b |
| C4DC | 1.038 ± 0.069 | 0.881 ± 0.072 a | 0.882 ± 0.072 a |
| C3DC-M | 0.012 ± 0.001 | 0.009 ± 0.001 a | 0.011 ± 0 |
| C6 | 0.034 ± 0.003 | 0.041 ± 0.003 | 0.034 ± 0.002 |
| C5DC | 0.029 ± 0.003 | 0.021 ± 0.002 a | 0.028 ± 0.002 b |
| C6DC | 0.007 ± 0.001 | 0.027 ± 0.003 a | 0.014 ± 0.001 a,b |
| C8 | 0.032 ± 0.003 | 0.045 ± 0.004 a | 0.042 ± 0.004 |
| C10 | 0.068 ± 0.007 | 0.1 ± 0.011 a | 0.097 ± 0.013 a |
| C12 | 0.033 ± 0.002 | 0.042 ± 0.004 a | 0.036 ± 0.002 |
| C14 | 0.068 ± 0.004 | 0.069 ± 0.005 | 0.054 ± 0.002 a,b |
| C16:1 | 0.092 ± 0.005 | 0.127 ± 0.012 a | 0.098 ± 0.005 b |
| C18:2 | 0.445 ± 0.031 | 0.456 ± 0.03 | 0.374 ± 0.026 a,b |
| C16 | 1.177 ± 0.075 | 1.295 ± 0.095 a | 1.039 ± 0.044 a,b |
| C18:1 | 1.002 ± 0.063 | 1.33 ± 0.091 a | 1.043 ± 0.055 b |
| C18 | 0.438 ± 0.037 | 0.37 ± 0.035 a | 0.329 ± 0.023 a |
| Metabolites | Pre-Surgery | 15 Days After | 90 Days After |
|---|---|---|---|
| CDCA | 0.117 ± 0.023 | 0.099 ± 0.023 | 0.118 ± 0.034 |
| DCA | 0.155 ± 0.02 | 0.089 ± 0.015 a | 0.183 ± 0.024 b |
| CA | 0.045 ± 0.013 | 0.035 ± 0.007 | 0.092 ± 0.047 |
| GCDCA | 0.315 ± 0.068 | 0.441 ± 0.088 | 0.688 ± 0.102 a |
| GDCA | 0.097 ± 0.02 | 0.135 ± 0.033 | 0.205 ± 0.053 |
| GUDCA | 0.033 ± 0.006 | 0.058 ± 0.015 | 0.034 ± 0.008 |
| GCA | 0.109 ± 0.032 | 0.107 ± 0.024 | 0.194 ± 0.03 |
| TCDCA | 0.039 ± 0.009 | 0.055 ± 0.011 | 0.091 ± 0.018 a |
| TDCA | 0.014 ± 0.003 | 0.02 ± 0.005 | 0.034 ± 0.01 |
| TUDCA | 0.002 ± 0.0007 | 0.002 ± 0.0008 | 0.001 ± 0.0001 |
| TCA | 0.022 ± 0.005 | 0.024 ± 0.004 | 0.033 ± 0.004 a |
| Metabolites | Pre-Surgery | 15 Days After | 90 Days After |
|---|---|---|---|
| Lysophosphatidylcholines | |||
| Lyso PC a C14:0 | 2.16 ± 0.07 | 1.7 ± 0.04 a | 1.78 ± 0.07 a |
| Lyso PC a C16:0 | 39.61 ± 2.21 | 40.09 ± 2.11 | 37.69 ± 1.67 |
| Lyso PC a C16:1 | 1.07 ± 0.07 | 0.68 ± 0.04 a | 0.85 ± 0.05 a,b |
| Lyso PC a C17:0 | 1.15 ± 0.04 | 1.09 ± 0.04 | 1.11 ± 0.05 |
| Lyso PC a C18:0 | 12.67 ± 0.72 | 8.47 ± 0.48 a | 9.72 ± 0.46 a,b |
| Lyso PC a C18:1 | 5.73 ± 0.27 | 4.7 ± 0.26 a | 5.69 ± 0.25 b |
| Lyso PC a C18:2 | 7.94 ± 0.41 | 4.96 ± 0.35 a | 6.24 ± 0.28 a,b |
| Lyso PC a C18:3 | 0.25 ± 0.01 | 0.12 ± 0.01 a | 0.15 ± 0.01 a,b |
| Lyso PC a C20:3 | 0.79 ± 0.05 | 0.39 ± 0.02 a | 0.47 ± 0.03 a,b |
| Lyso PC a C20:4 | 1.72 ± 0.09 | 1.93 ± 0.11 | 1.8 ± 0.1 |
| Lyso PC a C20:5 | 0.54 ± 0.02 | 0.46 ± 0.02 a | 0.42 ± 0.02 a |
| Lyso PC a C22:5 | 0.17 ± 0.01 | 0.14 ± 0.01 | 0.16 ± 0.01 |
| Lyso PC a C22:6 | 0.36 ± 0.02 | 0.38 ± 0.02 | 0.34 ± 0.02 b |
| Lyso PC a C24:0 | 1.61 ± 0.07 | 1.48 ± 0.06 | 1.4 ± 0.06 a |
| Lyso PC a C26:0 | 2.99 ± 0.18 | 2.13 ± 0.1 a | 2.26 ± 0.15 a |
| Lyso PC a C26:1 | 0.92 ± 0.03 | 0.83 ± 0.03 | 0.85 ± 0.03 |
| Lyso PC a C28:0 | 2.23 ± 0.14 | 1.43 ± 0.07 a | 1.61 ± 0.1 a |
| Lyso PC a C28:1 | 1.24 ± 0.07 | 1.07 ± 0.05 | 1.06 ± 0.06 a |
| Total Lyso PC | 83.14 ± 3.8 | 72.05 ± 3.38 a | 73.52 ± 2.99 a |
| Diacyl Phosphatidylcholines | |||
| PC aa C24:0 | 1.63 ± 0.08 | 1.6 ± 0.06 | 1.44 ± 0.05 a |
| PC aa C26:0 | 3.25 ± 0.14 | 2.66 ± 0.11 a | 2.85 ± 0.18 |
| PC aa C28:1 | 3.13 ± 0.1 | 2.63 ± 0.1 a | 2.62 ± 0.09 a |
| PC aa C30:0 | 5.96 ± 0.21 | 4.93 ± 0.17 a | 5.17 ± 0.15 a |
| PC aa C30:2 | 5.24 ± 0.15 | 5.7 ± 0.22 | 5.51 ± 0.18 |
| PC aa C32:0 | 33 ± 0.92 | 33.68 ± 1.08 | 34.68 ± 0.78 |
| PC aa C32:1 | 23.49 ± 1.22 | 20.82 ± 0.79 | 22.49 ± 0.92 |
| PC aa C32:2 | 6.6 ± 0.2 | 7.1 ± 0.31 | 7.1 ± 0.22 |
| PC aa C32:3 | 0.79 ± 0.03 | 0.82 ± 0.04 | 0.79 ± 0.03 |
| PC aa C34:1 | 215.38 ± 9.04 | 201.78 ± 7.9 | 204.45 ± 7.12 |
| PC aa C34:2 | 260.09 ± 9.01 | 218.43 ± 11.39 a | 218.85 ± 9.12 a |
| PC aa C34:3 | 9.48 ± 0.36 | 5.1 ± 0.24 a | 6.37 ± 0.38 a,b |
| PC aa C34:4 | 0.95 ± 0.05 | 0.46 ± 0.02 a | 0.54 ± 0.03 a,b |
| PC aa C36:0 | 36.82 ± 1.39 | 28.37 ± 1.25 a | 30.02 ± 1.49 a |
| PC aa C36:1 | 91.27 ± 3.25 | 65.32 ± 2.33 a | 68.49 ± 3.09 a |
| PC aa C36:2 | 164.82 ± 6.44 | 96.06 ± 4.7 a | 111.44 ± 5.75 a,b |
| PC aa C36:3 | 72.14 ± 3.49 | 46.63 ± 2.45 a | 50.28 ± 2.48 a |
| PC aa C36:4 | 73.74 ± 4.01 | 98.21 ± 6.19 a | 69.46 ± 2.28 b |
| PC aa C36:5 | 5.67 ± 0.43 | 3.5 ± 0.2 a | 3.55 ± 0.18 a |
| PC aa C36:6 | 1.32 ± 0.05 | 1.06 ± 0.04 a | 1.17 ± 0.06 b |
| PC aa C38:0 | 18.12 ± 0.76 | 10.17 ± 0.46 a | 11.91 ± 0.73 a,b |
| PC aa C38:1 | 45.05 ± 1.16 | 36.62 ± 1.14 a | 34.54 ± 1.34 a |
| PC aa C38:3 | 43.04 ± 1.72 | 30.8 ± 1.22 a | 32.09 ± 1 a |
| PC aa C38:4 | 50.57 ± 2.36 | 47.66 ± 2.67 | 42.51 ± 1.57 a |
| PC aa C38:5 | 19.43 ± 0.88 | 18.12 ± 0.98 | 17.73 ± 0.63 |
| PC aa C38:6 | 19.29 ± 0.74 | 22.87 ± 1.45 a | 18.51 ± 0.61 b |
| PC aa C40:1 | 6.12 ± 0.2 | 5.23 ± 0.16 a | 4.98 ± 0.15 a |
| PC aa C40:2 | 4.23 ± 0.17 | 3.82 ± 0.14 a | 3.85 ± 0.17 a |
| PC aa C40:3 | 3.82 ± 0.12 | 3.24 ± 0.1 a | 3.39 ± 0.09 a |
| PC aa C40:4 | 3.86 ± 0.15 | 3.14 ± 0.11 a | 3.03 ± 0.09 a |
| PC aa C40:5 | 5.73 ± 0.26 | 4.04 ± 0.19 a | 4.37 ± 0.18 a |
| PC aa C40:6 | 9.69 ± 0.39 | 7.48 ± 0.38 a | 7.3 ± 0.3 a |
| PC aa C42:0 | 2.4 ± 0.09 | 2.04 ± 0.07 a | 2.03 ± 0.08 a |
| PC aa C42:1 | 1.91 ± 0.07 | 1.65 ± 0.06 a | 1.71 ± 0.07 a |
| PC aa C42:2 | 1.44 ± 0.06 | 1.41 ± 0.06 | 1.33 ± 0.05 a |
| PC aa C42:4 | 0.74 ± 0.03 | 0.61 ± 0.02 a | 0.62 ± 0.02 a |
| PC aa C42:5 | 0.91 ± 0.04 | 0.85 ± 0.03 | 0.8 ± 0.03 a |
| PC aa C42:6 | 2.24 ± 0.09 | 2.32 ± 0.09 | 2.05 ± 0.1 b |
| Total PC | 1253.36 ± 40.09 | 1046.94 ± 39.25 a | 1036.32 ± 31.58 a |
| Ether Phosphatidylcholines | |||
| PC ae C30:0 | 1.18 ± 0.04 | 0.88 ± 0.03 a | 0.96 ± 0.04 a |
| PC ae C30:1 | 2.62 ± 0.07 | 2.54 ± 0.09 | 2.53 ± 0.08 |
| PC ae C30:2 | 1.04 ± 0.05 | 0.91 ± 0.04 | 0.87 ± 0.03 a |
| PC ae C32:1 | 3.95 ± 0.11 | 4.11 ± 0.15 | 4.38 ± 0.14 a |
| PC ae C32:2 | 1.02 ± 0.04 | 0.96 ± 0.03 | 1 ± 0.04 |
| PC ae C34:0 | 5.43 ± 0.16 | 4.79 ± 0.16 a | 4.76 ± 0.14 a |
| PC ae C34:1 | 11 ± 0.37 | 9.61 ± 0.4 a | 10.55 ± 0.38 b |
| PC ae C34:2 | 7.81 ± 0.2 | 5.34 ± 0.22 a | 6.71 ± 0.23 a,b |
| PC ae C34:3 | 3.46 ± 0.11 | 2.44 ± 0.11 a | 3.36 ± 0.11 b |
| PC ae C36:0 | 9.12 ± 0.37 | 7.76 ± 0.32 a | 8.71 ± 0.43 b |
| PC ae C36:1 | 38.55 ± 1.41 | 30.91 ± 1.33 a | 33.67 ± 1.53 a |
| PC ae C36:2 | 28.54 ± 1.05 | 21.57 ± 0.96 a | 23.54 ± 1.18 a |
| PC ae C36:3 | 5.15 ± 0.14 | 3.27 ± 0.14 a | 3.8 ± 0.12 a,b |
| PC ae C36:4 | 7.04 ± 0.27 | 5.43 ± 0.28 a | 5.65 ± 0.17 a |
| PC ae C36:5 | 3.65 ± 0.18 | 2.93 ± 0.16 a | 3.54 ± 0.15 b |
| PC ae C38:0 | 6.75 ± 0.26 | 4.48 ± 0.18 a | 5.16 ± 0.29 a,b |
| PC ae C38:1 | 21.9 ± 0.86 | 13.51 ± 0.58 a | 15.04 ± 0.87 a |
| PC ae C38:2 | 20.78 ± 0.81 | 12.98 ± 0.52 a | 14.92 ± 0.83 a,b |
| PC ae C38:3 | 12.32 ± 0.46 | 11.25 ± 0.53 | 10.29 ± 0.37 a |
| PC ae C38:4 | 8.86 ± 0.32 | 8.23 ± 0.42 | 7.2 ± 0.18 a,b |
| PC ae C38:5 | 6.3 ± 0.24 | 5.99 ± 0.34 | 5.91 ± 0.16 |
| PC ae C38:6 | 6.52 ± 0.2 | 5.15 ± 0.18 a | 5.6 ± 0.22 a |
| PC ae C40:1 | 14.42 ± 0.59 | 13.98 ± 0.56 | 12.48 ± 0.42 a,b |
| PC ae C40:2 | 10.94 ± 0.38 | 11.01 ± 0.41 | 10.01 ± 0.31 |
| PC ae C40:3 | 6.88 ± 0.27 | 5.97 ± 0.24 a | 5.73 ± 0.19 a |
| PC ae C40:4 | 4.41 ± 0.17 | 3.72 ± 0.17 a | 3.57 ± 0.12 a |
| PC ae C40:5 | 3.88 ± 0.13 | 3.94 ± 0.22 | 3.66 ± 0.12 |
| PC ae C40:6 | 2.93 ± 0.08 | 2.19 ± 0.07 a | 2.21 ± 0.14 a |
| PC ae C42:0 | 3.11 ± 0.15 | 2.11 ± 0.08 a | 2.13 ± 0.11 a |
| PC ae C42:1 | 6.57 ± 0.31 | 4.94 ± 0.19 a | 5 ± 0.22 a |
| PC ae C42:2 | 4.73 ± 0.2 | 4.03 ± 0.15 a | 4.07 ± 0.16 a |
| PC ae C42:3 | 2.81 ± 0.11 | 2.6 ± 0.1 | 2.54 ± 0.09 a |
| PC ae C42:4 | 1.61 ± 0.05 | 1.36 ± 0.06 a | 1.32 ± 0.05 a |
| PC ae C42:5 | 1.77 ± 0.05 | 1.52 ± 0.06 a | 1.49 ± 0.05 a |
| PC ae C44:3 | 0.86 ± 0.04 | 0.69 ± 0.03 a | 0.72 ± 0.03 a |
| PC ae C44:4 | 0.72 ± 0.03 | 0.62 ± 0.02 a | 0.62 ± 0.02 a |
| PC ae C44:5 | 1.01 ± 0.03 | 0.97 ± 0.04 | 0.86 ± 0.03 a,b |
| PC ae C44:6 | 1.33 ± 0.05 | 1.17 ± 0.04 a | 1.07 ± 0.04 a |
| Total ether PC | 280.94 ± 8.18 | 71.97 ± 2.56 a | 68.98 ± 2.17 a |
| Sphingomyelins | |||
| SM OH C14:1 | 6.93 ± 0.22 | 7.18 ± 0.26 | 7.03 ± 0.26 |
| SM OH C16:1 | 4.77 ± 0.14 | 5.9 ± 0.22 a | 5.86 ± 0.21 a |
| SM OH C22:1 | 15.81 ± 0.48 | 11.65 ± 0.38 a | 10.6 ± 0.43 a,b |
| SM OH C22:2 | 9.61 ± 0.23 | 9 ± 0.29 | 8.38 ± 0.24 a |
| SM OH C24:1 | 5.81 ± 0.16 | 5.67 ± 0.19 | 5.27 ± 0.15 a |
| SM C16:0 | 120.73 ± 2.77 | 136.64 ± 3.85 a | 141.02 ± 3.18 a |
| SM C16:1 | 14.83 ± 0.39 | 17.92 ± 0.59 a | 16.56 ± 0.48 a |
| SM C18:0 | 24.59 ± 0.73 | 34.76 ± 1.21 a | 35.81 ± 1.21 a |
| SM C18:1 | 11.5 ± 0.39 | 18.74 ± 0.8 a | 17.43 ± 0.66 a |
| SM C20:2 | 0.83 ± 0.03 | 0.78 ± 0.03 | 0.81 ± 0.02 |
| SM C22:3 | 3.05 ± 0.18 | 2.53 ± 0.13 a | 2.27 ± 0.09 a |
| SM C24:0 | 58.58 ± 1.66 | 55.81 ± 1.93 | 48.37 ± 1.48 a,b |
| SM C24:1 | 71.86 ± 1.66 | 74.49 ± 2.19 | 77.58 ± 1.82 a |
| SM C26:0 | 3.88 ± 0.09 | 3.66 ± 0.12 | 3.56 ± 0.09 a |
| SM C26:1 | 3.57 ± 0.09 | 3.49 ± 0.11 | 3.75 ± 0.09 b |
| Total SM | 356.34 ± 6.91 | 462.32 ± 12.25 a | 453.78 ± 8.33 a |
References
- Flegal, K.M.; Kit, B.K.; Orpana, H.; Graubard, B.I. Association of all-cause mortality with overweight and obesity using standard body mass index categories: A systematic review and meta-analysis. JAMA 2013, 309, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, E.A.; Linnan, L.A.; Tate, D.F.; Leese, P.J. A longitudinal study on the relationship between weight loss; medical expenditures; and absenteeism among overweight employees in the WAY to Health study. J. Occup. Environ. Med. 2009, 51, 1367–1373. [Google Scholar] [CrossRef] [PubMed]
- Eldar, S.; Heneghan, H.M.; Brethauer, S.A.; Schauer, P.R. Bariatric surgery for treatment of obesity. Int. J. Obes. 2011, 35 (Suppl. 3), S16–S21. [Google Scholar] [CrossRef] [PubMed]
- Catalán, V.; Gómez-Ambrosi, J.; Ramirez, B.; Rotellar, F.; Pastor, C.; Silva, C.; Rodríguez, A.; Gil, M.J.; Cienfuegos, J.A.; Frühbeck, G. Proinflammatory cytokines in obesity: Impact of type 2 diabetes mellitus and gastric bypass. Obes. Surg. 2007, 17, 1464–1474. [Google Scholar] [CrossRef] [PubMed]
- Holdstock, C.; Lind, L.; Engstrom, B.E.; Ohrvall, M.; Sundbom, M.; Larsson, A.; Karlsson, F.A. CRP reduction following gastric bypass surgery is most pronounced in insulin-sensitive subjects. Int. J. Obes. 2005, 29, 1275–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- le Roux, C.W.; Welbourn, R.; Werling, M.; Osborne, A.; Kokkinos, A.; Laurenius, A.; Lönroth, H.; Fändriks, L.; Ghatei, M.A.; Bloom, S.R.; et al. Gut hormones as mediators of appetite and weight loss after Roux-en-Y gastric bypass. Ann. Surg. 2007, 246, 780–785. [Google Scholar] [CrossRef] [PubMed]
- Rodieux, F.; Giusti, V.; D’Alessio, D.A.; Suter, M.; Tappy, L. Effects of gastric bypass and gastric banding on glucose kinetics and gut hormone release. Obesity 2008, 16, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.B.; Hagedorn, J.C.; Lawson, E.H.; Galanko, J.A.; Safadi, B.Y.; Curet, M.J.; Morton, J.M. Gastric bypass reduces biochemical cardiac risk factors. Surg. Obes. Relat. Dis. 2007, 3, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; Rochon, J.; et al. A branched-chain amino acid-related metabolic signature that differentiates obese and lean humans and contributes to insulin resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef] [PubMed]
- Rauschert, S.; Uhl, O.; Koletzko, B.; Kirchberg, F.; Mori, T.A.; Huang, R.C.; Beilin, L.J.; Hellmuth, C.; Oddy, W.H. Lipidomics Reveals Associations of Phospholipids With Obesity and Insulin Resistance in Young Adults. J. Clin. Endocrinol. Metab. 2016, 101, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Arora, T.; Velagapudi, V.; Pournaras, D.J.; Welbourn, R.; le Roux, C.W.; Orešič, M.; Bäckhed, F. Roux-en-Y Gastric Bypass Surgery Induces Early Plasma Metabolomic and Lipidomic Alterations in Humans Associated with Diabetes Remission. PLoS ONE 2015, 10, e0126401. [Google Scholar] [CrossRef] [PubMed]
- Graessler, J.; Qin, Y.; Zhong, H.; Zhang, J.; Licinio, J.; Wong, M.L.; Xu, A.; Chavakis, T.; Bornstein, A.B.; Ehrhart-Bornstein, M.; et al. Metagenomic sequencing of the human gut microbiome before and after bariatric surgery in obese patients with type 2 diabetes: Correlation with inflammatory and metabolic parameters. Pharmacogenomics J. 2013, 13, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Wijayatunga, N.N.; Sams, V.G.; Dawson, J.A.; Mancini, M.L.; Mancini, G.J.; Moustaid-Moussa, N. Roux-en-Y gastric bypass surgery alters serum metabolites and fatty acids in patients with morbid obesity. Diabetes Metab. Res. Rev. 2018, e3045. [Google Scholar] [CrossRef] [PubMed]
- Mikhalkova, D.; Holman, S.R.; Jiang, H.; Saghir, M.; Novak, E.; Coggan, A.R.; O’Connor, R.; Bashir, A.; Jamal, A.; Ory, D.S.; et al. Bariatric Surgery-Induced Cardiac and Lipidomic Changes in Obesity-Related Heart Failure with Preserved Ejection Fraction. Obesity 2018, 26, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Hall, E.M.; Flores, S.R.; De Jesús, V.R. Influence of Hematocrit and Total-Spot Volume on Performance Characteristics of Dried Blood Spots for Newborn Screening. Int. J. Neonatal Screen. 2015, 1, 69–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Yang, K.; Gross, R.W. Multi-dimensional mass spectrometry-based shotgun lipidomics and novel strategies for lipidomic analyses. Mass Spectrom. Rev. 2012, 31, 134–178. [Google Scholar] [CrossRef] [PubMed]
- Giesbertz, P.; Ecker, J.; Haag, A.; Spanier, B.; Daniel, H. An LC-MS/MS method to quantify acylcarnitine species including isomeric and odd-numbered forms in plasma and tissues. J. Lipid. Res. 2015, 56, 2029–2039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagliacozzi, D.; Mozzi, A.F.; Casetta, B.; Bertucci, P.; Bernardini, S.; Di Ilio, C.; Urbani, A.; Federici, G. Quantitative analysis of bile acids in human plasma by liquid chromatography-electrospray tandem mass spectrometry: A simple and rapid one-step method. Clin. Chem. Lab. Med. 2003, 41, 1633–1641. [Google Scholar] [CrossRef] [PubMed]
- Fiamoncini, J.; Yiorkas, A.M.; Gedrich, K.; Rundle, M.; Alsters, S.I.; Roeselers, G.; van den Broek, T.J.; Clavel, T.; Lagkouvardos, I.; Wopereis, S.; et al. Determinants of postprandial plasma bile acid kinetics in human volunteers. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G300–G312. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, R.; Zhao, X.; Weigert, C.; Simon, P.; Fehrenbach, E.; Fritsche, J.; Machann, J.; Schick, F.; Wang, J.; Hoene, M.M.; et al. Medium chain acylcarnitines dominate the metabolite pattern in humans under moderate intensity exercise and support lipid oxidation. PLoS ONE 2010, 5, e11519. [Google Scholar] [CrossRef] [PubMed]
- Krug, S.; Kastenmüller, G.; Stückler, F.; Rist, M.J.; Skurk, T.; Sailer, M.; Raffler, J.; Römisch-Margl, W.; Adamski, J.; Prehn, C.; et al. The dynamic range of the human metabolome revealed by challenges. FASEB J. 2012, 26, 2607–2619. [Google Scholar] [CrossRef] [PubMed]
- Fiamoncini, J.; Rundle, M.; Gibbons, H.; Thomas, E.L.; Geillinger-Kästle, K.; Bunzel, D.; Trezzi, J.P.; Kiselova-Kaneva, Y.; Wopereis, S.; Wahrheit, J.; et al. Plasma metabolome analysis identifies distinct human metabotypes in the postprandial state with different susceptibility to weight loss-mediated metabolic improvements. FASEB J. 2018, fj201800330R. [Google Scholar] [CrossRef] [PubMed]
- Kayser, B.D.; Lhomme, M.; Dao, M.C.; Ichou, F.; Bouillot, J.L.; Prifti, E.; Kontush, A.; Chevallier, J.M.; Aron-Wisnewsky, J.; Dugail, I.; et al. Serum lipidomics reveals early differential effects of gastric bypass compared with banding on phospholipids and sphingolipids independent of differences in weight loss. Int. J. Obes. 2017, 41, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.; Yu, H.; Zhao, X.; Bao, Y.; Hong, C.S.; Zhang, P.; Tu, Y.; Yin, P.; Gao, P.; Wei, L.; et al. Metabolomics Study of Roux-en-Y Gastric Bypass Surgery (RYGB) to Treat Type 2 Diabetes Patients Based on Ultraperformance Liquid Chromatography-Mass Spectrometry. J. Proteome. Res. 2016, 15, 1288–1299. [Google Scholar] [CrossRef] [PubMed]
- Schooneman, M.G.; Vaz, F.M.; Houten, S.M.; Soeters, M.R. Acylcarnitines: Reflecting or inflicting insulin resistance? Diabetes 2013, 62, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Camps, S.G.; Verhoef, S.P.; Westerterp, K.R. Weight loss, weight maintenance, and adaptive thermogenesis. Am. J. Clin. Nutr. 2013, 97, 990–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenbaum, M.; Hirsch, J.; Gallagher, D.A.; Leibel, R.L. Long-term persistence of adaptive thermogenesis in subjects who have maintained a reduced body weight. Am. J. Clin. Nutr. 2008, 88, 906–912. [Google Scholar] [CrossRef] [PubMed]
- Schooneman, M.G.; Napolitano, A.; Houten, S.M.; Ambler, G.K.; Murgatroyd, P.R.; Miller, S.R.; Hollak, C.E.; Tan, C.Y.; Virtue, S.; Vidal-Puig, A.; et al. Assessment of plasma acylcarnitines before and after weight loss in obese subjects. Arch. Biochem. Biophys. 2016, 606, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Lips, M.A.; Van Klinken, J.B.; van Harmelen, V.; Dharuri, H.K.; ’t Hoen, P.A.; Laros, J.F.; van Ommen, G.J.; Janssen, I.M.; Van Ramshorst, B.; Van Wagensveld, B.A.; et al. Roux-en-Y gastric bypass surgery; but not calorie restriction; reduces plasma branched-chain amino acids in obese women independent of weight loss or the presence of type 2 diabetes. Diabetes Care 2014, 37, 3150–3156. [Google Scholar] [CrossRef] [PubMed]
- Laferrère, B.; Reilly, D.; Arias, S.; Swerdlow, N.; Gorroochurn, P.; Bawa, B.; Bose, M.; Teixeira, J.; Stevens, R.D.; Wenner, B.R.; et al. Differential metabolic impact of gastric bypass surgery versus dietary intervention in obese diabetic subjects despite identical weight loss. Sci. Transl. Med. 2011, 3, 80re2. [Google Scholar]
- Newgard, C.B. Interplay between lipids and branched-chain amino acids in development of insulin resistance. Cell Metab. 2012, 15, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Pellicciari, R.; Pruzanski, M.; Auwerx, J.; Schoonjans, K. Targeting bile-acid signalling for metabolic diseases. Nat. Rev. Drug Discov. 2008, 7, 678–693. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, H.; Kasama, K.; Oshiro, T.; Watanabe, M.; Hirose, H.; Itoh, H. Serum bile acid along with plasma incretins and serum high-molecular weight adiponectin levels are increased after bariatric surgery. Metabolism 2009, 58, 1400–1407. [Google Scholar] [CrossRef] [PubMed]
- Steinert, R.E.; Peterli, R.; Keller, S.; Meyer-Gerspach, A.C.; Drewe, J.; Peters, T.; Beglinger, C. Bile acids and gut peptide secretion after bariatric surgery: A 1-year prospective randomized pilot trial. Obesity 2013, 21, E660–E668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefebvre, P.; Cariou, B.; Lien, F.; Kuipers, F.; Staels, B. Role of bile acids and bile acid receptors in metabolic regulation. Physiol. Rev. 2009, 89, 147–191. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; John, L.M.; Adams, S.H.; Yu, X.X.; Tomlinson, E.; Renz, M.; Williams, P.M.; Soriano, R.; Corpuz, R.; Moffat, B.; et al. Fibroblast growth factor 19 increases metabolic rate and reverses dietary and leptin-deficient diabetes. Endocrinology 2004, 145, 2594–2603. [Google Scholar] [CrossRef] [PubMed]
- De Giorgi, S.; Campos, V.; Egli, L.; Toepel, U.; Carrel, G.; Cariou, B.; Rainteau, D.; Schneiter, P.; Tappy, L.; Giusti, V. Long-term effects of Roux-en-Y gastric bypass on postprandial plasma lipid and bile acids kinetics in female non diabetic subjects: A cross-sectional pilot study. Clin. Nutr. 2015, 34, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.N.; Pfalzer, A.; Kaplan, L.M. Roux-en-Y gastric bypass normalizes the blunted postprandial bile acid excursion associated with obesity. Int. J. Obes. 2013, 37, 1553–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palleja, A.; Kashani, A.; Allin, K.H.; Nielsen, T.; Zhang, C.; Li, Y.; Brach, T.; Liang, S.; Feng, Q.; Jørgensen, N.B.; et al. Roux-en-Y gastric bypass surgery of morbidly obese patients induces swift and persistent changes of the individual gut microbiota. Genome Med. 2016, 8, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremaroli, V.; Karlsson, F.; Werling, M.; Ståhlman, M.; Kovatcheva-Datchary, P.; Olbers, T.; Fändriks, L.; le Roux, C.W.; Nielsen, J.; Bäckhed, F.; et al. Roux-en-Y Gastric Bypass and Vertical Banded Gastroplasty Induce Long-Term Changes on the Human Gut Microbiome Contributing to Fat Mass Regulation. Cell Metab. 2015, 22, 228–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palau-Rodriguez, M.; Tulipani, S.; Marco-Ramell, A.; Miñarro, A.; Jauregui, O.; Gonzalez-Dominguez, R.; Sanchez-Pla, A.; Ramos-Molina, B.; Tinahones, F.J.; Andres-Lacueva, C. Characterization of Metabolomic Profile Associated with Metabolic Improvement after Bariatric Surgery in Subjects with Morbid Obesity. J. Proteome. Res. 2018, 17, 2704–2714. [Google Scholar] [CrossRef] [PubMed]
- Garg, M.L.; Keelan, M.; Thomson, A.B.; Clandinin, M.T. Intestinal microsomes: Polyunsaturated fatty acid metabolism and regulation of enterocyte transport properties. Can. J. Physiol. Pharmacol. 1990, 68, 636–641. [Google Scholar] [CrossRef] [PubMed]
- Forbes, R.; Gasevic, D.; Watson, E.M.; Ziegler, T.R.; Lin, E.; Burgess, J.R.; Gletsu-Miller, N. Essential Fatty Acid Plasma Profiles Following Gastric Bypass and Adjusted Gastric Banding Bariatric Surgeries. Obes. Surg. 2016, 26, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- Christophe, A.; Vermeulen, A. Effects of weight loss on the fatty acid composition of serum lipids in obese women. Ann. Nutr. Metab. 1992, 36, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Kunesová, M.; Phinney, S.; Hainer, V.; Tvrzická, E.; Stich, V.; Parízková, J.; ŽÁk, J.; Stunkard, A. The responses of serum and adipose Fatty acids to a one-year weight reduction regimen in female obese monozygotic twins. Ann. N. Y. Acad. Sci. 2002, 967, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Mutch, D.M.; Fuhrmann, J.C.; Rein, D.; Wiemer, J.C.; Bouillot, J.L.; Poitou, C.; Clément, K. Metabolite profiling identifies candidate markers reflecting the clinical adaptations associated with Roux-en-Y gastric bypass surgery. PLoS ONE 2009, 4, e7905. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, S.; Han, C.Y.; Chiba, T.; McMillen, T.S.; Wang, S.A.; Haw, A.; Kirk, E.A.; O’Brien, K.D.; Chait, A. Dietary cholesterol worsens adipose tissue macrophage accumulation and atherosclerosis in obese LDL receptor-deficient mice. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Umemoto, T.; Subramanian, S.; Ding, Y.; Goodspeed, L.; Wang, S.; Han, C.Y.; Teresa, A.S.; Kim, J.; O’Brien, K.D.; Chait, A. Inhibition of intestinal cholesterol absorption decreases atherosclerosis but not adipose tissue inflammation. J. Lipid. Res. 2012, 53, 2380–2389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyberg, L.; Duan, R.D.; Nilsson, A. A mutual inhibitory effect on absorption of sphingomyelin and cholesterol. J. Nutr. Biochem. 2000, 11, 244–249. [Google Scholar] [CrossRef]
- Noh, S.K.; Koo, S.I. Milk sphingomyelin is more effective than egg sphingomyelin in inhibiting intestinal absorption of cholesterol and fat in rats. J. Nutr. 2004, 134, 2611–2616. [Google Scholar] [CrossRef] [PubMed]





| Medications | Number of Patients |
|---|---|
| Statins | 2 |
| Antihypertensives | 7 |
| Insulin | 1 |
| Metformin | 3 |
| Levothyroxine | 4 |
| Conditions | - |
| Tabagism | 7 |
| Low-grade/moderate steatosis | 17 |
| Pre-Surgery n = 26 | 90 Days after Surgery n = 26 | |
|---|---|---|
| Age | 36.2 ± 7.8 | - |
| Sex (Fem/Male) | 18/8 | - |
| Weight (kg) | 122 ± 22.4 | 96 ± 16.4 *** |
| % Weight loss | 20.9 ± 6.1 *** | |
| BMI (kg/m2) | 44.1 ± 3.6 | 35 ± 3.3 *** |
| % BMI loss | - | 20.7 ± 6.2 *** |
| Waist circ. (cm) | 121.5 ± 17.4 | 104.8 ± 12.6 ** |
| Hip circ. (cm) | 133.1 ± 11.6 | 122.3 ± 15.9 ** |
| Glucose (mg/dL) | 101.2 ± 22.5 | 84.36 ± 8.6 * |
| HbA1c (%) | 6.57 ± 2.2 | 5.5 ± 0.8 * |
| Cholesterol (mg/dL) | 203.68 ± 45.4 | 161.8 ± 40.3 *** |
| LDL-C (mg/dL) | 126 ± 38.9 | 98.7 ± 32.6 *** |
| HDL-C (mg/dL) | 42.7 ± 9.6 | 38.52 ± 10.2 * |
| Triglycerides (mg/dL) | 169.9 ± 97.2 | 121.3 ± 87.8 *** |
| γGT (U/L) | 41.8 ± 22.4 | 25.6 ± 19.4 * |
| ALT (U/L) | 23.2 ± 10.9 | 20.7 ± 6.3 |
| AST (U/L) | 35.2 ± 25.6 | 38.6 ± 42.6 |
| Creatinine (mg/dL) | 0.81 ± 0.15 | 0.71 ± 0,1 * |
| Uric acid (mg/dL) | 6.1 ± 2.1 | 5.1 ± 1.4 ** |
| Vit D (pg/dL) | 23.2 ± 7.8 | 24.6 ± 7.8 |
| Folic acid (ng/dL) | 10.4 ± 3 | 10.2 ± 5.4 |
| Ferritin (ng/dL) | 156 ± 130.5 | 150.2 ± 101.3 |
| PCR (mg/dL) | 5.01 ± 5.9 | 4.65 ± 8.5 |
| T4 (ug/dL) | 8.3 ± 1.8 | 7.5 ± 2.5 |
| TSH (mU/L) | 3.8 ± 4.6 | 2.8 ± 2.5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fiamoncini, J.; Fernandes Barbosa, C.; Arnoni Junior, J.R.; Araújo Junior, J.C.; Taglieri, C.; Szego, T.; Gelhaus, B.; Possolo de Souza, H.; Daniel, H.; Martins de Lima, T. Roux-en-Y Gastric Bypass Surgery Induces Distinct but Frequently Transient Effects on Acylcarnitine, Bile Acid and Phospholipid Levels. Metabolites 2018, 8, 83. https://doi.org/10.3390/metabo8040083
Fiamoncini J, Fernandes Barbosa C, Arnoni Junior JR, Araújo Junior JC, Taglieri C, Szego T, Gelhaus B, Possolo de Souza H, Daniel H, Martins de Lima T. Roux-en-Y Gastric Bypass Surgery Induces Distinct but Frequently Transient Effects on Acylcarnitine, Bile Acid and Phospholipid Levels. Metabolites. 2018; 8(4):83. https://doi.org/10.3390/metabo8040083
Chicago/Turabian StyleFiamoncini, Jarlei, Carina Fernandes Barbosa, José Rubens Arnoni Junior, José Celestino Araújo Junior, Cinthia Taglieri, Tiago Szego, Barbara Gelhaus, Heraldo Possolo de Souza, Hannelore Daniel, and Thais Martins de Lima. 2018. "Roux-en-Y Gastric Bypass Surgery Induces Distinct but Frequently Transient Effects on Acylcarnitine, Bile Acid and Phospholipid Levels" Metabolites 8, no. 4: 83. https://doi.org/10.3390/metabo8040083