
Structural and Functional Characterization of Orcokinin B-like Neuropeptides in the Cuttlefish (Sepia officinalis)

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
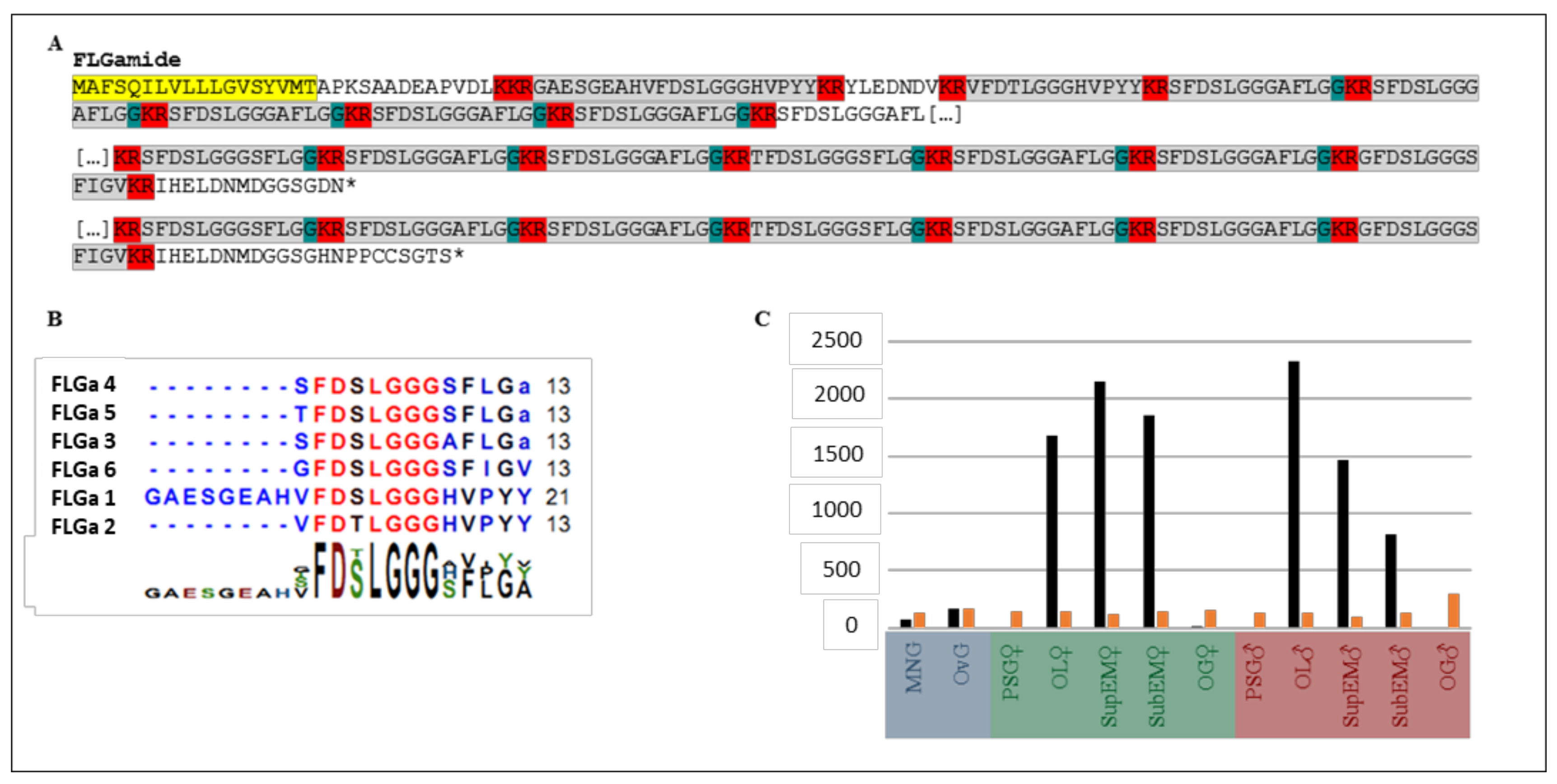
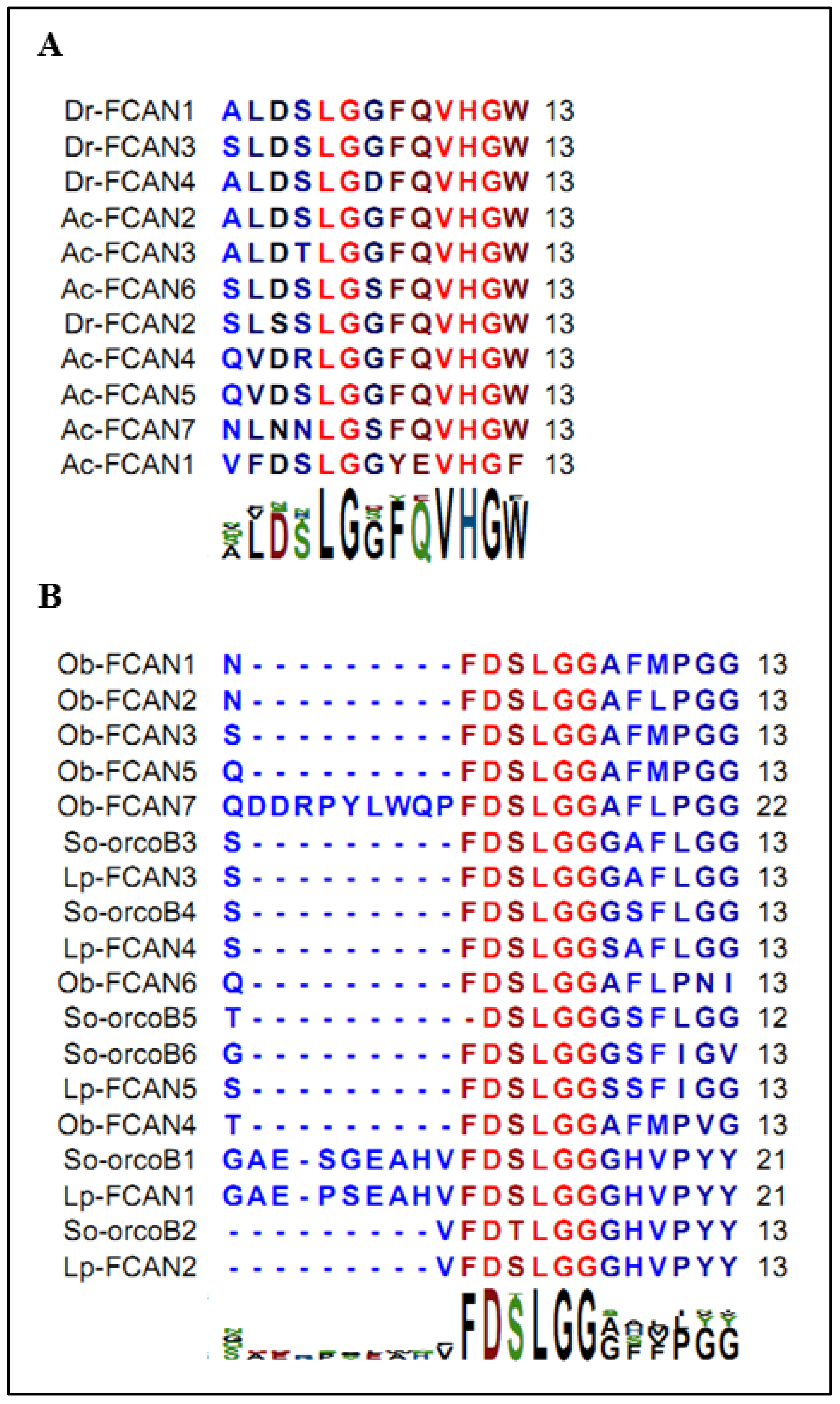
2.1. Identification of FLGamide Precursors
2.2. Tissue Mapping by Mass Spectrometry
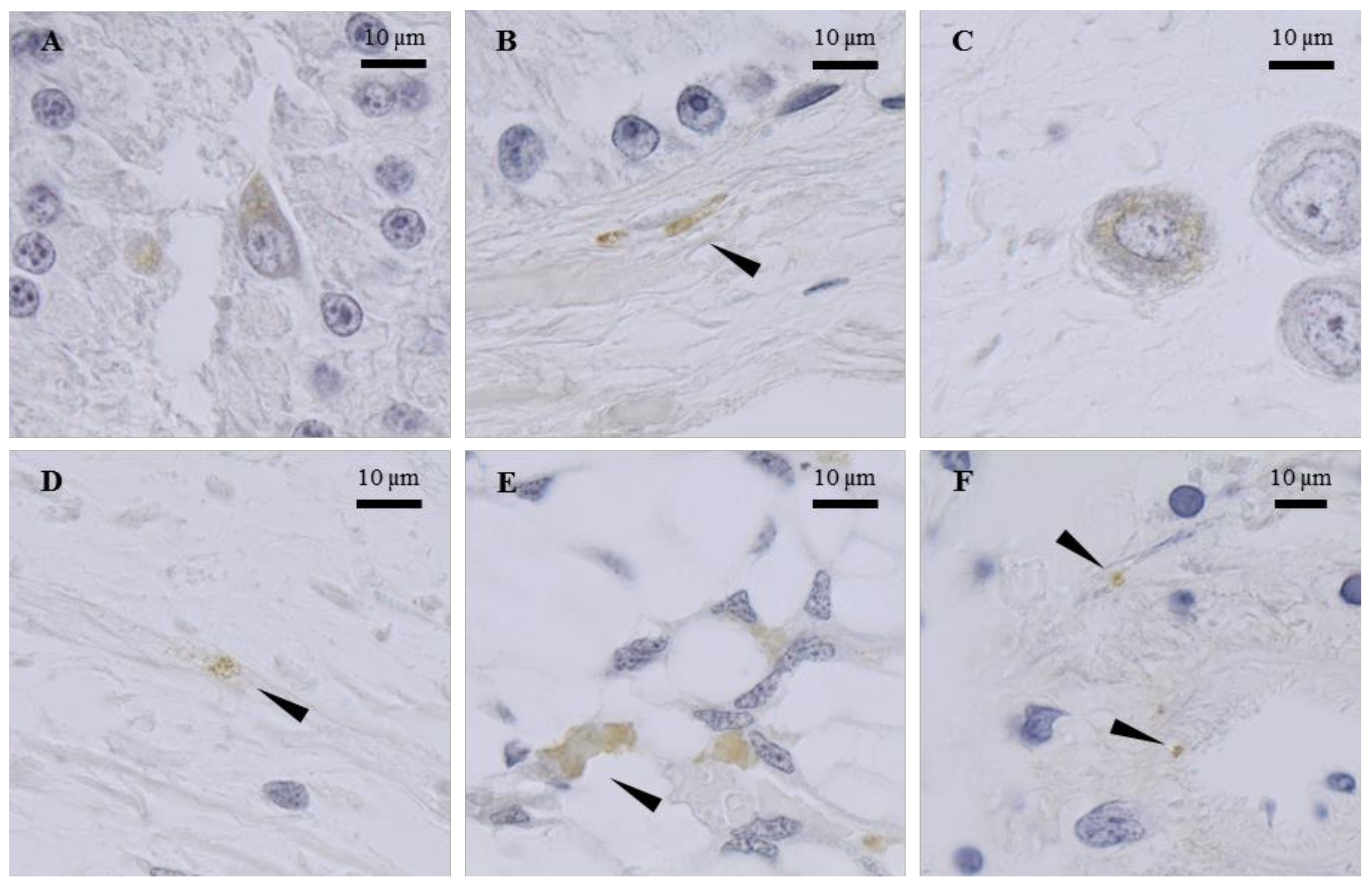
2.3. Tissue Mapping by Immunocytochemistry
2.4. Structure Determination
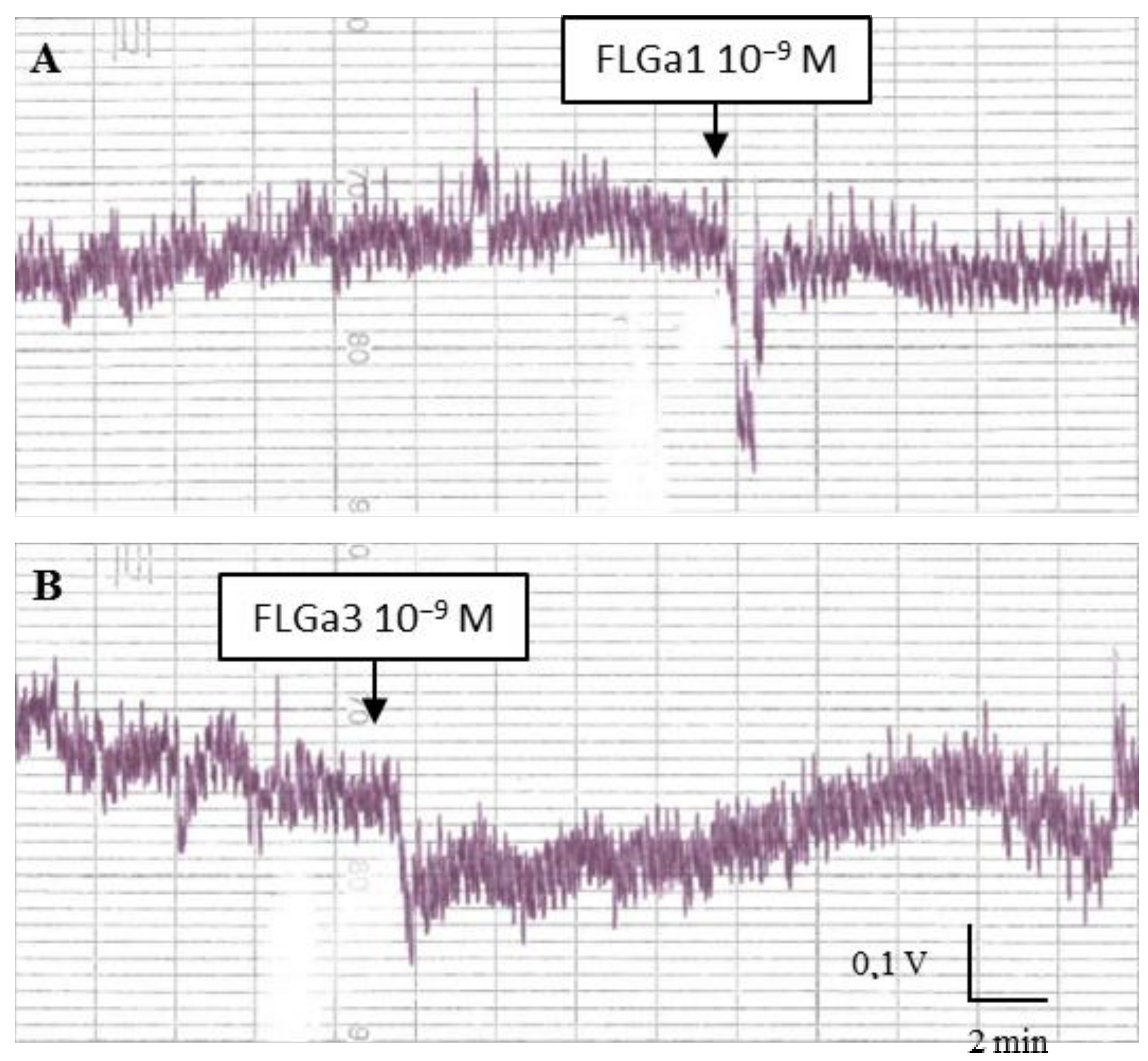
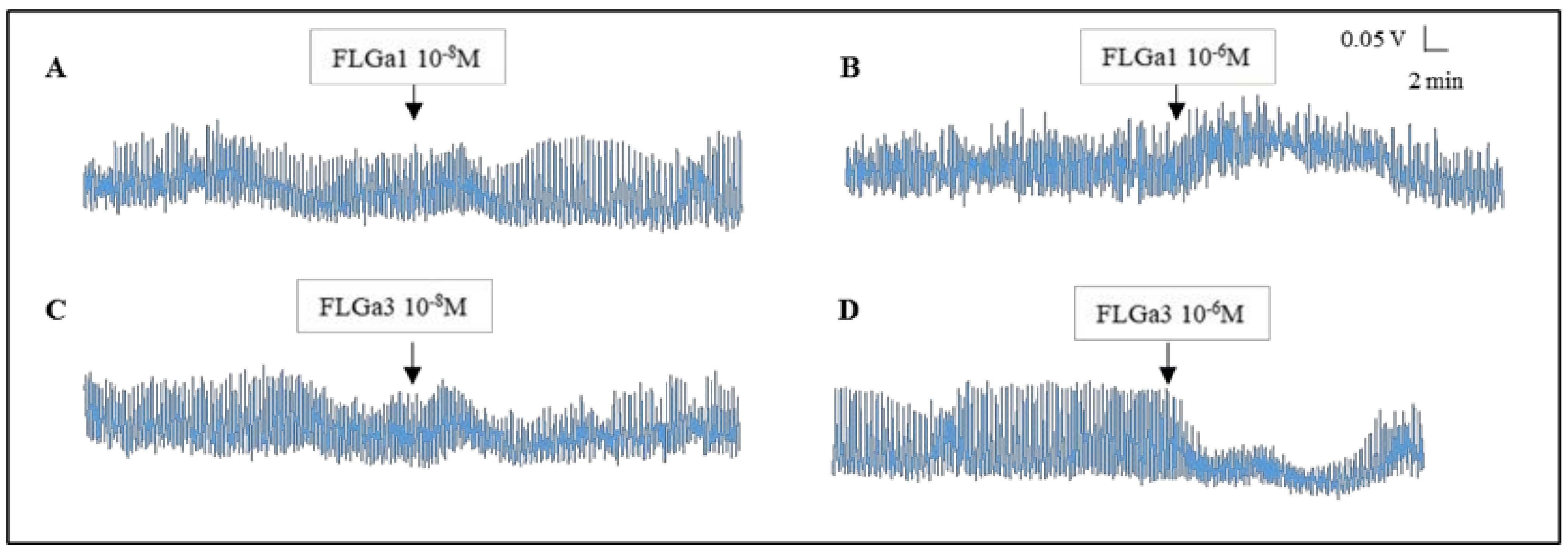
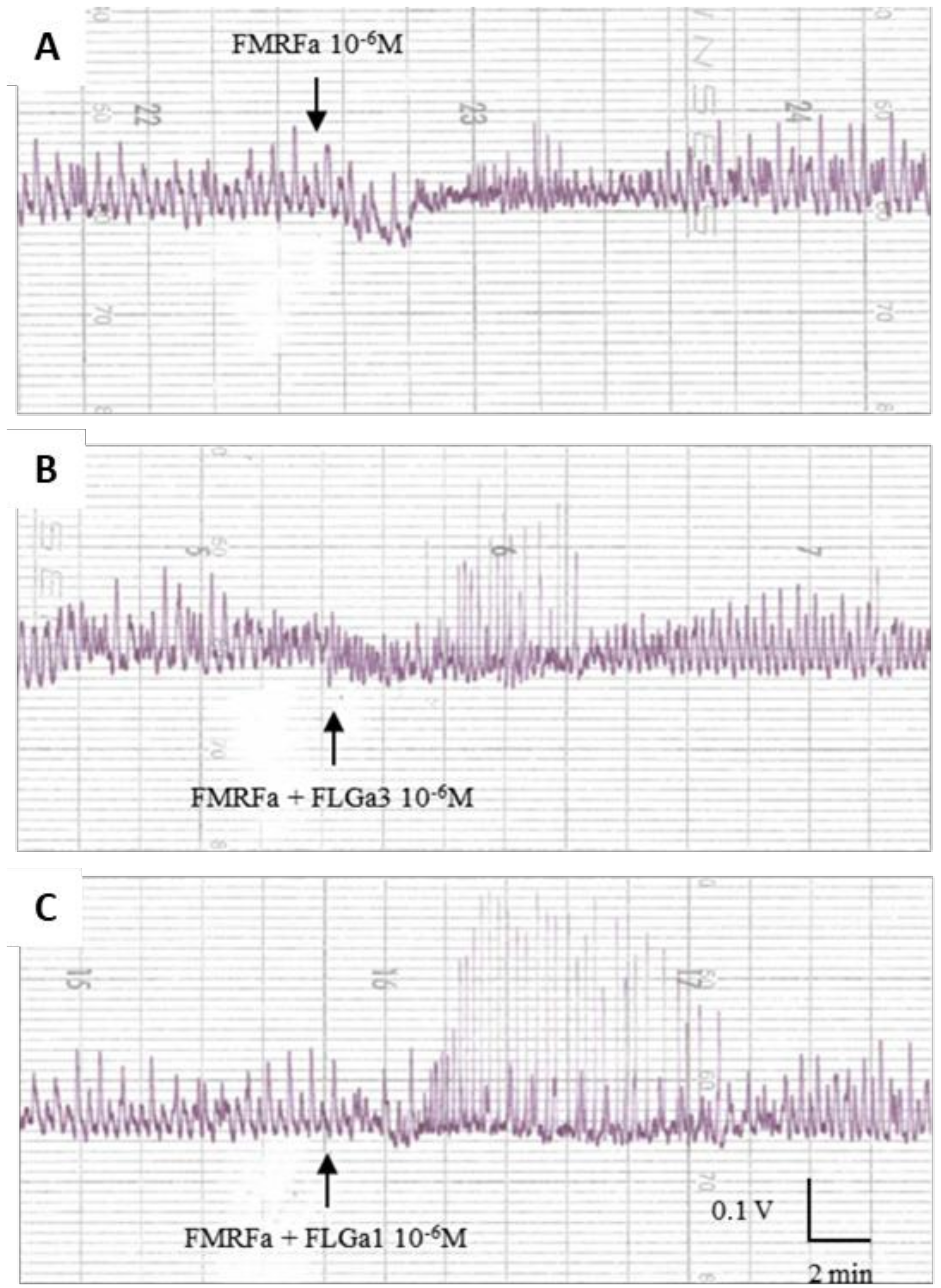
2.5. Biological Activity
3. Discussion
4. Materials and Methods
4.1. Animals and Tissue Collection
4.2. In Silico Analyses
4.3. Tissue Mapping by Mass Spectrometry
4.3.1. Extraction
4.3.2. NanoLC-MALDI-TOF/TOF Analysis
Sample Preparation for Mass Spectrometry Analysis
Mass Spectrometry Analysis
Peptide Sequencing and Protein Precursor Identification
4.4. Tissue Mapping by Immunocytochemistry
4.5. Peptide Synthesis
4.6. NMR and Structure Determination
4.6.1. NMR Measurements
4.6.2. Structure Calculations
4.7. Myotropic Bioassay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zatylny, C.; Gagnon, J.; Boucaud-Camou, E.; Henry, J. The SepOvotropin: A new ovarian peptide regulating oocyte transport in Sepia officinalis. Biochem. Biophys. Res. Commun. 2000, 276, 1013–1018. [Google Scholar] [CrossRef]
- Bernay, B.; Gagnon, J.; Henry, J. Egg capsule secretion in invertebrates: A new ovarian regulatory peptide identified by mass spectrometry comparative screening in Sepia officinalis. Biochem. Biophys. Res. Commun. 2004, 314, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Bernay, B.; Baudy-Floc’h, M.; Zanuttini, B.; Gagnon, J.; Henry, J. Identification of SepCRP analogues in the cuttlefish Sepia officinalis: A novel family of ovarian regulatory peptides. Biochem. Biophys. Res. Commun. 2005, 338, 1037–1047. [Google Scholar] [CrossRef] [PubMed]
- Zatylny-Gaudin, C.; Henry, J. Egg-Laying in the Cuttlefish Sepia officinalis. In Biological Resources of Water; InTechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Henry, J.; Cornet, V.; Bernay, B.; Zatylny-Gaudin, C. Identification and expression of two oxytocin/vasopressin-related peptides in the cuttlefish Sepia officinalis. Peptides 2013, 46, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Henry, J.; Zatylny, C.; Boucaud-Camou, E. Peptidergic control of egg-laying in the cephalopod Sepia officinalis: Involvement of FMRFamide and FMRFamide-related peptides. Peptides 1999, 20, 1061–1070. [Google Scholar] [CrossRef]
- Henry, J.; Zatylny, C. Identification and tissue mapping of APGWamide-related peptides in Sepia officinalis using LC-ESI-MS/MS. Peptides 2002, 23, 1031–1037. [Google Scholar] [CrossRef]
- Bernay, B.; Baudy-Floc’h, M.; Gagnon, J.; Henry, J. Ovarian jelly-peptides (OJPs), a new family of regulatory peptides identified in the cephalopod Sepia officinalis. Peptides 2006, 27, 1259–1268. [Google Scholar] [CrossRef]
- Zatylny, C.; Gagnon, J.; Boucaud-Camou, E.; Henry, J. ILME: A Waterborne Pheromonal Peptide Released by the Eggs of Sepia officinalis. Biochem. Biophys. Res. Commun. 2000, 275, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Enault, J.; Zatylny-Gaudin, C.; Bernay, B.; Lefranc, B.; Leprince, J.; Baudy-Floc’h, M.; Henry, J. A complex set of sex pheromones identified in the cuttlefish Sepia officinalis. PLoS ONE 2012, 7, e46531. [Google Scholar] [CrossRef]
- Cornet, V.; Henry, J.; Goux, D.; Duval, E.; Bernay, B.; Le Corguillé, G.; Corre, E.; Zatylny-Gaudin, C. How egg case proteins can protect cuttlefish offspring? PLoS ONE 2015, 10, e0132836. [Google Scholar] [CrossRef] [Green Version]
- Boletzky, S. Fecundity variation in relation to intermittent or chromic spawning in the cuttlefish, Sepia officinalis (Mollusca, Cephalopoda). Bull. Mar. Sci. 1987, 40, 382–387. [Google Scholar]
- Van In, V.; Ntalamagka, N.; O’Connor, W.; Wang, T.; Powell, D.; Cummins, S.F.; Elizur, A. Reproductive neuropeptides that stimulate spawning in the Sydney Rock Oyster (Saccostrea glomerata). Peptides 2016, 82, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Arch, S. Biosynthesis of the egg-laying hormone (ELH) in the bag cell neurons of Aplysia californica. J. Gen. Physiol. 1972, 60, 102–119. [Google Scholar] [CrossRef] [Green Version]
- Chiu, A.Y.; Hunkapiller, M.W.; Heller, E.; Stuart, D.K.; Hood, L.E.; Strumwasser, F. Purification and primary structure of the neuropeptide egg-laying hormone of Aplysia californica. Proc. Natl. Acad. Sci. USA 1979, 76, 6656–6660. [Google Scholar] [CrossRef] [Green Version]
- Chiu, A.; Strumwasser, F. An immunohistochemical study of the neuropeptidergic bag cells of Aplysia. J. Neurosci. 1981, 1, 812–826. [Google Scholar] [CrossRef]
- Ebberink, R.H.; van Loenhout, H.; Geraerts, W.P.; Joosse, J. Purification and amino acid sequence of the ovulation neurohormone of Lymnaea stagnalis. Proc. Natl. Acad. Sci. USA 1985, 82, 7767–7771. [Google Scholar] [CrossRef] [Green Version]
- Geraerts, W.P.M.; Cheeseman, P.; Ebberink, R.H.M.; Nuyt, K.; Hogenes, T.M. Partial purification and characterization of the ovulation hormone of the freshwater pulmonate snail Lymnaea stagnalis. Gen. Comp. Endocrinol. 1983, 51, 471–476. [Google Scholar] [CrossRef]
- Vreugdenhil, E.; Geraerts, W.P.M.; Jackson, J.F.; Joosse, J. The molecular basis of the neuro-endocrine control of egg-laying behaviour in Lymnaea. Peptides 1985, 6, 465–470. [Google Scholar] [CrossRef]
- York, P.S.; Cummins, S.F.; Degnan, S.M.; Woodcroft, B.J.; Degnan, B.M. Marked changes in neuropeptide expression accompany broadcast spawnings in the gastropod Haliotis asinina. Front. Zool. 2012, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Zatylny-Gaudin, C.; Cornet, V.; Leduc, A.; Zanuttini, B.; Corre, E.; Le Corguillé, G.; Bernay, B.; Garderes, J.; Kraut, A.; Couté, Y.; et al. Neuropeptidome of the cephalopod Sepia officinalis: Identification, tissue mapping, and expression pattern of neuropeptides and neurohormones during egg laying. J. Proteome Res. 2016, 15, 48–67. [Google Scholar] [CrossRef]
- Wüthrich, K. NMR of Proteins and Nucleic Acids; Wiley: New York, NY, USA, 1986. [Google Scholar]
- Alexandrowicz, J.S. The neurosecretory system of the vena cava in Cephalopoda II. Sepia officinalis and Octopus vulgaris. J. Mar. Biol. 1965, 45, 209–228. [Google Scholar] [CrossRef]
- Alexandrowicz, J.S. The Neurosecretory System of the Vena Cava in Cephalopoda I. Eledone Cirrosa. J. Mar. Biol. 1964, 44, 111–132. [Google Scholar] [CrossRef]
- Alexandrowicz, J.S. Innervation of the Hearts of Sepia officinalis. Acta Zool. 1960, 41, 65–100. [Google Scholar] [CrossRef]
- Tompset, D.H. Sepia in Liverpool Marine Biology Committee Memoirs; The University Press of Liverpool: Liverpool, UK, 1939. [Google Scholar]
- Kim, S.; Martin, K.C. Neuron-wide RNA transport combines with netrin-mediated local translation to spatially regulate the synaptic proteome. eLife 2015, 4, e04158. [Google Scholar] [CrossRef] [PubMed]
- Merianda, T.; Twiss, J. Peripheral nerve axons contain machinery for co-translational secretion of axonally-generated proteins. Neurosci. Bull. 2013, 29, 493–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Donald, J.; Hanna, P.; Nuurai, P.; Sobhon, P. Comparative distribution of a putative egg-laying hormone in neural and reproductive tissues of four Decapoda crustaceans. Invertebr. Neurosci. 2009, 9, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Sobhon, P.; Withyachumnarnkul, B.; Hanna, P. Identification of a putative egg-laying hormone in neural and ovarian tissues of the black tiger shrimp, Penaeus monodon, using immunocytochemistry. Invertebr. Neurosci. 2006, 6, 41–46. [Google Scholar] [CrossRef]
- Saitongdee, P.; Apisawetakan, S.; Anunruang, N.; Poomthong, T.; Hanna, P.; Sobhon, P. Egg-laying-hormone immunoreactivity in the neural ganglia and ovary of Haliotis asinina Linnaeus. Invertebr. Neurosci. 2005, 5, 165–172. [Google Scholar] [CrossRef]
- Endress, M.; Zatylny-Gaudin, C.; Corre, E.; Le Corguillé, G.; Benoist, L.; Leprince, J.; Lefranc, B.; Bernay, B.; Leduc, A.; Rangama, J.; et al. Crustacean cardioactive peptides: Expression, localization, structure, and a possible involvement in regulation of egg-laying in the cuttlefish Sepia officinalis. Gen. Comp. Endocrinol. 2018, 260, 67–79. [Google Scholar] [CrossRef]
- Zatylny, C.; Durantou, F.; Boucaud-Camou, E.; Henry, J. Evidence of 5-hydroxytryptamine synthesis in the follicles of Sepia officinalis and direct involvement in the control of egg-laying. Mol. Reprod. Dev. 2000, 55, 182–188. [Google Scholar] [CrossRef]
- Da Costa, G.; Bondon, A.; Coutant, J.; Curmi, P.; Monti, J.-P. Intermolecular interactions between the neurotensin and the third extracellular loop of human neurotensin 1 receptor. J. Biomol. Struct. Dyn. 2013, 31, 1381–1392. [Google Scholar] [CrossRef]
- Schwyzer, R. Molecular mechanism of opioid receptor selection. Biochemistry 1986, 25, 6335–6342. [Google Scholar] [CrossRef]
- Ons, S.; Bellés, X.; Maestro, J.L. Orcokinins contribute to the regulation of vitellogenin transcription in the cockroach Blattella germanica. J. Insect Physiol. 2015, 82, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, R.A.; Aragão, C.; Frias, P.A.; Sykes, A.V. The use of different anaesthetics as welfare promoters during short-term human manipulation of European cuttlefish (Sepia officinalis) juveniles. Aquaculture 2012, 370–371, 130–135. [Google Scholar] [CrossRef]
- Leprince, J.; Oulyadi, H.; Vaudry, D.; Masmoudi, O.; Gandolfo, P.; Patte, C.; Costentin, J.; Fauchère, J.-L.; Davoust, D.; Vaudry, H.; et al. Synthesis, conformational analysis and biological activity of cyclic analogs of the octadecaneuropeptide ODN. Eur. J. Biochem. 2001, 268, 6045–6057. [Google Scholar] [CrossRef]
- Chatenet, D.; Dubessy, C.; Boularan, C.; Scalbert, E.; Pfeiffer, B.; Renard, P.; Lihrmann, I.; Pacaud, P.; Tonon, M.C.; Vaudry, H.; et al. Structure-activity relationships of a novel series of urotensin II analogues: Identification of a urotensin II antagonist. J. Med. Chem. 2006, 49, 7234–7238. [Google Scholar] [CrossRef]
- Mo, H.; Harwood, J.S.; Yang, D.; Post, C.B. A simple method for NMR t1 noise suppression. J. Magn. Reson. 2017, 276, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Vranken, W.F.; Boucher, W.; Stevens, T.J.; Fogh, R.H.; Pajon, A.; Llinas, M.; Ulrich, E.L.; Markley, J.L.; Ionides, J.; Laue, E.D. The CCPN data model for NMR spectroscopy: Development of a software pipeline. Proteins Struct. Funct. Bioinforma 2005, 59, 687–696. [Google Scholar] [CrossRef]
- Johnson, B.A. Using NMRView to Visualize and Analyze the NMR Spectra of Macromolecules. In Protein NMR Techniques; Humana Press: Rahway, NJ, USA, 2004; pp. 313–352. [Google Scholar]
- Case, D.A.; Cheatham, T.E.; Darden, T.; Gohlke, H.; Luo, R.; Merz, K.M.; Onufriev, A.; Simmerling, C.; Wang, B.; Woods, R.J. The Amber biomolecular simulation programs. J. Comput. Chem. 2005, 26, 1668–1688. [Google Scholar] [CrossRef] [Green Version]
- Koradi, R.; Billeter, M.; Wüthrich, K. MOLMOL: A program for display and analysis of macromolecular structures. J. Mol. Graph. 1996, 14, 51–55. [Google Scholar] [CrossRef]











{kind=link}
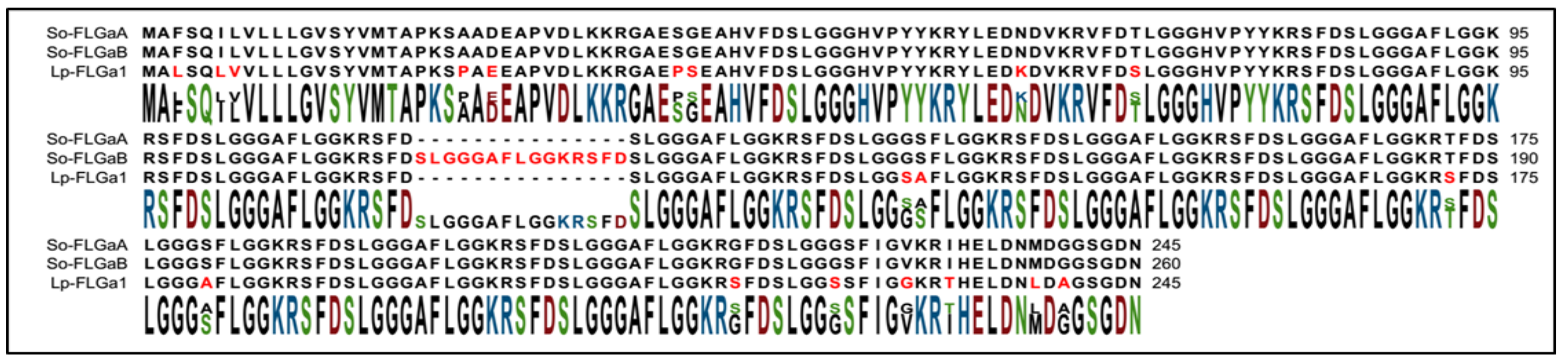{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conformational Restraints |
|---|
| Distance constraints: 119 |
| Restraints violations |
| >0.3 Å: 0 |
| Mean global r.m.s.d. (Å) |
| Heavy atoms (residues 4–14): 0.26 |
| Backbone atoms (residues 4–14): 0.746 |
| Residue | HN | HA | HB2 | HB3 | HG2 | HG3 | HD1 | HD2 | Others |
|---|---|---|---|---|---|---|---|---|---|
| Gly1 | - | 3.92/3.92 | |||||||
| Ala2 | 8.74 | 4.38 | 1.44 | ||||||
| Glu3 | 8.58 | 4.40 | 2.03 | 2.14 | |||||
| Ser4 | 8.34 | 4.46 | 3.94 | 4.03 | |||||
| Gly5 | 8.68 | 4.03/4.03 | |||||||
| Glu6 | 8.44 | 4.19 | 2.07 | 2.07 | 2.43 | 2.43 | |||
| Ala7 | 8.24 | 4.18 | 1.44 | ||||||
| His8 | 8.24 | 4.64 | 3.24 | 3.32 | 7.34 | HE1 8.66 | |||
| Val9 | 8.14 | 3.81 | 2.08 | 0.78 | 0.94 | ||||
| Phe10 | 8.08 | 4.44 | 3.13 | 3.25 | 7.28 | 7.28 | HE1/2 7.28 HZ 7.17 | ||
| Asp11 | 8.32 | 4.55 | 2.91 | 2.91 | |||||
| Ser12 | 8.04 | 4.41 | 3.92 | 3.97 | |||||
| Leu13 | 7.84 | 4.36 | 1.81 | 1.81 | 1.64 | 0.91 | 0.91 | ||
| Gly14 | 8.13 | 3.99 | 3.99 | ||||||
| Gly15 | 8.22 | 3.93 | 3.93 | ||||||
| Gly16 | 8.32 | 3.91 | 3.91 | ||||||
| His17 | 8.12 | 3.18 | 3.30 | 7.31 | HE1 8.63 | ||||
| Val18 | 8.27 | 4.35 | 2.13 | 1.02 | 1.02 | ||||
| Pro19 | - | 4.41 | 1.82 | 2.17 | 1.92 | 1.98 | 3.57 | 3.89 | |
| Tyr20 | 7.80 | 4.38 | 3.02 | 3.02 | 7.06 | 7.06 | HE1/2 6.82 | ||
| Tyr21 | 7.60 | 4.49 | 2.93 | 3.02 | 7.06 | 7.06 | HE1/2 6.82 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Endress, M.; Zatylny-Gaudin, C.; Leprince, J.; Lefranc, B.; Corre, E.; Le Corguillé, G.; Bernay, B.; Leduc, A.; Rangama, J.; Mouret, L.; et al. Structural and Functional Characterization of Orcokinin B-like Neuropeptides in the Cuttlefish (Sepia officinalis). Mar. Drugs 2022, 20, 505. https://doi.org/10.3390/md20080505
Endress M, Zatylny-Gaudin C, Leprince J, Lefranc B, Corre E, Le Corguillé G, Bernay B, Leduc A, Rangama J, Mouret L, et al. Structural and Functional Characterization of Orcokinin B-like Neuropeptides in the Cuttlefish (Sepia officinalis). Marine Drugs. 2022; 20(8):505. https://doi.org/10.3390/md20080505
Chicago/Turabian StyleEndress, Maxime, Céline Zatylny-Gaudin, Jérôme Leprince, Benjamin Lefranc, Erwan Corre, Gildas Le Corguillé, Benoît Bernay, Alexandre Leduc, Jimmy Rangama, Liza Mouret, and et al. 2022. "Structural and Functional Characterization of Orcokinin B-like Neuropeptides in the Cuttlefish (Sepia officinalis)" Marine Drugs 20, no. 8: 505. https://doi.org/10.3390/md20080505