
Differential Accumulation of Metabolites and Transcripts Related to Flavonoid, Styrylpyrone, and Galactolipid Biosynthesis in Equisetum Species and Tissue Types



Abstract
:1. Introduction
2. Results and Discussion
2.1. Experimental Design Considerations
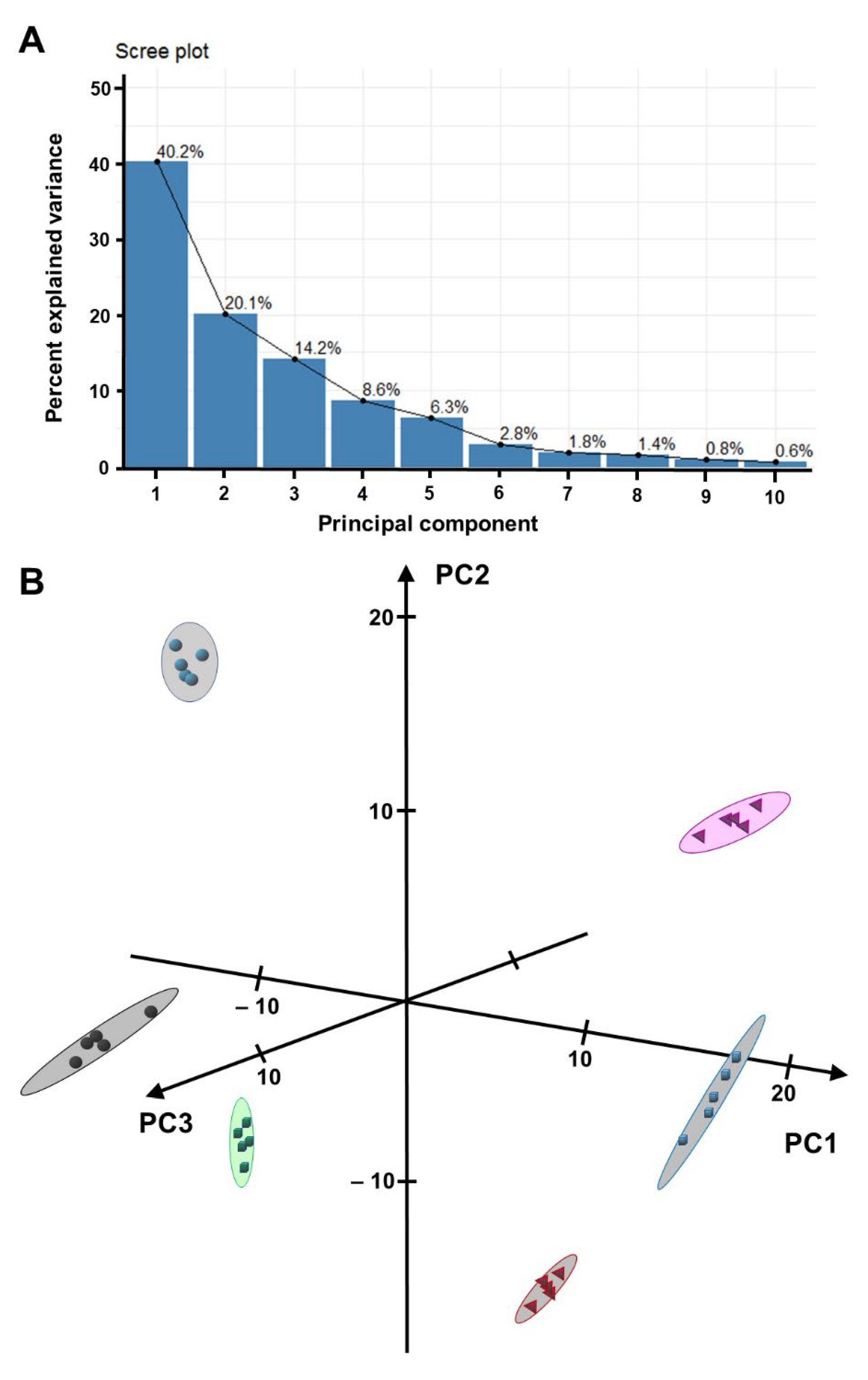
2.2. Sample Differentiation Based on HPLC–QTOF-MS Data
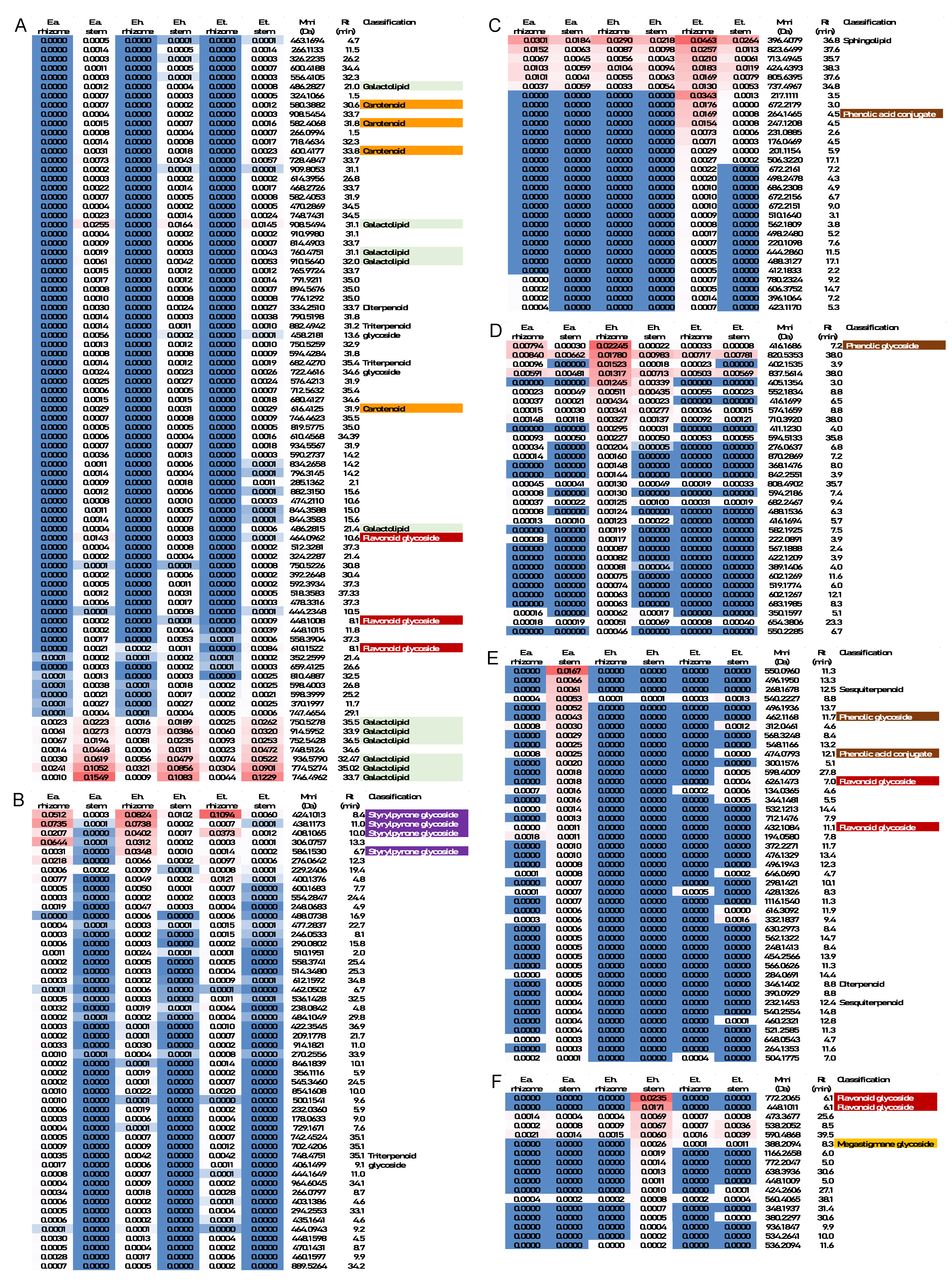
2.3. Annotation of Metabolites That Contribute Substantially to the Separation of Samples by PCA
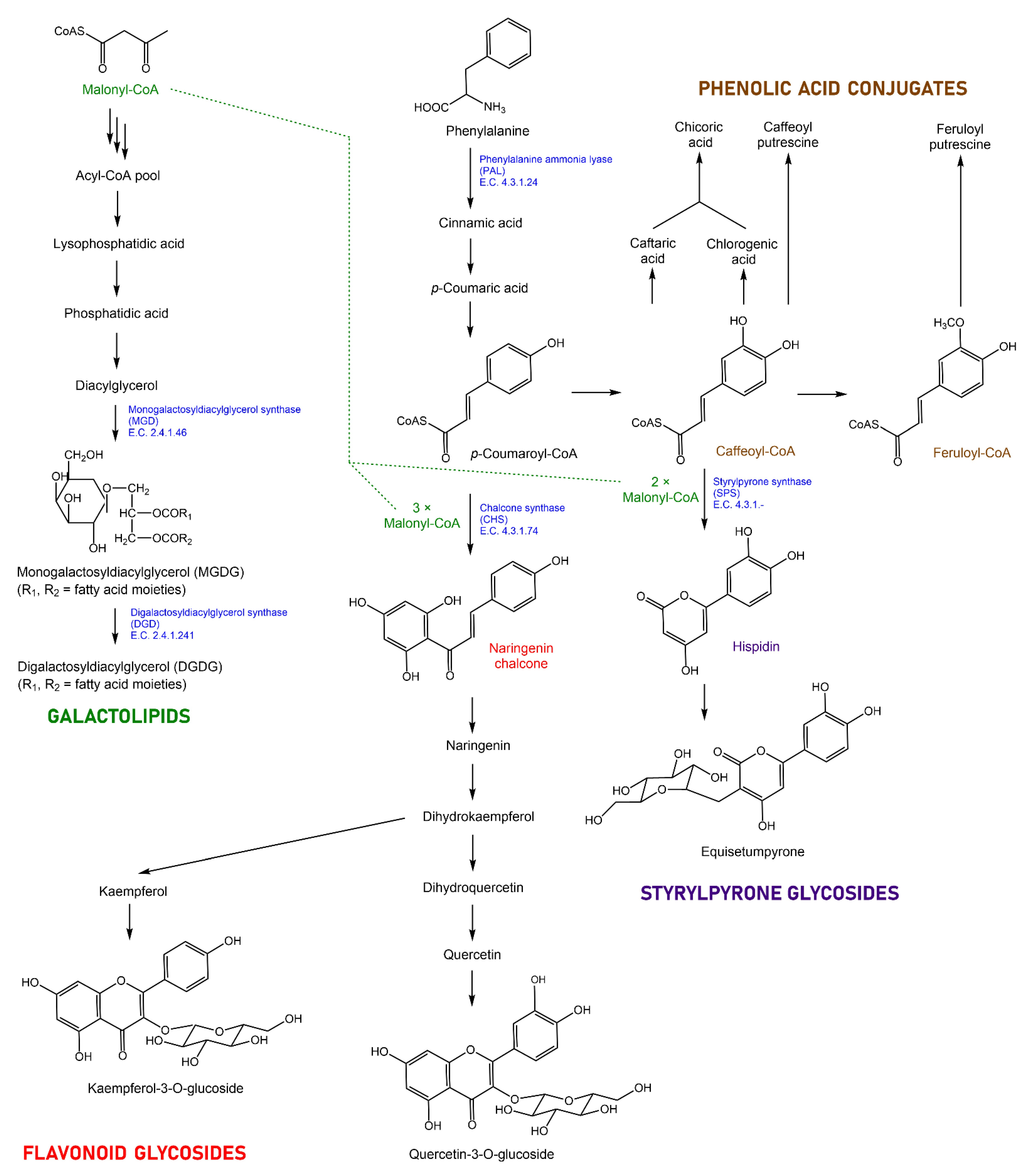
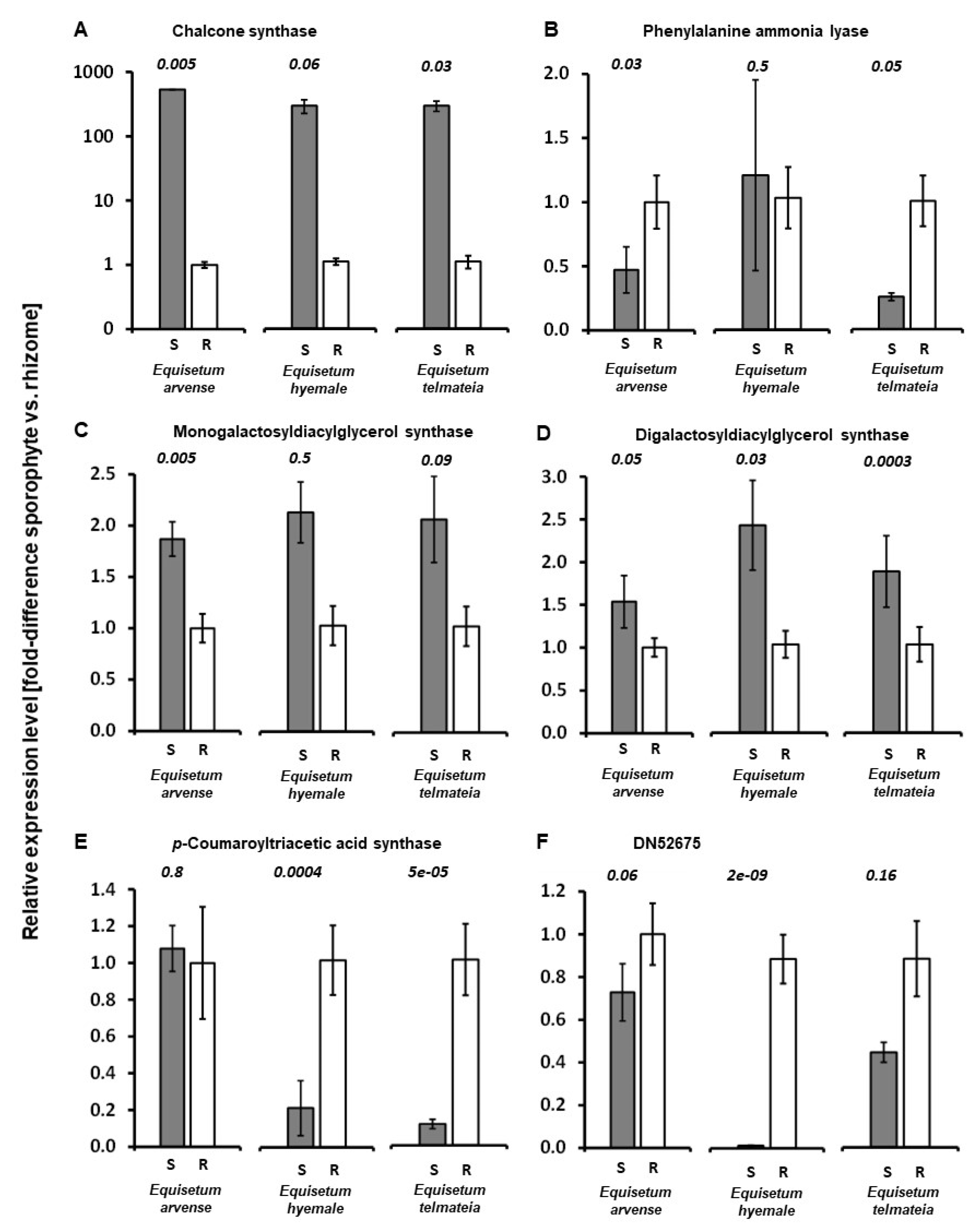
2.4. Expression Patterns of Genes Putatively Involved in the Biosynthesis of Specialized Metabolites in Equisetum
3. Materials and Methods
3.1. Plant Growth
3.2. Tissue Extraction and Analysis by HPLC–QTOF-MS
3.3. HPLC–QTOF-MS Data Processing and Statistical Analyses
3.4. RNA Extraction, RNA-Seq, and Data Processing
3.5. RNA Extraction, First-Strand cDNA Synthesis, qPCR, and Data Processing
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elgorriaga, A.; Escapa, I.H.; Rothwell, G.W.; Tomescu, A.M.; Rubén Cúneo, N. Origin of Equisetum: Evolution of horsetails (Equisetales) within the major euphyllophyte clade Sphenopsida. Am. J. Bot. 2018, 105, 1286–1303. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.W.; Puttick, M.N.; Donoghue, P.C. Origin of horsetails and the role of whole-genome duplication in plant macroevolution. Proc. R. Soc. B 2019, 286, 20191662. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, E.C. The Development, Structure, and Affinities of the Genus Equisetum; Kessinger Publishing: Whitefish, MT, USA, 1899; Volume 5, 48p. [Google Scholar]
- Carneiro, D.M.; Jardim, T.V.; Araújo, Y.C.L.; Arantes, A.C.; de Sousa, A.C.; Barroso, W.K.S.; Sousa, A.L.L.; da Cunha, L.C.; Cirilo, H.N.C.; Bara, M.T.F. Equisetum arvense: New Evidences Supports Medical use in Daily Clinic. Pharmacogn. Rev. 2019, 13, 51. [Google Scholar] [CrossRef]
- Glet, E.; Gutschmidt, J.; Glet, P. Das Alkaloid im Equisetum palustre. Z. Physiol. Chem. 1936, 244, 229–234. [Google Scholar] [CrossRef]
- Karrer, P.; Eugster, C. Über ein Alkaloid aus Equisetum palustre. Helv. Chim. Acta 1948, 31, 1062–1066. [Google Scholar] [CrossRef] [PubMed]
- Eugster, C.H.; Griot, R.; Karrer, P. Weiteres über die Sumpfschachtelhalmbasen. Helv. Chim. Acta 1953, 36, 1387–1400. [Google Scholar] [CrossRef]
- Hauteville, M.; Chopin, J.; Geiger, H.; Schuler, L. Protogenkwanin, a new flavonoid from Equisetum arvense L. Tetrahedron 1981, 37, 377–381. [Google Scholar] [CrossRef]
- Kutney, J.P.; Hall, J.E. Constituents from Equisetum telmateia: The structures of equisporoside and equisporol. Phytochemistry 1971, 10, 3287–3289. [Google Scholar] [CrossRef]
- Saleh, N.; Majak, W.; Towers, G. Flavonoids of Equisetum species. Phytochemistry 1972, 11, 1095–1099. [Google Scholar] [CrossRef]
- Suzuki, K.-I.; Homma, T. Isolation and chemical structure of flavonoids from the horsetails (Equisetum Arvense L.). J. Adv. Sci. 1997, 9, 104–105. [Google Scholar] [CrossRef]
- Syrchina, A.; Gorokhova, V.; Tyukavkina, N.; Babkin, V.; Voronkov, M. Flavonoid glycosides of spore-bearing stems of Equisetum arvense. Chem. Nat. Compd. 1980, 16, 245–248. [Google Scholar] [CrossRef]
- Veit, M.; Bauer, K.; Beckert, C.; Kast, B.; Geiger, H.; Czygan, F.-C. Phenolic characters of British hybrid taxa in Equisetum subgenus Equisetum. Biochem. Syst. Ecol. 1995, 23, 79–87. [Google Scholar] [CrossRef]
- Veit, M.; Bauer, K.; Geiger, H.; Czygan, F.-C. Flavonoids of Equisetum hybrids in the subgenus Equisetum. Planta Med. 1992, 58, 697. [Google Scholar] [CrossRef]
- Veit, M.; Beckert, C.; Höhne, C.; Bauer, K.; Geiger, H. Interspecific and intraspecific variation of phenolics in the genus Equisetum subgenus Equisetum. Phytochemistry 1995, 38, 881–891. [Google Scholar] [CrossRef]
- Veit, M.; Czygan, F.-C.; Geiger, H. Seasonal and intraspecific variations in flavonoids of Equisetum arvense. Planta Med. 1991, 57, A3–A4. [Google Scholar] [CrossRef]
- Veit, M.; Czygan, F.-C.; Geiger, H.; Markham, K. Flavonoids in Equisetum x litorale. Planta Med. 1990, 56, 578–579. [Google Scholar] [CrossRef]
- Veit, M.; Czygan, F.-C.; Witte, L.; Markham, K.; Geiger, H. New Kaempferol glycosides from Equisetum species. Z. Für Nat. B 1993, 48, 1398–1400. [Google Scholar] [CrossRef]
- Veit, M.; Geiger, H.; Czygan, F.-C.; Markham, K.R. Malonylated flavone 5-O-glucosides in the barren sprouts of Equisetum arvense. Phytochemistry 1990, 29, 2555–2560. [Google Scholar] [CrossRef]
- Veit, M.; Strack, D.; Czygan, F.-C.; Wray, V.; Witte, L. Di-E-caffeoyl-meso-tartaric acid in the barren sprouts of Equisetum arvense. Phytochemistry 1991, 30, 527–529. [Google Scholar] [CrossRef]
- Veit, M.; Weidner, C.; Strack, D.; Wray, V.; Witte, L.; Czygan, F.-C. The distribution of caffeic acid conjugates in the Equisetaceae and some ferns. Phytochemistry 1992, 31, 3483–3485. [Google Scholar] [CrossRef]
- Weidner, C.; Veit, M.; Czygan, F.-C. Accumulation dynamics of caffeic acid conjugates in Equisetum arvense. Planta Med. 1991, 57, A37. [Google Scholar] [CrossRef]
- Cyronak, M.J.; Britton, G.; Simpson, K.L. Rhodoxanthin, the red pigment of Equisetum arvense shoots. Phytochemistry 1977, 16, 612–613. [Google Scholar] [CrossRef]
- D’Agostino, M.; Dini, A.; Pizza, C.; Senatore, F.; Aquino, R. Sterols from Equisetum arvense. Boll. Della Soc. Ital. Di Biol. Sper. 1984, 60, 2241–2245. [Google Scholar]
- Takatsuto, S.; Abe, H.; Gamoh, K. Evidence for Brassinosteroids in Strobilus of Equisetum arvense L. Agr. Biol. Chem. Tokyo 1990, 54, 1057–1059. [Google Scholar] [CrossRef]
- Takatsuto, S.; Abe, H. Sterol Composition of the Strobilus of Equisetum arvense L. Biosci. Biotechnol. Biochem. 1992, 56, 834–835. [Google Scholar] [CrossRef] [Green Version]
- Veit, M.; Geiger, H.; Kast, B.; Beckert, C.; Horn, C.; Markham, K.R.; Wong, H.; Czygan, F.-C. Styrylpyrone glucosides from Equisetum. Phytochemistry 1995, 39, 915–917. [Google Scholar] [CrossRef]
- Veit, M.; Geiger, H.; Wray, V.; Abou-Mandour, A.; Rozdzinski, W.; Witte, L.; Strack, D.; Czygan, F.-C. Equisetumpyrone, a styrylpyrone glucoside in gametophytes from Equisetum arvense. Phytochemistry 1993, 32, 1029–1032. [Google Scholar] [CrossRef]
- Cheng, J.T.; He, J.; Li, X.Y.; Wu, X.D.; Shao, L.D.; Dong, L.B.; Deng, X.; Gao, X.; Peng, L.Y.; Cheng, X. Three New Sucrose Fatty Acid Esters from Equisetum hiemale L. Helv. Chim. Acta 2012, 95, 1158–1163. [Google Scholar] [CrossRef]
- Ang, A.M.G.; Peteros, N.P.; Uy, M.M. Antioxidant and Toxicity Assay-guided Isolation of Herniarin from Equisetum debile Roxb.(Equisetaceae). Asian J. Biol. Life Sci. 2019, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Brune, T.; Haas, K. Equisetum species show uniform epicuticular wax structures but diverse composition patterns. AoB Plants 2011, 2011, plr009. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.-F.; Chen, Y.-H.; Sung, P.-J.; Wang, G.-H.; Liou, J.-R.; Wang, S.-Y.; Chang, S.-T.; Zu, Y.G.; Chiang, M.Y.; Fu, Y.-J. Equisetumone, a novel 4-5-olide secocaryophyllane sesquiterpene from Equisetum palustre. RSC Adv. 2014, 4, 45749–45752. [Google Scholar] [CrossRef]
- Chang, J.; Xuan, L.-J.; Xu, Y.-M. Three new phenolic glycosides from the fertile sprouts of Equisetum arvense. J. Integr. Plant Biol. 2001, 43, 193. [Google Scholar]
- Jin, M.; Zhang, C.; Zheng, T.; Yao, D.; Shen, L.; Luo, J.; Jiang, Z.; Ma, J.; Jin, X.-J.; Cui, J. A new phenyl glycoside from the aerial parts of Equisetum hyemale. Nat. Prod. Res. 2014, 28, 1813–1818. [Google Scholar] [CrossRef]
- Kanchanapoom, T.; Otsuka, H.; Ruchirawat, S. Megastigmane glucosides from Equisetum debile and E. diffusum. Chem. Pharm. Bull. 2007, 55, 1277–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, J.-M.; Qiu, Y.-H.; Tan, X.-Q.; Tan, C.-H.; Xiao, K. Chemical constituents of Equisetum debile. J. Asian Nat. Prod. Res. 2011, 13, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Thai, T.H.; Hung, N.Q.; Van Minh, C.; Cuong, N.X.; Yen, P.H.; Huong, L.M.; Van Kiem, P. Chemical constituents of Equisetum debile and their cytotoxic activity. Nat. Prod. Commun. 2008, 3, 1903–1906. [Google Scholar] [CrossRef] [Green Version]
- Wiedenfeld, H.; Cetto, A.A.; Amador, C.P. Flavonol glycosides from Equisetum myriochaetum. Biochem. Syst. Ecol. 2000, 28, 395–397. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.H.; Tan, C.H.; Jiang, S.H.; Zhu, D.Y. Debilosides A–C: Three New Megastigmane Glucosides from Equisetum debile. Helv. Chim. Acta 2006, 89, 1422–1426. [Google Scholar] [CrossRef]
- Sandhu, N.S.; Kaur, S.; Chopra, D. Equisetum arvense: Pharmacology and phytochemistry-a review. Asian J. Pharm. Clin. Res. 2010, 3, 146–150. [Google Scholar]
- Hohlfeld, M.; Veit, M.; Strack, D. Hydroxycinnamoyltransferases involved in the accumulation of caffeic acid esters in gametophytes and shoots of Equisetum arvense. Plant Physiol. 1996, 111, 1153–1159. [Google Scholar] [CrossRef] [Green Version]
- Beckert, C.; Horn, C.; Schnitzler, J.-P.; Lehning, A.; Heller, W.; Veit, M. Styrylpyrone biosynthesis in Equisetum arvense. Phytochemistry 1997, 44, 275–283. [Google Scholar] [CrossRef]
- Herderich, M.; Beckert, C.; Veit, M. Establishing styrylpyrone synthase activity in cell free extracts obtained from gametophytes of Equisetum arvense L. by high performance liquid chromatography–tandem mass spectrometry. Phytochem. Anal. Int. J. Plant Chem. Biochem. Tech. 1997, 8, 194–197. [Google Scholar] [CrossRef]
- Liou, G.; Chiang, Y.-C.; Wang, Y.; Weng, J.-K. Mechanistic basis for the evolution of chalcone synthase catalytic cysteine reactivity in land plants. J. Biol. Chem. 2018, 293, 18601–18612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuthbertson, D.; Piljac-Žegarac, J.; Lange, B.M. Validation of a microscale extraction and high-throughput UHPLC-QTOF-MS analysis method for huperzine A in Huperzia. Biomed. Chromatogr. 2012, 26, 1191–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuthbertson, D.J.; Johnson, S.R.; Piljac-Zegarac, J.; Kappel, J.; Schafer, S.; Wust, M.; Ketchum, R.E.; Croteau, R.B.; Marques, J.V.; Davin, L.B.; et al. Accurate mass-time tag library for LC/MS-based metabolite profiling of medicinal plants. Phytochemistry 2013, 91, 187–197. [Google Scholar] [CrossRef] [Green Version]
- Lange, B.M. The evolution of plant secretory structures and emergence of terpenoid chemical diversity. Annu. Rev. Plant Biol. 2015, 66, 139–159. [Google Scholar] [CrossRef]
- Šamec, D.; Pierz, V.; Srividya, N.; Wüst, M.; Lange, B.M. Assessing chemical diversity in Psilotum nudum (L.) Beauv., a Pantropical whisk fern that has lost many of its fern-like characters. Front. Plant Sci. 2019, 10, 868. [Google Scholar] [CrossRef]
- Nokhsorov, V.V.; Dudareva, L.V.; Senik, S.V.; Chirikova, N.K.; Petrov, K.A. Influence of Extremely Low Temperatures of the Pole of Cold on the Lipid and Fatty-Acid Composition of Aerial Parts of the Horsetail Family (Equisetaceae). Plants 2021, 10, 996. [Google Scholar] [CrossRef]
- Czeczuga, B. Carotenoid contents in leaves grown under various light intensities. Biochem. Syst. Ecol. 1987, 15, 523–527. [Google Scholar] [CrossRef]
- Veit, M.; Abou-Mandour, A.; Czygan, F.-C. Phenolics from gametophytes of Equisetum arvense. Planta Med. 1991, 57, A36. [Google Scholar] [CrossRef]
- Francescato, L.N.; Debenedetti, S.L.; Schwanz, T.G.; Bassani, V.L.; Henriques, A.T. Identification of phenolic compounds in Equisetum giganteum by LC–ESI-MS/MS and a new approach to total flavonoid quantification. Talanta 2013, 105, 192–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid metabolism in plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef] [Green Version]
- Barros, J.; Dixon, R.A. Plant phenylalanine/tyrosine ammonia-lyases. Trends Plant Sci. 2020, 25, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Pluskal, T.; Torrens-Spence, M.P.; Fallon, T.R.; De Abreu, A.; Shi, C.H.; Weng, J.-K. The biosynthetic origin of psychoactive kavalactones in kava. Nat. Plants 2019, 5, 867–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colpitts, C.C. An Investigation into Plant Type III Polyketide Synthases: A Styrylpyrone Synthase from Equisetum Hyemale and Anther-Specific Chalcone Synthase-Like Enzymes from Physcomitrella Patens and Arabidopsis Thaliana; Faculty of Graduate Studies and Research, University of Regina: Regina, SK, Canada, 2009. [Google Scholar]
- RStudio Home Page. Available online: https://www.rstudio.com (accessed on 1 April 2022).
- The Comprehensive R Archive Network Home Page. Available online: https://cran.r-project.org/ (accessed on 1 April 2022).
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B. PubChem in 2021: New data content and improved web interfaces. Nucleic Acids Res. 2021, 49, D1388–D1395. [Google Scholar] [CrossRef]
- Afendi, F.M.; Okada, T.; Yamazaki, M.; Hirai-Morita, A.; Nakamura, Y.; Nakamura, K.; Ikeda, S.; Takahashi, H.; Altaf-Ul-Amin, M.; Darusman, L.K. KNApSAcK family databases: Integrated metabolite–plant species databases for multifaceted plant research. Plant Cell Physiol. 2012, 53, e1. [Google Scholar] [CrossRef] [Green Version]
- Fischedick, J.T.; Johnson, S.R.; Ketchum, R.E.B.; Croteau, R.B.; Lange, B.M. NMR spectroscopic search module for Spektraris, an online resource for plant natural product identification—Taxane diterpenoids from Taxus x media cell suspension cultures as a case study. Phytochemistry 2015, 113, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Sud, M.; Fahy, E.; Cotter, D.; Azam, K.; Vadivelu, I.; Burant, C.; Edison, A.; Fiehn, O.; Higashi, R.; Nair, K.S. Metabolomics Workbench: An international repository for metabolomics data and metadata, metabolite standards, protocols, tutorials and training, and analysis tools. Nucleic Acids Res. 2016, 44, D463–D470. Available online: https://www.metabolomicsworkbench.org/ (accessed on 1 April 2022). [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Lefever, S.; Hellemans, J.; Pattyn, F.; Przybylski, D.R.; Taylor, C.; Geurts, R.; Untergasser, A.; Vandesompele, J.; consortium, R. RDML: Structured language and reporting guidelines for real-time quantitative PCR data. Nucleic Acids Res. 2009, 37, 2065–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Untergasser, A.; Ruijter, J.M.; Benes, V.; van den Hoff, M.J. Web-based LinRegPCR: Application for the visualization and analysis of (RT)-qPCR amplification and melting data. BMC Bioinform. 2021, 22, 398. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accurate Mass—Time Tag | Monoisotopic Mass (Measured/Calculated) | Δppm | Molecular Formula | MS (ESI-Positive) (* Most Abundant) | Tentative Annotation | References; Evidence Level, Standard Source |
|---|---|---|---|---|---|---|
| Phenolic acids, phenolic acid conjugates, and phenolic glycosides | ||||||
| BML-LCMS-19- 1.71–148.0521 | 148.0521/148.0524 | 2.29 | C9H8O2 | [M + H]+ 149.0594 [M + NH4]+ 166.0867 * | Cinnamic acid | [13,15]; 2 |
| BML-LCMS-19- 3.40–312.0438 | 312.0438/312.0481 | 2.98 | C13H12O9 | [M + Na]+ 335.0371 * | Caffeoyl tartaric acid (caftaric acid) | [13,15]; 4, Sigma Aldrich 88,656 |
| BML-LCMS-19- 4.53–264.1465 | 264.1465/264.1474 | 1.34 | C14H20N2O3 | [M + H]+ 265.1552 * [M + Na]+ 287.1366 | Feruloylputrescine | No reference; 1 |
| BML-LCMS-19- 7.24–416.1686 | 416.1686/416.1682 | 0.62 | C19H28O10 | [M + Na]+ 439.1579 * | Equisetumoside A or B | [33]; (2) |
| BML-LCMS-19- 7.79–414.1533 | 414.1533/414.1526 | 1.38 | C19H26O10 | [M + Na]+ 437.1426 * | Equisetumoside D | [37]; (2) |
| BML-LCMS-19- 10.64–310.1246 | 310.1246/310.1205 | 0.55 | C19H18O4 | [M + H]+ 311.1281 * | Coniferylcinnamate | No reference; 1 |
| BML-LCMS-19- 12.15–474.0793 | 474.0793/474.0798 | 0.60 | C22H18O12 | [M + Na]+ 497.0690 * | Dicaffeoyltartaric acid (chicoric acid) | [13,15,20,51]; (4) Cayman 24,960 |
| Flavonoid glycosides | ||||||
| BML-LCMS-19- 6.08–772.2065 | 772.2065/772.2062 | 0.93 | C33H40O21 | [M + H]+ 773.2147 * | Kaempferol 3-O-sophoroside-7-O-glucoside | [13,15]; 3 |
| BML-LCMS-19- 7.05–626.1473 | 626.1473/626.1483 | 1.93 | C27H30O17 | [M + H]+ 627.1545 * | Quercetin-3,7-di-O-glucoside | [13,15]; 3 |
| BML-LCMS-19- 8.06–448.1008 | 448.1008/448.1006 | 0.37 | C21H20O11 | [M + H]+ 449.1079 * | Luteolin-5-O-glucoside | [13,15,51,52]; 4, Sigma Aldrich 1,370,837 |
| BML-LCMS-19- 8.06–610.1522 | 610.1522/610.1534 | 0.37 | C27H30O16 | [M + H]+ 611.1614 * [M + Na]+ 633.1432 | Kaempferol-3,7-O-di-glucoside | [13,15]; 3 |
| BML-LCMS-18- 8.20–756.2103 | 756.2103/756.2113 | 0.75 | C33H40O20 | [M + H]+ 757.2181 * | Kaempferol-3-O-rutinoside-7-O-glucoside | [13,15]; 3 |
| BML-LCMS-19- 9.95–610.1540 | 610.1540/610.1534 | 1.23 | C27H30O16 | [M + H]+ 611.1622 [M + Na]+ 633.1438 * | Kaempferol-3-O-sophoroside | [13,15]; 3 |
| BML-LCMS-19- 10.60–464.0962 | 464.0962/464.0955 | 1.02 | C21H20O12 | [M + H]+ 465.1036 * [M + Na]+ 487.0849 | Quercetin 3-glucoside (isoquercitrin) | [13,15,51,52]; 4, Sigma Aldrich 00140585 |
| BML-LCMS-19- 11.78–448.1015 | 448.1015/448.1006 | 0.01 | C21H20O11 | [M + Na]+ 471.0900 * | Kaempferol-3-O-glucoside (astragalin) | [13,15,51,52]; 4, Cayman 25,060 |
| Styrylpyrone glycosides | ||||||
| BML-LCMS-19- 6.68–586.1530 | 586.1530/586.1534 | 0.51 | C25H30O16 | [M + H]+ 587.2013 * | 3-Hydroxyhispidin-3,4’-di-O-glucoside | [52]; 3 |
| BML-LCMS-19- 8.39–424.1013 | 424.1013/424.1006 | 1.37 | C19H20O11 | [M + H]+ 425.1085 * [M + Na]+ 447.0903 | Equisetumpyrone | [28,52]; 3 |
| BML-LCMS-19- 9.97–408.1065 | 408.1065/408.1056 | 1.31 | C19H20O10 | [M + H]+ 409.1136 * [M + Na]+ 431.0957 | 3’-Deoxyequisetumpyrone | [27]; 3 |
| BML-LCMS-19- 10.97–438.1173 | 438.1173/438.1162 | 1.63 | C20H22O11 | [M + H]+ 439.1245 * [M + Na]+ 461.1064 | 4’-O-Methylequisetumpyrone | [27]; 3 |
| Carotenoids and apocarotenoids | ||||||
| BML-LC-MS-18- 8.28–388.2094 | 388.2094/388.2097 | 0.72 | C19H32O8 | [M + Na]+ 411.1991 * | Debiloside B | [39]; 2 |
| BML-LCMS-19- 30.60–580.3882 | 580.3882/580.3916 | 1.88 | C40H52O3 | [M + H]+ 581.3978 * | Carotenoid | No reference; 1 |
| BML-LMS-19- 31.75–582.4068 | 582.4068/582.4073 | 0.03 | C40H54O3 | [M + H]+ 583.4142 * | Carotenoid | No reference; 1 |
| BML-LCMS-19- 31.91–616.4125 | 616.4125/616.4128 | 0.05 | C40H56O5 | [M + Na]+ 639.4013 * | Carotenoid | No reference; 1 |
| BML-LCMS-19- 33.83–600.4177 | 600.4177/600.4179 | 0.63 | C40H56O4 | [M + H]+ 601.4246 * | Violaxanthin | [53]; 4, Sigma Aldrich 52,444 |
| Lipids | ||||||
| BML-LCMS-19- 21.01–486.2827 | 486.2827/486.2829 | 2.55 | C25H42O9 | [M + NH4]+ 504.3157 [M + Na]+ 509.2718 * | 16:3-Glycosylmonoacylglycerol | No reference; 1 |
| BML-LCMS-19- 21.36–486.2815 | 486.2815/486.2829 | 2.83 | C25H42O9 | [M + NH4]+ 504.3157 [M + Na]+ 509.2714 * [M + K]+ 525.2461 | 16:3-Glycosylmonoacylglycerol | No reference; 1 |
| BML-LCMS-19- 31.15–908.5494 | 908.5494/908.5497 | 0.79 | C49H80O15 | [M + NH4]+ 926.6327 [M + Na]+ 931.5382 * | Digalactosyldiacylglycerol (34:6) | [49]; 4, Avanti 840,524 (mix) |
| BML-LCMS-19- 31.97–910.5640 | 910.5640/910.5654 | 1.55 | C49H82O15 | [M + Na]+ 933.5536 * [M + K]+ 949.5267 | Digalactosyldiacylglycerol (34:5) | [49]; 4, Avanti 840,524 (mix) |
| BML-LCMS-19- 32.47–936.5790 | 936.5790/936.5810 | 1.36 | C51H84O15 | [M + NH4]+ 954.6140 [M + Na]+ 959.5708 * | Digalactosyldiacylglycerol (36:6) | [49]; 4, Avanti 840,524 (mix) |
| BML-LCMS-19- 33.31–938.5952 | 938.5952/938.5967 | 0.15 | C51H86O15 | [M + Na]+ 961.5872 * | Digalactosyldiacylglycerol (36:5) | No reference; 1 |
| BML-LCMS-19- 33.70–746.4962 | 746.4962/746.4969 | 0.03 | C43H70O10 | [M + NH4]+ 764.5311 * [M + Na]+ 769.4859 | Monogalactosyldiacylglycerol (34:6) | [49]; 4, Avanti 840,523 (mix) |
| BML-LCMS-19- 33.94–914.5952 | 914.5952/914.5967 | 0.71 | C49H86O15 | [M + NH4]+ 932.6303 [M + Na]+ 937.5848 * | Digalactosyldiacylglycerol (34:3) | [49]; 4, Avanti 840,524 (mix) |
| BML-LCMS-19- 34.19–940.6076 | 940.6076/940.6123 | 0.19 | C51H88O15 | [M + Na]+ 963.6026 * | Digalactosyldiacylglycerol (36:4) | [49]; 4, Avanti 840,524 (mix) |
| BML-LCMS-19- 34.77–916.6117 | 916.6117/916.6123 | 0.94 | C49H88O15 | [M + NH4]+ 934.6454 [M + Na]+ 939.6016 * | Digalactosyldiacylglycerol (34:2) | [49]; 4, Avanti 840,524 (mix) |
| BML-LC-MS-18- 35.02–774.5274 | 774.5274/774.5282 | 0.34 | C45H74O10 | [M + NH4]+ 792.5619 * [M + Na]+ 797.5171 | Monogalactosyldiacylglycerol (36:6) | [49]; 4, Avanti 840,523 (mix) |
| BML-LC-MS-18- 35.46–750.5278 | 750.5278/750.5282 | 0.66 | C43H74O10 | [M + NH4]+ 768.5588 [M + Na]+ 773.5169 * | Monogalactosyldiacylglycerol (34:4) | [49]; 4, Avanti 840,523 (mix) |
| BML-LC-MS-18- 35.86–776.5443 | 776.5443/776.5438 | 0.61 | C45H76O10 | [M + Na]+ 799.5325 * | Monogalactosyldiacylglycerol (36:5) | [49]; 4, Avanti 840,523 (mix) |
| BML-LC-MS-18- 36.54–752.5428 | 752.5428/752.5438 | 1.04 | C43H76O10 | [M + NH4]+ 770.5775 [M + Na]+ 775.5325 * | Monogalactosyldiacylglycerol (34:3) | [49]; 4, Avanti 840,523 (mix) |
| BML-LC-MS-18- 36.72–778.5575 | 778.5575/778.5595 | 1.13 | C45H78O10 | [M + H]+ 779.5910 [M + NH4]+ 796.5930 [M + Na]+ 801.5480 * | Monogalactosyldiacylglycerol (36:4) | [49]; 4, Avanti 840,523 (mix) |
| BML-LC-MS-18- 37.32–754.5557 | 754.5557/754.5595 | 1.70 | C43H78O10 | [M + NH4]+ 772.5921 [M + Na]+ 777.5483 * | Monogalactosyldiacylglycerol (36:2) | No reference; 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parrish, A.N.; Lange, I.; Šamec, D.; Lange, B.M. Differential Accumulation of Metabolites and Transcripts Related to Flavonoid, Styrylpyrone, and Galactolipid Biosynthesis in Equisetum Species and Tissue Types. Metabolites 2022, 12, 403. https://doi.org/10.3390/metabo12050403
Parrish AN, Lange I, Šamec D, Lange BM. Differential Accumulation of Metabolites and Transcripts Related to Flavonoid, Styrylpyrone, and Galactolipid Biosynthesis in Equisetum Species and Tissue Types. Metabolites. 2022; 12(5):403. https://doi.org/10.3390/metabo12050403
Chicago/Turabian StyleParrish, Amber N., Iris Lange, Dunja Šamec, and Bernd Markus Lange. 2022. "Differential Accumulation of Metabolites and Transcripts Related to Flavonoid, Styrylpyrone, and Galactolipid Biosynthesis in Equisetum Species and Tissue Types" Metabolites 12, no. 5: 403. https://doi.org/10.3390/metabo12050403