
The Upper Airway Microbiota, Environmental Exposures, Inflammation, and Disease


1
Department of Biomedical Science, University of Guelph, Guelph, ON N1G 2W1, Canada
2
Department of Medicine, Division of Pulmonary & Critical Care Medicine, State University of New York Downstate Medical Centre, Brooklyn, NY 11203, USA
3
Department of Biology, University of Western Ontario, London, ON N6A 5B7, Canada
*
Author to whom correspondence should be addressed.
Medicina 2021, 57(8), 823; https://doi.org/10.3390/medicina57080823
Submission received: 15 July 2021
/
Revised: 9 August 2021
/
Accepted: 10 August 2021
/
Published: 14 August 2021
(This article belongs to the Special Issue Respiratory Immune Responses during Infection and Pollution Inhalation)
Abstract
:Along with playing vital roles in pathogen exclusion and immune system priming, the upper airways (UAs) and their microbiota are essential for myriad physiological functions such as conditioning and transferring inhaled air. Dysbiosis, a microbial imbalance, is linked with various diseases and significantly impedes the quality of one’s life. Daily inhaled exposures and/or underlying conditions contribute to adverse changes to the UA microbiota. Such variations in the microbial community exacerbate UA and pulmonary disorders via modulating inflammatory and immune pathways. Hence, exploring the UA microbiota’s role in maintaining homeostasis is imperative. The microbial composition and subsequent relationship with airborne exposures, inflammation, and disease are crucial for strategizing innovating UA diagnostics and therapeutics. The development of a healthy UA microbiota early in life contributes to normal respiratory development and function in the succeeding years. Although different UA cavities present a unique microbial profile, geriatrics have similar microbes across their UAs. This lost community segregation may contribute to inflammation and disease, as it stimulates disadvantageous microbial–microbial and microbial–host interactions. Varying inflammatory profiles are associated with specific microbial compositions, while the same is true for many disease conditions and environmental exposures. A shift in the microbial composition is also detected upon the administration of numerous therapeutics, highlighting other beneficial and adverse side effects. This review examines the role of the UA microbiota in achieving homeostasis, and the impact on the UAs of environmental airborne pollutants, inflammation, and disease.
Keywords:
upper airways; microbiota; dysbiosis; inflammation; airborne exposures; age; environmental exposure; disease1. Introduction
The upper airways (UAs) compose the respiratory system’s primary elements to conduct and process air to the lungs [1]. An average adult transfers over 7000 L of air daily through their UAs [1,2]. Hence, the airways have developed a complex immune system to filter the unsterile inhaled air [2,3]. Daily environmental contaminants, such as pollutants and pathogens, cause a shift in UA homeostasis, thereby activating the innate immune response and leading to a local inflammatory reaction [3,4]. The UAs harbor complex microbial communities to aid with their immense physiological responsibility. Similar to the gut microbiota, the upper airway symbionts are diverse, including pathogenic, commensal, and symbiotic microorganisms [2,5,6,7]. Collectively, the human body serves as a host for about 100 trillion microbes [7,8]. They endow us with crucial qualities such as aiding in metabolism, pathogen exclusion, and immune system training [5,7,9,10]. Dysbiosis, or loss of the beneficial and commensal microbes that prevent the colonization of opportunistic pathogens, is associated with local inflammation and exacerbation of many pulmonary disorders, such as chronic obstructive pulmonary disease (COPD), asthma, chronic rhinosinusitis (CRS), and various UA infections (UAIs). UAIs are a leading cause of loss of quality of life, and often death [11]. An average adult experiences two to four annual respiratory infections [11]. Consequently, UAIs constitute a yearly burden of 75 million physician visits, 150 million sick days away from work, and more than USD 10 billion in insurance costs, solely in American adults [11].
Several environmental factors and/or pre-existing conditions contribute to UA dysbiosis. This review examines the role of the UA microbiota in achieving homeostasis, and the impact of environmental airborne pollutants, inflammation, and disease on the UAs and their symbionts. It is also important to note that age, weight, and diet are also important factors for microbiota composition, diversity, and stability [12].
2. Upper Airway Microbial Niches
The UAs are defined as the nasal cavity, sinuses, nasopharynx, oropharynx, and laryngopharynx (Figure 1), all of which display variable niche parameters [6,13]. The development and maturation of the UAs is a complicated multistage process that occurs both pre- and postnatally [6,9,14,15]. The specialized resident microbial community plays a vital role in this post-natal development, as germ-free murine models indicate deranged UA physiology [6,9,14]. Microbial interactions lead to the development of highly specialized systems containing diverse niches across the UA cavities, with varying physiological functions [6,9,14,16]. For example, air-filled spaces such as the nasopharyngeal tonsils (adenoids), amongst others, serve as a major location for pathogen recognition and defense responses [17,18]. Each of these UA regions is subjected to specific microbial cellular and physiological gradients [14,19]. Differences in temperature, pH, mucus secretion, and relative oxygen concentration regulate bacterial colonization in the UAs [7,8,14,19].
The nasal cavity is constantly interacting with the external environment [18,20]. It is lined with diverse epithelial cell types, providing varying microenvironments for the microbiota, and leading to high microbial diversity in nasal niches [6,7,8,18,20]. Within the nasal cavity, the turbinates increase the surface area and thus regulate temperature and humidity [21]. As primary elements of the UAs, the anterior nares and nasal vestibule contain sebaceous glands and vibrissae to help moderate their dry and contaminated microhabitat [1,22]. The vibrissae filter large particles (>3 μm) from inhaled air, while smaller particles (0.5–3 μm) are trapped in the mucus layer [1,18,22].
Located adjacent to the nasal vestibule is the middle meatus [1,23]. This region serves as the drainage area for the UA sinuses (anterior ethmoids, maxillary, and frontal sinuses) [18,23]. The UA sinuses are air-filled cavities that are important for warming and humidifying inhaled air, as well as pathogen recognition and many other speculated functions [17,18,23]. These UA segments are composed of ciliated columnar epithelium, producing mucus that is then transported into the nasal cavity [18]. This mucosal drainage establishes local microhabitats in the nasal cavity with unique microbial populations [2,6,17,18]. Located at the ceiling of the nasal cavity is the olfactory area, which is composed of pseudostratified columnar olfactory epithelium that is modified to contain many neurons [6,21,24]. This area is also of interest, as there is a correlation between olfactory function and the composition of the local microbiota [24].
3. The UA Microbiota
Our body’s ecological microhabitats harbor all kinds of microorganisms, such as bacteria, viruses, and other eukaryotes [6,25]. Here, we will primarily restrict our discussion to bacteria in this review, since the bulk of UA microbiota research covers this kingdom. The development of a healthy UA microbiota depends on multiple innate factors such as genetics; however, lifestyle and environmental exposures also help determine the microbial composition [6,25]. Within healthy UAs, varying microbial species and biomass are observed across the nasal, nasopharyngeal, and oropharyngeal cavities [26]. The most abundant phyla in the human respiratory tract are Proteobacteria, Firmicutes, and Bacteroidetes [27]. However, the specific microbial composition may change across an individual’s lifetime (see Table 1) [8,22,25,28,29,30].
3.1. UA Microbiota in Infants
The microbiota develops within the first two years of life and subsequently ensures normal UA development and function [33]. Studies have shown that abnormal colonization of the UAs during infancy contributes to UA diseases such as asthma and other UAIs [33,34]. UA microbiota assemblage begins at birth, and resembles the environment exposed to during birth ((i.e., maternal vaginal (normal birth) or skin microbiota (cesarean)) [5,35]. If an infant is born via cesarean, their nasopharyngeal microbiota represents their mother’s skin microbiota, whereas if born via the vaginal route, their microbiota will resemble the maternal urogenital microbiota [5,35]. Following birth, the UA microbiota is maintained and nourished by feeding [5,36]. Breastfed infants support stable Dolosigranulum/Corynebacterium profiles, whereas formula-fed infants show increased Staphylococcus aureus profiles [5,36]. The microbiota of breast-fed infants seems to offer a stronger protective effect against UAIs [5,36]. Similarly, a Moraxella species (spp.)-dominated UA microbial profile in children has a protective effect against UAIs, yet there are exceptions, with some Moraxella spp. that facilitate viral-associated wheezing in young infants [9,33,34,35]. Additionally, increased nasopharyngeal signatures of Streptococcus can predict asthma in children [9,33,34,35]. Overall observations in the first 2 years of life suggest that the combination of Dolosigranulum, Moraxella, and Corynebacterium enhance protective respiratory physiology to a greater degree relative to a Haemophilus and Streptococcus dominated profiles in infants [35,37]. These latter profiles were associated with UAIs of viral and bacterial (Haemophilus influenza and Streptococcus pneumoniae) origin, as well as an elevated risk for development of respiratory-disease pathologies early in life [35,38,39].
3.2. UA Microbiota in Adults
In comparison to infants, a healthy adult’s UA microbiota has a higher bacterial load and lower diversity [9,19,40,41]. At the anterior nares, Actinobacteria, Firmicutes, and low levels of anaerobic Bacteroidetes have been detected (see Table 1) [5,24,40,42,43,44]. Different regions in the nasal cavity, such as the middle meatus, present a trumping diversity relative to the anterior nares [31]. The anterior nares are colonized by a greater proportion of Firmicutes and Actinobacteria relative to the middle meatus (Table 1) [31], whereas the middle meatus harbors more Proteobacteria in comparison to the anterior nares (Table 1) [31]. Contrastingly, the adult nasopharyngeal microbiota is similar to their anterior nares microbiota at a phylum level [19]. A healthy adult’s paranasal sinuses include bacterial genera such as Staphylococcus, Corynebacterium, and Propionibacterium (Table 1) [45]. Overall, the healthy adult UA microbiota is distinct, depending on the sampling site (middle meatus, oral cavity, oropharynx) [6,46].
3.3. UA Microbiota in the Geriatric Population
As humans age, the distinctive variations defining the microbiota in different regions of the UA gradually diminish [28]. These alterations in the microbial communities are thought to start between the ages of 40–65 years [9,28]. The nasal cavity microbiota that was once dominated by Cutibacterium, Corynebacterium, and Staphylococcus become (at >65 years) more representative of an oropharyngeal microbial community [9,28]. This age-associated melting pot in the UA microbiota may be explained as a consequence of the aging immune system [28], whereby the process of immunosenescence leads to increases in proinflammatory markers (i.e., TNF-α and IL-6) [47]. This may lead to loss of species richness, the opening of new environmental niches, and increased disease susceptibility [28,47]. As we age, the UA niches overlap, leading to increased microbial interactions (See Figure 2) [47]. Microbes continuously interact with the host, but also interact with other species living in their niches [48,49,50]. Such interactions are mostly beneficial; however, when niches overlap nutrients, there is increased competition, driving adverse interbacterial interactions [51,52].
4. UA Inflammation
Inflammation subjects the UA microbiota to selective pressure and may lead to dysbiosis, as it causes alterations in UA niches (see Figure 3) [58]. Clinical features of inflammation mostly stem from increased blood flow to the afflicted region in the UAs [58]. Such clinical features include increased vascular permeability, which facilitates the release of materials and immune cells, as well as increased mucus secretion and mucociliary clearance [58]. Even though the immune response plays a beneficial and protective role, altered inflammation may also lead to further harm, such as tissue injury and edema in the UAs, as well as altered niche parameters and dysbiosis [9,58].
The inflammatory immune response in the UAs is one of the multiple players, including cells such as granulocytes, lymphocytes, and antigen-presenting cells (APCs) [3,58]. Neutrophils are the most abundant granulocyte in the UAs, and their count is positively correlated with a higher degree of inflammation [3,58]. Neutrophilia affects resident S. aureus by causing the expression of more pathogenic profiles and the development of immune-evasion strategies [59,60]. Apart from direct inflammation, this also leads to an increase in mucus viscosity, changing the normal UA microhabitat and thereby contributing to UA dysbiosis [59,60,61]. Signatures of Staphylococcus, Planococcaceae, and Lactococcus were detected in the nasal microbiota of individuals with neutrophilia [62]. In addition, a higher neutrophil count in the mucosa is negatively correlated with microbial diversity [63]. Lymphocytes, which are also detected at higher levels during a period of inflammation, are associated with specific bacterial subsets [62]. A high lymphocyte count in nasal lavage samples was associated with increased colonization of Staphylococcus, Rothia, and Enterococcus within the nasal microbiota [62]. Conversely, the microbiota may also influence the host immune and inflammatory response. Commonly detected in the UA microbiota, Propionibacterium can produce bacteriocin, an antimicrobial peptide (AMP) that mitigates the inflammation associated with the overgrowth of pathogenic bacteria [64].
Noncellular mediators such as AMPs, cytokines, chemokines, immunoglobulins, arachidonic acid metabolites, and reactive oxygen species (ROS) also contribute to inflammation and thereby perturbate the UA microbiota. The nasal microbiota of subjects with increased concentrations of IL-6 is enriched with Moraxella [62]. High levels of IL-8 were associated with an increased presence of Staphylococcus, Veillonella, Lachnospiraeae, Bacteroides, and Planococcaceae in the nasal microbiota [62].
Disease states that drive UA chronic inflammation, such as cystic fibrosis (CF), obstructive sleep apnea, CRS, and asthma, also impact the UA inflammatory profile and microbiota [65]. CF patients with higher colonization of S. aureus within their UAs had distinct levels of inflammatory biomarkers in nasal lavage samples [65]. Notable levels of IL-6, IL-8, IL-1β, MMP-9, and neutrophil elastase were detected [65]. This suggests that increased colonization of S. aureus in diseased UAs could further contribute to the inflammatory response [65]. S. aureus in low-infection colony numbers triggers the secretion of the anti-inflammatory cytokine IL-10, yet when present in higher colony numbers, it reduces IL-10, leading to a more proinflammatory response [65,66,67].
5. Environmental Exposures and the UA Microbiota
Acute changes in environmental factors cause perturbations to the UA microbiota [30,68,69]. In this section, we will define the main environmental exposures that contribute to UA microbiota dysbiosis.
5.1. Seasonal Influences
Seasons contribute to changes in the UA microbiota [69]. Infants and children presented a higher incidence of UAI and S. pneumoniae colonization throughout winter [70,71,72]. Wintertime also increased the abundance of Proteobacteria and increased carriage of Fusobacteria and Cyanobacteria [73], whereas infants with respiratory infections have increased carriage of Haemophilus across spring and summer [33]. During the autumn–winter seasons, infants with respiratory infections presented with decreased carriage of Moraxella [33]. In adults, increased presence of M. catarrhalis and coronavirus is detected during the winter, and Klebsiella pneumoniae presents with higher densities in the summertime [72].
5.2. Air Pollution
Air pollution, which is a mixture of harmful gases and particulate matter (PM), is a major contributor to dysbiosis and many respiratory diseases [74,75]. Globally, almost 90% of people live in areas with suboptimal air quality, according to the WHO. Air pollutants, such as nitrogen and sulfur oxides, as well as ozone, directly irritate the UA epithelium by causing increased oxidative stress and local inflammation [76]. Carcinogens such as diesel emissions and household pollutants arise from burning fuels such as wood and coal [77,78]. PM are foreign substances (i.e., a mixture of solid particles and liquid droplets) found in air that are defined based on size and aerodynamic qualities [79,80]. Larger PM (~10 μm) may get stuck in the UA, while smaller PM (~4 μm) is more likely to travel down the bronchioles and alveoli [79,80]. Short-term exposure to PM influenced the nasal microbiota of 40 healthy subjects by decreasing indices of bacterial diversity, which resulted in increased susceptibility to UAIs (Table 2) [81]. Similarly, exposure to high concentrations of PM for three consecutive days was followed by an increased pharyngeal abundance of particular taxa (Table 2) [82]. Air pollution (PM2.5 and NO2) is associated with Ružička dissimilarity and the abundance of Corynebacteriaceae (NO2 only) in healthy infants [83].
5.3. Cigarette Smoke Exposure
Cigarette smoke is a major contributor to disease and directly contacts the UA surfaces [30,82,86,90,91]. It directly impacts the microbiota; for example, causing oxidation of antimicrobial activity [82]. This change in niche parameters drives the colonization of Gram-positive anaerobic microbes (Table 2) in a smoker’s nasopharynx [30]. The recolonization of smokers’ UAs may also include pathogens associated with UAIs and endocarditis [30]. Notably, a cigarette may also serve as a source of opportunistic pathogens, as some commercially available cigarettes have bacterial signatures (Table 2) [86]. Passive smokers also suffered from smoke-induced dysbiosis (Table 2) [84,85]. Higher levels of S. pneumoniae were detected in children with smoking parents [86]. There was also an observed increase in the prevalence of various UAIs [82,87,88]. Cigarette smoke enhances bacterial attachment, colonization, and biofilm formation in various mechanisms, including the increased production of the bacterial fimbrial protein, FimA [29,82,92,93]. Cigarette smoke alters host physiological functions such as mucociliary clearance, which in turn impairs the innate immune response to pathogens and the UA microenvironments [32,91,94]. Fortunately, smoke-induced dysbiosis can be alleviated by smoke cessation for approximately one year [29,82,85]. Partial recovery of UA microbiota and physiology was also detected [30,82,95].
6. Disease Pathology and Dysbiosis
6.1. UA Diseases and Dysbiosis
CRS, defined as inflammation within the UA sinuses for more than a period of 12 weeks, is well studied in relation to the UA microbiota [96]. In CRS patients, there was a decrease in UA microbial diversity, and their nasal microbiota was frequently dominated by coagulase-negative bacteria (Table 3) [45,97,98]. Increased prevalence of anaerobic pockets in UA niches of CRS patients may explain the decline in microbial diversity and increased presence of anaerobic bacteria [98,99,100]. Streptococcic-dominated UAs evoked proinflammatory, T-helper 1 (TH1) responses and encoded an ansamycin biosynthesis gene pathway [101]. Similarly, increased colonization of Pseudomonadaceae evoked a TH1 proinflammatory response and activated tryptophan metabolism gene pathways instead [101]. The development of nasal polyps was correlated with increased colonization of Corynebacteriaceae and enhanced IL-15 expression [101].
Additionally, the nasal microbiota potentially plays a vital role in regulating the local immune response, and consequently the pathophysiology of allergic rhinitis (AR) [99]. Samples from the middle meatus and nasal vestibule showed varying microbial profiles in AR patients vs. healthy controls [117]. Spring pollen leads to eosinophilia and increased AR symptoms, and is associated with higher middle meatus microbial biodiversity [99].
Genetic disorders such as CF and primary ciliary dyskinesia (CD) also impact the UA microbiota, as they affect an individual’s mucociliary clearance [118,119]. Children with CF present with thickening of the UA mucus layer and increased Staphylococcus spp., while other beneficial bacteria in their microbiota decrease in abundance [37,119]. CD is a combination of disorders that result in deranged ciliary action and decreased ability to clear mucus from the UA. People presenting with CD frequently suffer from CRS and are at increased risk of UAI and asthma development [118]. Equally, CF patients frequently develop CRS with the presence of nasal polyps [119].
Recently studies have proved that the nasal microbiota has a role in the onset and severity of asthma [33]. Distinct nasopharyngeal microbiota predicts the risk and severity of asthma-related inflammation [33]. During the first year of life, increased nasopharyngeal colonization of Streptococcus spp. is a strong predictor of future asthma development [33], and the nasopharyngeal microbiota in children with asthma differs from healthy controls (Table 3) [120]. Adults with exacerbated and non-exacerbated asthma have distinguished nasal microbiota [104,105]. Taxa from Bacteroidetes and Proteobacteria dominated the nasal microbiota of asthma patients (Table 3) [117,121]. Bacterial species abundance and glycerolipid metabolism varied depending on asthma state (Table 3) [103,104]. Similarly, individuals suffering from obstructive sleep apnea (OSA) have a distinct nasal microbiota [62]. Neutrophilic samples were enriched with Planococcaceae, Lactococcus, and Staphylococcus, while samples that had a high lymphocyte count or higher levels of IL-8 were enriched with Streptococcus, Rothia, and Enterococcus. Such alterations were also not impacted by three months of treatment of continuous positive airway pressure. Hence, the microbial diversity and composition distinctions in OSA patients correlated with inflammatory biomarkers.
6.2. UAIs and Dysbiosis
Different UAIs correlate with specific microbial profiles (Table 3). Positive and negative changes to the microbiota may be attributed to intramicrobial interaction and the immune response against a pathogen [122,123,124]. Influenza A virus infections modify the UA microbiota by increasing the presence of pathogenic bacteria [123,124]. Influenza virus infections in adults are associated with increased nasal carriage of Streptococcus pneumoniae and S. aureus [106]. S. pneumoniae secrete proteases that stimulate viral hemagglutinin activation and may also modulate the host immune response to facilitate influenza A viral infection [125,126]. Elevated S. pneumoniae density in the UAs increases the risk of pneumococcal pneumoniae and is associated with influenza virus, rhinovirus, and adenovirus infections [109]. In infants, a nasopharyngeal microbiota dominated by Haemophilus presented higher rhinovirus-A spp. infections [110]. Rhinovirus-C spp. infections are more likely to occur in infants with a Moraxella-dominated nasopharynx [110]. Moreover, a predominant Haemophilus nasopharyngeal microbiota is associated with delayed clearance of respiratory syncytial virus in infants hospitalized for bronchitis [127]. Equally, in health infants with human rhinovirus (HRV) infections, a study found a significant difference in the Shannon diversity index between rare and frequently infected HRV groups, and higher bacteria density in the frequently infected group [128].
7. UA Microbiota Therapeutics
Unsurprisingly, multiple therapeutics impact the UA microbiota, such as saline rinses, intranasal corticosteroids, probiotics, and antibiotics. Treatments such as antibiotics directly impact the microbiota with their antimicrobial properties and lead to a decline in UA microbial diversity and biomass [129,130]. The administration of antibiotics such as beta-lactams and mupirocin are associated with increases in Gram-negative bacteria [33,129,131,132]. Additionally, some commensal bacteria can tolerate such conditions and become pathogenic following antibiotic use [133,134]. This shift in the UA microbiota due to antibiotic treatment is assumed to last for a minimum of two weeks post-treatment, according to samples from the anterior nares [134]. Intranasal corticosteroids such as mometasone furoate monohydrate also decrease the biomass and biodiversity of the UA microbiota [129,130,135]. Intranasal administration of corticosteroids correlates with decreases in the overall UA microbial biomass by suppressing several taxa (i.e., Moraxella spp., streptococci) whilst promoting others, such as Staphylococci [129,130,135]. Intranasal vaccinations also play a role in disturbing the UA microbiota. Specifically, vaccination against S. pneumoniae alters the carriage of S. aureus and H. influenzae, yet S. pneumoniae colonization remains unchanged [136]. Intranasal live attenuated influenza vaccine significantly increased the nasal microbial taxa richness and increased the variation in influenza-specific IgA production [137]. Nasal rinses are used today to prevent and treat UAIs, and are typically a saline mixture with variable pH, but could also be as simple as distilled or tap water [138]. The inexpensive, simple method has a very low direct impact on the UA microbiota [138]. However, in some cases, contamination due to the use of tap or well water has been shown to lead to mycobacterial infections [138,139,140,141]. Surgical procedures in the UAs used to treat refractory sinusitis and polyposis also impact the microbiota [138]. The procedure enlarges the sinus ostia, improves mucociliary clearance, and facilitates topical therapy access [142]. It influences UA physiology by reducing the ambient temperature and humidity within the airways [142,143]. Patients are prescribed antibiotic and/or probiotic adjuvant therapeutics to prevent pathologic recolonization within the UAs, and the procedural outcomes are mostly positive [131,143].
Probiotics are a promising therapeutic to dysbiosis and are used in many diseases, such as asthma and CRS [144]. Probiotics are living beneficial bacteria that are administered directly into the nasal cavity to provide health benefits to the host [141,144,145]. However, the efficacy of probiotics is dependent on their ability to colonize specific regions of the UA epithelium [146]. Recently, probiotics were suggested as an adjuvant therapy to antimicrobial treatments to help mitigate their negative effects on microbial diversity and biomass [144,146]. They are sometimes referred to as keystone species due to their ability to act as pioneers and help re-establish the normal UA microbiota [147]. In turn, probiotics induce adaptations that improve the epithelial barrier and its function [144,146,148,149]. Positive interactions with host immune components induce physiological improvement of the UA epithelium by regulating signaling transduction pathways [144,146,148,149]. The residing UA symbionts interact with probiotic species, increasing antimicrobial production and inhibiting pathogen overgrowth via changing pH of the niche [147,150,151]. Notably, Enterococcus faecalis administration in children with acute sinusitis reduces the frequency and duration of disease [146,152]. Probiotics possess immunomodulatory functions, including regulation of circulating lymphocytes and cytokines [153,154,155,156]. Lactobacillus rhamnosus exemplifies this by increasing Th1 cells and decreasing Th2 cells in rodent models [157,158]. Moreover, probiotics modulate inflammatory reactions against specific immunogens by regulating lymphocyte responses and mucosal IgA levels [159].
8. Limitations and Future Research
Most of the UA microbiota knowledge is obtained through cultivation assays and next-generation sequencing. Both short reads provide basic information about the UA microbial diversity and taxonomic composition of the UA microbiota. Overcoming said limitations requires long-read technologies such as Oxford Nanopore [160] or Pacific Bioscience technology [161]. These will permit the sequencing of the entire 16S rRNA gene and provide more accurate results in terms of species- or strain-level profiling. Shotgun metagenomics offers insight into the microbiota and its functions. This technology also allows the assembly of draft uncultured microbial genomes associated with human health or disease. Sampling of the disease-afflicted region improves study findings. Positively correlated microbial candidates with both the site and incidence of a disease may help relieve its burden if removed. In opposition, negatively correlated disease candidates may play a protective role and may be administered as probiotics. Hence, it is crucial to address the limitations of sampling from less-accessible cavities of the UAs; appropriate sampling tools will minimize contamination from neighboring sites. Intramicrobial and microbial-host interactions are also research areas that require more attention. Since they will benefit our research designs in the pursuit of therapeutics and causal connection with UA disorders. Nonetheless, proving causality and displaying the efficacy and feasibility of treatments requires approaches such as animal model development and arranging more clinical trials. Finally, we have focused mostly on bacterial changes in the UA microbiota, but emerging evidence has revealed the composition of the lung virome, the global viral communities present in the airways [162]. Equally, recent data suggests that the microbiota compositions in critically severe COVID-19 patients were likely due to intubation and mechanical ventilation [111]. A recent study in children presenting with a positive SARS-CoV-2 infection demonstrated that both the upper respiratory tract and the gut microbiota were altered. The alteration of the microbiota in these children was dominated by the genus Pseudomonas (see Table 3), and remained altered up to 25–58 days in different individuals [112]. As children do not experience the complications associated with adult COVID-19, these microbiota profiles may give important insight into the role of the microbiota in disease susceptibility. This same group also demonstrated that bacterial diversity was lower in adult COVID-19 patients than healthy controls [113]. Importantly, microbiota diversity was greater enriched in mild COVID-19 patients, which suggests that a more complex microbiota may aid in recovery from COVID-19. A recent study in the USA also observed significant microbiota changes in an adult COVID-19 population [114]. Several UA immune responses are linked to COVID-19 outcomes, including changes in human epididymis secretory protein 4 [163]. However, further analysis of COVID-19′s influence on the UA microbiota and the influence of the UA microbiota on COVID-19 severity need further addressing. For further reading on microbiota and immune responses in COVID-19 upper airway, we recommend the review by Di Stadio et al. [164].
9. Conclusions
Overall, the UA microbiota literature suggests that the development of a diverse microbiota across the UAs aids in maintaining healthy and disease-free UAs. Age, diet, environmental exposures, infections, and therapeutics all contribute to UA microbiota and health. The loss of UA community segregation appears to contribute to inflammation and disease that results in disadvantageous microbial–microbial and microbial–host interactions. Maintaining UA microbiota homeostasis may represent a major approach for future therapeutics. New approaches to sampling and analysing microbiota diversity will aid in our understanding of UA diseases. Future scientific advancements to recover the healthy microbiota may become a common choice in the treatment of patients with UA diseases.
Author Contributions
Conceptualization, Z.E. and P.G.; writing—original draft preparation, Z.E.; writing—review and editing, Z.E., P.S. and P.G.; visualization, Z.E.; supervision, P.G.; project administration, P.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant made available to P.G. from the Flight Attendant Medical Research Institute (No. CIA160005).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Neil MacLusky for his invaluable input.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sahin-Yilmaz, A.; Naclerio, R.M. Anatomy and physiology of the upper airway. Proc. Am. Thorac. Soc. 2011, 8, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Kumpitsch, C.; Koskinen, K.; Schopf, V.; Moissl-Eichinger, C. The microbiome of the upper respiratory tract in health and disease. BMC Biol. 2019, 17, 87. [Google Scholar] [CrossRef] [Green Version]
- Hartl, D.; Tirouvanziam, R.; Laval, J.; Greene, C.M.; Habiel, D.; Sharma, L.; Yildirim, A.O.; Dela Cruz, C.S.; Hogaboam, C.M. Innate Immunity of the Lung: From Basic Mechanisms to Translational Medicine. J. Innate Immun. 2018, 10, 487–501. [Google Scholar] [CrossRef]
- Gandhi, V.D.; Vliagoftis, H. Airway epithelium interactions with aeroallergens: Role of secreted cytokines and chemokines in innate immunity. Front. Immunol. 2015, 6, 147. [Google Scholar] [CrossRef] [Green Version]
- de Steenhuijsen Piters, W.A.; Sanders, E.A.; Bogaert, D. The role of the local microbial ecosystem in respiratory health and disease. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proctor, D.M.; Relman, D.A. The Landscape Ecology and Microbiota of the Human Nose, Mouth, and Throat. Cell Host Microbe 2017, 21, 421–432. [Google Scholar] [CrossRef] [Green Version]
- Lloyd-Price, J.; Abu-Ali, G.; Huttenhower, C. The healthy human microbiome. Genome Med. 2016, 8, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costello, E.K.; Lauber, C.L.; Hamady, M.; Fierer, N.; Gordon, J.I.; Knight, R. Bacterial community variation in human body habitats across space and time. Science 2009, 326, 1694–1697. [Google Scholar] [CrossRef] [Green Version]
- Schenck, L.P.; Surette, M.G.; Bowdish, D.M. Composition and immunological significance of the upper respiratory tract microbiota. FEBS Lett. 2016, 590, 3705–3720. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Mahurkar, A.; Rahnavard, G.; Crabtree, J.; Orvis, J.; Hall, A.B.; Brady, A.; Creasy, H.H.; McCracken, C.; Giglio, M.G.; et al. Strains, functions and dynamics in the expanded Human Microbiome Project. Nat. Cell Biol. 2017, 550, 61–66. [Google Scholar] [CrossRef]
- Garibaldi, R.A. Epidemiology of community-acquired respiratory tract infections in adults. Incidence, etiology, and impact. Am. J. Med. 1985, 78, 32–37. [Google Scholar] [CrossRef]
- Ballini, A.; Scacco, S.; Boccellino, M.; Santacroce, L.; Arrigoni, R. Microbiota and Obesity: Where Are We Now? Biology 2020, 9, 415. [Google Scholar] [CrossRef]
- De Boeck, I.; Wittouck, S.; Wuyts, S.; Oerlemans, E.F.M.; van den Broek, M.F.L.; Vandenheuvel, D.; Vanderveken, O.; Lebeer, S. Comparing the Healthy Nose and Nasopharynx Microbiota Reveals Continuity As Well As Niche-Specificity. Front. Microbiol. 2017, 8, 2372. [Google Scholar] [CrossRef] [Green Version]
- Man, W.H.; de Steenhuijsen Piters, W.A.; Bogaert, D. The microbiota of the respiratory tract: Gatekeeper to respiratory health. Nat. Rev. Microbiol. 2017, 15, 259–270. [Google Scholar] [CrossRef]
- Frayman, K.B.; Armstrong, D.S.; Grimwood, K.; Ranganathan, S.C. The airway microbiota in early cystic fibrosis lung disease. Pediatr. Pulmonol. 2017, 52, 1384–1404. [Google Scholar] [CrossRef] [PubMed]
- Wostmann, B.S. The germfree animal in nutritional studies. Annu. Rev. Nutr. 1981, 1, 257–279. [Google Scholar] [CrossRef] [PubMed]
- Harabuchi, Y.; Hamamoto, M.; Shirasaki, H.; Asakura, K.; Matsuyama, H.; Kataura, A. Specific immune response of the adenoids to a respiratory antigen. Am. J. Otolaryngol. 1989, 10, 138–142. [Google Scholar] [CrossRef]
- Jones, N. The nose and paranasal sinuses physiology and anatomy. Adv. Drug Deliv. Rev. 2001, 51, 5–19. [Google Scholar] [CrossRef]
- Stearns, J.C.; Davidson, C.J.; McKeon, S.; Whelan, F.J.; Fontes, M.E.; Schryvers, A.B.; Bowdish, D.M.; Kellner, J.D.; Surette, M.G. Culture and molecular-based profiles show shifts in bacterial communities of the upper respiratory tract that occur with age. ISME J. 2015, 9, 1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassis, C.M.; Tang, A.L.; Young, V.B.; Pynnonen, M.A. The nasal cavity microbiota of healthy adults. Microbiome 2014, 2, 27. [Google Scholar] [CrossRef] [Green Version]
- Pang, B.; Yee, K.K.; Lischka, F.W.; Rawson, N.E.; Haskins, M.E.; Wysocki, C.J.; Craven, B.A.; Van Valkenburgh, B. The influence of nasal airflow on respiratory and olfactory epithelial distribution in felids. J. Exp. Biol. 2016, 219, 1866–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camarinha-Silva, A.; Jauregui, R.; Pieper, D.H.; Wos-Oxley, M.L. The temporal dynamics of bacterial communities across human anterior nares. Environ. Microbiol. Rep. 2012, 4, 126–132. [Google Scholar] [CrossRef]
- Geurkink, N. Nasal anatomy, physiology, and function. J. Allergy Clin. Immunol. 1983, 72, 123–128. [Google Scholar] [CrossRef]
- Koskinen, K.; Reichert, J.L.; Hoier, S.; Schachenreiter, J.; Duller, S.; Moissl-Eichinger, C.; Schopf, V. The nasal microbiome mirrors and potentially shapes olfactory function. Sci. Rep. 2018, 8, 1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, M. Microbial Inhabitants of Humans: Their Ecology and Role in Health and Disease; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Wang, H.; Dai, W.; Feng, X.; Zhou, Q.; Wang, H.; Yang, Y.; Li, S.; Zheng, Y. Microbiota Composition in Upper Respiratory Tracts of Healthy Children in Shenzhen, China, Differed with Respiratory Sites and Ages. Biomed. Res. Int. 2018, 2018, 6515670. [Google Scholar] [CrossRef] [PubMed]
- Ubags, N.D.J.; Marsland, B.J. Mechanistic insight into the function of the microbiome in lung diseases. Eur. Respir. J. 2017, 50. [Google Scholar] [CrossRef]
- Whelan, F.J.; Verschoor, C.P.; Stearns, J.C.; Rossi, L.; Luinstra, K.; Loeb, M.; Smieja, M.; Johnstone, J.; Surette, M.G.; Bowdish, D.M. The loss of topography in the microbial communities of the upper respiratory tract in the elderly. Ann. Am. Thorac. Soc. 2014, 11, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Wos-Oxley, M.L.; Plumeier, I.; von Eiff, C.; Taudien, S.; Platzer, M.; Vilchez-Vargas, R.; Becker, K.; Pieper, D.H. A poke into the diversity and associations within human anterior nare microbial communities. ISME J. 2010, 4, 839–851. [Google Scholar] [CrossRef]
- Charlson, E.S.; Chen, J.; Custers-Allen, R.; Bittinger, K.; Li, H.; Sinha, R.; Hwang, J.; Bushman, F.D.; Collman, R.G. Disordered microbial communities in the upper respiratory tract of cigarette smokers. PLoS ONE 2010, 5, e15216. [Google Scholar] [CrossRef] [Green Version]
- Yan, M.; Pamp, S.J.; Fukuyama, J.; Hwang, P.H.; Cho, D.Y.; Holmes, S.; Relman, D.A. Nasal microenvironments and interspecific interactions influence nasal microbiota complexity and S. aureus carriage. Cell Host Microbe 2013, 14, 631–640. [Google Scholar] [CrossRef] [Green Version]
- Castranova, V.; Huffman, L.J.; Judy, D.J.; Bylander, J.E.; Lapp, L.N.; Weber, S.L.; Blackford, J.A.; Dey, R.D. Enhancement of nitric oxide production by pulmonary cells following silica exposure. Environ. Health Perspect 1998, 106 (Suppl. 5), 1165–1169. [Google Scholar] [CrossRef] [PubMed]
- Teo, S.M.; Mok, D.; Pham, K.; Kusel, M.; Serralha, M.; Troy, N.; Holt, B.J.; Hales, B.J.; Walker, M.L.; Hollams, E.; et al. The infant nasopharyngeal microbiome impacts severity of lower respiratory infection and risk of asthma development. Cell Host Microbe 2015, 17, 704–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Linstow, M.L.; Schonning, K.; Hoegh, A.M.; Sevelsted, A.; Vissing, N.H.; Bisgaard, H. Neonatal airway colonization is associated with troublesome lung symptoms in infants. Am. J. Respir. Crit. Care Med. 2013, 188, 1041–1042. [Google Scholar] [CrossRef]
- Biesbroek, G.; Tsivtsivadze, E.; Sanders, E.A.; Montijn, R.; Veenhoven, R.H.; Keijser, B.J.; Bogaert, D. Early respiratory microbiota composition determines bacterial succession patterns and respiratory health in children. Am. J. Respir. Crit. Care Med. 2014, 190, 1283–1292. [Google Scholar] [CrossRef]
- Biesbroek, G.; Bosch, A.A.; Wang, X.; Keijser, B.J.; Veenhoven, R.H.; Sanders, E.A.; Bogaert, D. The impact of breastfeeding on nasopharyngeal microbial communities in infants. Am. J. Respir. Crit. Care Med. 2014, 190, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Prevaes, S.M.; de Winter-de Groot, K.M.; Janssens, H.M.; de Steenhuijsen Piters, W.A.; Tramper-Stranders, G.A.; Wyllie, A.L.; Hasrat, R.; Tiddens, H.A.; van Westreenen, M.; van der Ent, C.K.; et al. Development of the Nasopharyngeal Microbiota in Infants with Cystic Fibrosis. Am. J. Respir. Crit. Care Med. 2016, 193, 504–515. [Google Scholar] [CrossRef]
- Luna, P.N.; Hasegawa, K.; Ajami, N.J.; Espinola, J.A.; Henke, D.M.; Petrosino, J.F.; Piedra, P.A.; Sullivan, A.F.; Camargo, C.A., Jr.; Shaw, C.A.; et al. The association between anterior nares and nasopharyngeal microbiota in infants hospitalized for bronchiolitis. Microbiome 2018, 6, 2. [Google Scholar] [CrossRef]
- Vissing, N.H.; Chawes, B.L.; Bisgaard, H. Increased risk of pneumonia and bronchiolitis after bacterial colonization of the airways as neonates. Am. J. Respir. Crit. Care Med. 2013, 188, 1246–1252. [Google Scholar] [CrossRef]
- Camarinha-Silva, A.; Jauregui, R.; Chaves-Moreno, D.; Oxley, A.P.; Schaumburg, F.; Becker, K.; Wos-Oxley, M.L.; Pieper, D.H. Comparing the anterior nare bacterial community of two discrete human populations using Illumina amplicon sequencing. Environ. Microbiol. 2014, 16, 2939–2952. [Google Scholar] [CrossRef]
- de Steenhuijsen Piters, W.A.; Huijskens, E.G.; Wyllie, A.L.; Biesbroek, G.; van den Bergh, M.R.; Veenhoven, R.H.; Wang, X.; Trzcinski, K.; Bonten, M.J.; Rossen, J.W.; et al. Dysbiosis of upper respiratory tract microbiota in elderly pneumonia patients. ISME J. 2016, 10, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Mihindukulasuriya, K.A.; Gao, H.; La Rosa, P.S.; Wylie, K.M.; Martin, J.C.; Kota, K.; Shannon, W.D.; Mitreva, M.; Sodergren, E.; et al. Exploration of bacterial community classes in major human habitats. Genome Biol. 2014, 15, R66. [Google Scholar] [CrossRef]
- Zhou, Y.; Gao, H.; Mihindukulasuriya, K.A.; La Rosa, P.S.; Wylie, K.M.; Vishnivetskaya, T.; Podar, M.; Warner, B.; Tarr, P.I.; Nelson, D.E.; et al. Biogeography of the ecosystems of the healthy human body. Genome Biol. 2013, 14, R1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, J.; Byrd, A.L.; Deming, C.; Conlan, S.; Program, N.C.S.; Kong, H.H.; Segre, J.A. Biogeography and individuality shape function in the human skin metagenome. Nature 2014, 514, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, J.M.; Amedee, R.G. Contrasting the microbiomes from healthy volunteers and patients with chronic rhinosinusitis. Am. J. Rhinol. Allergy 2014, 28, 182. [Google Scholar] [CrossRef] [PubMed]
- Dickson, R.P.; Erb-Downward, J.R.; Freeman, C.M.; McCloskey, L.; Beck, J.M.; Huffnagle, G.B.; Curtis, J.L. Spatial Variation in the Healthy Human Lung Microbiome and the Adapted Island Model of Lung Biogeography. Ann. Am. Thorac. Soc. 2015, 12, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Capri, M.; Monti, D.; Giunta, S.; Olivieri, F.; Sevini, F.; Panourgia, M.P.; Invidia, L.; Celani, L.; Scurti, M.; et al. Inflammaging and anti-inflammaging: A systemic perspective on aging and longevity emerged from studies in humans. Mech. Ageing Dev. 2007, 128, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Morris, B.E.; Henneberger, R.; Huber, H.; Moissl-Eichinger, C. Microbial syntrophy: Interaction for the common good. FEMS Microbiol. Rev. 2013, 37, 384–406. [Google Scholar] [CrossRef]
- Ramsey, M.M.; Freire, M.O.; Gabrilska, R.A.; Rumbaugh, K.P.; Lemon, K.P. Staphylococcus aureus Shifts toward Commensalism in Response to Corynebacterium Species. Front. Microbiol. 2016, 7, 1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stubbendieck, R.M.; May, D.S.; Chevrette, M.G.; Temkin, M.I.; Wendt-Pienkowski, E.; Cagnazzo, J.; Carlson, C.M.; Gern, J.E.; Currie, C.R. Competition among Nasal Bacteria Suggests a Role for Siderophore-Mediated Interactions in Shaping the Human Nasal Microbiota. Appl. Environ. Microbiol. 2019, 85, e02406-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bomar, L.; Brugger, S.D.; Yost, B.H.; Davies, S.S.; Lemon, K.P. Corynebacterium accolens Releases Antipneumococcal Free Fatty Acids from Human Nostril and Skin Surface Triacylglycerols. mBio 2016, 7, e01725-15. [Google Scholar] [CrossRef] [Green Version]
- Wollenberg, M.S.; Claesen, J.; Escapa, I.F.; Aldridge, K.L.; Fischbach, M.A.; Lemon, K.P. Propionibacterium-produced coproporphyrin III induces Staphylococcus aureus aggregation and biofilm formation. mBio 2014, 5, e01286-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zipperer, A.; Konnerth, M.C.; Laux, C.; Berscheid, A.; Janek, D.; Weidenmaier, C.; Burian, M.; Schilling, N.A.; Slavetinsky, C.; Marschal, M.; et al. Human commensals producing a novel antibiotic impair pathogen colonization. Nature 2016, 535, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, J.A.; Riley, T.V.; Mee, B.J. Staphylococcus aureus but not Staphylococcus epidermidis can acquire iron from transferrin. Microbiology (Read.) 1995, 141 Pt 1, 197–203. [Google Scholar] [CrossRef] [Green Version]
- Krismer, B.; Liebeke, M.; Janek, D.; Nega, M.; Rautenberg, M.; Hornig, G.; Unger, C.; Weidenmaier, C.; Lalk, M.; Peschel, A. Nutrient limitation governs Staphylococcus aureus metabolism and niche adaptation in the human nose. PLoS Pathog. 2014, 10, e1003862. [Google Scholar] [CrossRef]
- Schaller, M.; Loewenstein, M.; Borelli, C.; Jacob, K.; Vogeser, M.; Burgdorf, W.H.; Plewig, G. Induction of a chemoattractive proinflammatory cytokine response after stimulation of keratinocytes with Propionibacterium acnes and coproporphyrin III. Br. J. Dermatol. 2005, 153, 66–71. [Google Scholar] [CrossRef]
- Borelli, C.; Merk, K.; Schaller, M.; Jacob, K.; Vogeser, M.; Weindl, G.; Berger, U.; Plewig, G. In vivo porphyrin production by P. acnes in untreated acne patients and its modulation by acne treatment. Acta Derm. Venereol. 2006, 86, 316–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, G.L.; Holt, P.G. The concept of airway inflammation. Am. J. Respir. Crit. Care Med. 2000, 162, S2–S6. [Google Scholar] [CrossRef]
- Sorensen, O.E.; Borregaard, N. Neutrophil extracellular traps—The dark side of neutrophils. J. Clin. Investig. 2016, 126, 1612–1620. [Google Scholar] [CrossRef] [Green Version]
- Jhelum, H.; Sori, H.; Sehgal, D. A novel extracellular vesicle-associated endodeoxyribonuclease helps Streptococcus pneumoniae evade neutrophil extracellular traps and is required for full virulence. Sci. Rep. 2018, 8, 7985. [Google Scholar] [CrossRef]
- Ritchie, N.D.; Ijaz, U.Z.; Evans, T.J. IL-17 signalling restructures the nasal microbiome and drives dynamic changes following Streptococcus pneumoniae colonization. BMC Genom. 2017, 18, 807. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.G.; Sulaiman, I.; Wang, J.; Shen, N.; Clemente, J.C.; Li, Y.; Laumbach, R.J.; Lu, S.E.; Udasin, I.; Le-Hoang, O.; et al. Severe Obstructive Sleep Apnea Is Associated with Alterations in the Nasal Microbiome and an Increase in Inflammation. Am. J. Respir. Crit. Care Med. 2019, 199, 99–109. [Google Scholar] [CrossRef]
- Kirst, M.E.; Baker, D.; Li, E.; Abu-Hasan, M.; Wang, G.P. Upper versus lower airway microbiome and metagenome in children with cystic fibrosis and their correlation with lung inflammation. PLoS ONE 2019, 14, e0222323. [Google Scholar] [CrossRef] [PubMed]
- Psaltis, A.J.; Wormald, P.J. Therapy of Sinonasal Microbiome in CRS: A Critical Approach. Curr. Allergy Asthma Rep. 2017, 17, 59. [Google Scholar] [CrossRef] [PubMed]
- Janhsen, W.K.; Arnold, C.; Hentschel, J.; Lehmann, T.; Pfister, W.; Baier, M.; Boer, K.; Hunniger, K.; Kurzai, O.; Hipler, U.C.; et al. Colonization of CF patients’ upper airways with S. aureus contributes more decisively to upper airway inflammation than P. aeruginosa. Med. Microbiol. Immunol. 2016, 205, 485–500. [Google Scholar] [CrossRef]
- Schwartz, J.S.; Peres, A.G.; Mfuna Endam, L.; Cousineau, B.; Madrenas, J.; Desrosiers, M. Topical probiotics as a therapeutic alternative for chronic rhinosinusitis: A preclinical proof of concept. Am. J. Rhinol. Allergy 2016, 30, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Wagner Mackenzie, B.; Waite, D.W.; Hoggard, M.; Douglas, R.G.; Taylor, M.W.; Biswas, K. Bacterial community collapse: A meta-analysis of the sinonasal microbiota in chronic rhinosinusitis. Environ. Microbiol. 2017, 19, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Manisalidis, I.; Stavropoulou, E.; Stavropoulos, A.; Bezirtzoglou, E. Environmental and Health Impacts of Air Pollution: A Review. Front. Public Health 2020, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murdoch, K.M.; Mitra, B.; Lambert, S.; Erbas, B. What is the seasonal distribution of community acquired pneumonia over time? A systematic review. Australas Emerg. Nurs. J. 2014, 17, 30–42. [Google Scholar] [CrossRef] [Green Version]
- Numminen, E.; Chewapreecha, C.; Turner, C.; Goldblatt, D.; Nosten, F.; Bentley, S.D.; Turner, P.; Corander, J. Climate induces seasonality in pneumococcal transmission. Sci. Rep. 2015, 5, 11344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinberger, D.M.; Grant, L.R.; Steiner, C.A.; Weatherholtz, R.; Santosham, M.; Viboud, C.; O’Brien, K.L. Seasonal drivers of pneumococcal disease incidence: Impact of bacterial carriage and viral activity. Clin. Infect. Dis. 2014, 58, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Hassoun, A.; Huff, M.D.; Weisman, D.; Chahal, K.; Asis, E.; Stalons, D.; Grigorenko, E.; Green, J.; Malone, L.L.; Clemmons, S.; et al. Seasonal variation of respiratory pathogen colonization in asymptomatic health care professionals: A single-center, cross-sectional, 2-season observational study. Am. J. Infect. Control. 2015, 43, 865–870. [Google Scholar] [CrossRef] [Green Version]
- Bogaert, D.; Keijser, B.; Huse, S.; Rossen, J.; Veenhoven, R.; van Gils, E.; Bruin, J.; Montijn, R.; Bonten, M.; Sanders, E. Variability and diversity of nasopharyngeal microbiota in children: A metagenomic analysis. PLoS ONE 2011, 6, e17035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, F.J.; Fussell, J.C. Air pollution and airway disease. Clin. Exp. Allergy 2011, 41, 1059–1071. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Chu, J.; Li, Y.; Kong, X. The influence of air pollution on respiratory microbiome: A link to respiratory disease. Toxicol. Lett. 2020, 334, 14–20. [Google Scholar] [CrossRef]
- Guarnieri, M.; Balmes, J.R. Outdoor air pollution and asthma. Lancet 2014, 383, 1581–1592. [Google Scholar] [CrossRef] [Green Version]
- Carter, E.; Norris, C.; Dionisio, K.L.; Balakrishnan, K.; Checkley, W.; Clark, M.L.; Ghosh, S.; Jack, D.W.; Kinney, P.L.; Marshall, J.D.; et al. Assessing Exposure to Household Air Pollution: A Systematic Review and Pooled Analysis of Carbon Monoxide as a Surrogate Measure of Particulate Matter. Environ. Health Perspect. 2017, 125, 076002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simkovich, S.M.; Goodman, D.; Roa, C.; Crocker, M.E.; Gianella, G.E.; Kirenga, B.J.; Wise, R.A.; Checkley, W. The health and social implications of household air pollution and respiratory diseases. NPJ Prim. Care Respir. Med. 2019, 29, 12. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.J.; Zheng, X.Y.; Chung, K.F.; Zhong, N.S. Impact of air pollution on the burden of chronic respiratory diseases in China: Time for urgent action. Lancet 2016, 388, 1939–1951. [Google Scholar] [CrossRef]
- Whyand, T.; Hurst, J.R.; Beckles, M.; Caplin, M.E. Pollution and respiratory disease: Can diet or supplements help? A review. Respir. Res. 2018, 19, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariani, J.; Favero, C.; Spinazze, A.; Cavallo, D.M.; Carugno, M.; Motta, V.; Bonzini, M.; Cattaneo, A.; Pesatori, A.C.; Bollati, V. Short-term particulate matter exposure influences nasal microbiota in a population of healthy subjects. Environ. Res. 2018, 162, 119–126. [Google Scholar] [CrossRef]
- Qin, T.; Zhang, F.; Zhou, H.; Ren, H.; Du, Y.; Liang, S.; Wang, F.; Cheng, L.; Xie, X.; Jin, A.; et al. High-Level PM2.5/PM10 Exposure Is Associated With Alterations in the Human Pharyngeal Microbiota Composition. Front. Microbiol. 2019, 10, 54. [Google Scholar] [CrossRef] [Green Version]
- Gisler, A.; Korten, I.; de Hoogh, K.; Vienneau, D.; Frey, U.; Decrue, F.; Gorlanova, O.; Soti, A.; Hilty, M.; Latzin, P.; et al. Associations of air pollution and greenness with the nasal microbiota of healthy infants: A longitudinal study. Environ. Res. 2021, 202, 111633. [Google Scholar] [CrossRef]
- Brook, I.; Gober, A.E. Recovery of potential pathogens in the nasopharynx of healthy and otitis media-prone children and their smoking and nonsmoking parents. Ann. Otol. Rhinol. Laryngol. 2008, 117, 727–730. [Google Scholar] [CrossRef]
- Greenberg, D.; Givon-Lavi, N.; Broides, A.; Blancovich, I.; Peled, N.; Dagan, R. The contribution of smoking and exposure to tobacco smoke to Streptococcus pneumoniae and Haemophilus influenzae carriage in children and their mothers. Clin. Infect. Dis. 2006, 42, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, A.R.; Berger, S.; Vogel, T.M. Human pathogens abundant in the bacterial metagenome of cigarettes. Environ. Health Perspect. 2010, 118, 351–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Sun, Y.; An, Y.; Wang, R.; Lin, H.; Liu, M.; Li, S.; Ma, M.; Xiao, C. Air pollution during the winter period and respiratory tract microbial imbalance in a healthy young population in Northeastern China. Environ. Pollut. 2019, 246, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Chen, R.; Wang, C.; Wang, W.; Jiang, J.; Wu, W.; Cai, J.; Zhao, Z.; Xu, X.; Kan, H. Ozone exposure leads to changes in airway permeability, microbiota and metabolome: A randomised, double-blind, crossover trial. Eur. Respir. J. 2020, 56. [Google Scholar] [CrossRef] [PubMed]
- Rylance, J.; Kankwatira, A.; Nelson, D.E.; Toh, E.; Day, R.B.; Lin, H.; Gao, X.; Dong, Q.; Sodergren, E.; Weinstock, G.M.; et al. Household air pollution and the lung microbiome of healthy adults in Malawi: A cross-sectional study. BMC Microbiol. 2016, 16, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garmendia, J.; Morey, P.; Bengoechea, J.A. Impact of cigarette smoke exposure on host-bacterial pathogen interactions. Eur. Respir. J. 2012, 39, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Phipps, J.C.; Aronoff, D.M.; Curtis, J.L.; Goel, D.; O’Brien, E.; Mancuso, P. Cigarette smoke exposure impairs pulmonary bacterial clearance and alveolar macrophage complement-mediated phagocytosis of Streptococcus pneumoniae. Infect. Immun. 2010, 78, 1214–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagaitkar, J.; Demuth, D.R.; Daep, C.A.; Renaud, D.E.; Pierce, D.L.; Scott, D.A. Tobacco upregulates P. gingivalis fimbrial proteins which induce TLR2 hyposensitivity. PLoS ONE 2010, 5, e9323. [Google Scholar] [CrossRef] [PubMed]
- Goldstein-Daruech, N.; Cope, E.K.; Zhao, K.Q.; Vukovic, K.; Kofonow, J.M.; Doghramji, L.; Gonzalez, B.; Chiu, A.G.; Kennedy, D.W.; Palmer, J.N.; et al. Tobacco smoke mediated induction of sinonasal microbial biofilms. PLoS ONE 2011, 6, e15700. [Google Scholar] [CrossRef] [Green Version]
- Ratner, A.J.; Lysenko, E.S.; Paul, M.N.; Weiser, J.N. Synergistic proinflammatory responses induced by polymicrobial colonization of epithelial surfaces. Proc. Natl. Acad. Sci. USA 2005, 102, 3429–3434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, S.J. Smoking cessation may improve the microbial flora profile. Thorax 2007, 62, 676. [Google Scholar]
- Abreu, N.A.; Nagalingam, N.A.; Song, Y.; Roediger, F.C.; Pletcher, S.D.; Goldberg, A.N.; Lynch, S.V. Sinus microbiome diversity depletion and Corynebacterium tuberculostearicum enrichment mediates rhinosinusitis. Sci. Transl. Med. 2012, 4, 151ra124. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.B.; Hong, S.W.; Kim, D.K.; Jeon, S.G.; Kim, K.R.; Cho, S.H.; Gho, Y.S.; Jee, Y.K.; Kim, Y.K. Decreased diversity of nasal microbiota and their secreted extracellular vesicles in patients with chronic rhinosinusitis based on a metagenomic analysis. Allergy 2014, 69, 517–526. [Google Scholar] [CrossRef]
- Ramakrishnan, V.R.; Hauser, L.J.; Feazel, L.M.; Ir, D.; Robertson, C.E.; Frank, D.N. Sinus microbiota varies among chronic rhinosinusitis phenotypes and predicts surgical outcome. J. Allergy Clin. Immunol. 2015, 136, 334–342.e331. [Google Scholar] [CrossRef] [PubMed]
- Aurora, R.; Chatterjee, D.; Hentzleman, J.; Prasad, G.; Sindwani, R.; Sanford, T. Contrasting the microbiomes from healthy volunteers and patients with chronic rhinosinusitis. JAMA Otolaryngol. Head Neck Surg. 2013, 139, 1328–1338. [Google Scholar] [CrossRef]
- Lal, D.; Keim, P.; Delisle, J.; Barker, B.; Rank, M.A.; Chia, N.; Schupp, J.M.; Gillece, J.D.; Cope, E.K. Mapping and comparing bacterial microbiota in the sinonasal cavity of healthy, allergic rhinitis, and chronic rhinosinusitis subjects. Int. Forum Allergy Rhinol. 2017, 7, 561–569. [Google Scholar] [CrossRef]
- Copeland, E.; Leonard, K.; Carney, R.; Kong, J.; Forer, M.; Naidoo, Y.; Oliver, B.G.G.; Seymour, J.R.; Woodcock, S.; Burke, C.M.; et al. Chronic Rhinosinusitis: Potential Role of Microbial Dysbiosis and Recommendations for Sampling Sites. Front. Cell Infect. Microbiol. 2018, 8, 57. [Google Scholar] [CrossRef]
- Cope, E.K.; Goldberg, A.N.; Pletcher, S.D.; Lynch, S.V. Compositionally and functionally distinct sinus microbiota in chronic rhinosinusitis patients have immunological and clinically divergent consequences. Microbiome 2017, 5, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoggard, M.; Waldvogel-Thurlow, S.; Zoing, M.; Chang, K.; Radcliff, F.J.; Wagner Mackenzie, B.; Biswas, K.; Douglas, R.G.; Taylor, M.W. Inflammatory Endotypes and Microbial Associations in Chronic Rhinosinusitis. Front. Immunol. 2018, 9, 2065. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.J.; Kim, S.H.; Lee, M.J.; Kim, B.K.; Song, W.J.; Park, H.W.; Cho, S.H.; Hong, S.J.; Chang, Y.S.; Kim, B.S. Different upper airway microbiome and their functional genes associated with asthma in young adults and elderly individuals. Allergy 2019, 74, 709–719. [Google Scholar] [CrossRef]
- Fazlollahi, M.; Lee, T.D.; Andrade, J.; Oguntuyo, K.; Chun, Y.; Grishina, G.; Grishin, A.; Bunyavanich, S. The nasal microbiome in asthma. J. Allergy Clin. Immunol. 2018, 142, 834–843.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lastours, V.; Malosh, R.; Ramadugu, K.; Srinivasan, U.; Dawid, S.; Ohmit, S.; Foxman, B. Co-colonization by Streptococcus pneumoniae and Staphylococcus aureus in the throat during acute respiratory illnesses. Epidemiol. Infect. 2016, 144, 3507–3519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges, L.; Giongo, A.; Pereira, L.M.; Trindade, F.J.; Gregianini, T.S.; Campos, F.S.; Ghedin, E.; da Veiga, A.B.G. Comparison of the nasopharynx microbiome between influenza and non-influenza cases of severe acute respiratory infections: A pilot study. Health Sci. Rep. 2018, 1, e47. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.; Xie, G.; Zhou, Q.; Qiu, C.; Li, J.; Hu, Q.; Dai, W.; Li, D.; Zheng, Y.; Wen, F. Distinct Nasopharyngeal and Oropharyngeal Microbiota of Children with Influenza A Virus Compared with Healthy Children. Biomed. Res. Int. 2018, 2018, 6362716. [Google Scholar] [CrossRef] [Green Version]
- Wolter, N.; Tempia, S.; Cohen, C.; Madhi, S.A.; Venter, M.; Moyes, J.; Walaza, S.; Malope-Kgokong, B.; Groome, M.; du Plessis, M.; et al. High nasopharyngeal pneumococcal density, increased by viral coinfection, is associated with invasive pneumococcal pneumonia. J. Infect. Dis. 2014, 210, 1649–1657. [Google Scholar] [CrossRef] [Green Version]
- Toivonen, L.; Camargo, C.A., Jr.; Gern, J.E.; Bochkov, Y.A.; Mansbach, J.M.; Piedra, P.A.; Hasegawa, K. Association between rhinovirus species and nasopharyngeal microbiota in infants with severe bronchiolitis. J. Allergy Clin. Immunol. 2019, 143, 1925–1928.e7. [Google Scholar] [CrossRef] [Green Version]
- Miao, Q.; Schwarz, S.; Schwarz, G. Responding to COVID-19: Community volunteerism and coproduction in China. World Dev. 2021, 137, 105128. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Liu, P.; Zhang, T.; Wu, Q.; Zeng, M.; Ma, Y.; Jin, X.; Xu, J.; Zhang, Z.; Zhang, C. Progressive deterioration of the upper respiratory tract and the gut microbiomes in children during the early infection stages of COVID-19. J. Genet. Genom. 2021. [Google Scholar] [CrossRef]
- Xu, R.; Lu, R.; Zhang, T.; Wu, Q.; Cai, W.; Han, X.; Wan, Z.; Jin, X.; Zhang, Z.; Zhang, C. Temporal association between human upper respiratory and gut bacterial microbiomes during the course of COVID-19 in adults. Commun. Biol. 2021, 4, 240. [Google Scholar] [CrossRef]
- Rosas-Salazar, C.; Kimura, K.S.; Shilts, M.H.; Strickland, B.A.; Freeman, M.H.; Wessinger, B.C.; Gupta, V.; Brown, H.M.; Rajagopala, S.V.; Turner, J.H.; et al. SARS-CoV-2 infection and viral load are associated with the upper respiratory tract microbiome. J. Allergy Clin. Immunol. 2021, 147, 1226–1233.e2. [Google Scholar] [CrossRef]
- Aljabr, W.; Alruwaili, M.; Penrice-Randal, R.; Alrezaihi, A.; Harrison, A.J.; Ryan, Y.; Bentley, E.; Jones, B.; Alhatlani, B.Y.; AlShahrani, D.; et al. Amplicon and Metagenomic Analysis of Middle East Respiratory Syndrome (MERS) Coronavirus and the Microbiome in Patients with Severe MERS. mSphere 2021, e0021921. [Google Scholar] [CrossRef]
- Fan, R.R.; Howard, L.M.; Griffin, M.R.; Edwards, K.M.; Zhu, Y.; Williams, J.V.; Vidal, J.E.; Klugman, K.P.; Gil, A.I.; Lanata, C.F.; et al. Nasopharyngeal Pneumococcal Density and Evolution of Acute Respiratory Illnesses in Young Children, Peru, 2009–2011. Emerg. Infect. Dis. 2016, 22, 1996–1999. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.H.; Poroyko, V.; Watanabe, S.; Jiang, D.; Lane, J.; deTineo, M.; Baroody, F.M.; Naclerio, R.M.; Pinto, J.M. Seasonal allergic rhinitis affects sinonasal microbiota. Am. J. Rhinol. Allergy 2014, 28, 281–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommer, J.U.; Schafer, K.; Omran, H.; Olbrich, H.; Wallmeier, J.; Blum, A.; Hormann, K.; Stuck, B.A. ENT manifestations in patients with primary ciliary dyskinesia: Prevalence and significance of otorhinolaryngologic co-morbidities. Eur. Arch. Otorhinolaryngol. 2011, 268, 383–388. [Google Scholar] [CrossRef]
- Schraven, S.P.; Wehrmann, M.; Wagner, W.; Blumenstock, G.; Koitschev, A. Prevalence and histopathology of chronic polypoid sinusitis in pediatric patients with cystic fibrosis. J. Cyst. Fibros. 2011, 10, 181–186. [Google Scholar] [CrossRef]
- Perez-Losada, M.; Crandall, K.A.; Freishtat, R.J. Two sampling methods yield distinct microbial signatures in the nasopharynges of asthmatic children. Microbiome 2016, 4, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finegold, S.M.; Flynn, M.J.; Rose, F.V.; Jousimies-Somer, H.; Jakielaszek, C.; McTeague, M.; Wexler, H.M.; Berkowitz, E.; Wynne, B. Bacteriologic findings associated with chronic bacterial maxillary sinusitis in adults. Clin. Infect. Dis. 2002, 35, 428–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.H.; Gordon, A.; Shedden, K.; Kuan, G.; Ng, S.; Balmaseda, A.; Foxman, B. The respiratory microbiome and susceptibility to influenza virus infection. PLoS ONE 2019, 14, e0207898. [Google Scholar] [CrossRef] [Green Version]
- Lynch, S.V. Viruses and microbiome alterations. Ann. Am. Thorac. Soc. 2014, 11 (Suppl. 1), S57–S60. [Google Scholar] [CrossRef]
- Lee, K.H.; Gordon, A.; Foxman, B. The role of respiratory viruses in the etiology of bacterial pneumonia: An ecological perspective. Evol. Med. Public Health 2016, 2016, 95–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Short, K.R.; Habets, M.N.; Hermans, P.W.; Diavatopoulos, D.A. Interactions between Streptococcus pneumoniae and influenza virus: A mutually beneficial relationship? Future Microbiol. 2012, 7, 609–624. [Google Scholar] [CrossRef] [PubMed]
- McCullers, J.A.; McAuley, J.L.; Browall, S.; Iverson, A.R.; Boyd, K.L.; Henriques Normark, B. Influenza enhances susceptibility to natural acquisition of and disease due to Streptococcus pneumoniae in ferrets. J. Infect. Dis. 2010, 202, 1287–1295. [Google Scholar] [CrossRef] [Green Version]
- Mansbach, J.M.; Hasegawa, K.; Piedra, P.A.; Avadhanula, V.; Petrosino, J.F.; Sullivan, A.F.; Espinola, J.A.; Camargo, C.A. Haemophilus-Dominant Nasopharyngeal Microbiota Is Associated with Delayed Clearance of Respiratory Syncytial Virus in Infants Hospitalized for Bronchiolitis. J. Infect. Dis. 2019, 219, 1804–1808. [Google Scholar] [CrossRef]
- Korten, I.; Ramsey, K.; Mika, M.; Usemann, J.; Frey, U.; Hilty, M.; Latzin, P. Nasal Microbiota and Respiratory Tract Infections: The Role of Viral Detection. Am. J. Respir. Crit. Care Med. 2019, 199, 919–922. [Google Scholar] [CrossRef]
- Ramakrishnan, V.R.; Holt, J.; Nelson, L.F.; Ir, D.; Robertson, C.E.; Frank, D.N. Determinants of the Nasal Microbiome: Pilot Study of Effects of Intranasal Medication Use. Allergy Rhinol. (Provid.) 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perl, T.M.; Cullen, J.J.; Wenzel, R.P.; Zimmerman, M.B.; Pfaller, M.A.; Sheppard, D.; Twombley, J.; French, P.P.; Herwaldt, L.A.; Mupirocin and the Risk of Staphylococcus Aureus Study Team. Intranasal mupirocin to prevent postoperative Staphylococcus aureus infections. N. Engl. J. Med. 2002, 346, 1871–1877. [Google Scholar] [CrossRef]
- Smith, T.L.; Litvack, J.R.; Hwang, P.H.; Loehrl, T.A.; Mace, J.C.; Fong, K.J.; James, K.E. Determinants of outcomes of sinus surgery: A multi-institutional prospective cohort study. Otolaryngol. Head Neck Surg. 2010, 142, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Feazel, L.M.; Robertson, C.E.; Ramakrishnan, V.R.; Frank, D.N. Microbiome complexity and Staphylococcus aureus in chronic rhinosinusitis. Laryngoscope 2012, 122, 467–472. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.M.; Qiao, X.L.; Ai, L.; Zhai, J.J.; Wang, X.X. Isolation of antimicrobial resistant bacteria in upper respiratory tract infections of patients. 3 Biotech. 2016, 6, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, G.B.; Shaw, D.; Marsh, R.L.; Carroll, M.P.; Serisier, D.J.; Bruce, K.D. Respiratory microbiota: Addressing clinical questions, informing clinical practice. Thorax 2015, 70, 74–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bode, L.G.; Kluytmans, J.A.; Wertheim, H.F.; Bogaers, D.; Vandenbroucke-Grauls, C.M.; Roosendaal, R.; Troelstra, A.; Box, A.T.; Voss, A.; van der Tweel, I.; et al. Preventing surgical-site infections in nasal carriers of Staphylococcus aureus. N. Engl. J. Med. 2010, 362, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Bosch, A.; van Houten, M.A.; Bruin, J.P.; Wijmenga-Monsuur, A.J.; Trzcinski, K.; Bogaert, D.; Rots, N.Y.; Sanders, E.A.M. Nasopharyngeal carriage of Streptococcus pneumoniae and other bacteria in the 7th year after implementation of the pneumococcal conjugate vaccine in the Netherlands. Vaccine 2016, 34, 531–539. [Google Scholar] [CrossRef] [Green Version]
- Salk, H.M.; Simon, W.L.; Lambert, N.D.; Kennedy, R.B.; Grill, D.E.; Kabat, B.F.; Poland, G.A. Taxa of the Nasal Microbiome Are Associated with Influenza-Specific IgA Response to Live Attenuated Influenza Vaccine. PLoS ONE 2016, 11, e0162803. [Google Scholar] [CrossRef] [Green Version]
- Bastier, P.L.; Lechot, A.; Bordenave, L.; Durand, M.; de Gabory, L. Nasal irrigation: From empiricism to evidence-based medicine. A review. Eur. Ann. Otorhinolaryngol. Head Neck Dis. 2015, 132, 281–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauser, L.J.; Ir, D.; Kingdom, T.T.; Robertson, C.E.; Frank, D.N.; Ramakrishnan, V.R. Evaluation of bacterial transmission to the paranasal sinuses through sinus irrigation. Int. Forum Allergy Rhinol. 2016, 6, 800–806. [Google Scholar] [CrossRef] [PubMed]
- Tichenor, W.S.; Thurlow, J.; McNulty, S.; Brown-Elliott, B.A.; Wallace, R.J., Jr.; Falkinham, J.O., 3rd. Nontuberculous Mycobacteria in household plumbing as possible cause of chronic rhinosinusitis. Emerg. Infect. Dis. 2012, 18, 1612–1617. [Google Scholar] [CrossRef]
- Yoder, J.S.; Straif-Bourgeois, S.; Roy, S.L.; Moore, T.A.; Visvesvara, G.S.; Ratard, R.C.; Hill, V.R.; Wilson, J.D.; Linscott, A.J.; Crager, R.; et al. Primary amebic meningoencephalitis deaths associated with sinus irrigation using contaminated tap water. Clin. Infect. Dis. 2012, 55, e79–e85. [Google Scholar] [CrossRef] [Green Version]
- Jain, R.; Hoggard, M.; Biswas, K.; Zoing, M.; Jiang, Y.; Douglas, R. Changes in the bacterial microbiome of patients with chronic rhinosinusitis after endoscopic sinus surgery. Int. Forum Allergy Rhinol. 2017, 7, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Tan, N.C.; Foreman, A.; Jardeleza, C.; Douglas, R.; Tran, H.; Wormald, P.J. The multiplicity of Staphylococcus aureus in chronic rhinosinusitis: Correlating surface biofilm and intracellular residence. Laryngoscope 2012, 122, 1655–1660. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Siu, J.; Shrestha, K.; Inthavong, K.; Shang, Y.; Douglas, R. Particle deposition in the paranasal sinuses following endoscopic sinus surgery. Comput. Biol. Med. 2020, 116, 103573. [Google Scholar] [CrossRef]
- Martens, K.; Pugin, B.; De Boeck, I.; Spacova, I.; Steelant, B.; Seys, S.F.; Lebeer, S.; Hellings, P.W. Probiotics for the airways: Potential to improve epithelial and immune homeostasis. Allergy 2018, 73, 1954–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cope, E.K.; Lynch, S.V. Novel microbiome-based therapeutics for chronic rhinosinusitis. Curr. Allergy Asthma Rep. 2015, 15, 504. [Google Scholar] [CrossRef]
- Sagar, S.; Vos, A.P.; Morgan, M.E.; Garssen, J.; Georgiou, N.A.; Boon, L.; Kraneveld, A.D.; Folkerts, G. The combination of Bifidobacterium breve with non-digestible oligosaccharides suppresses airway inflammation in a murine model for chronic asthma. Biochim. Biophys. Acta 2014, 1842, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Mack, D.R.; Ahrne, S.; Hyde, L.; Wei, S.; Hollingsworth, M.A. Extracellular MUC3 mucin secretion follows adherence of Lactobacillus strains to intestinal epithelial cells in vitro. Gut 2003, 52, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Ganzle, M.G.; Lin, X.B.; Ruan, L.; Sun, M. Diversity and dynamics of bacteriocins from human microbiome. Environ. Microbiol. 2015, 17, 2133–2143. [Google Scholar] [CrossRef] [PubMed]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nat. Rev. Microbiol. 2010, 8, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Kitz, R.; Martens, U.; Zieseniß, E.; Enck, P.; Rose, M. Probiotic, E. faecalis—Adjuvant therapy in children with recurrent rhinosinusitis. Open Med. 2012, 7, 362–365. [Google Scholar] [CrossRef] [Green Version]
- Pochard, P.; Gosset, P.; Grangette, C.; Andre, C.; Tonnel, A.B.; Pestel, J.; Mercenier, A. Lactic acid bacteria inhibit TH2 cytokine production by mononuclear cells from allergic patients. J. Allergy Clin. Immunol. 2002, 110, 617–623. [Google Scholar] [CrossRef] [Green Version]
- von der Weid, T.; Bulliard, C.; Schiffrin, E.J. Induction by a lactic acid bacterium of a population of CD4(+) T cells with low proliferative capacity that produce transforming growth factor beta and interleukin-10. Clin. Diagn Lab. Immunol. 2001, 8, 695–701. [Google Scholar] [CrossRef] [Green Version]
- Shandilya, U.K.; Jadhav, S.; Panwar, V.; Kansal, V.K. Probiotics: Potent Immunomodulatory Tool Against Allergy. Probiotics Antimicrob. Proteins 2011, 3, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Christensen, H.R.; Frokiaer, H.; Pestka, J.J. Lactobacilli differentially modulate expression of cytokines and maturation surface markers in murine dendritic cells. J. Immunol. 2002, 168, 171–178. [Google Scholar] [CrossRef]
- Spacova, I.; Petrova, M.I.; Fremau, A.; Pollaris, L.; Vanoirbeek, J.; Ceuppens, J.L.; Seys, S.; Lebeer, S. Intranasal administration of probiotic Lactobacillus rhamnosus GG prevents birch pollen-induced allergic asthma in a murine model. Allergy 2019, 74, 100–110. [Google Scholar] [CrossRef]
- Wu, C.T.; Chen, P.J.; Lee, Y.T.; Ko, J.L.; Lue, K.H. Effects of immunomodulatory supplementation with Lactobacillus rhamnosus on airway inflammation in a mouse asthma model. J. Microbiol. Immunol. Infect. 2016, 49, 625–635. [Google Scholar] [CrossRef] [Green Version]
- Toh, Z.Q.; Anzela, A.; Tang, M.L.; Licciardi, P.V. Probiotic therapy as a novel approach for allergic disease. Front. Pharmacol. 2012, 3, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Zhang, Y.; Ying, C.; Wang, D.; Du, C. Nanopore-based fourth-generation DNA sequencing technology. Genom. Proteom. Bioinform. 2015, 13, 4–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earl, J.P.; Adappa, N.D.; Krol, J.; Bhat, A.S.; Balashov, S.; Ehrlich, R.L.; Palmer, J.N.; Workman, A.D.; Blasetti, M.; Sen, B.; et al. Species-level bacterial community profiling of the healthy sinonasal microbiome using Pacific Biosciences sequencing of full-length 16S rRNA genes. Microbiome 2018, 6, 190. [Google Scholar] [CrossRef] [Green Version]
- Porto, B.N.; Moraes, T.J. The triad: Respiratory microbiome–virus–immune response in the pathophysiology of pulmonary viral infections. Expert Rev. Respir. Med. 2021, 15, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Schirinzi, A.; Cazzolla, A.P.; Lovero, R.; Lo Muzio, L.; Testa, N.F.; Ciavarella, D.; Palmieri, G.; Pozzessere, P.; Procacci, V.; Di Serio, F.; et al. New Insights in Laboratory Testing for COVID-19 Patients: Looking for the Role and Predictive Value of Human epididymis secretory protein 4 (HE4) and the Innate Immunity of the Oral Cavity and Respiratory Tract. Microorganisms 2020, 8, 1718. [Google Scholar] [CrossRef] [PubMed]
- Di Stadio, A.; Costantini, C.; Renga, G.; Pariano, M.; Ricci, G.; Romani, L. The Microbiota/Host Immune System Interaction in the Nose to Protect from COVID-19. Life (Basel) 2020, 10, 345. [Google Scholar] [CrossRef]
Figure 1.
The anatomy of the upper airways. The UAs are formed by many different cavities, including the nasal cavity, sinuses, nasopharynx, oropharynx, and laryngopharynx.
Figure 1.
The anatomy of the upper airways. The UAs are formed by many different cavities, including the nasal cavity, sinuses, nasopharynx, oropharynx, and laryngopharynx.

Figure 2.
Possible microbial interactions in the UAs. The secretions of Staphylococcus lugdunesis (i.e., lugdunin, a thiazolidine) inhibits the growth of S. aureus [53]. Similarly, Corynebacterium spp. shifts S. aureus towards a more commensal state by attenuating its virulence components, such as the agr operon and genes involved in its hemolytic activity [50,54,55]. Moreover, Corynebacterium and Cutibacterium spp. affect the growth of S. aureus, as the most commonly secreted porphyrin by Cutibacterium spp. facilitates S. aureus aggregation and biofilm formation with other UA microbes [52,56,57].
Figure 2.
Possible microbial interactions in the UAs. The secretions of Staphylococcus lugdunesis (i.e., lugdunin, a thiazolidine) inhibits the growth of S. aureus [53]. Similarly, Corynebacterium spp. shifts S. aureus towards a more commensal state by attenuating its virulence components, such as the agr operon and genes involved in its hemolytic activity [50,54,55]. Moreover, Corynebacterium and Cutibacterium spp. affect the growth of S. aureus, as the most commonly secreted porphyrin by Cutibacterium spp. facilitates S. aureus aggregation and biofilm formation with other UA microbes [52,56,57].

Figure 3.
Dysbiosis, inflammation, and altered niche parameters heavily impact respiratory physiology. These three factors are at continuous interplay. However, the direct mechanisms that may further elucidate this three-way relationship have not been defined.
Figure 3.
Dysbiosis, inflammation, and altered niche parameters heavily impact respiratory physiology. These three factors are at continuous interplay. However, the direct mechanisms that may further elucidate this three-way relationship have not been defined.


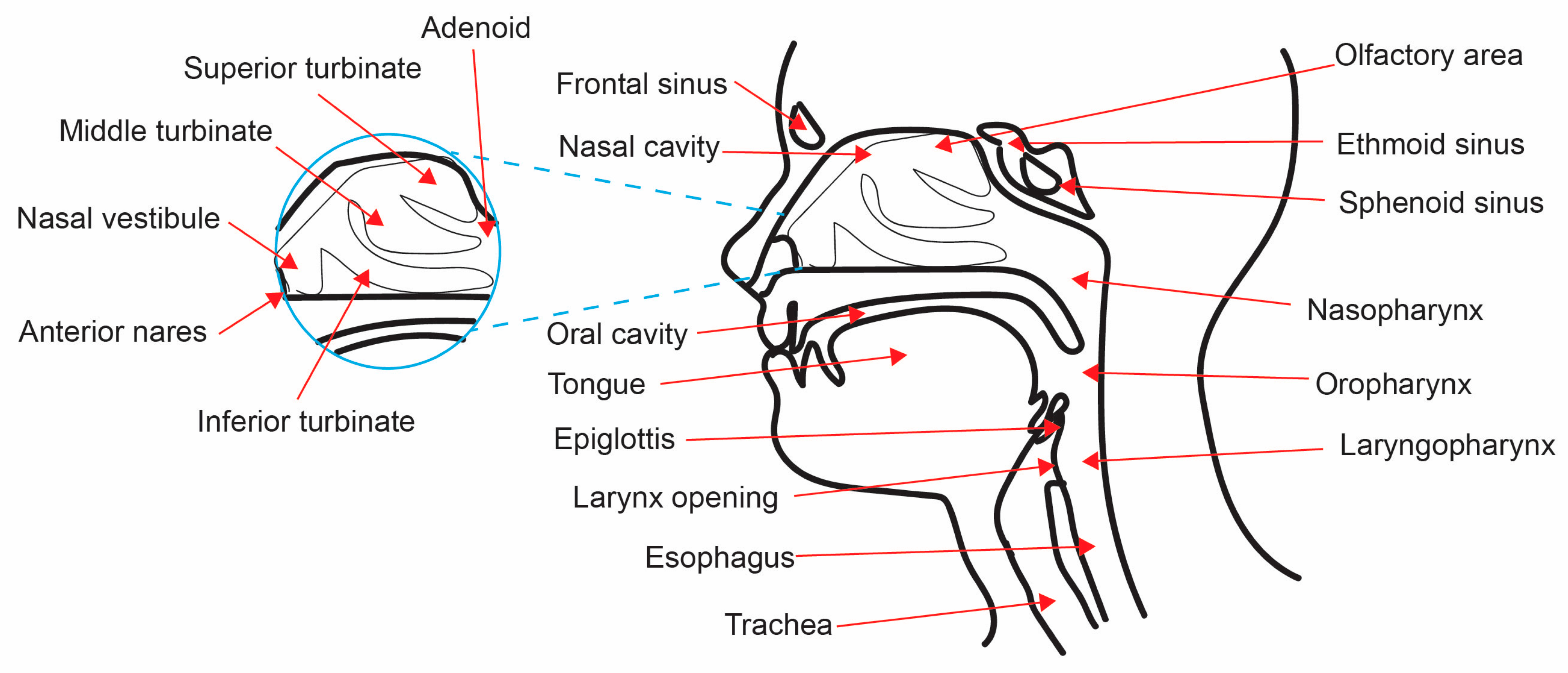{kind=link}
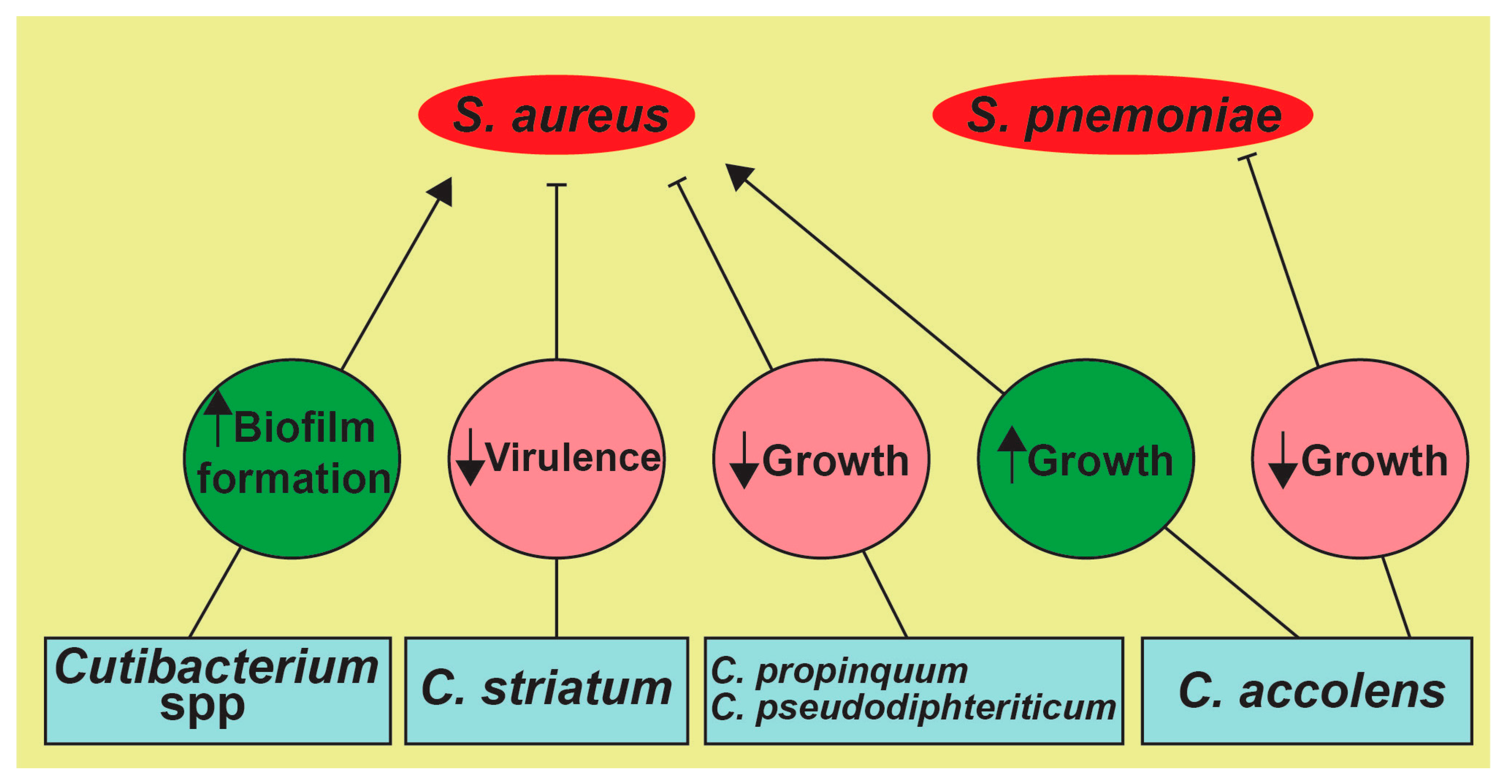{kind=link}
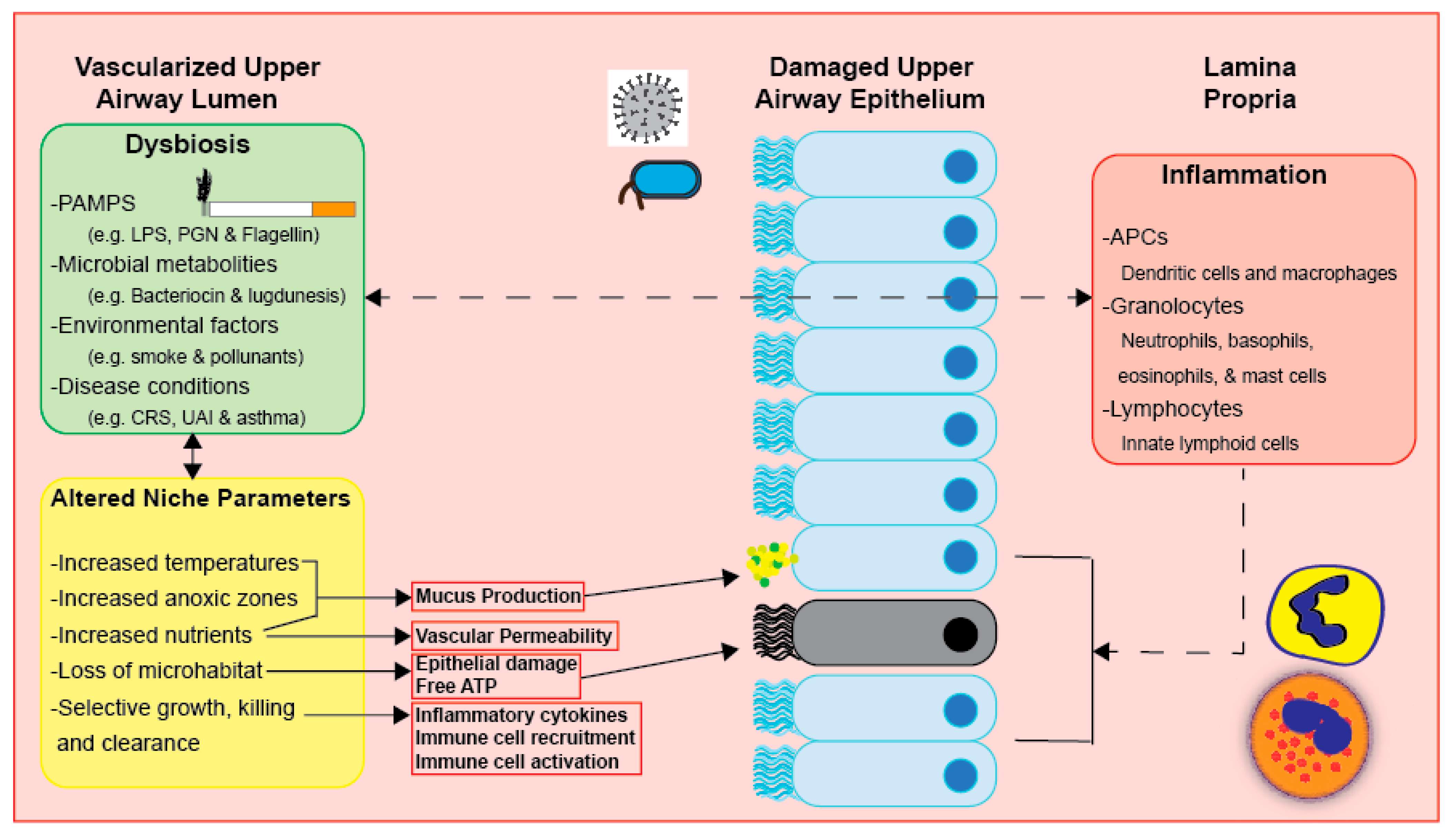{kind=link}
Table 1.
Detected UA bacteria across age in a healthy population.
| Healthy Population | Study | Sampling Site and Collection Method | Detected Bacteria |
|---|---|---|---|
| Infants | [26] | Anterior nares Nasal swab | Corynebacterium, Propionibacterium, Bifidobacterium, Streptococcus, Staphylococcus, Dolosigranulum, Moraxella |
| [26] | Nasopharynx Nasopharyngeal swab | Corynebacterium, Propionibacterium, Bifidobacterium, Bacteroides, Staphylococcus, Faecalibacterium, Streptococcus, Moraxella | |
| [26] | Oropharynx Oropharyngeal swab | Prevotella, Streptococcus, Vaillonella, Haemophilus, Moraxella, Neisseria | |
| Adults | [31] | Anterior nares Nasal swab | Corynebacterium, Propionibacterium, Prevotella, Dolosigranulum, Staphylococcus, Streptococcus, Moraxella, Escherichia shigella |
| [31] | Middle meatus Middle meatus swab | Corynebacterium, Propionibacterium, Prevotella, Dolosigranulum, Staphylococcus, Streptococcus, Moraxella, Escherichia shigella | |
| [32] | Sinus Sinus swab | Corynebacterium, Propionibacterium, Prevotella, Staphylococcus, Anaerococcus, Peptoniphilus, Ralstonia | |
| [9] | Nasopharynx Nasopharyngeal swab | Corynebacterium, Propionibacterium, Bifidobacterium, Prevotella, Sphingobacterium, Staphylococcus, Faecalibacterium, Streptococcus, Pseudomonas, Haemophilus | |
| [9] | Oropharynx Oropharyngeal swab | Corynebacterium, Rothia, Prevotella, Porphyromonas, Streptococcus, Vaillonella, Haemophilus, Moraxella | |
| Elderly | [28] | Anterior nares Nasal swab | Corynebacterium, Propionibacterium, Bifidobacterium, Prevotella, Bacteroides, Streptococcus, Staphylococcus, Vaillonella, Moraxella, Pseudomonas |
| [28] | Nasopharynx Nasopharyngeal swab | Corynebacterium, Propionibacterium, Bifidobacterium, Prevotella, Bacteroides, Streptococcus, Staphylococcus, Vaillonella, Moraxella, Pseudomonas | |
| [28] | Oropharynx Oropharyngeal swab | Corynebacterium, Propionibacterium, Bifidobacterium, Prevotella, Bacteroides, Streptococcus, Staphylococcus, Vaillonella, Moraxella, Pseudomonas |
Table 2.
Environmental exposures and the changes in the UA microbiota.
| Exposure | Study | Population and Sampling Site | Detected Bacteria |
|---|---|---|---|
| Active cigarette smoke | [30] | Adults Nasopharynx | ↓ Actinomycetaceae, ↓ Corynebacteriaceae, ↓ Coriobacteriaceae, ↑ Eggerthella, ↓ Flexibacteriaceae, ↓ Flavobacteriaceae, ↑ Porphromonadaceae, ↓ Leuconostocaceae, ↑ Erysipelotrichaceae, ↑ Aerococcaceae, ↑ Eubacteriaceae, ↑ Incertae Sedis XIII, ↑ Peptostretococcaceae, ↑ Ruminococcaceae, ↑ Lachnospiraceae I.S. spp., ↑ Anaerovorax, ↑ Dorea, ↑ Erysipelotrichaceae I.S., ↑ Eubacterium spp., ↑ Abiotrophia spp., ↓ Rhodocyclaceae, ↓ Rhodobacteraceae, ↓ Enterobacteriaceae, ↓ Alcalgenaceae, ↓ Methylophilacea, ↓ Shigella spp., ↑ Pasteurellaceae, ↑ Haemophilus spp. |
| [84] | Adults Nasopharynx | ↑ Streptococcus pneumoniae, ↑ Streptococcus pyogenes, ↑ H. influenzae, ↑ M. catarrhalis | |
| Passive cigarette smoke | [85] | Infants (<60 months) in the USA Nasopharynx | ↑ Streptococcus pneumoniae |
| Tobacco samples from cigarettes | [86] | Cigarettes | Bacillus, Clostridium, Enterococcus, Staphylococcus, Acinetobacter, Burkholderia, Klebsiella, Pseudomonas aeruginosa, Serratia, Campylobacter, Proteus |
| Air pollutants: PM2.5, PM10, SO2, NO, O3 | [87] | Healthy young adults in northeastern China Oropharynx | ↓ Prevotella, ↓ Fusobacterium, ↓ Capnocytophaga, ↓ Veillonella, ↓ Campylobacter |
| Air pollutants: PM2.5, NO2 | [83] | Healthy infants Nasal swabs | ↓ Corynebacteriaceae (NO2 only) |
| Air pollutants: PM2.5, PM10 | [81] | Healthy subjects in Italy Nasal cavity | (−) Rothia, (−) Corynebacterium, (−) Flavobacterium, (−) Finegoldia, (−) Oribacterium, (−) Streptococcus, (−) Neisseria, (−) Haemophilus, (−) Actinobacillus, (+) Moraxella |
| [82] | Vendors at an open-air farmer’s market in China Oropharynx | ↑ Leptotrichia, ↓ Prevotella 7, ↑ Streptococcus, ↑ Staphylococcus, ↑ Haemophilus, ↑ Moraxella | |
| Air pollutants: O3 | [88] | Healthy young adults in China Nasal cavity | ↑ Moraxellaceae, ↑ Pseudomonadaceae |
| Household air pollutants | [89] | Healthy adults in Malawi Nasal cavity | ↓ Tropheryma, ↑ Streptococcus, ↓ Neisseria, ↑ Petrobacter |
Symbols: (↑ or ↓) represent the higher or lower presence of said bacteria detected upon exposure, respectively, while (− and +) indicate a negative or positive correlation with the said disease condition, respectively.
Table 3.
Disease status determines the UA microbial composition.
| Exposure | Study | Population and Sampling Site | Detected Bacteria |
|---|---|---|---|
| Chronic Rhinosinusitis (CRS) | [99] | Adults Middle meatus | ↑ Corynebacterium, ↑ Curtobacteria, ↑ Staphylococcus, ↑ Pseudomonas |
| [102] | Adults Sinus | Corynebacteriaceae, Staphylococcaceae, Streptococcaceae, Pseudomonadaceae | |
| [100] | Adults with nasal polyps Middle meatus | (+) Fusobacterium, (+) Streptococcus, (+) Haemophilus | |
| [100] | Adults without nasal polyps Middle meatus | Corynebacterium, Staphylococcus, Alloiococcus | |
| [101] | Adults Middle Meatus | ↓ Corynebacterium, ↑ Porphyromonas, ↑ Prevotella, ↑ Anaerococcus, ↑ Lactobacillus, ↑ Finegoldia, ↑ Peptoniphilus, ↑ Dialister, ↑ Parvimonas, ↓ Staphylococcus, ↓ Dolosigranulum | |
| [103] | Adults Nasopharynx | ↓ Corynebacterium, ↓ Propionibacterium, ↓ Anaerococcus, ↓ Finegoldia, ↓ Peptoniphilus, ↓ Staphylococcus | |
| Asthma | [104] | Adults (18–45 years old) in Korea Nasopharynx | ↓ Corynebacterium, ↓ Moraxella |
| Non-Exacerbated Asthma | [105] | Adults (mean age 36 years old) Nasopharynx | ↑ Dialister |
| Exacerbated Asthma | [105] | Adults (mean age 36 years old) Nasopharynx | ↑ Gardnerella, ↑ Prevotella, ↑ Alkanindiges |
| Obstructive Sleep Apnea | [62] | Adults in the USA and Spain Nasopharynx | ↑ Streptococcus, ↑ Prevotella, ↑ Veillonella |
| Influenza A Virus | [106] | Infants (under the age of 2 years old) Oropharynx | ↑ Streptococcus pneumoniae, ↑ Staphylococcus aureus |
| [107] | Patients (up to 52 years old) Nasopharynx | ↑ Corynebacterium spp., ↑ Streptococcus, ↓ Pseudomonas | |
| [108] | Children in China Nasopharynx/Oropharynx | ↑ Dolosigranulum spp., ↑ Staphylococcus aureus, ↑ Phyllobacterium spp., ↑ Moraxella spp. | |
| Adenovirus | [109] | Patients (mean age 24–25 years) in South Africa Nasopharynx | ↑ Streptococcus pneumoniae |
| Respiratory Syncytial Virus | [110] | Hospitalized infants (under 1 year of age) in the USA Nasopharynx | ↑ Haemophilus spp. |
| SARS-CoV-2 | [111] | Elderly patients in China Oropharynx | ↑ Klebsiella pneumoniae, ↑ Enterococcus spp., ↑ Staphylococcus |
| [112] | Children (7–139 months old) Posterior nasopharynx and oropharynx | ↓ Bacteroidetes, ↓ Firmicutes, ↑ Proteobacteria, ↑ Pseudomonas, ↑ Herbaspirillum, ↑ Burkholderia, and ↑ Comamonadaceae_U | |
| [113] | Adults (17–68 years old) Throat swabs | ↑ Bacteroidales, ↑ Fusobacterium, ↑ Porphyromonas, ↑ Prevotella, and ↑ Neisseria | |
| [114] | >18-year-old, nonhospitalized COVID-19 patients Midturbinate swab | ↑ Peptoniphilus lacrimalis, ↑ Campylobacter hominis, ↑ Prevotella 9 copri, ↑ Anaerococcus, ↓ Corynebacterium, ↓ Staphylococcus haemolyticus, ↓ Prevotella disiens, and ↓ Corynebacterium | |
| Middle East Respiratory Syndrome (MERS) Coronavirus | [115] | Adults (47–74 years old) in Saudi Arabia Oropharyngeal swabs and tracheal aspirates | (+) Acinetobacter baumannii, (+) Pseudomonas aeruginosa, and (+) Streptococcus pneumoniae |
| [116] | Young children (under 3 years old) in Peru Nasopharynx | ↑ Streptococcus pneumoniae, ↑ Staphylococcus aureus | |
| Rhinovirus-A | [109] | Patients (mean age 24–25 years) in South Africa Nasopharynx | ↑ Streptococcus pneumoniae, ↑ Staphylococcus aureus |
| [110] | Hospitalized infants (under 1 year of age) in the USA Nasopharynx | ↑ Haemophilus spp. | |
| Rhinovirus-C | [116] | Young children (under 3 years old) in Peru Nasopharynx | ↑ Streptococcus pneumoniae, ↑ Staphylococcus aureus |
| [109] | Patients (mean age 24–25 years) in South Africa Nasopharynx | ↑ Streptococcus pneumoniae, ↑ Staphylococcus aureus | |
| [110] | Hospitalized infants (under 1 year of age) in the USA Nasopharynx | ↑ Moraxella spp. |
Symbols: (↑ or ↓) represent the higher or lower presence of said bacteria detected upon exposure, respectively, while (+) indicate a positive correlation with the said disease condition, respectively.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Elgamal, Z.; Singh, P.; Geraghty, P. The Upper Airway Microbiota, Environmental Exposures, Inflammation, and Disease. Medicina 2021, 57, 823. https://doi.org/10.3390/medicina57080823
AMA Style
Elgamal Z, Singh P, Geraghty P. The Upper Airway Microbiota, Environmental Exposures, Inflammation, and Disease. Medicina. 2021; 57(8):823. https://doi.org/10.3390/medicina57080823
Chicago/Turabian StyleElgamal, Ziyad, Pratyush Singh, and Patrick Geraghty. 2021. "The Upper Airway Microbiota, Environmental Exposures, Inflammation, and Disease" Medicina 57, no. 8: 823. https://doi.org/10.3390/medicina57080823