
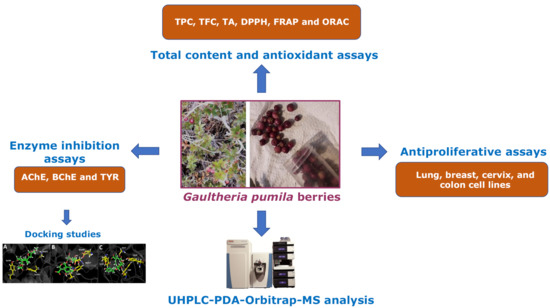
UHPLC-MS Chemical Fingerprinting and Antioxidant, Antiproliferative, and Enzyme Inhibition Potential of Gaultheria pumila Berries


 ,
,  ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material
2.3. Extraction Procedures
2.4. UHPLC–DAD–MS Instrument
2.5. LC Parameters and MS Parameters
2.6. Total Phenolic (TP), Total Flavonoid (TF), and Total Anthocyanins (TA) Content
2.7. Antioxidant Activity
2.7.1. DPPH Scavenging Activity
2.7.2. Ferric-Reducing Antioxidant Power Assay (FRAP)
2.7.3. Oxygen Radical Absorbance Capacity (ORAC) Assay
2.8. Determination of Cholinesterase Inhibition
2.9. Tyrosinase Inhibition Assay
2.10. Docking Studies
2.11. Antiproliferative Activity
2.12. Statistical Analysis
3. Results and Discussion
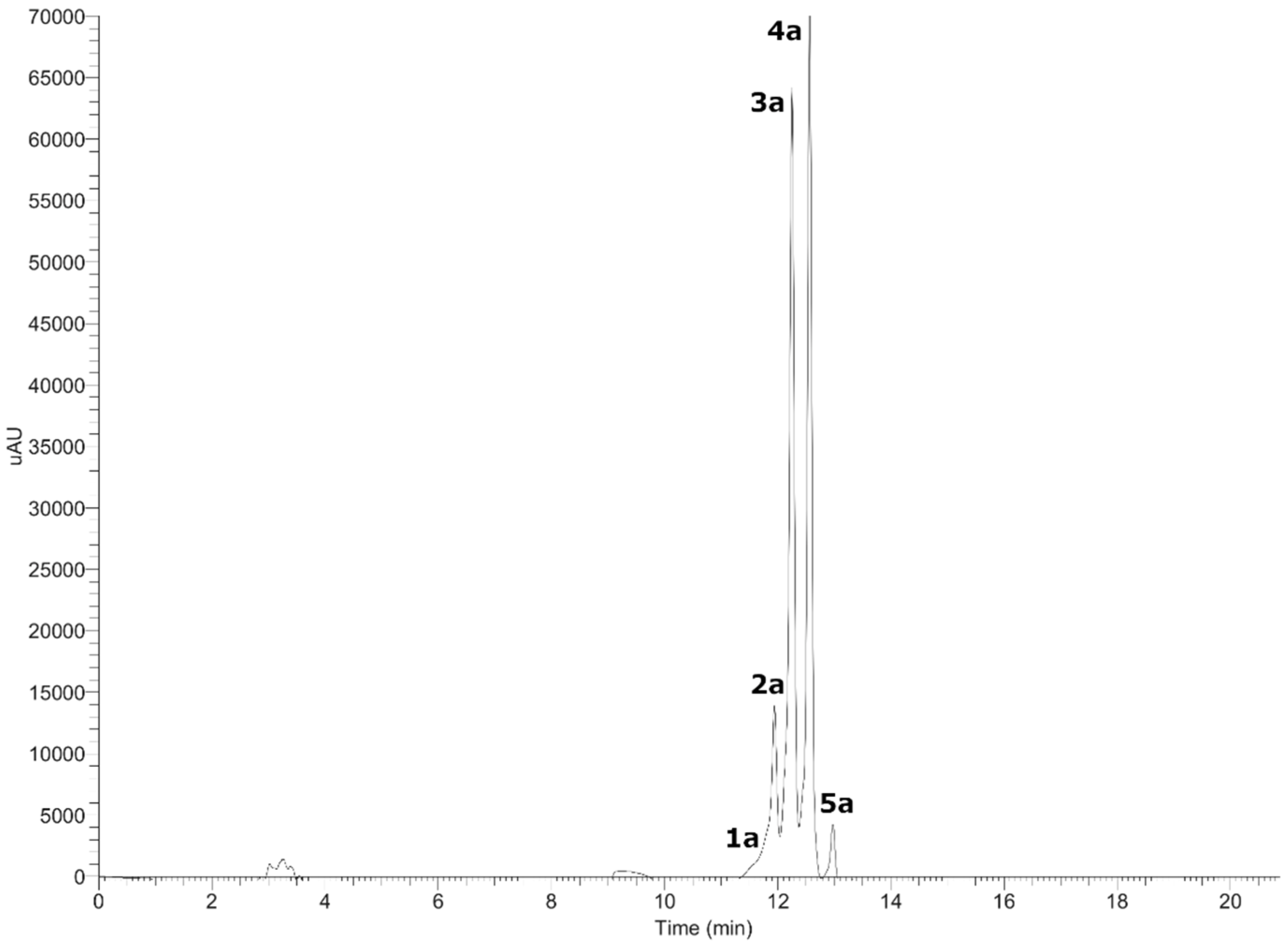
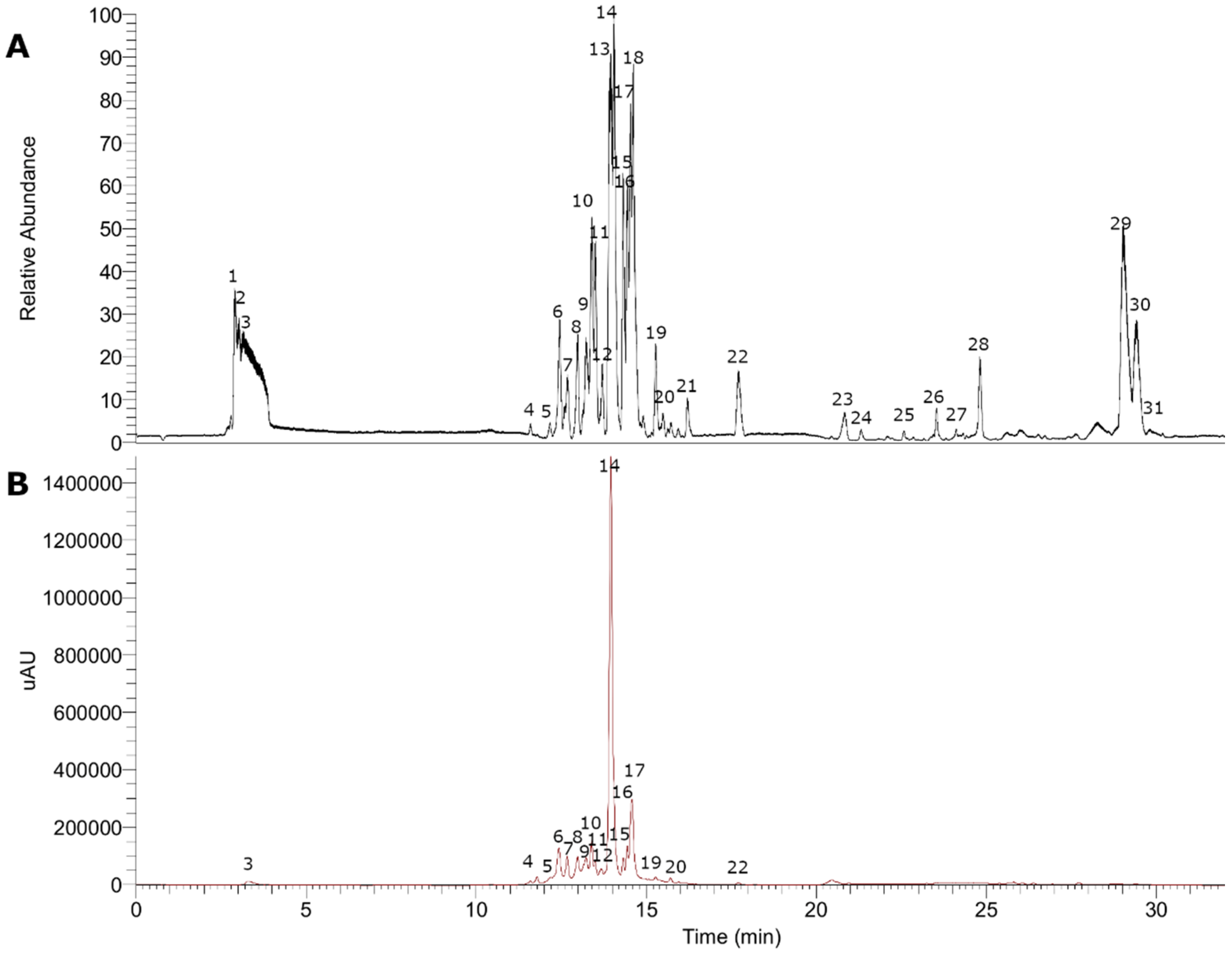
3.1. UHPLC–MS Analysis of Gaultheria Pumila Extract
3.1.1. Anthocyanins
3.1.2. Flavonoids and Derivatives
3.1.3. Phenolic Acids
3.1.4. Fatty Acids
3.1.5. Terpenoids
3.2. Total Phenolic, Flavonoid, and Anthocyanin Content, and Antioxidant Activity
3.3. Enzymatic Inhibitory Activity
3.4. Docking Studies
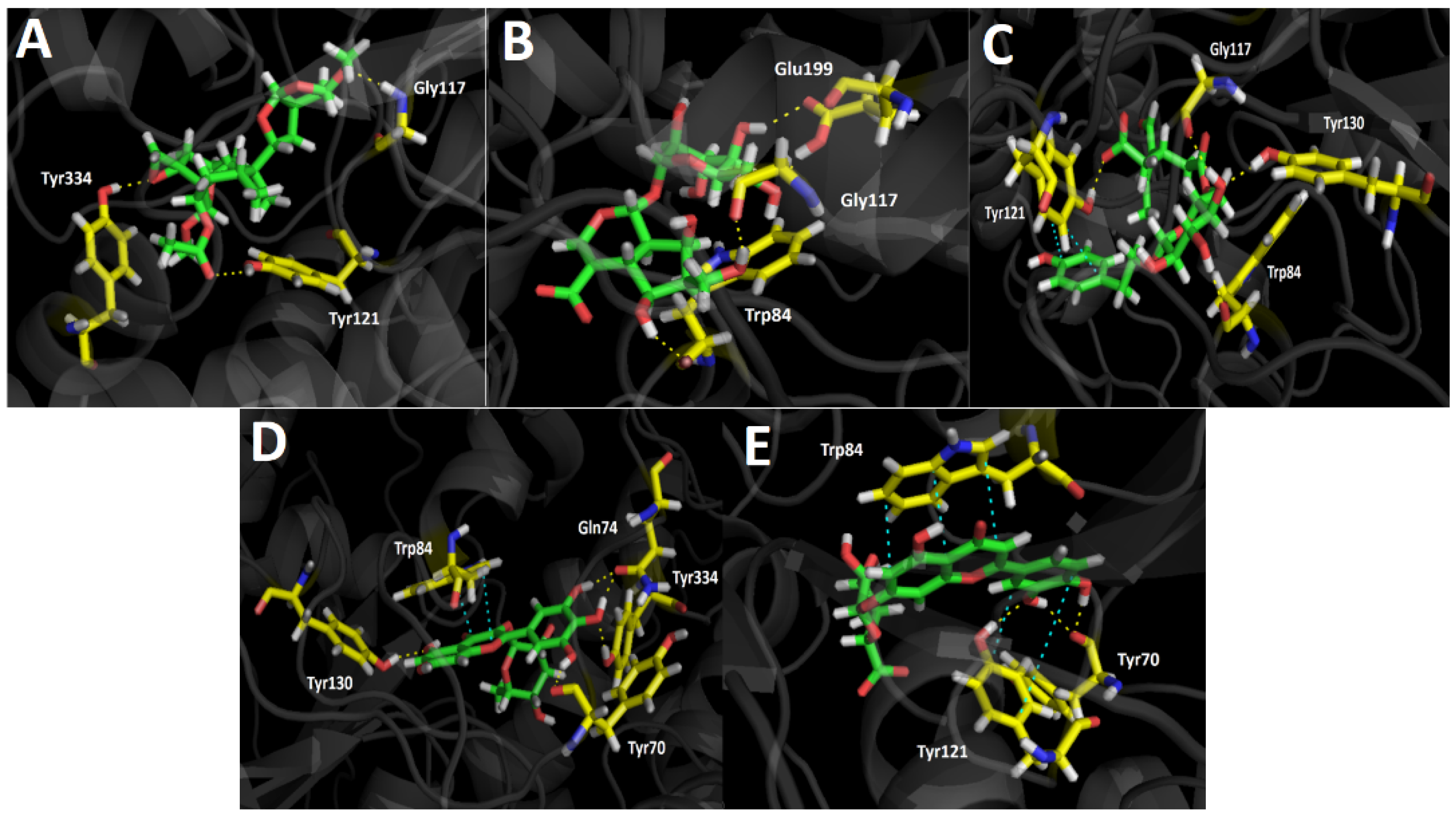
3.4.1. Acetylcholinesterase (TcAChE) Docking Results
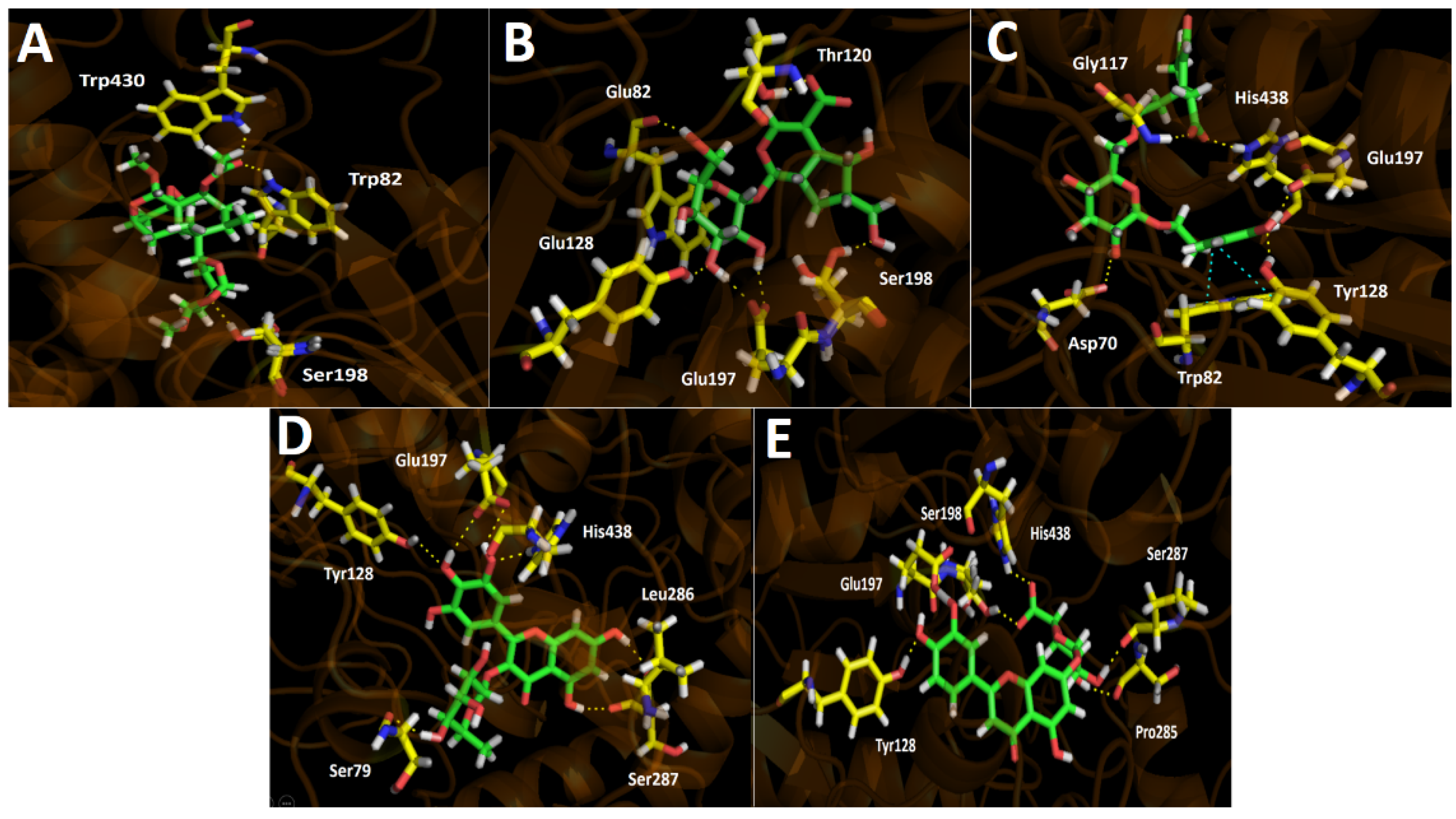
3.4.2. Butyrylcholinesterase (hBuChE) Docking Results
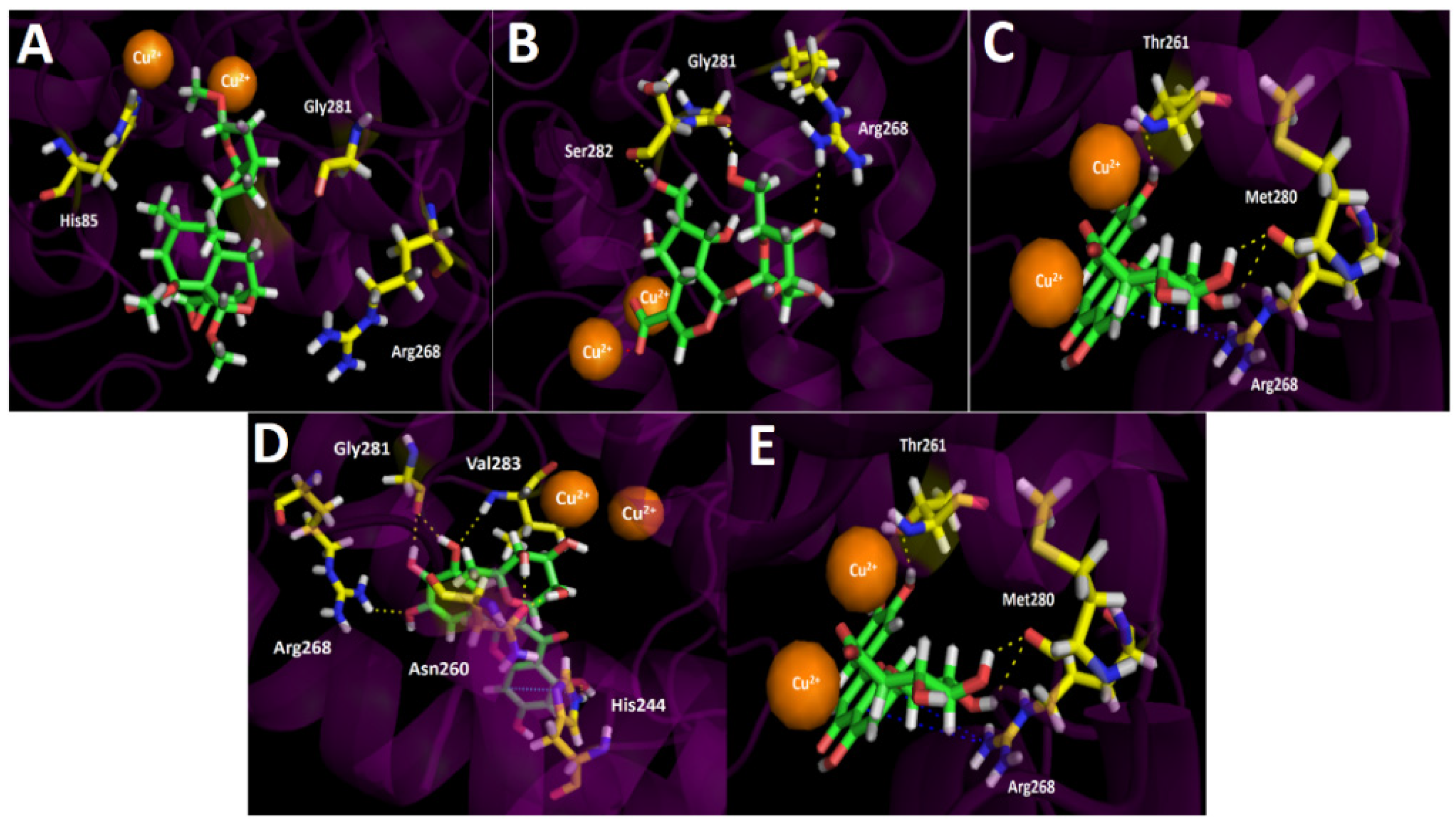
3.4.3. Tyrosinase Docking Results
3.5. Antiproliferative Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Silva, S.; Costa, E.M.; Veiga, M.; Morais, R.M.; Calhau, C.; Pintado, M. Health promoting properties of blueberries: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 181–200. [Google Scholar] [CrossRef]
- Speer, H.; D’Cunha, N.M.; Alexopoulos, N.I.; McKune, A.J.; Naumovski, N. Anthocyanins and Human Health—A Focus on Oxidative Stress, Inflammation and Disease. Antioxidants 2020, 9, 366. [Google Scholar] [CrossRef] [PubMed]
- López, J.; Vera, C.; Bustos, R.; Florez-Mendez, J. Native berries of Chile: A comprehensive review on nutritional aspects, functional properties, and potential health benefits. J. Food Meas. Charact. 2021, 15, 1139–1160. [Google Scholar] [CrossRef]
- Luo, B.; Gu, R.; Kennelly, E.J.; Long, C. Gaultheria Ethnobotany and Bioactivity: Blueberry Relatives with Anti-inflammatory, Antioxidant, and Anticancer Constituents. Curr. Med. Chem. 2017, 25, 5168–5176. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-R.; Qiao, W.-L.; Liu, Z.-Z.; Wang, X.-H.; Jiang, R.; Li, S.-Y.; Shi, R.-B.; She, G.-M. Gaultheria: Phytochemical and Pharmacological Characteristics. Molecules 2013, 18, 12071–12108. [Google Scholar] [CrossRef]
- Pandey, B.P.; Thapa, R.; Upreti, A. Chemical composition, antioxidant and antibacterial activities of essential oil and methanol extract of Artemisia vulgaris and Gaultheria fragrantissima collected from Nepal. Asian Pac. J. Trop. Med. 2017, 10, 952–959. [Google Scholar] [CrossRef]
- Michel, P.; Dobrowolska, A.; Kicel, A.; Owczarek, A.; Bazylko, A.; Granica, S.; Piwowarski, J.P.; Olszewska, M.A. Polyphenolic profile, antioxidant and anti-inflammatory activity of eastern teaberry (Gaultheria procumbens L.) leaf extracts. Molecules 2014, 19, 20498–20520. [Google Scholar] [CrossRef] [Green Version]
- Shen, B.-B.; Yang, Y.-P.; Yasamin, S.; Liang, N.; Su, W.; Chen, S.-H.; Wang, X.-J.; Wang, W. Analysis of the Phytochemistry and Bioactivity of the Genus Polygonum of Polygonaceae. Digit. Chin. Med. 2018, 1, 19–36. [Google Scholar] [CrossRef]
- Ferguson, A.; Carvalho, E.; Gourlay, G.; Walker, V.; Martens, S.; Salminen, J.P.; Constabel, C.P. Phytochemical analysis of salal berry (Gaultheria shallon Pursh.), a traditionally-consumed fruit from western North America with exceptionally high proanthocyanidin content. Phytochemistry 2018, 147, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Teillier, S.; Escobar, F. Revision of the genus of gaultheria L. (Ericaceae) in Chile. Gayana Bot. 2013, 70, 136–153. [Google Scholar] [CrossRef]
- Mieres-Castro, D.; Schmeda-Hirschmann, G.; Theoduloz, C.; Gómez-Alonso, S.; Pérez-Navarro, J.; Márquez, K.; Jiménez-Aspee, F. Antioxidant activity and the isolation of polyphenols and new iridoids from Chilean Gaultheria phillyreifolia and G. poeppigii berries. Food Chem. 2019, 291, 167–179. [Google Scholar] [CrossRef]
- Oyarzún, P.; Cornejo, P.; Gómez-Alonso, S.; Ruiz, A. Influence of Profiles and Concentrations of Phenolic Compounds in the Coloration and Antioxidant Properties of Gaultheria poeppigii Fruits from Southern Chile. Plant Foods Hum. Nutr. 2020, 75, 532–539. [Google Scholar] [CrossRef]
- Mieres-Castro, D.; Schmeda-Hirschmann, G.; Theoduloz, C.; Rojas, A.; Piderit, D.; Jiménez-Aspee, F. Isolation and characterization of secondary metabolites from Gaultheria tenuifolia berries. J. Food Sci. 2020, 85, 2792–2802. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, A.; Hermosín-Gutiérrez, I.; Vergara, C.; von Baer, D.; Zapata, M.; Hitschfeld, A.; Obando, L.; Mardones, C. Anthocyanin profiles in south Patagonian wild berries by HPLC-DAD-ESI-MS/MS. Food Res. Int. 2013, 51, 706–713. [Google Scholar] [CrossRef]
- Ruiz, A.; Bustamante, L.; Vergara, C.; Von Baer, D.; Hermosín-Gutiérrez, I.; Obando, L.; Mardones, C. Hydroxycinnamic acids and flavonols in native edible berries of South Patagonia. Food Chem. 2015, 167, 84–90. [Google Scholar] [CrossRef]
- Villagra, E.; Campos-Hernandez, C.; Cáceres, P.; Cabrera, G.; Bernardo, Y.; Arencibia, A.; Carrasco, B.; Caligari, P.D.S.; Pico, J.; García-Gonzales, R. Morphometric and phytochemical characterization of chaura fruits (Gaultheria pumila): A native Chilean berry with commercial potential. Biol. Res. 2014, 47, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocchetti, G.; Chiodelli, G.; Giuberti, G.; Ghisoni, S.; Baccolo, G.; Blasi, F.; Montesano, D.; Trevisan, M.; Lucini, L. UHPLC-ESI-QTOF-MS profile of polyphenols in Goji berries (Lycium barbarum L.) and its dynamics during in vitro gastrointestinal digestion and fermentation. J. Funct. Foods 2018, 40, 564–572. [Google Scholar] [CrossRef]
- Liu, S.; Marsol-Vall, A.; Laaksonen, O.; Kortesniemi, M.; Yang, B. Characterization and Quantification of Nonanthocyanin Phenolic Compounds in White and Blue Bilberry (Vaccinium myrtillus) Juices and Wines Using UHPLC-DAD-ESI-QTOF-MS and UHPLC-DAD. J. Agric. Food Chem. 2020, 68, 7734–7744. [Google Scholar] [CrossRef]
- Barrientos, R.; Fernández-Galleguillos, C.; Pastene, E.; Simirgiotis, M.; Romero-Parra, J.; Ahmed, S.; Echeverría, J. Metabolomic Analysis, Fast Isolation of Phenolic Compounds, and Evaluation of Biological Activities of the Bark from Weinmannia trichosperma Cav. (Cunoniaceae). Front. Pharmacol. 2020, 11, 780. [Google Scholar] [CrossRef]
- Barrientos, R.E.; Ahmed, S.; Cortés, C.; Fernández-Galleguillos, C.; Romero-Parra, J.; Simirgiotis, M.J.; Echeverría, J. Chemical Fingerprinting and Biological Evaluation of the Endemic Chilean Fruit Greigia sphacelata (Ruiz and Pav.) Regel (Bromeliaceae) by UHPLC-PDA-Orbitrap-Mass Spectrometry. Molecules 2020, 25, 3750. [Google Scholar] [CrossRef]
- Brito, A.; Areche, C.; Sepúlveda, B.; Kennelly, E.J.; Simirgiotis, M.J. Anthocyanin characterization, total phenolic quantification and antioxidant features of some chilean edible berry extracts. Molecules 2014, 19, 10936–10955. [Google Scholar] [CrossRef] [Green Version]
- Simirgiotis, M.; Quispe, C.; Bórquez, J.; Areche, C.; Sepúlveda, B. Fast Detection of Phenolic Compounds in Extracts of Easter Pears (Pyrus communis) from the Atacama Desert by Ultrahigh-Performance Liquid Chromatography and Mass Spectrometry (UHPLC–Q/Orbitrap/MS/MS). Molecules 2016, 21, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larrazábal-Fuentes, M.J.; Fernández-Galleguillos, C.; Palma-Ramírez, J.; Romero-Parra, J.; Sepúlveda, K.; Galetovic, A.; González, J.; Paredes, A.; Bórquez, J.; Simirgiotis, M.J.; et al. Chemical Profiling, Antioxidant, Anticholinesterase, and Antiprotozoal Potentials of Artemisia copa Phil. (Asteraceae). Front. Pharmacol. 2020, 11, 594174. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Aspee, F.; Quispe, C.; Soriano, M.d.P.C.; Fuentes Gonzalez, J.; Hüneke, E.; Theoduloz, C.; Schmeda-Hirschmann, G. Antioxidant activity and characterization of constituents in copao fruits (Eulychnia acida Phil., Cactaceae) by HPLC–DAD–MS/MSn. Food Res. Int. 2014, 62, 286–298. [Google Scholar] [CrossRef]
- Simirgiotis, M.J.; Bórquez, J.; Schmeda-Hirschmann, G. Antioxidant capacity, polyphenolic content and tandem HPLC–DAD–ESI/MS profiling of phenolic compounds from the South American berries Luma apiculata and L. chequén. Food Chem. 2013, 139, 289–299. [Google Scholar] [CrossRef]
- Ramirez, J.E.; Zambrano, R.; Sepúlveda, B.; Kennelly, E.J.; Simirgiotis, M.J. Anthocyanins and antioxidant capacities of six Chilean berries by HPLC-HR-ESI-ToF-MS. Food Chem. 2015, 176, 106–114. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-throughput assay of oxygen radical absorbance capacity (ORAC) using a multichannel liquid handling system coupled with a microplate fluorescence reader in 96-well format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef] [PubMed]
- Zengin, G.; Uysal, S.; Ceylan, R.; Aktumsek, A. Phenolic constituent, antioxidative and tyrosinase inhibitory activity of Ornithogalum narbonense L. from Turkey: A phytochemical study. Ind. Crop. Prod. 2015, 70, 1–6. [Google Scholar] [CrossRef]
- Petersson, G.A.; Bennett, A.; Tensfeldt, T.G.; Al-Laham, M.A.; Shirley, W.A.; Mantzaris, J. A complete basis set model chemistry. I. The total energies of closed-shell atoms and hydrides of the first-row elements. J. Chem. Phys. 1988, 89, 2193–2218. [Google Scholar] [CrossRef]
- Adamo, C.; Barone, V. Toward reliable density functional methods without adjustable parameters: The PBE0 model Seeking for parameter-free double-hybrid functionals: The PBE0-DH model Accurate excitation energies from time-dependent density functional theory: Assessing the PBE0. J. Chem. Phys. 1999, 110, 6158–6170. [Google Scholar] [CrossRef]
- Greenblatt, H.M.; Kryger, G.; Lewis, T.; Silman, I.; Sussman, J.L. Structure of acetylcholinesterase complexed with (−)-galanthamine at 2.3 A resolution. FEBS Lett. 1999, 463, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Nachon, F.; Carletti, E.; Ronco, C.; Trovaslet, M.; Nicolet, Y.; Jean, L.; Renard, P.Y. Crystal structures of human cholinesterases in complex with huprine W and tacrine: Elements of specificity for anti-Alzheimer’s drugs targeting acetyl- and butyryl-cholinesterase. Biochem. J. 2013, 453, 393–399. [Google Scholar] [CrossRef] [Green Version]
- Ismaya, W.T.; Rozeboom, H.J.; Weijn, A.; Mes, J.J.; Fusetti, F.; Wichers, H.J.; Dijkstra, B.W. Crystal structure of agaricus bisporus mushroom tyrosinase: Identity of the tetramer subunits and interaction with tropolone. Biochemistry 2011, 50, 5477–5486. [Google Scholar] [CrossRef] [Green Version]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo, Q.A.; Triana, J.; Eiroa, J.L.; Calcul, L.; Rivera, E.; Wojtas, L.; Padrón, J.M.; Boberieth, L.; Keramane, M.; Abel-Santos, E.; et al. ent-Labdane Diterpenoids from the Aerial Parts of Eupatorium obtusissmum. J. Nat. Prod. 2016, 79, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Pal, D.; Sur, S.; Mandal, S.; Saha, P.; Panda, C.K. Pongapin and Karanjin, furanoflavanoids of Pongamia pinnata, induce G2/M arrest and apoptosis in cervical cancer cells by differential reactive oxygen species modulation, DNA damage, and nuclear factor kappa-light-chain-enhancer of activated B cell signal. Phyther. Res. 2019, 33, 1084–1094. [Google Scholar] [CrossRef] [PubMed]
- Picariello, G.; Sciammaro, L.; Siano, F.; Volpe, M.G.; Puppo, M.C.; Mamone, G. Comparative analysis of C-glycosidic flavonoids from Prosopis spp. and Ceratonia siliqua seed germ flour. Food Res. Int. 2017, 99, 730–738. [Google Scholar] [CrossRef]
- Liu, J.; Hefni, M.E.; Witthöft, C.M. Characterization of Flavonoid Compounds in Common Swedish Berry Species. Foods 2020, 9, 358. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.L.; Liu, Z.Z.; Xie, M.; Zhang, X.; Yang, Y.; Yan, C.; Qiao, W.L.; Liu, W.R.; Jiang, R.; Wang, X.H.; et al. Salicylic Acid Derivatives and Other Components from Gaultheria trichoclada. Chem. Nat. Compd. 2016, 52, 301–303. [Google Scholar] [CrossRef]
- Hajdú, Z.; Hohmann, J.; Forgo, P.; Martinek, T.; Dervarics, M.; Zupkó, I.; Falkay, G.; Cossuta, D.; Máthé, I. Diterpenoids and flavonoids from the fruits of Vitex agnus-castus and antioxidant activity of the fruit extracts and their constituents. Phyther. Res. 2007, 21, 391–394. [Google Scholar] [CrossRef]
- Ono, M.; Masuoka, C.; Koto, M.; Tateishi, M.; Komatsu, H.; Kobayashi, H.; Igoshi, K.; Ito, Y.; Okawa, M.; Nohara, T. Antioxidant ortho-benzoyloxyphenyl acetic acid ester, vaccihein A, from the fruit of rabbiteye blueberry (Vaccinium ashei). Chem. Pharm. Bull. 2002, 50, 1416–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willems, J.L.; Khamis, M.M.; Mohammed Saeid, W.; Purves, R.W.; Katselis, G.; Low, N.H.; El-Aneed, A. Analysis of a series of chlorogenic acid isomers using differential ion mobility and tandem mass spectrometry. Anal. Chim. Acta 2016, 933, 164–174. [Google Scholar] [CrossRef]
- Jia, S.S.; Ma, C.M.; Li, Y.H.; Hao, J.H. Glycosides of phenolic acid and flavonoids from the leaves of Glycyrrhiza uralensis Ficsh. Yao Xue Xue Bao 1992, 27, 441–444. [Google Scholar]
- Llorent-Martinez, E.J.; Spinola, V.; Gouveia, S.; Castilho, P.C. HPLC-ESI-MSn characterization of phenolic compounds, terpenoid saponins, and other minor compounds in Bituminaria bituminosa. Ind. Crop. Prod. 2015, 69, 80–90. [Google Scholar] [CrossRef]
- Petreska, J.; Stefkov, G.; Kulevanova, S.; Alipieva, K.; Bankova, V.; Stefova, M. Phenolic Compounds of Mountain Tea from the Balkans: LC/DAD/ESI/MSn Profile and Content. Nat. Prod. Commun. 2011, 6, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Tajner-Czopek, A.; Gertchen, M.; Rytel, E.; Kita, A.; Kucharska, A.Z.; Sokół-Łętowska, A. Study of antioxidant activity of some medicinal plants having high content of caffeic acid derivatives. Antioxidants 2020, 9, 412. [Google Scholar] [CrossRef]
- Shrestha, A.; Rezk, A.; Said, I.H.; von Glasenapp, V.; Smith, R.; Ullrich, M.S.; Schepker, H.; Kuhnert, N. Comparison of the polyphenolic profile and antibacterial activity of the leaves, fruits and flowers of Rhododendron ambiguum and Rhododendron cinnabarinum. BMC Res. Notes 2017, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kyriacou, M.C.; El-Nakhel, C.; Pannico, A.; Graziani, G.; Soteriou, G.A.; Giordano, M.; Zarrelli, A.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Genotype-Specific Modulatory Effects of Select Spectral Bandwidths on the Nutritive and Phytochemical Composition of Microgreens. Front. Plant Sci. 2019, 10, 1501. [Google Scholar] [CrossRef]
- Nadeem, M.; Mumtaz, M.W.; Danish, M.; Rashid, U.; Mukhtar, H.; Irfan, A.; Anwar, F.; Saari, N. UHPLC-QTOF-MS/MS metabolites profiling and antioxidant/antidiabetic attributes of Cuscuta reflexa grown on Casearia tomentosa: Exploring phytochemicals role via molecular docking. Int. J. Food Prop. 2020, 23, 918–940. [Google Scholar] [CrossRef]
- Gómez-Romero, M.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Metabolite profiling and quantification of phenolic compounds in methanol extracts of tomato fruit. Phytochemistry 2010, 71, 1848–1864. [Google Scholar] [CrossRef] [PubMed]
- Razeq, F.M.; Kosma, D.K.; França, D.; Rowland, O.; Molina, I. Extracellular lipids of Camelina sativa: Characterization of cutin and suberin reveals typical polyester monomers and unusual dicarboxylic fatty acids. Phytochemistry 2021, 184, 112665. [Google Scholar] [CrossRef]
- Chen, C.Y.; Lin, R.J.; Huang, J.C.; Wu, Y.H.; Cheng, M.J.; Hung, H.C.; Lo, W.L. Chemical constituents from the whole plant of Gaultheria itoana Hayata. Chem. Biodivers. 2009, 6, 1737–1743. [Google Scholar] [CrossRef]
- Davini, E.; Esposito, P.; Iavarone, C.; Trogolo, C. Structure and configuration of unedide, an iridoid glucoside from Arbutus unedo. Phytochemistry 1981, 20, 1583–1585. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, L.; Gao, J.; Wu, C.; Han, L.; Liu, E.; Shi, P.; Gao, X.; Wang, T. New secoiridoids from the fruits of Ligustrum lucidum Ait with triglyceride accumulation inhibitory effects. Fitoterapia 2013, 91, 107–112. [Google Scholar] [CrossRef]
- Dai, L.-P.; Zhang, L.-X.; Liu, Y.-L.; Wu, H.; Liu, R.-X.; Zhao, M.; Tian, S.-S.; Jiang, X.; Chen, S.-Q. Isolation and purification of diterpenoids from the aerial parts of Isodon excisoides target-guided by UPLC-LTQ-Orbitrap-MS. Nat. Prod. Res. 2018, 32, 2424–2430. [Google Scholar] [CrossRef]
- Bozov, P.I.; Coll, J. Neo-Clerodane Diterpenoids from Scutellaria altissima. Nat. Prod. Commun. 2015, 10, 13–16. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wei, T.; Zhou, X. Inhibitory Effects of Cortex Dictamni Aqueous Extract on Dipeptidyl Peptidase I and Chymase Activities and the Screening of Active Ingredients in Cortex Dictamni Based on Molecular Docking Technique. J. Biol. Life Sci. 2021, 12, 19–30. [Google Scholar] [CrossRef]
- Quesada-Romero, L.; Fernández-Galleguillos, C.; Bergmann, J.; Amorós, M.-E.; Jiménez-Aspee, F.; González, A.; Simirgiotis, M.; Rossini, C. Phenolic Fingerprinting, Antioxidant, and Deterrent Potentials of Persicaria maculosa Extracts. Molecules 2020, 25, 3054. [Google Scholar] [CrossRef] [PubMed]
- Mcdougall, G.J.; Austin, C.; Van Schayk, E.; Martin, P. Salal (Gaultheria shallon) and aronia (Aronia melanocarpa) fruits from Orkney: Phenolic content, composition and effect of wine-making. Food Chem. 2016, 205, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Bravo, K.; Alzate, F.; Osorio, E. Fruits of selected wild and cultivated Andean plants as sources of potential compounds with antioxidant and anti-aging activity. Ind. Crop. Prod. 2016, 85, 341–352. [Google Scholar] [CrossRef]
- Colak, N.; Primetta, A.K.; Riihinen, K.R.; Jaakola, L.; Grúz, J.; Strnad, M.; Torun, H.; Ayaz, F.A. Phenolic compounds and antioxidant capacity in different-colored and non-pigmented berries of bilberry (Vaccinium myrtillus L.). Food Biosci. 2017, 20, 67–78. [Google Scholar] [CrossRef]
- Céspedes, C.L.; Valdez-Morales, M.; Avila, J.G.; El-Hafidi, M.; Alarcón, J.; Paredes-López, O. Phytochemical profile and the antioxidant activity of Chilean wild black-berry fruits, Aristotelia chilensis (Mol) Stuntz (Elaeocarpaceae). Food Chem. 2010, 119, 886–895. [Google Scholar] [CrossRef]
- Cásedas, G.; Les, F.; González-Burgos, E.; Gómez-Serranillos, M.P.; Smith, C.; López, V. Cyanidin-3-O-glucoside inhibits different enzymes involved in central nervous system pathologies and type-2 diabetes. South Afr. J. Bot. 2019, 120, 241–246. [Google Scholar] [CrossRef]
- Llorent-Martínez, E.J.; Zengin, G.; Lobine, D.; Molina-García, L.; Mollica, A.; Mahomoodally, M.F. Phytochemical characterization, in vitro and in silico approaches for three Hypericum species. New J. Chem. 2018, 42, 5204–5214. [Google Scholar] [CrossRef]
- Nile, A.; Gansukh, E.; Park, G.S.; Kim, D.H.; Hariram Nile, S. Novel insights on the multi-functional properties of flavonol glucosides from red onion (Allium cepa L) solid waste—In vitro and in silico approach. Food Chem. 2021, 335, 127650. [Google Scholar] [CrossRef] [PubMed]
- Beara, I.N.; Torović, L.D.; Pintać, D.; Majkić, T.M.; Orčić, D.Z.; Mimica-Dukić, N.M.; Lesjak, M.M. Polyphenolic profile, antioxidant and neuroprotective potency of grape juices and wines from Fruška Gora region (Serbia). Int. J. Food Prop. 2018, 20, S2552–S2568. [Google Scholar] [CrossRef] [Green Version]
- Tkacz, K.; Wojdyło, A.; Turkiewicz, I.P.; Ferreres, F.; Moreno, D.A.; Nowicka, P. UPLC-PDA-Q/TOF-MS profiling of phenolic and carotenoid compounds and their influence on anticholinergic potential for AChE and BuChE inhibition and on-line antioxidant activity of selected Hippophaë rhamnoides L. cultivars. Food Chem. 2019, 309, 125766. [Google Scholar] [CrossRef]
- Budryn, G.; Grzelczyk, J.; Jaśkiewicz, A.; Żyżelewicz, D.; Pérez-Sánchez, H.; Cerón-Carrasco, J.P. Evaluation of butyrylcholinesterase inhibitory activity by chlorogenic acids and coffee extracts assed in ITC and docking simulation models. Food Res. Int. 2018, 109, 268–277. [Google Scholar] [CrossRef]
- Alam, F.; Saqib, Q.N.U.; Shah, A.J.; Ashraf, M.; Al Ain, Q. Gut modulatory and butyrylcholinesterase inhibitory activities of Gaultheria trichophylla. Pharm. Biol. 2016, 54, 2917–2921. [Google Scholar] [CrossRef] [Green Version]
- Salehi, B.; Sharifi-Rad, J.; Cappellini, F.; Reiner, Ž.; Zorzan, D.; Imran, M.; Sener, B.; Kilic, M.; El-Shazly, M.; Fahmy, N.M.; et al. The Therapeutic Potential of Anthocyanins: Current Approaches Based on Their Molecular Mechanism of Action. Front. Pharmacol. 2020, 11, 1300. [Google Scholar] [CrossRef] [PubMed]
- Viegas, O.; Faria, M.A.; Sousa, J.B.; Vojtek, M.; Gonçalves-Monteiro, S.; Suliburska, J.; Diniz, C.; Ferreira, I.M.P.L.V.O. Delphinidin-3-O-glucoside inhibits angiogenesis via VEGFR2 downregulation and migration through actin disruption. J. Funct. Foods 2019, 54, 393–402. [Google Scholar] [CrossRef]
- León-González, A.J.; Sharif, T.; Kayali, A.; Abbas, M.; Dandache, I.; Etienne-Selloum, N.; Kevers, C.; Pincemail, J.; Auger, C.; Chabert, P.; et al. Delphinidin-3-O-glucoside and delphinidin-3-O-rutinoside mediate the redox-sensitive caspase 3-related pro-apoptotic effect of blackcurrant juice on leukaemia Jurkat cells. J. Funct. Foods 2015, 17, 847–856. [Google Scholar] [CrossRef]
- Li, Y.; Li, X.; Cuiping, C.; Pu, R.; Weihua, Y. Study on the Anticancer Effect of an Astragaloside- And Chlorogenic Acid-Containing Herbal Medicine (RLT-03) in Breast Cancer. Evid. Based Complement. Altern. Med. 2020, 2020, 1515081. [Google Scholar] [CrossRef]
- Cincin, Z.B.; Unlu, M.; Kiran, B.; Bireller, E.S.; Baran, Y.; Cakmakoglu, B. Molecular mechanisms of quercitrin-induced apoptosis in non-small cell lung cancer. Arch. Med. Res. 2014, 45, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.M.; Deng, X.T.; Zhou, J.; Li, Q.P.; Ge, X.X.; Miao, L. Pharmacological basis and new insights of quercetin action in respect to its anti-cancer effects. Biomed. Pharmacother. 2020, 121, 109604. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Z.; Jin, J.; Zhu, S.X.; He, G.Q.; Li, S.H.; Wang, J.; Cai, Y. Quercetin pretreatment enhances the radiosensitivity of colon cancer cells by targeting Notch-1 pathway. Biochem. Biophys. Res. Commun. 2020, 523, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Iqubal, M.K.; Imtiyaz, K.; Saleem, S.; Mittal, S.; Rizvi, M.M.A.; Ali, J.; Baboota, S. Topical nanostructured lipid carrier gel of quercetin and resveratrol: Formulation, optimization, in vitro and ex vivo study for the treatment of skin cancer. Int. J. Pharm. 2020, 587, 119705. [Google Scholar] [CrossRef]
- Teekaraman, D.; Elayapillai, S.P.; Viswanathan, M.P.; Jagadeesan, A. Quercetin inhibits human metastatic ovarian cancer cell growth and modulates components of the intrinsic apoptotic pathway in PA-1 cell line. Chem. Biol. Interact. 2019, 300, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Ni, G.; Tang, Y.; Li, M.; He, Y.; Rao, G. Synthesis of scutellarein derivatives with a long aliphatic chain and their biological evaluation against human cancer cells. Molecules 2018, 23, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | RT | λ Max (nm) | Elemental Composition [M + H]+ [M − H]− | Measured Mass (m/z) | Theorical Mass (m-z) | Accuracy (ppm) | MSn Ions | Tentative Identification |
|---|---|---|---|---|---|---|---|---|
| 1a | 11.45 | 234–282–520 | C20H19O11+ | 435.10989 | 435.09219 | 40.685 | 303.05676, 247.12254, 178.13483, 155.15498, 130.00858 | Delphinidin-3-O-arabinoside |
| 2a | 11.94 | 239–522 | C21H21O12+ | 465.12381 | 465.10275 | 45.144 | 303.05667, 247.12254, 183.10243, 130.15967 | Delphinidin 3-O-galactoside * |
| 3a | 12.26 | 280–521 | C21H21O11+ | 449.12598 | 449.10784 | 40.394 | 287.06052, 247.12250, 171.09984, 147.04466 | Cyanidin 3-O-galactoside * |
| 4a | 12.58 | 281–517 | C20H19O10+ | 419.11234 | 419.09727 | 35.949 | 287.06049, 209.04585, 147.04463 | Cyanidin-3-O-arabinoside * |
| 5a | 12.99 | 238–309–516 | C22H23O11+ | 463.10559 | 463.12349 | −38.648 | 301.16412, 225.81639, 171.09319 | Peonidin-3-O-glucoside |
| 1 | 2.94 | 232 | C18H17O9− | 377.08575 | 377.08781 | −2.542 | 279.19812, 218.78490, 162.83897 | Vaccihein A |
| 2 | 3.06 | 232 | C16H23O12− | 407.11966 | 407.11950 | 3.089 | 295.51428, 156.00233, 119.60213 | Unedide |
| 3 | 3.38 | 256 | C19H11O6− | 335.05420 | 335.05611 | −2.431 | 268.80099, 215.03267, 160.84152 | Pongapin |
| 4 | 11.58 | 234–297 | C16 H17O9− | 353.08820 | 353.08781 | 4.224 | 314.02936, 191.05566, 135.04482, 109.02880 | Caffeoylquinic acid derivative |
| 5 | 12.17 | 236–282 | C12H13O8− | 285.06189 | 285.06159 | 4.898 | 219.44096, 167.07101, 108.02096 | Uralenneoside |
| 6 | 12.47 | 236–299 | C16H17O9− | 353.08804 | 353.08781 | 3.771 | 707.18433, 191.05592 | Chlorogenic acid |
| 7 | 12.67 | 236–282–517 | C20H17O10− | 417.08307 | 417.08272 | 3.469 | 163.03966, 128.04340, 119.04950 | Coumaric acid glucoside |
| 8 | 12.98 | 236–312 | C26H27O14− | 563.14105 | 563.14063 | 2.696 | 281.06680, 191.05582, 115.91994 | Isoschaftoside |
| 9 | 13.25 | 237–323 | C16H15O8− | 335.07755 | 335.07724 | 4.196 | 213.96364, 179.03458, 135.04459 | 5-O-Caffeoylshikimic acid |
| 10 | 13.39 | 238–326 | C25H27O14− | 551.14050 | 551.14063 | 1.757 | 179.03439, 135.04460 | Caffeic acid derivative |
| 11 | 13.50 | 254–354 | C21H19O13− | 479.08319 | 479.08311 | 2.448 | 317.09708, 289.08298, 213.96367, 115.91998 | Myricetin-O-hexoside |
| 12 | 13.70 | 238 | C20H17O12− | 449.07294 | 449.07255 | 3.313 | 319.04620, 183.02954, 134.89410 | Methyl gallate derivative |
| 13 | 13.96 | 241–304 | C25H27O13− | 535.14545 | 535.14571 | 1.556 | 357.11948, 163.03975, 119.04951 | p-coumaroyl monotropein isomer |
| 14 | 14.05 | 255–352 | C21H19O12− | 463.08838 | 463.08820 | 2.759 | 385.11417, 281.06665, 163.03955 | Myricetin-O-rhamnoside |
| 15 | 14.34 | 243–351 | C20H17O11− | 433.07782 | 433.07763 | 2.961 | 300.02753, 279.05096, 151.00316, 138.07101, 107.01299 | Quercetin 3-O-ribose |
| 16 | 14.41 | 255–351 | C20H17O11− | 433.07776 | 433.07763 | 2.822 | 300.02744, 151.03909 134.89404, 115.91995 | Quercetin 3-O-arabinoside * |
| 17 | 14.53 | 255–347 | C20H17O11− | 433.07773 | 433.07763 | 2.753 | 301.03571, 263.05963, 151.00317, 115.92000 | Quercetin 3-O-xyloside |
| 18 | 14.62 | 245–317 | C21H19O11− | 447.09341 | 447.09329 | 2.734 | 300.02750, 251.05937, 115.92000, 151.00310 | Quercitrin |
| 19 | 15.28 | 244–339 | C21H17O12− | 461.07294 | 461.07255 | 3.227 | 285.04007, 174.95566, 151.03946, 132.86728, 108.02094 | Luteolin-7-glucuronide |
| 20 | 15.49 | 244 | C21H17O12− | 461.07297 | 461.07255 | 3.292 | 309.17999, 195.96376 151.00304, 123.04469 | Galloyl derivative |
| 21 | 16.21 | 244 | C23H29O11− | 481.17169 | 481.17154 | 2.602 | 255.82225, 160.84169, 115.92001 | Nuzhenal B |
| 22 | 17.71 | 248 | C15H9O7− | 301.03555 | 301.03538 | 4.222 | 285.04120, 236.97911, 151.00301, 121.02882, 107.01293 | Quercetin |
| 23 | 20.86 | 245 | C27H45O13− | 577.28729 | 577.28657 | 3.157 | 450.53427, 425.21106, 207.85930 | Procyanidin B-type dimer |
| 24 | 21.32 | 247 | C18H33O5− | 329.23383 | 329.23335 | 4.797 | 197.80783, 160.84171, 122.02425 | Pinellic acid |
| 25 | 22.58 | 250 | C20H29O6− | 365.19760 | 365.19696 | 4.751 | 314.38947, 221.08304, 153.09119 | Enmenol |
| 26 | 23.56 | 254–290 | C23H19O12− | 487.08856 | 487.08820 | 3.054 | 427.03278, 311.22318, 191.10770 | Caffeoyl feruloyl tartaric acid |
| 27 | 24.13 | 255 | C18H33O4− | 313.23877 | 313.23843 | 4.578 | 284.97467, 174.95573, 149.92661, 116.95750 | Octadecanedioic acid |
| 28 | 24.81 | 257 | C20H39O3− | 327.29071 | 327.29047 | 4.089 | 266.28955, 206.63348, 136.34750 | 2-Hydroxyeicosanoic acid |
| 29 | 29.03 | 258 | C24H35O8− | 451.23380 | 451.23374 | 2.561 | 297.15295, 265.14801, 235.09744 | Scupolin I |
| 30 | 29.41 | 258 | C18H15O7− | 343.08267 | 343.08233 | 4.199 | 279.16360, 265.14801, 233.08183, 162.83871 | Eupatorin |
| 31 | 29.81 | 258 | C27H47O14− | 595.28961 | 595.29713 | −10.789 | 253.98213, 198.76825, 144.82085 | Dictamnoside M |
| Assay | Gaultheria pumila | Standard |
|---|---|---|
| Total phenolics A | 189.2 ± 0.2 | - |
| Total flavonoids B | 51.8 ± 0.1 | - |
| Total anthocyanin C | 47.3 ± 0.2 | - |
| FRAP D | 134.1 ± 0.1 | - |
| ORAC E | 4251.6 ± 16.9 | - |
| DPPH F | 92.8 ± 0.1 | Gallic acid: 0.55 ± 0.1 |
| Assay | AChE Inhibition IC50 (µg/mL) | BChE Inhibition IC50 (µg/mL) | Tyrosinase Inhibition IC50 (µg/mL) |
|---|---|---|---|
| Gaultheria pumila | 7.7 ± 0.3 | 34.5 ± 0.5 | 3.3 ± 0.2 |
| Galanthamine | 0.3 ± 0.3 | 3.82 ± 0.2 | - |
| Kojic acid | - | - | 10.0 ± 1.9 |
| Compound | Binding Energy (kcal/mol) Acetylcholinesterase | Binding Energy (kcal/mol) Butyrylcholinesterase | Binding Energy (kcal/mol) Tyrosinase |
|---|---|---|---|
| Scupolin I (29) | −10.582 | −7.361 | −4.303 |
| Unedide (2) | −12.081 | −10.972 | −10.643 |
| Nuzhenal B (21) | −15.289 | −11.079 | −10.576 |
| Myricetin O-rhamnoside (14) | −15.175 | −11.581 | −10.528 |
| Luteolin 7-glucuronide (19) | −13.757 | −11.777 | −11.323 |
| Galantamine | −12.989 | −7.125 | - |
| Kojic acid | - | - | −6.050 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Galleguillos, C.; Quesada-Romero, L.; Puerta, A.; Padrón, J.M.; Souza, E.; Romero-Parra, J.; Simirgiotis, M.J. UHPLC-MS Chemical Fingerprinting and Antioxidant, Antiproliferative, and Enzyme Inhibition Potential of Gaultheria pumila Berries. Metabolites 2021, 11, 523. https://doi.org/10.3390/metabo11080523
Fernández-Galleguillos C, Quesada-Romero L, Puerta A, Padrón JM, Souza E, Romero-Parra J, Simirgiotis MJ. UHPLC-MS Chemical Fingerprinting and Antioxidant, Antiproliferative, and Enzyme Inhibition Potential of Gaultheria pumila Berries. Metabolites. 2021; 11(8):523. https://doi.org/10.3390/metabo11080523
Chicago/Turabian StyleFernández-Galleguillos, Carlos, Luisa Quesada-Romero, Adrián Puerta, José M. Padrón, Ernane Souza, Javier Romero-Parra, and Mario J. Simirgiotis. 2021. "UHPLC-MS Chemical Fingerprinting and Antioxidant, Antiproliferative, and Enzyme Inhibition Potential of Gaultheria pumila Berries" Metabolites 11, no. 8: 523. https://doi.org/10.3390/metabo11080523