
Cyanobacteria—From the Oceans to the Potential Biotechnological and Biomedical Applications


 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Preclinical and Clinical Trials of Metabolites from Marine Cyanobacteria
2.1. Bioactive Constituents of Marine Cyanobacteria
2.1.1. Antioxidant and Antiobesity Supplements from Cyanobacteria
2.1.2. Cytotoxic Agents from Cyanobacteria
2.1.3. Antiparasite Agents
2.1.4. Antiviral Natural Products with Anti-SARS-CoV-2 Potential from Cyanobacteria
2.2. Clinical Trials of Metabolites from Marine Cyanobacteria
3. Applications of Cyanobacteria in Biotechnology
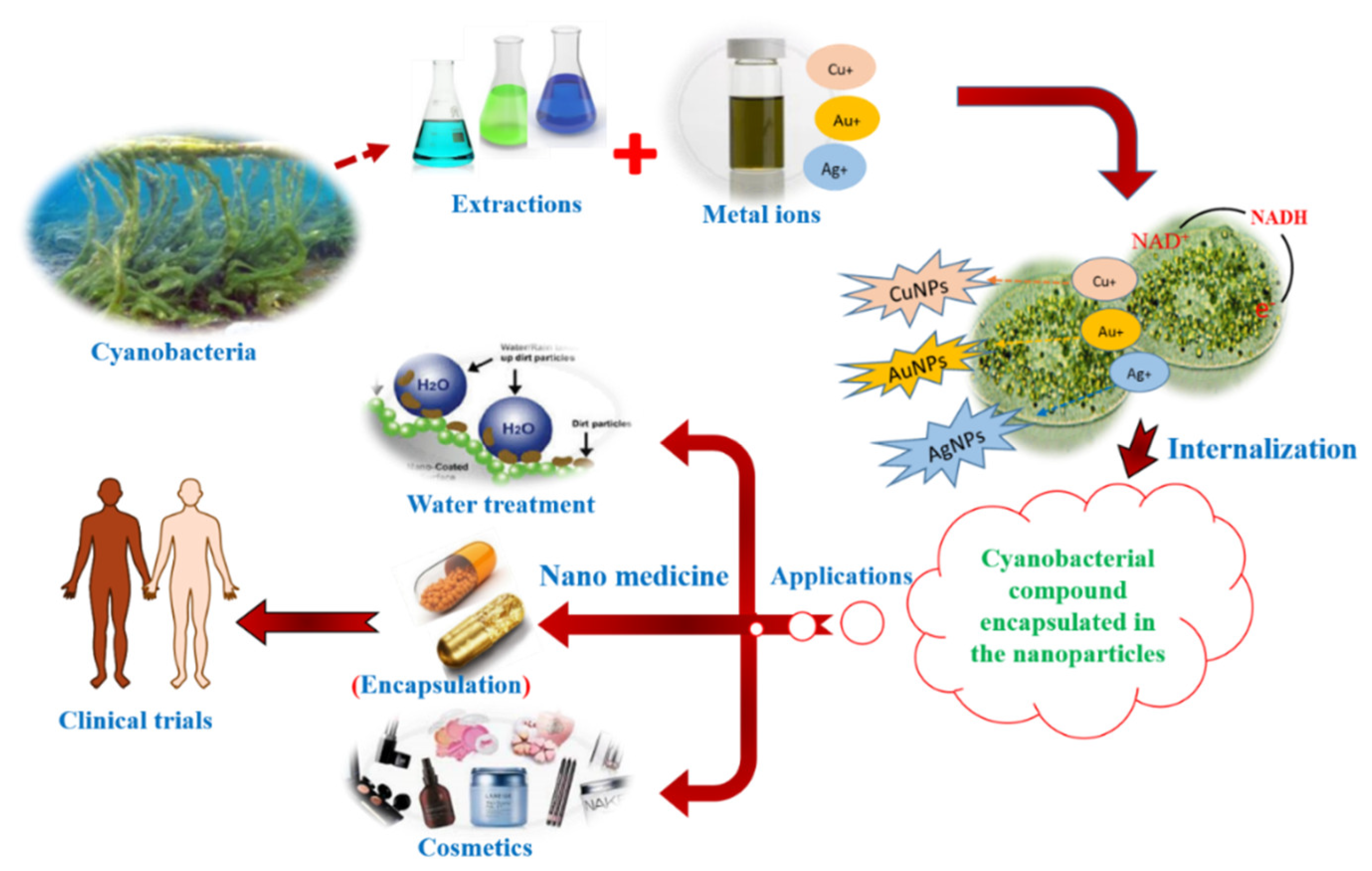
3.1. NanoBiotechnological Use of Cyanobacterial Extracts and Metabolites
3.2. Cyanobacteria: Foes or Friend of Skins, Their Use in Cosmetics
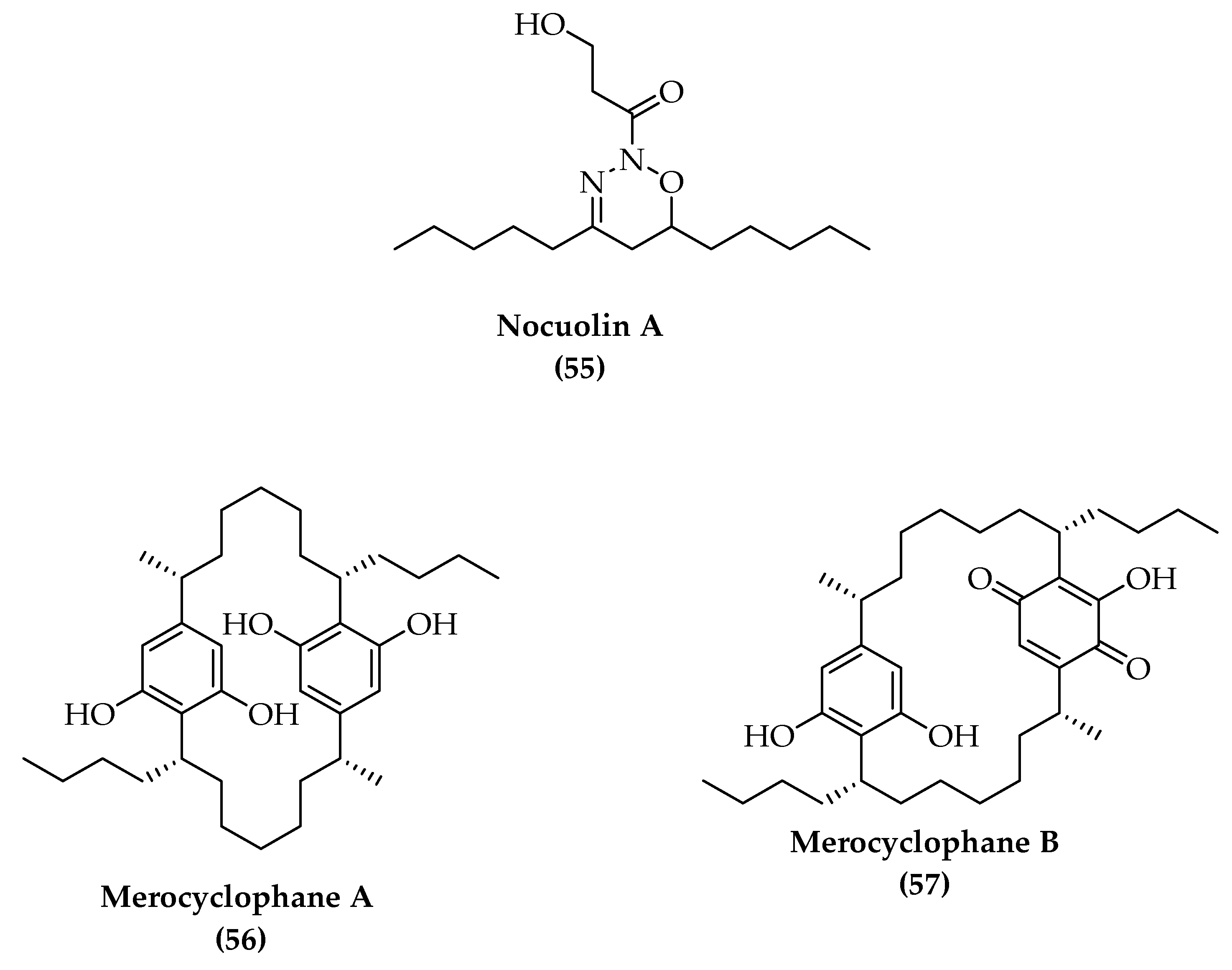
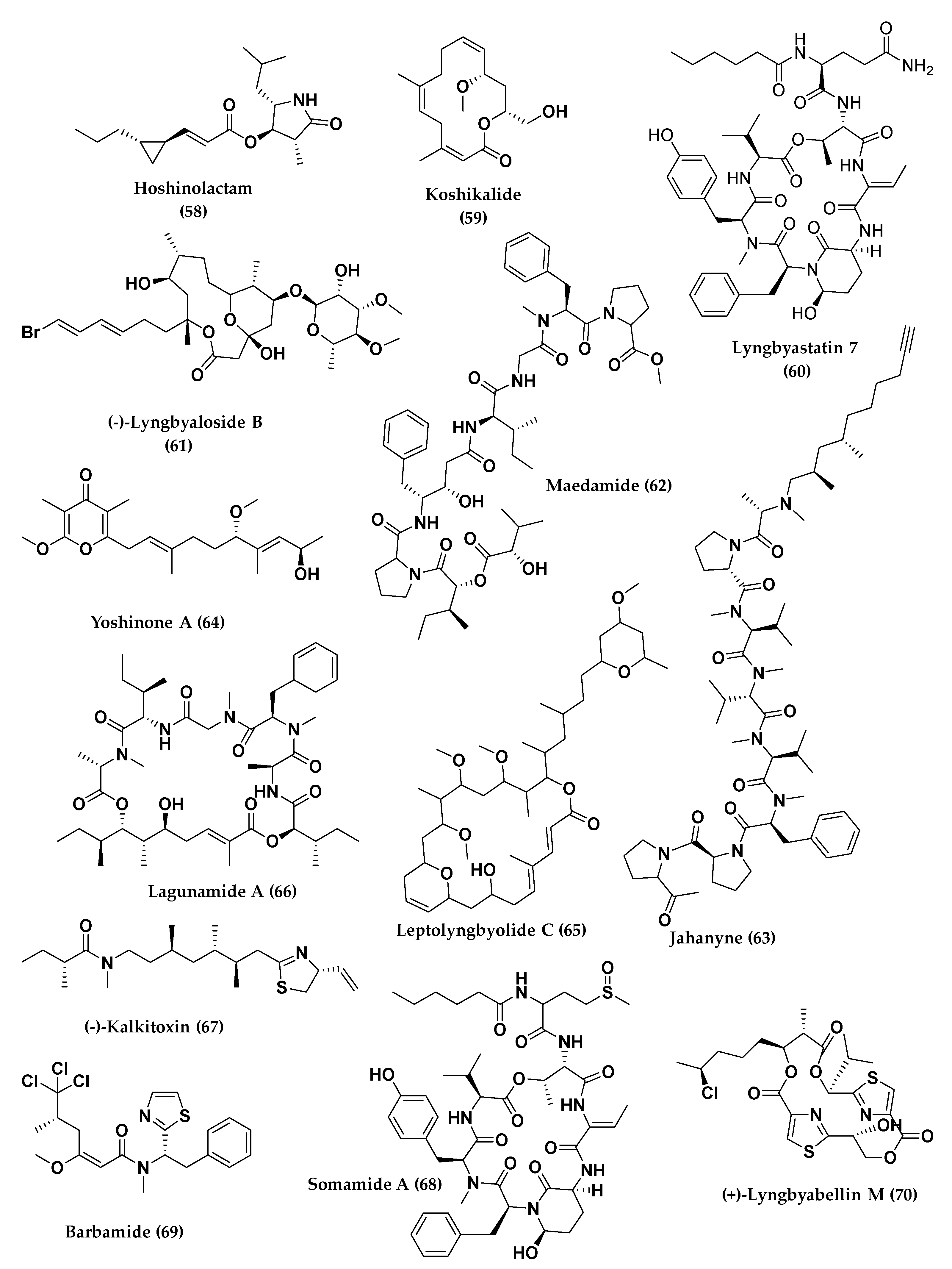
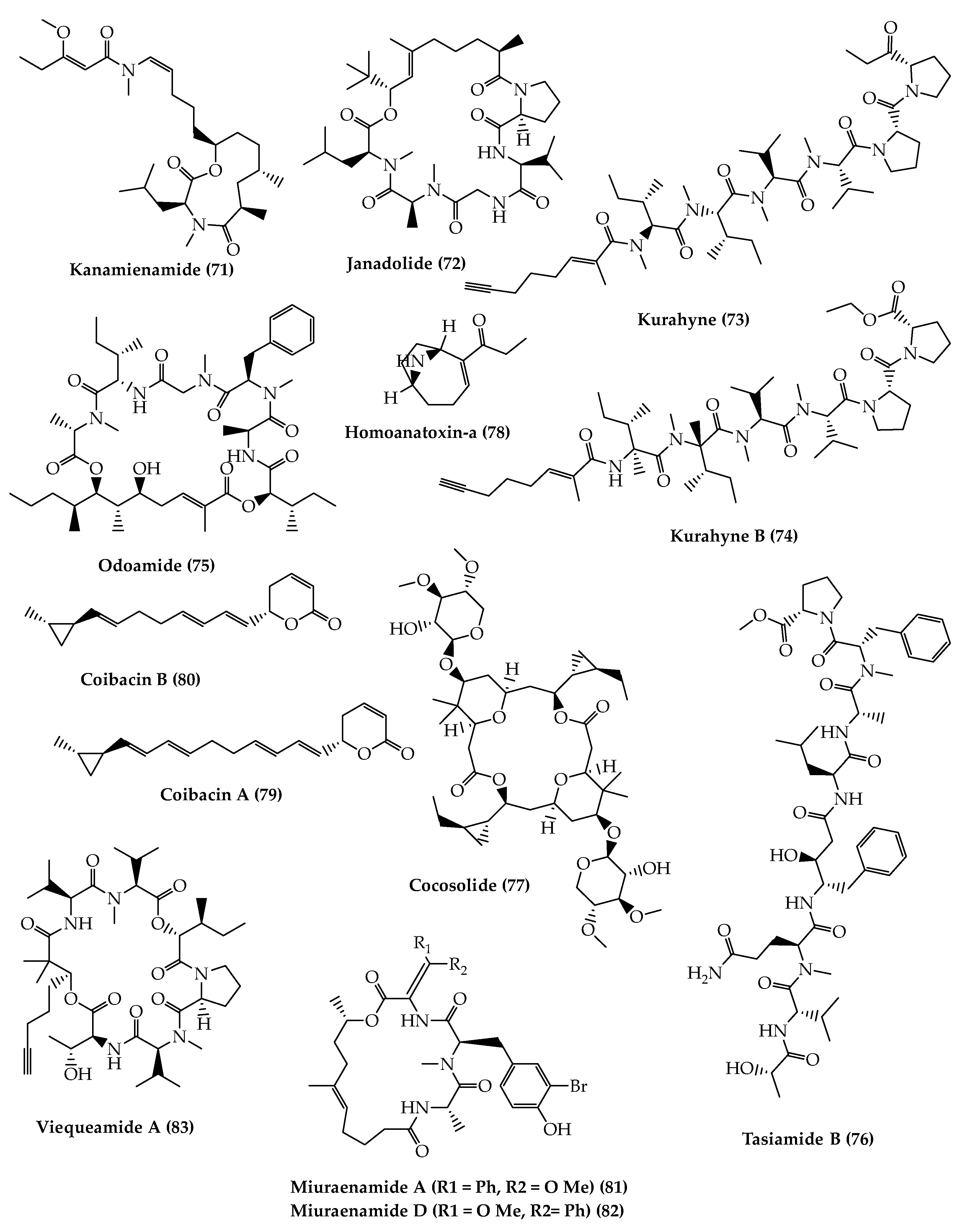
4. Total Synthesis and Stereochemical Determination of Marine Cyanobacteria Bioactive Compounds
4.1. Depsipeptides
4.2. Polyketides Peptide
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Golubic, S.; Seong-Joo, L. Early cyanobacterial fossil record: Preservation, palaeoenvironments and identification. Eur. J. Phycol. 1999, 34, 339–348. [Google Scholar] [CrossRef]
- Singh, J.S.; Kumar, A.; Rai, A.N.; Singh, D.P. Cyanobacteria: A precious bio-resource in agriculture, ecosystem, and environmental sustainability. Front. Microbiol. 2016, 7, 529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fogg, G.E.; Stewart, W.D.P.; Fay, P.; Walsby, A.E. The Blue-Green Algae; Academic Press: New York, NY, USA, 1973; p. 459. [Google Scholar]
- Flombaum, P.; Gallegos, J.L.; Gordillo, R.A.; Rincón, J.; Zabala, L.L.; Jiao, N.; Karl, D.M.; Li, W.K.W.; Lomas, M.W.; Veneziano, D. Present and future global distributions of the marine cyanobacteria Prochlorococcus and Synechococcus. Proc. Natl. Acad. Sci. USA 2013, 110, 9824–9829. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.; Garcia, M.; Costa-Rodrigues, J.; Costa, M.S.; Ribeiro, M.J.; Fernandes, M.H.; Barros, P.; Barreiro, A.; Vasconcelos, V.; Martins, R. Exploring bioactive properties of marine cyanobacteria isolated from the Portuguese coast: High potential as a source of anticancer compounds. Mar. Drugs 2014, 12, 98–114. [Google Scholar] [CrossRef] [Green Version]
- Bergman, B.; Sandh, G.; Lin, S.; Larsson, J.; Carpenter, E.J. Trichodesmium–a widespread marine cyanobacterium with unusual nitrogen fixation properties. FEMS Microbiol. Rev. 2013, 37, 286–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rastogi, R.P.; Madamwar, D.; Incharoensakdi, A. Bloom dynamics of cyanobacteria and their toxins: Environmental health impacts and mitigation strategies. Front. Microbiol. 2015, 6, 1254. [Google Scholar] [CrossRef] [Green Version]
- Bocchi, S.; Malgioglio, A. Azolla-Anabaena as a biofertilizer for rice paddy fields in the Po Valley, a temperate rice area in Northern Italy. Int. J. Agron. 2010, 2010, 5. [Google Scholar] [CrossRef] [Green Version]
- Lincoln, E.P.; Wilkie, A.C.; French, B.T. Cyanobacterial process for renovating dairy wastewater. Biomass Bioenergy 1996, 10, 63–68. [Google Scholar] [CrossRef]
- Radakovits, R.; Jinkerson, R.E.; Darzins, A.; Posewitz, M.C. Genetic engineering of algae for enhanced biofuel production. Eukaryot. Cell 2010, 9, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Pichel, F.; Ramírez-Reinat, E.; Gao, Q. Microbial excavation of solid carbonates powered by P-type ATPase-mediated transcellular Ca2+ transport. Proc. Natl. Acad. Sci. USA 2010, 107, 21749–21754. [Google Scholar] [CrossRef] [Green Version]
- Whitton, B.A.; Potts, M. The Ecology of Cyanobacteria: Their Diversity in Time and Space; Springer Science & Business Media: New York, NY, USA; Boston, MA, USA; Dordrecht, The Netherlands; London, UK; Moscow, Russia, 2007. [Google Scholar]
- Najafi, A.; Moradinasab, M.; Nabipour, I. First record of microbiomes of sponges collected from the Persian Gulf, using tag pyrosequencing. Front. Microbiol. 2018, 9, 1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpenter, E.J.; Foster, R.A. Marine cyanobacterial symbioses. In Cyanobacteria in Symbiosis; Springer: New York, NY, USA; Boston, MA, USA; Dordrecht, The Netherlands; London, UK; Moscow, Russia, 2002; pp. 11–17. [Google Scholar]
- Gozari, M.; Bahador, N.; Mortazavi, M.S.; Eftekhar, E.; Jassbi, A.R. An “olivomycin A” derivative from a sponge-associated Streptomyces sp. strain SP 85. 3 Biotech 2019, 9, 439. [Google Scholar] [CrossRef]
- Kaasalainen, U.; Fewer, D.P.; Jokela, J.; Wahlsten, M.; Sivonen, K.; Rikkinen, J. Cyanobacteria produce a high variety of hepatotoxic peptides in lichen symbiosis. Proc. Natl. Acad. Sci. USA 2012, 109, 5886–5891. [Google Scholar] [CrossRef] [Green Version]
- Anjum, K.; Abbas, S.Q.; Akhter, N.; Shagufta, B.I.; Shah, S.A.A.; Hassan, S.S.U. Emerging biopharmaceuticals from bioactive peptides derived from marine organisms. Chem. Biol. Drug. Des. 2017, 90, 12–30. [Google Scholar] [CrossRef]
- Zanchett, G.; Oliveira-Filho, E.C. Cyanobacteria and cyanotoxins: From impacts on aquatic ecosystems and human health to anticarcinogenic effects. Toxins 2013, 5, 1896–1917. [Google Scholar] [CrossRef]
- Gutiérrez, M.; Suyama, T.L.; Engene, N.; Wingerd, J.S.; Matainaho, T.; Gerwick, W.H. Apratoxin D, a potent cytotoxic cyclodepsipeptide from papua new guinea collections of the marine cyanobacteria Lyngbya majuscula and Lyngbya sordida. J. Nat. Prod. 2008, 71, 1099–1103. [Google Scholar] [CrossRef]
- Linington, R.G.; Edwards, D.J.; Shuman, C.F.; McPhail, K.L.; Matainaho, T.; Gerwick, W.H. Symplocamide A, a Potent Cytotoxin and Chymotrypsin Inhibitor from the Marine Cyanobacterium Symploca sp. J. Nat. Prod. 2008, 71, 22–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
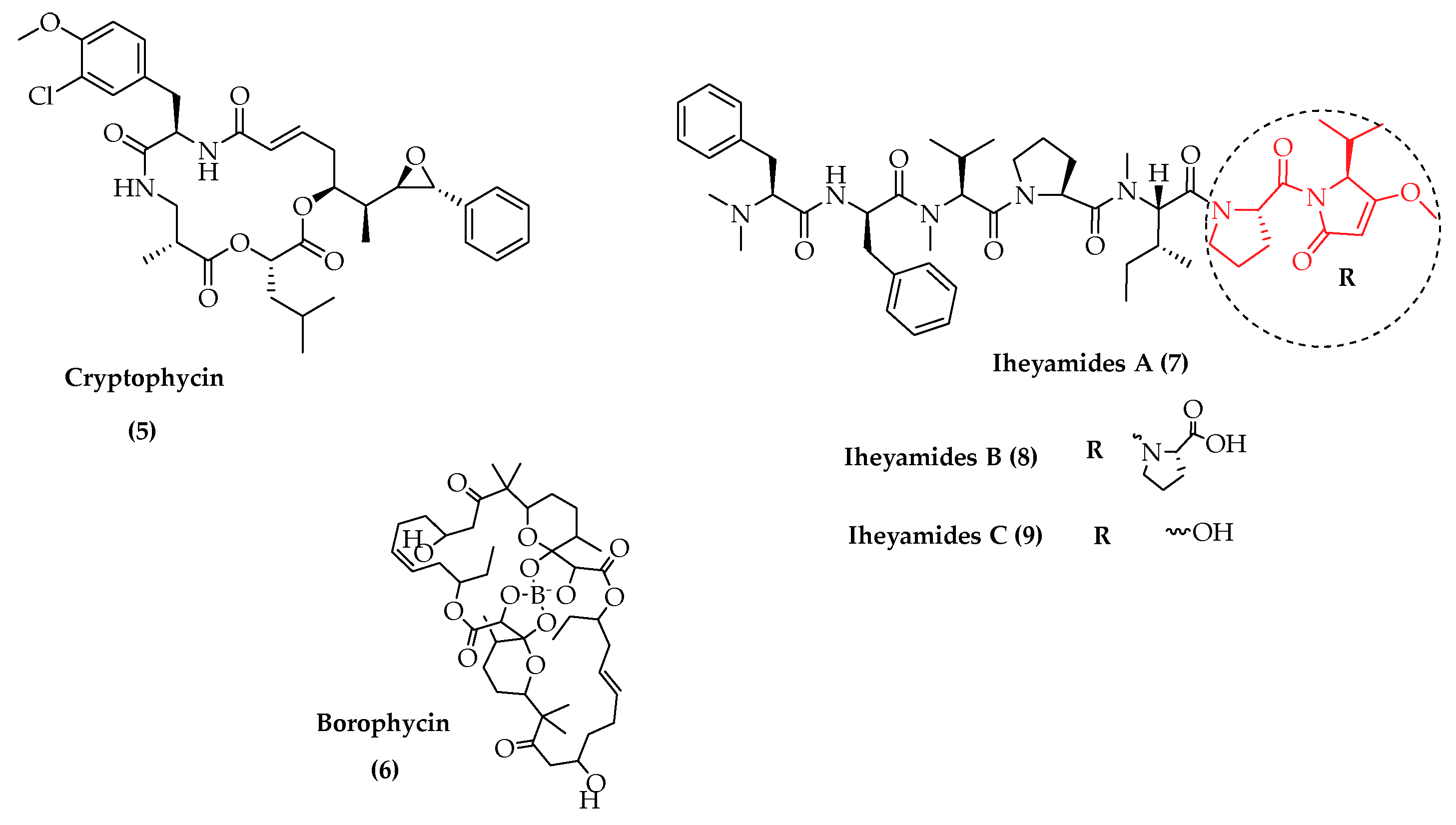
- Kurisawa, N.; Iwasaki, A.; Jeelani, G.; Nozaki, T.; Suenaga, K. Iheyamides A–C, Antitrypanosomal Linear Peptides Isolated from a Marine Dapis sp. Cyanobacterium. J. Nat. Prod. 2020, 83, 1684–1690. [Google Scholar] [CrossRef] [PubMed]
- Demay, J.; Bernard, C.; Reinhardt, A.; Marie, B. Natural products from cyanobacteria: Focus on beneficial activities. Mar. Drugs 2019, 17, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.C.; Monroe, E.A.; Eisman, E.B.; Gerwick, L.; Sherman, D.H.; Gerwick, W.H. The unique mechanistic transformations involved in the biosynthesis of modular natural products from marine cyanobacteria. Nat. Prod. Rep. 2010, 27, 1048–1065. [Google Scholar] [CrossRef]
- Iwasaki, A.; Tadenuma, T.; Sumimoto, S.; Ohshiro, T.; Ozaki, K.; Kobayashi, K.; Teruya, T.; Tomoda, H.; Suenaga, K. Biseokeaniamides A, B, and C, sterol O-acyltransferase inhibitors from an Okeania sp. marine cyanobacterium. J. Nat. Prod. 2017, 80, 1161–1166. [Google Scholar] [CrossRef] [PubMed]
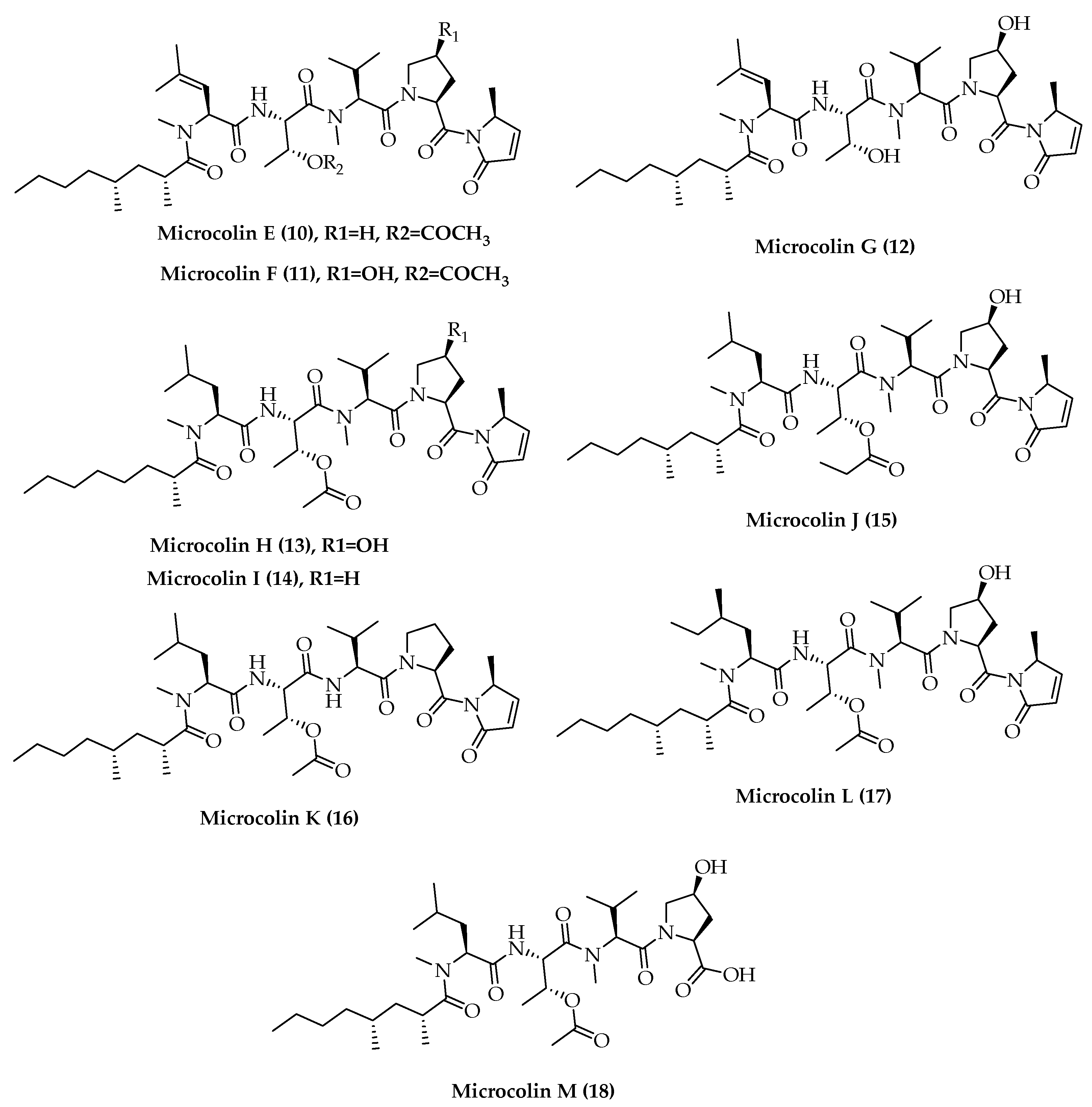
- Yu, H.-B.; Glukhov, E.; Li, Y.; Iwasaki, A.; Gerwick, L.; Dorrestein, P.C.; Jiao, B.-H.; Gerwick, W.H. Cytotoxic Microcolin Lipopeptides from the Marine Cyanobacterium Moorea producens. J. Nat. Prod. 2019, 82, 2608–2619. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, K.; Iwasaki, A.; Sezawa, D.; Fujimura, H.; Nozaki, T.; Saito-Nakano, Y.; Suenaga, K.; Teruya, T. Isolation and Total Synthesis of Mabuniamide, a Lipopeptide from an Okeania sp. Marine Cyanobacterium. J. Nat. Prod. 2019, 82, 2907–2915. [Google Scholar] [CrossRef] [PubMed]
- Uzair, B.; Tabassum, S.; Rasheed, M.; Rehman, S.F. Exploring marine cyanobacteria for lead compounds of pharmaceutical importance. Sci. World J. 2012, 2012, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, M.K. Antimicrobial Activity of Nostoc calcicola (Cyanobacteria) isolated from central India against human pathogens. Asian J. Pharm. 2016, 10. [Google Scholar] [CrossRef]
- Shah, S.; Akhter, N.; Auckloo, B.; Khan, I.; Lu, Y.; Wang, K.; Wu, B.; Guo, Y.-W.J.M.d. Structural diversity, biological properties and applications of natural products from cyanobacteria. A review. Mar. Drugs 2017, 15, 354. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.F.; Ratnayake, R.R.; Meerajini, K.; Wasantha Kumara, K.L. Antioxidant properties in some selected cyanobacteria isolated from fresh water bodies of Sri Lanka. Food Sci. Nutr. 2016, 4, 753–758. [Google Scholar] [CrossRef] [Green Version]
- Sijtsma, L.; De Swaaf, M.E. Biotechnological production and applications of the ω-3 polyunsaturated fatty acid docosahexaenoic acid. Appl. Microbiol. Biotechnol. 2004, 64, 146–153. [Google Scholar] [CrossRef]
- Singh, R.; Parihar, P.; Singh, M.; Bajguz, A.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Uncovering potential applications of cyanobacteria and algal metabolites in biology, agriculture and medicine: Current status and future prospects. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Abed, R.; Dobretsov, S.; Sudesh, K. Application of cyanobacteria in biotechnology. J. Appl. Microbiol. 2008, 106, 1–12. [Google Scholar] [CrossRef]
- Ananyev, G.M.; Skizim, N.J.; Dismukes, G.C. Enhancing biological hydrogen production from cyanobacteria by removal of excreted products. J. Biotechnol. 2012, 162, 97–104. [Google Scholar] [CrossRef]
- Zahra, Z.; Choo, D.H.; Lee, H.; Parveen, A. Cyanobacteria: Review of current potentials and applications. Environments 2020, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- Bajpai, V.; Shukla, S.; Kang, S.-M.; Hwang, S.; Song, X.; Huh, Y.; Han, Y.-K.J.M.d. Developments of cyanobacteria for nano-marine drugs: Relevance of nanoformulations in cancer therapies. Mar. Drugs 2018, 16, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patra, J.K.; Das, G.; Fraceto, L.F.; Campos, E.V.R.; Rodriguez-Torres, M.d.p.; Acosta–Torres, L.S.; Diaz-Torres, L.A.; Grillo, R.; Swamy, M.K.; Sharma, S.; et al. Nano based drug delivery systems: Recent developments and future prospects. J. Nanobiotechnol. 2018, 16, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morone, J.; Lopes, G.; Preto, M.; Vasconcelos, V.; Martins, R. Exploitation of filamentous and picoplanktonic cyanobacteria for cosmetic applications: Potential to improve skin structure and preserve dermal matrix components. Mar. Drugs 2020, 18, 486. [Google Scholar] [CrossRef] [PubMed]
- Morone, J.; Alfeus, A.; Vasconcelos, V.; Martins, R. Revealing the potential of cyanobacteria in cosmetics and cosmeceuticals—A new bioactive approach. Algal Res. 2019, 41, 101541. [Google Scholar] [CrossRef]
- Mourelle, M.; Gómez, C.; Legido, J.J.C. The potential use of marine microalgae and cyanobacteria in cosmetics and thalassotherapy. Cosmetics 2017, 4, 46. [Google Scholar] [CrossRef] [Green Version]
- Burja, A.M.; Banaigs, B.; Abou-Mansour, E.; Burgess, J.G.; Wright, P.C. Marine cyanobacteria—A prolific source of natural products. Tetrahedron 2001, 57, 9347–9377. [Google Scholar] [CrossRef]
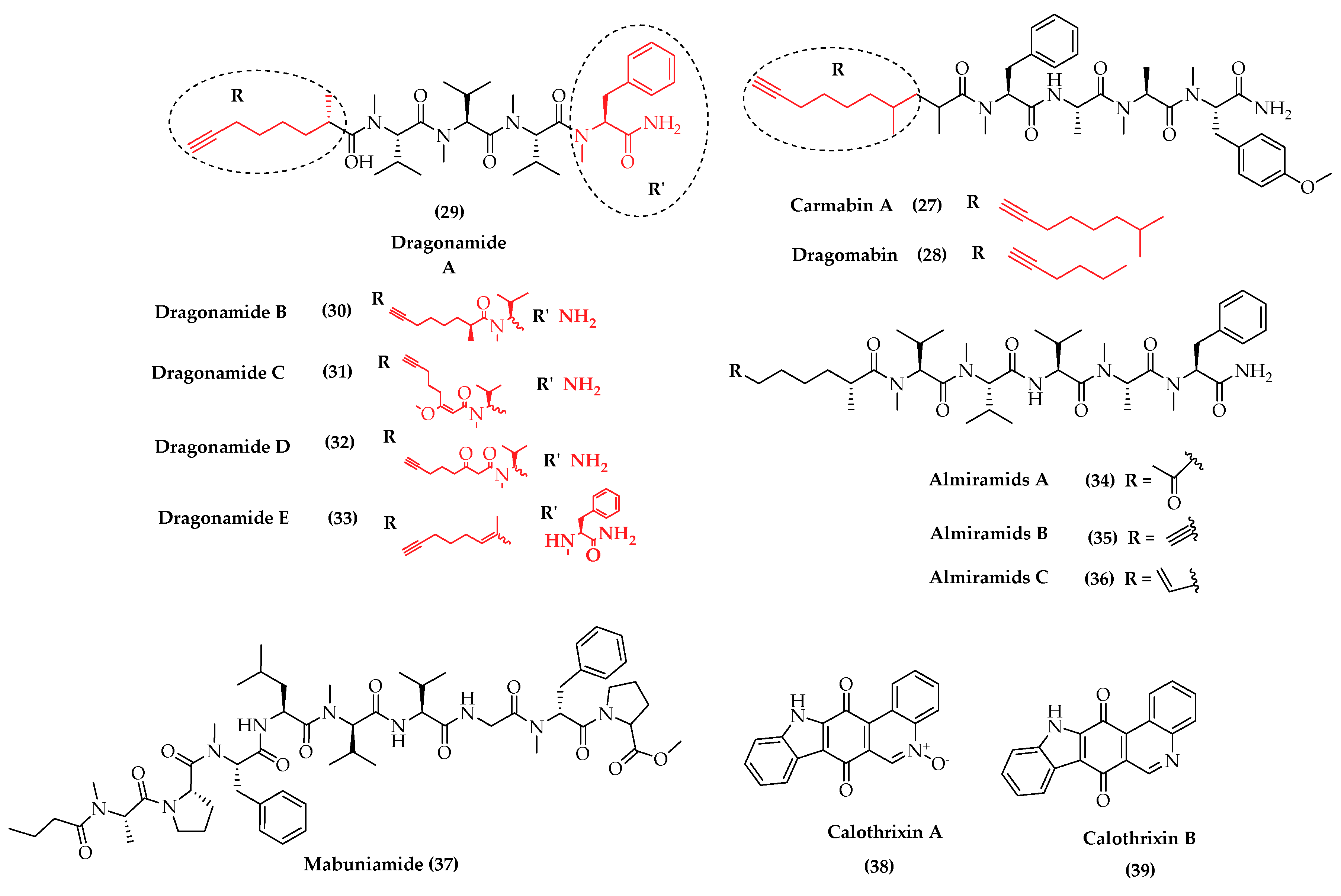
- Gunasekera, S.P.; Ross, C.; Paul, V.J.; Matthew, S.; Luesch, H. Dragonamides C and D, linear lipopeptides from the marine cyanobacterium brown Lyngbya polychroa. J. Nat. Prod. 2008, 71, 887–890. [Google Scholar] [CrossRef]
- Kwan, J.C.; Rocca, J.R.; Abboud, K.A.; Paul, V.J.; Luesch, H. Total structure determination of grassypeptolide, a new marine cyanobacterial cytotoxin. Org. Lett. 2008, 10, 789–792. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Sinha, R.P. Biotechnological and industrial significance of cyanobacterial secondary metabolites. Biotechnol. Adv. 2009, 27, 521–539. [Google Scholar] [CrossRef] [PubMed]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2015, 15, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
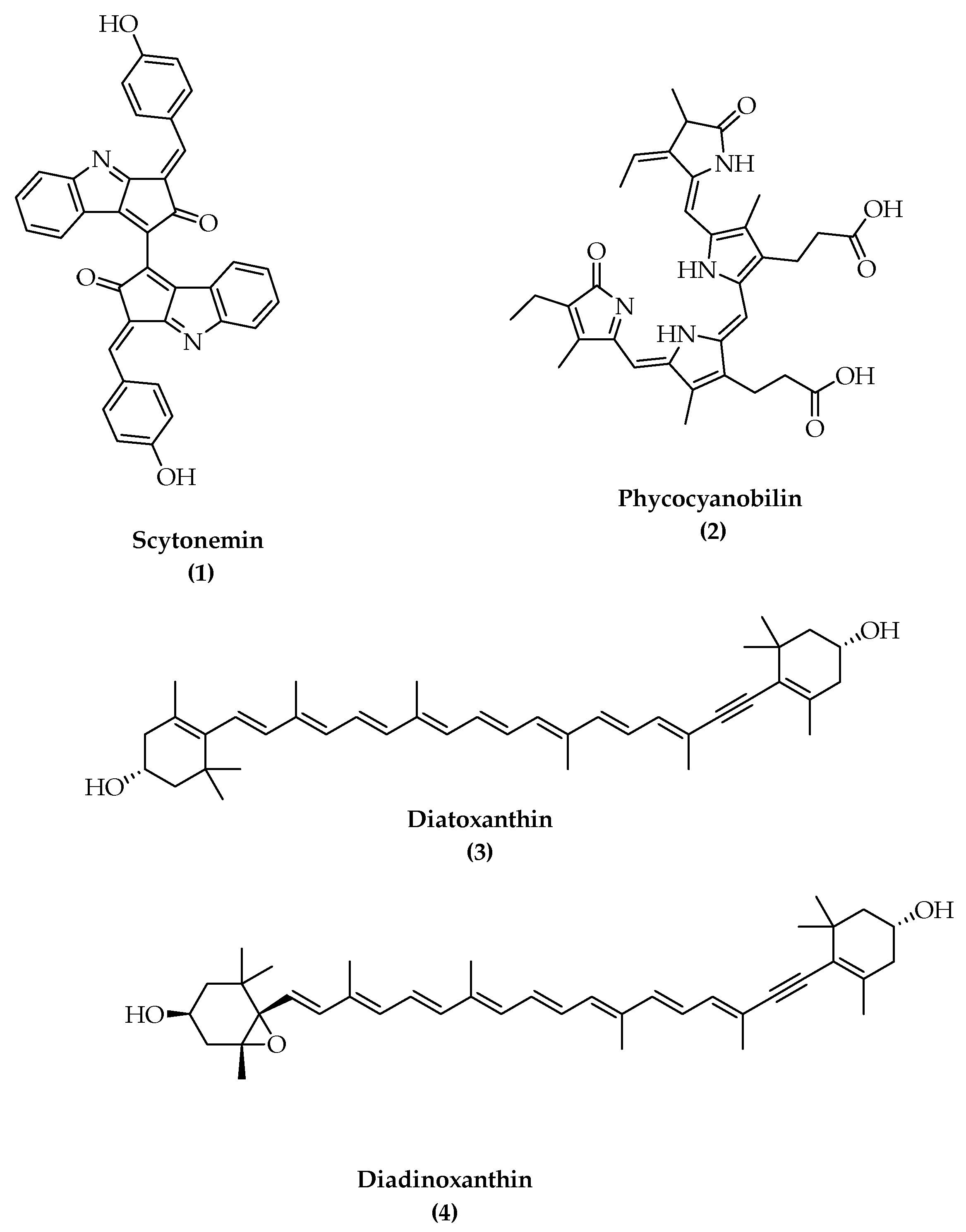
- Pathak, J.; Richa, R.; Sonker, A.S.; Kannaujiya, V.K.; Sinha, R.P. Isolation and partial purification of scytonemin and mycosporine-like amino acids from biological crusts. J. Chem. Pharm. Res. 2015, 7, 362–371. [Google Scholar]
- Rastogi, R.P.; Sinha, R.P.; Incharoensakdi, A. Partial characterization, UV-induction and photoprotective function of sunscreen pigment, scytonemin from Rivularia sp. HKAR-4. Chemosphere 2013, 93, 1874–1878. [Google Scholar] [CrossRef]
- Dillon, J.G.; Castenholz, R.W. The synthesis of the UV-screening pigment, scytonemin, and photosynthetic performance in isolates from closely related natural populations of cyanobacteria (Calothrix sp.). Environ. Microbiol. 2003, 5, 484–491. [Google Scholar] [CrossRef]
- Stevenson, C.S.; Capper, E.A.; Roshak, A.K.; Marquez, B.; Grace, K.; Gerwick, W.H.; Jacobs, R.S.; Marshall, L.A. Scytonemin—A marine natural product inhibitor of kinases key in hyperproliferative inflammatory diseases. Inflamm. Res. 2002, 51, 112–114. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K.; Nazifi, E.; Hirai, Y.; Wada, N.; Matsugo, S.; Sakamoto, T. The cyanobacterial UV-absorbing pigment scytonemin displays radical-scavenging activity. J. Gen. Appl. Microbiol. 2012, 58, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, R.P.; Incharoensakdi, A. Characterization of UV-screening compounds, mycosporine-like amino acids, and scytonemin in the cyanobacterium Lyngbya sp. CU2555. FEMS Microbiol. Ecol. 2014, 87, 244–256. [Google Scholar] [CrossRef] [Green Version]
- Takamatsu, S.; Hodges, T.W.; Rajbhandari, I.; Gerwick, W.H.; Hamann, M.T.; Nagle, D.G. Marine natural products as novel antioxidant prototypes. J. Nat. Prod. 2003, 66, 605–608. [Google Scholar] [CrossRef] [Green Version]
- Suh, H.J.; Lee, H.W.; Jung, J. Mycosporine glycine protects biological systems against photodynamic damage by quenching singlet oxygen with a high efficiency. Photochem. Photobiol. 2003, 78, 109–113. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.-P.-P.; Sulaiman Rahman, H. Antioxidant and oxidative stress: A mutual interplay in age-related diseases. Front. Pharm. 2018, 9, 1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aydaş, S.B.; Ozturk, S.; Aslım, B. Phenylalanine ammonia lyase (PAL) enzyme activity and antioxidant properties of some cyanobacteria isolates. Food Chem. 2013, 136, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, A.; Andrade, M.A.; Menezes, C.; Vilarinho, F.; Dias, E. Antioxidant and cytoprotective properties of cyanobacteria: Potential for biotechnological applications. Toxins 2020, 12, 548. [Google Scholar] [CrossRef] [PubMed]
- Minic, S.L.; Milcic, M.; Stanic-Vucinic, D.; Radibratovic, M.; Sotiroudis, T.G.; Nikolic, M.R.; Velickovic, T.Ć. Phycocyanobilin, a bioactive tetrapyrrolic compound of blue-green alga Spirulina, binds with high affinity and competes with bilirubin for binding on human serum albumin. RSC Adv. 2015, 5, 61787–61798. [Google Scholar] [CrossRef] [Green Version]
- Mishra, P.; Singh, V.P.; Prasad, S.M. Spirulina and its nutritional importance: A possible approach for development of functional food. Biochem. Pharmacol. 2014, 3, e171. [Google Scholar]
- Deo, S.K.; Pandey, R.; Jha, S.K.; Singh, J.; Sodhi, K.S. Spirulina: The single cell protein. Indo Am. J. Pharm. Res. 2014, 4, 221–2217. [Google Scholar]
- Miranda, M.S.; Cintra, R.G.; Barros, S.B.d.M.; Mancini-Filho, J. Antioxidant activity of the microalga Spirulina maxima. Braz. J. Med. Biol. Res. 1998, 31, 1075–1079. [Google Scholar] [CrossRef]
- Al-Dhabi, N.A.; Valan Arasu, M. Quantification of phytochemicals from commercial Spirulina products and their antioxidant activities. J. Evid. Based Complement. Altern. Med. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Yousefi, R.; Mottaghi, A.; Saidpour, A. Spirulina platensis effectively ameliorates anthropometric measurements and obesity-related metabolic disorders in obese or overweight healthy individuals: A randomized controlled trial. Complement. Ther. Med. 2018, 40, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Sommella, E.; Conte, G.M.; Salviati, E.; Pepe, G.; Bertamino, A.; Ostacolo, C.; Sansone, F.; Prete, F.D.; Aquino, R.P.; Campiglia, P. Fast profiling of natural pigments in different Spirulina (Arthrospira platensis) dietary supplements by DI-FT-ICR and evaluation of their antioxidant potential by pre-column DPPH-UHPLC assay. Molecules 2018, 23, 1152. [Google Scholar] [CrossRef] [Green Version]
- Moore, R.E.; Corbett, T.H.; Patterson, G.M.L.; Valeriote, F.A. The search for new antitumor drugs from blue-green algae. Curr. Pharm. Des. 1996, 2, 317–330. [Google Scholar]
- Hemscheidt, T.; Puglisi, M.P.; Larsen, L.K.; Patterson, G.M.L.; Moore, R.E.; Rios, J.L.; Clardy, J. Structure and biosynthesis of borophycin, a new boeseken complex of boric acid from a marine strain of the blue-green alga Nostoc linckia. J. Org. Chem. 1994, 59, 3467–3471. [Google Scholar] [CrossRef]
- Torres, F.A.E.; Passalacqua, T.G.; Velásquez, A.M.A.; de Souza, R.A.; Colepicolo, P.; Graminha, M.A.S. New drugs with antiprotozoal activity from marine algae: A review. Rev. Bras. Farmacogn. 2014, 24, 265–276. [Google Scholar] [CrossRef] [Green Version]
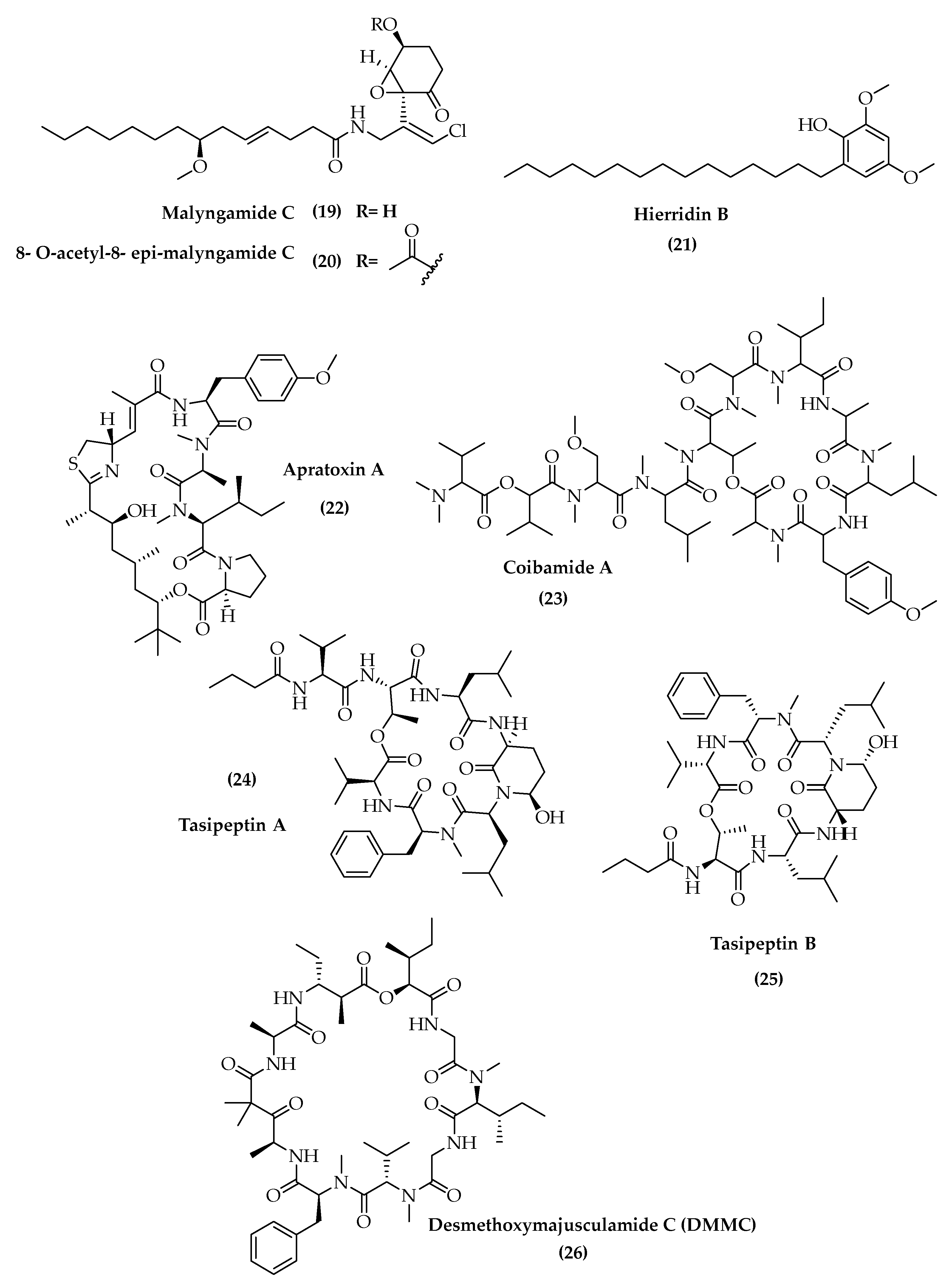
- Gross, H.; McPhail, K.; Goeger, D.; Valeriote, F.; Gerwick, W. Two cytotoxic stereoisomers of malyngamide C, 8- epi-malyngamide C and 8- O-acetyl-8- epi-malyngamide C, from the marine cyanobacterium Lyngbya majuscula. Phytochemistry 2010, 71, 1729–1735. [Google Scholar] [CrossRef] [Green Version]
- Bernardo, P.H.; Chai, C.L.L.; Heath, G.A.; Mahon, P.J.; Smith, G.D.; Waring, P.; Wilkes, B.A. Synthesis, electrochemistry, and bioactivity of the cyanobacterial calothrixins and related quinones. J. Med. Chem. 2004, 47, 4958–4963. [Google Scholar] [CrossRef]
- Leao, P.N.; Costa, M.; Ramos, V.; Pereira, A.R.; Fernandes, V.C.; Domingues, V.F.; Gerwick, W.H.; Vasconcelos, V.M.; Martins, R. Antitumor activity of hierridin B, a cyanobacterial secondary metabolite found in both filamentous and unicellular marine strains. PLoS ONE 2013, 8, e69562. [Google Scholar] [CrossRef] [Green Version]
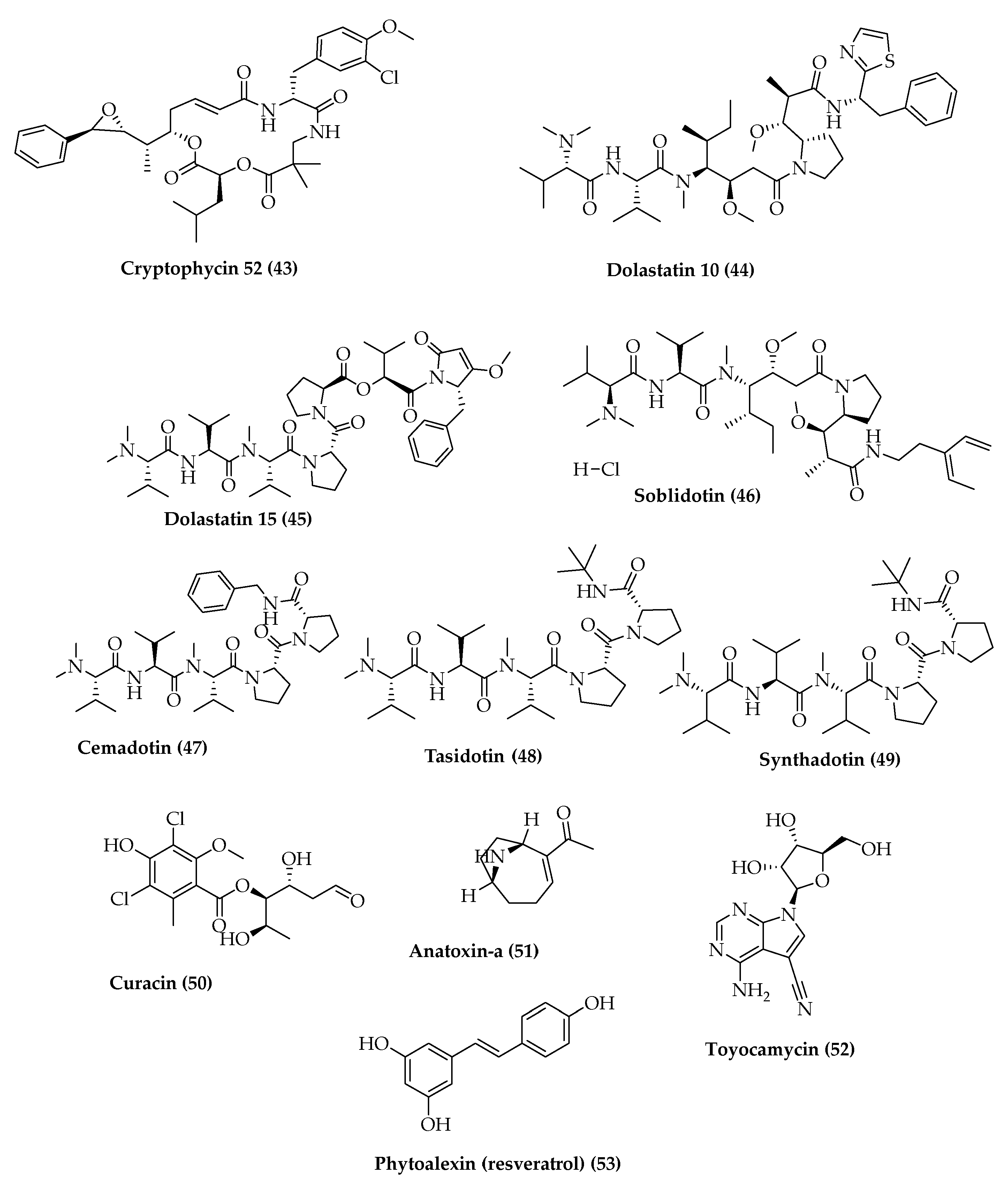
- Luesch, H.; Moore, R.E.; Paul, V.J.; Mooberry, S.L.; Corbett, T.H. Isolation of dolastatin 10 from the marine cyanobacterium Symploca species VP642 and total stereochemistry and biological evaluation of its analogue symplostatin 1. J. Nat. Prod. 2001, 64, 907–910. [Google Scholar] [CrossRef] [PubMed]
- McPhail, K.L.; Medina, R.A.; Gerwick, W.H.; Goeger, D.E.; Capeon, T.L. Isolation, Purification, and Structure Elucidation of the Antiproliferative Compound Coibamide A. U.S. Patent 20120028905-A1, 2 February 2012. [Google Scholar]
- Gerwick, W.H.; Tan, L.T.; Sitachitta, N. Nitrogen-containing metabolites from marine cyanobacteria. Alkaloids Chem. Biol. 2001, 57, 75–184. [Google Scholar] [PubMed]
- Williams, P.G.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J. Tasipeptins A and B: New cytotoxic depsipeptides from the marine cyanobacterium Symploca sp. J. Nat. Prod. 2003, 66, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Simmons, T.L.; Nogle, L.M.; Media, J.; Valeriote, F.A.; Mooberry, S.L.; Gerwick, W.H. Desmethoxymajusculamide C, a cyanobacterial depsipeptide with potent cytotoxicity in both cyclic and ring-opened forms. J. Nat. Prod. 2009, 72, 1011–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McPhail, K.L.; Correa, J.; Linington, R.G.; González, J.; Ortega-Barría, E.; Capson, T.L.; Gerwick, W.H. Antimalarial linear lipopeptides from a Panamanian strain of the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2007, 70, 984–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balunas, M.J.; Linington, R.G.; Tidgewell, K.; Fenner, A.M.; Urena, L.-D.; Togna, G.D.; Kyle, D.E.; Gerwick, W.H. Dragonamide E, a modified linear lipopeptide from Lyngbya majuscula with antileishmanial activity. J. Nat. Prod. 2010, 73, 60–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, L.M.; Lopez, D.; Vesely, B.A.; Della Togna, G.; Gerwick, W.H.; Kyle, D.E.; Linington, R.G. Almiramides A− C: Discovery and development of a new class of leishmaniasis lead compounds. J. Med. Chem. 2010, 53, 4187–4197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rickards, R.W.; Rothschild, J.M.; Willis, A.C.; de Chazal, N.M.; Kirk, J.; Kirk, K.; Saliba, K.J.; Smith, G.D. Calothrixins A and B, novel pentacyclic metabolites from Calothrix cyanobacteria with potent activity against malaria parasites and human cancer cells. Tetrahedron 1999, 55, 13513–13520. [Google Scholar] [CrossRef]
- Lee, J.-B.; Hayashi, T.; Hayashi, K.; Sankawa, U.; Maeda, M.; Nemoto, T.; Nakanishi, H. Further purification and structural analysis of calcium spirulan from Spirulina platensis. J. Nat. Prod. 1998, 61, 1101–1104. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Hayashi, K.; Maeda, M.; Kojima, I. Calcium spirulan, an inhibitor of enveloped virus replication, from a blue-green alga Spirulina platensis. J. Nat. Prod. 1996, 59, 83–87. [Google Scholar] [CrossRef]
- Hayashi, K.; Hayashi, T.; Kojima, I. A Natural sulfated polysaccharide, calcium spirulan, isolated from Spirulina platensis: In vitro and ex vivo evaluation of anti-herpes simplex virus and anti-human immunodeficiency virus activities. AIDS Res. Hum. Retrovir. 1996, 12, 1463–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rechter, S.; König, T.; Auerochs, S.; Thulke, S.; Walter, H.; Dörnenburg, H.; Walter, C.; Marschall, M. Antiviral activity of Arthrospira-derived spirulan-like substances. Antivir. Res. 2006, 72, 197–206. [Google Scholar] [CrossRef]
- Kanekiyo, K.; Lee, J.-B.; Hayashi, K.; Takenaka, H.; Hayakawa, Y.; Endo, S.; Hayashi, T. Isolation of an antiviral polysaccharide, nostoflan, from a terrestrial cyanobacterium, Nostoc flagelliforme. J. Nat. Prod. 2005, 68, 1037–1041. [Google Scholar] [CrossRef]
- Lokhande, K.B.; Apte, G.R.; Shrivastava, A.; Singh, A.; Pal, J.K.; Swamy, K.V.; Gupta, R.K. Sensing the interactions between carbohydrate-binding agents and N-linked glycans of SARS-CoV-2 spike glycoprotein using molecular docking and simulation studies. J. Biomol. Struct. Dyn. 2020, 1–19. [Google Scholar] [CrossRef]
- Pendyala, B.; Patras, A.; Dash, C. Phycobilins as potent food bioactive broad-spectrum inhibitor compounds against Mpro and PLpro of SARS-CoV-2 and other coronaviruses: A preliminary Study. bioRxiv 2020. [Google Scholar] [CrossRef]
- Naidoo, D.; Roy, A.; Kar, P.; Mutanda, T.; Anandraj, A. Cyanobacterial metabolites as promising drug leads against the Mpro and PLpro of SARS-CoV-2: An in silico analysis. J. Biomol. Struct. Dyn. 2020, 1–13. [Google Scholar] [CrossRef]
- Schwarzer, D.; Finking, R.; Marahiel, M.A. Nonribosomal peptides: From genes to products. Nat. Prod. Rep. 2003, 20, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Lichota, A.; Gwozdzinski, K. Anticancer Activity of Natural Compounds from Plant and Marine Environment. Int. J. Mol. Sci. 2018, 19, 3533. [Google Scholar] [CrossRef] [Green Version]
- Gheda, S.F.; Ismail, G.A. Natural products from some soil cyanobacterial extracts with potent antimicrobial, antioxidant and cytotoxic activities. Anais Acad. Bras. Ciênc. 2019, 92, e20190934. [Google Scholar] [CrossRef] [PubMed]
- Jordan, M.A.; Wilson, L. Microtubules and actin filaments: Dynamic targets for cancer chemotherapy. Curr. Opin. Cell Biol. 1998, 10, 123–130. [Google Scholar] [CrossRef]
- Dixit, R.B.; Suseela, M.R. Cyanobacteria: Potential candidates for drug discovery. Antonie Leeuwenhoek 2013, 103, 947–961. [Google Scholar] [CrossRef]
- Tan, L.T. Bioactive natural products from marine cyanobacteria for drug discovery. Phytochemistry 2007, 68, 954–979. [Google Scholar] [CrossRef]
- Drugbank. Available online: https://www.drugbank.ca/drugs/DB12730 (accessed on 20 December 2020).
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT00005579 (accessed on 20 December 2020).
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT00003626 (accessed on 20 December 2020).
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT00003914 (accessed on 20 December 2020).
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT00003557 (accessed on 20 December 2020).
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT00003778 (accessed on 20 December 2020).
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT00003693 (accessed on 20 December 2020).
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT00003677 (accessed on 20 December 2020).
- Liu, L.; Rein, K.S. New peptides isolated from Lyngbya species: A review. Mar. Drugs 2010, 8, 1817–1837. [Google Scholar] [CrossRef] [Green Version]
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT00068211 (accessed on 20 December 2020).
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT00082134 (accessed on 20 December 2020).
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT00078455 (accessed on 21 December 2020).
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT00431223 (accessed on 21 December 2020).
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT00546052 (accessed on 21 December 2020).
- Drugbank. Available online: http://www.drugbank.ca/drugs/DB13916 (accessed on 21 December 2020).
- Patterson, G.M.L.; Bolis, C.M. Fungal cellwall polysaccharides elicit an antifungal secondary metabolite (phytoalexin) in the cyanobacterium scytonema ocelutum 2. J. Phycol. 1997, 33, 54–60. [Google Scholar] [CrossRef]
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT01677611 (accessed on 21 December 2020).
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT00064220 (accessed on 21 December 2020).
- Wu, Q.; Liu, L.; Miron, A.; Klímová, B.; Wan, D.; Kuča, K. The antioxidant, immunomodulatory, and anti-inflammatory activities of Spirulina: An overview. Arch. Toxicol. 2016, 90, 1817–1840. [Google Scholar] [CrossRef]
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT02886676 (accessed on 21 December 2020).
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT02817620 (accessed on 21 December 2020).
- Aráoz, R.; Molgó, J.; de Marsac, N.T. Neurotoxic cyanobacterial toxins. Toxicon 2010, 56, 813–828. [Google Scholar] [CrossRef] [PubMed]
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT02916966 (accessed on 21 December 2020).
- Raja, R.; Hemaiswarya, S.; Ganesan, V.; Carvalho, I.S. Recent developments in therapeutic applications of Cyanobacteria. Crit. Rev. Microbiol. 2016, 42, 394–405. [Google Scholar] [CrossRef]
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT02928042 (accessed on 21 December 2020).
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT03219931 (accessed on 21 December 2020).
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT02241161 (accessed on 21 December 2020).
- Clinicaltrials. Available online: https://www.clinicaltrials.gov/NCT03004196 (accessed on 21 December 2020).
- Lotfi, H.; Sheervalilou, R.; Zarghami, N. An update of the recombinant protein expression systems of Cyanovirin-N and challenges of preclinical development. BioImpacts 2018, 8, 139. [Google Scholar] [CrossRef] [Green Version]
- Beltran, E.C.; Neilan, B.A.J.A.; Microbiology, E. Geographical segregation of the neurotoxin-producing cyanobacterium Anabaena circinalis. Appl. Environ. Microbiol. 2000, 66, 4468–4474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sierosławska, A.; Rymuszka, A.; Kalinowska, R.; Skowroński, T.; Bownik, A.; Pawlik-Skowrońska, B. Toxicity of cyanobacterial bloom in the eutrophic dam reservoir (Southeast Poland). Environ. Toxicol. Chem. Int. J. 2010, 29, 556–560. [Google Scholar] [CrossRef]
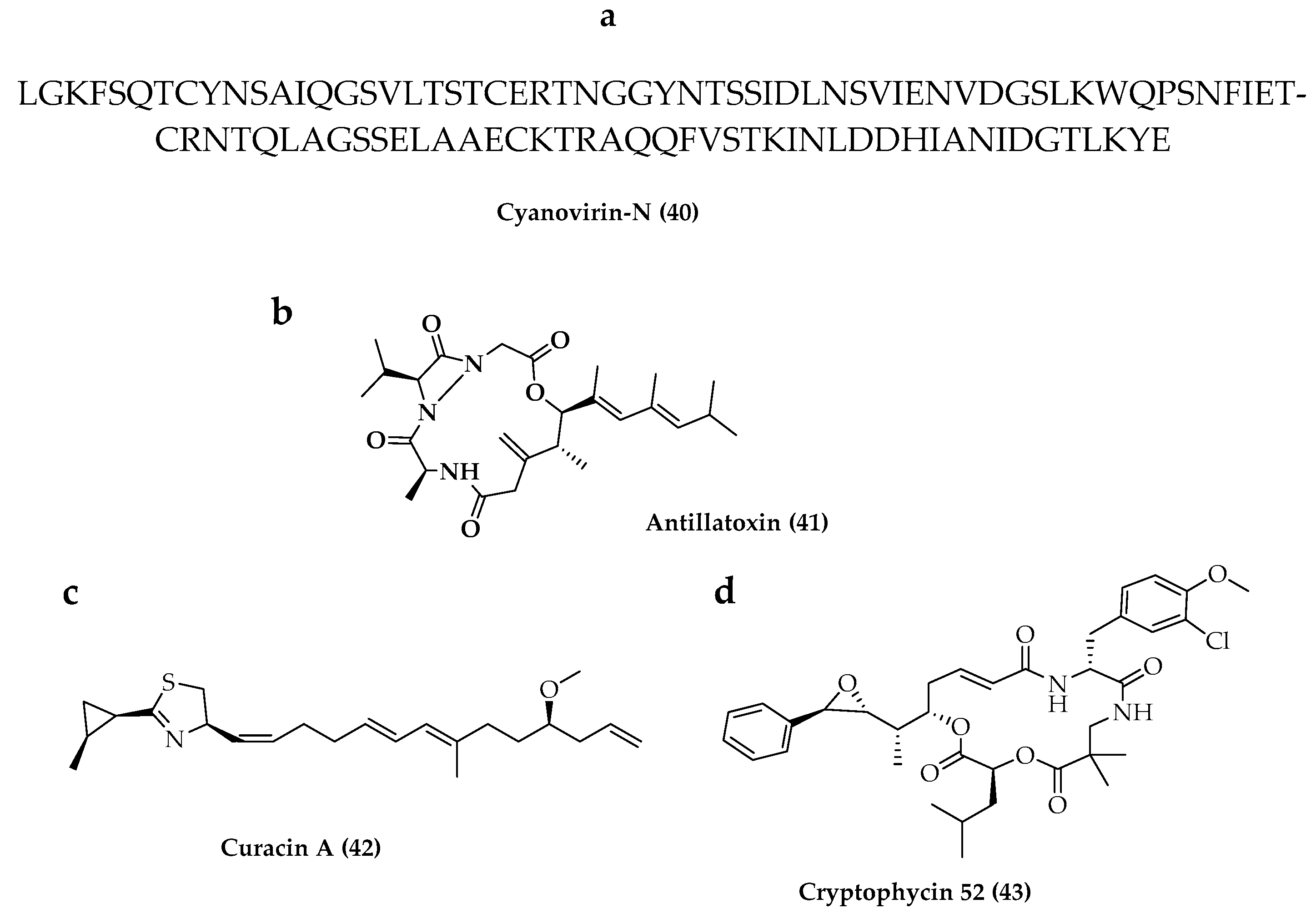
- Boyd, M.R.; Gustafson, K.R.; McMahon, J.B.; Shoemaker, R.H.; O’Keefe, B.R.; Mori, T.; Gulakowski, R.J.; Wu, L.; Rivera, M.I.; Laurencot, C.M.; et al. Discovery of cyanovirin-N, a novel human immunodeficiency virus-inactivating protein that binds viral surface envelope glycoprotein gp120: Potential applications to microbicide development. Antivir. Res. 1997, 41, 1521–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gustafson, K.R.; Sowder, R.C., II; Henderson, L.E.; Cardellina, J.H., II; McMahon, J.B.; Rajamani, U.; Pannell, L.K.; Boyd, M.R. Isolation, primary sequence determination, and disulfide bond structure of cyanovirin-n, an anti-hiv (human immunodeficiency virus) protein from the cyanobacterium nostoc ellipsosporum. Biochem. Biophys. Res. Commun. 1997, 238, 223–228. [Google Scholar] [CrossRef]
- Buffa, V.; Stieh, D.; Mamhood, N.; Hu, Q.; Fletcher, P.; Shattock, R.J. Cyanovirin-N potently inhibits human immunodeficiency virus type 1infection in cellular and cervical explant models. J. Gen. Virol. 2009, 90, 234–243. [Google Scholar] [CrossRef] [Green Version]
- Stewart, J.B.; Bornemann, V.; JIAN, L.; Moore, R.E.; Caplan, F.R.; Karuso, H.; Larsen, L.K.; Patterson, G.M. Cytotoxic, fungicidal nucleosides from blue green algae belonging to the Scytonemataceae. J. Antibiot. 1988, 41, 1048–1056. [Google Scholar] [CrossRef] [Green Version]
- Bastea, L.I.; Hollant, L.M.; Döppler, H.R.; Reid, E.M.; Storz, P. Sangivamycin and its derivatives inhibit Haspin-Histone H3-survivin signaling and induce pancreatic cancer cell death. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Stockwin, L.H.; Sherry, X.Y.; Stotler, H.; Hollingshead, M.G.; Newton, D.L. ARC (NSC 188491) has identical activity to Sangivamycin (NSC 65346) including inhibition of both P-TEFb and PKC. BMC Cancer 2009, 9, 63. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Fewer, D.P.; Sivonen, K. Genome mining demonstrates the widespread occurrence of gene clusters encoding bacteriocins in cyanobacteria. PLoS ONE 2011, 6, e22384. [Google Scholar] [CrossRef] [Green Version]
- Desriac, F.; Defer, D.; Bourgougnon, N.; Brillet, B.; Le Chevalier, P.; Fleury, Y. Bacteriocin as weapons in the marine animal-associated bacteria warfare: Inventory and potential applications as an aquaculture probiotic. Mar. Drugs 2010, 8, 1153–1177. [Google Scholar] [CrossRef] [Green Version]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef]
- Ishii, K.; Katoh, T.; Okuwaki, Y.; Hayashi, O. Influence of dietary Spirulina platensis on IgA level in human saliva. J. Kagawa Nutr. Univ. 1999, 30, 27–33. [Google Scholar]
- Tantong, S.; Incharoensakdi, A.; Sirikantaramas, S.; Lindblad, P. Purification, potential of synechocystis PCC 6803 as a novel cyanobacterial chassis for heterologous expression of enzymes in the trans-resveratrol biosynthetic pathway. Protein Expr. Purif. 2016, 121, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Vallianou, N.G.; Evangelopoulos, A.; Kazazis, C. Resveratrol and diabetes. Rev. Diabetic Stud. 2013, 10, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ott, P.A.; Pavlick, A.C.; Johnson, D.B.; Hart, L.L.; Infante, J.R.; Luke, J.J.; Lutzky, J.; Rothschild, N.; Spitler, L.; Cowey, C.L. A phase 2 study of glembatumumab vedotin (GV), an antibody-drug conjugate (ADC) targeting gpNMB, in advanced melanoma. European Society for Medical Oncology. Ann. Oncol. 2016, 27, vi393. [Google Scholar] [CrossRef] [Green Version]
- Deng, C.; Pan, B.; O’Connor, O.A. Brentuximab vedotin. Clin. Cancer Res. 2013, 19, 22–27. [Google Scholar] [CrossRef] [Green Version]
- Kulasooriya, S.A. Cyanobacteria: Pioneers of planet earth. Ceylon J. Sci. (Bio. Sci.) 2011, 40, 71–88. [Google Scholar] [CrossRef] [Green Version]
- Kulasooriya, S.A.; Magana-Arachchi, D.N. Nitrogen fixing cyanobacteria: Their diversity, ecology and utilisation with special reference to rice cultivation. J. Nat. Sci. Foundat. Sri Lanka 2016, 44, 111–128. [Google Scholar] [CrossRef] [Green Version]
- Vijayakumar, S.; Menakha, M. Pharmaceutical applications of cyanobacteria—A review. J. Acute Med. 2015, 5, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Kumar, J.; Singh, D.; Tyagi, M.B.; Kumar, A. Cyanobacteria: Applications in biotechnology. In Cyanobacteria; Elsevier: Amsterdam, The Netherlands, 2019; pp. 327–346. [Google Scholar]
- Angermayr, S.A.; Hellingwerf, K.J.; Lindblad, P.; de Mattos, M.J.T. Energy biotechnology with cyanobacteria. Curr. Opin. Biotechnol. 2009, 20, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- De Farias Silva, C.E.; Bertucco, A. Bioethanol from microalgae and cyanobacteria: A review and technological outlook. Process Biochem. 2016, 51, 1833–1842. [Google Scholar] [CrossRef]
- Deviram, G.; Mathimani, T.; Anto, S.; Ahamed, T.S.; Ananth, D.A.; Pugazhendhi, A. Applications of microalgal and cyanobacterial biomass on a way to safe, cleaner and a sustainable environment. J. Clean. Prod. 2020, 253, 119770. [Google Scholar] [CrossRef]
- Mirsasaani, S.S.; Hemati, M.; Dehkord, E.S.; Yazdi, G.T.; Poshtiri, D.A. Nanotechnology and nanobiomaterials in dentistry. In Nanobiomaterials in Clinical Dentistry; Elsevier: Amsterdam, The Netherlands, 2019; pp. 19–37. [Google Scholar]
- Nair, H.B.; Sung, B.; Yadav, V.R.; Kannappan, R.; Chaturvedi, M.M.; Aggarwal, B.B. Delivery of antiinflammatory nutraceuticals by nanoparticles for the prevention and treatment of cancer. Biochem. Pharmacol. 2010, 80, 1833–1843. [Google Scholar] [CrossRef] [Green Version]
- Mohanpuria, P.; Rana, N.K.; Yadav, S.K. Biosynthesis of nanoparticles: Technological concepts and future applications. J. Nanopart. Res. 2008, 10, 507–517. [Google Scholar] [CrossRef]
- Elghazawy, N.H.; Hefnawy, A.; Sedky, N.K.; El-Sherbiny, I.M.; Arafa, R.K. Preparation and nanoformulation of new quinolone scaffold-based anticancer agents: Enhancing solubility for better cellular delivery. Eur. J. Pharm. Sci. 2017, 105, 203–211. [Google Scholar] [CrossRef]
- Kubik, T.; Bogunia-Kubik, K.; Sugisaka, M. Nanotechnology on duty in medical applications. Curr. Pharm. Biotechnol. 2005, 6, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, S.R.; Santhiyagu, P.; Ramasamy, R.; Arivalagan, P.; Kumar, G.; Ethiraj, K.; Ramaswamy, B.R. Seaweeds: A resource for marine bionanotechnology. Enzyme Microb. Technol. 2016, 95, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Manivasagan, P.; Bharathiraja, S.; Moorthy, M.S.; Oh, Y.-O.; Seo, H.; Oh, J. Marine biopolymer-based nanomaterials as a novel platform for theranostic applications. Polym. Rev. 2017, 57, 631–667. [Google Scholar] [CrossRef]
- Cardoso, M.J.; Costa, R.R.; Mano, J.F. Marine origin polysaccharides in drug delivery systems. Mar. Drugs 2016, 14, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lengke, M.F.; Fleet, M.E.; Southam, G. Biosynthesis of silver nanoparticles by filamentous cyanobacteria from a silver (I) nitrate complex. Langmuir 2007, 23, 2694–2699. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.; Berthold, D.; Puranik, P.; Gantar, M. Screening of cyanobacteria and microalgae for their ability to synthesize silver nanoparticles with antibacterial activity. Biotechnol. Rep. 2015, 5, 112–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MubarakAli, D.; Sasikala, M.; Gunasekaran, M.; Thajuddin, N. Biosynthesis and characterization of silver nanoparticles using marine cyanobacterium, Oscillatoria willei NTDM01. Dig. J. Nanomater. Biostruct. 2011, 6, 385–390. [Google Scholar]
- Brayner, R.; Barberousse, H.; Hemadi, M.; Djedjat, C.; Yéprémian, C.; Coradin, T.; Livage, J.; Fiévet, F.; Couté, A. Cyanobacteria as bioreactors for the synthesis of Au, Ag, Pd, and Pt nanoparticles via an enzyme-mediated route. J. Nanosci. Nanotechnol. 2007, 7, 2696–2708. [Google Scholar] [CrossRef]
- Shanmugam, R.; Chelladurai, M.; Paulmar, K.; Vanaja, M.; Gnanajobitha, G.; Gurusamy, A. Intracellular and extracellular biosynthesis of silver nanoparticles by using marine bacteria Vibrio alginolyticus. Nanosci. Nanotechnol. 2013, 3, 21–25. [Google Scholar]
- Delneuville, C.; Danloy, E.P.; Wang, L.; Su, B.-L. Single cyanobacteria@ silica porous microcapsules via a sol–gel layer by layer for heavy-metal remediation. J. Sol. Gel. Sci. Technol. 2019, 89, 244–254. [Google Scholar] [CrossRef]
- Zhang, Y.S.; Zhang, Y.-N.; Zhang, W. Cancer-on-a-chip systems at the frontier of nanomedicine. Drug Dis. Today 2017, 22, 1392–1399. [Google Scholar] [CrossRef] [PubMed]
- Meléndez-Martínez, A.J.; Stinco, C.M.; Mapelli-Brahm, P. Skin carotenoids in public health and nutricosmetics: The emerging roles and applications of the UV radiation-absorbing colourless carotenoids phytoene and phytofluene. Nutrients 2019, 11, 1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halpern, A.C.; Kopp, L.J. Awareness, knowledge and attitudes to non-melanoma skin cancer and actinic keratosis among the general public. Int. J. Dermatol. 2005, 44, 107–111. [Google Scholar] [CrossRef]
- Conde, F.R.; Churio, M.S.; Previtali, C.M. The photoprotector mechanism of mycosporine-like amino acids. Excited-state properties and photostability of porphyra-334 in aqueous solution. J. Photochem. Photobiol. B Biol. 2000, 56, 139–144. [Google Scholar] [CrossRef]
- Dunlap, W.C.; Chalker, B.E.; Bandaranayake, W.M.; Wu Won, J.J. Nature’s sunscreen from the Great Barrier Reef, Australia. Int. J. Cosmet. Sci. 1998, 20, 41–51. [Google Scholar] [CrossRef]
- Bhatia, S.; Garg, A.; Sharma, K.; Kumar, S.; Sharma, A.; Purohit, A.P. Mycosporine and mycosporine-like amino acids: A paramount tool against ultra violet irradiation. Pharmacogn. Rev. 2011, 5, 138. [Google Scholar] [CrossRef] [Green Version]
- Li, E.P.H.; Min, H.J.; Belk, R.W. Skin lightening and beauty in four Asian cultures. ACR N. Am. Adv. 2008, 35, 444–449. [Google Scholar]
- Joshi, L.S.; Pawar, H.A. Herbal cosmetics and cosmeceuticals: An overview. Nat. Prod. Chem. Res. 2015, 3, 170. [Google Scholar] [CrossRef]
- Chanchal, D.; Swarnlata, S. Novel approaches in herbal cosmetics. J. Cosmet. Dermatol. 2008, 7, 89–95. [Google Scholar] [CrossRef]
- Dreno, B.; Araviiskaia, E.; Berardesca, E.; Bieber, T.; Hawk, J.; Sanchez-Viera, M.; Wolkenstein, P. The science of dermocosmetics and its role in dermatology. J. Eur. Acad. Dermatol. Venereol. 2014, 28, 1409–1417. [Google Scholar] [CrossRef]
- Datta, H.S.; Paramesh, R. Trends in aging and skin care: Ayurvedic concepts. J. Ayurveda Integr. Med. 2010, 1, 110. [Google Scholar] [CrossRef] [Green Version]
- Ou, H.-L.; Schumacher, B. DNA damage responses and p53 in the aging process. Blood 2018, 131, 488–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dizdaroglu, M.; Jaruga, P. Mechanisms of free radical-induced damage to DNA. Free Radic. Res. 2012, 46, 382–419. [Google Scholar] [CrossRef] [PubMed]
- Liochev, S.I. Reactive oxygen species and the free radical theory of aging. Free Radic. Biol. Med. 2013, 60, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, M.; Shabbir, A.; Wojcikowski, K.; Wohlmuth, H.; Gobe, G.C. The antioxidant effects of Radix Astragali (Astragalus membranaceus and related species) in protecting tissues from injury and disease. Curr. Drug Targets 2016, 17, 1331–1340. [Google Scholar] [CrossRef]
- Fougère, B.; Boulanger, E.; Nourhashémi, F.; Guyonnet, S.; Cesari, M. RETRACTED: Chronic Inflammation: Accelerator of Biological Aging. J. Gerontol. A 2017, 72, 1218–1225. [Google Scholar] [CrossRef] [Green Version]
- Maru, G.B.; Gandhi, K.; Ramchandani, A.; Kumar, G. The role of inflammation in skin cancer. In Inflammation and Cancer; Springer: New York, NY, USA; Boston, MA, USA; Dordrecht, The Netherlands; London, UK; Moscow, Russia, 2014; pp. 437–469. [Google Scholar]
- Radice, M.; Manfredini, S.; Ziosi, P.; Dissette, V.; Buso, P.; Fallacara, A.; Vertuani, S. Herbal extracts, lichens and biomolecules as natural photo-protection alternatives to synthetic UV filters. A systematic review. Fitoterapia 2016, 114, 144–162. [Google Scholar] [CrossRef]
- Derikvand, P.; Llewellyn, C.A.; Purton, S. Cyanobacterial metabolites as a source of sunscreens and moisturizers: A comparison with current synthetic compounds. Eur. J. Phycol. 2017, 52, 43–56. [Google Scholar] [CrossRef]
- Gunes, S.; Tamburaci, S.; Dalay, M.C.; Deliloglu Gurhan, I. in vitro evaluation of Spirulina platensis extract incorporated skin cream with its wound healing and antioxidant activities. Pharm. Biol. 2017, 55, 1824–1832. [Google Scholar] [CrossRef] [Green Version]
- Saha, R. Cosmeceuticals and herbal drugs: Practical uses. Int. J. Pharm. Sci. Res. 2012, 3, 59–65. [Google Scholar]
- Hashtroudi, M.S.; Shariatmadari, Z.; Riahi, H.; Ghassempour, A. Analysis of Anabaena vaginicola and Nostoc calcicola from Northern Iran, as rich sources of major carotenoids. Food Chem. 2013, 136, 1148–1153. [Google Scholar] [CrossRef]
- Hamed, I. The evolution and versatility of microalgal biotechnology: A review. Compr. Rev. Food Sci. F 2016, 15, 1104–1123. [Google Scholar] [CrossRef]
- Yabuta, Y.; Hashimoto, E.; Takeuchi, T.; Sakaki, S.; Yamaguchi, Y.; Takenaka, H.; Watanabe, F. Characterization of a hot water extract of an edible cyanobacterium Nostochopsis sp. for use as an ingredient in cosmetics. Food Sci. Technol. Res. 2014, 20, 505–507. [Google Scholar] [CrossRef] [Green Version]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzyme Inhibit. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef] [Green Version]
- Ngatu, N.R.; Okajima, M.K.; Yokogawa, M.; Hirota, R.; Eitoku, M.; Muzembo, B.A.; Dumavibhat, N.; Takaishi, M.; Sano, S.; Kaneko, T. Anti-inflammatory effects of sacran, a novel polysaccharide from Aphanothece sacrum, on 2, 4, 6-trinitrochlorobenzene–induced allergic dermatitis in vivo. Ann. Allergy Asthma Immunol. 2012, 108, 117–122. [Google Scholar] [CrossRef]
- Okamoto, S.; Iwasaki, A.; Ohno, O.; Suenaga, K. Isolation and structure of kurahyne B and total synthesis of the kurahynes. J. Nat. Prod. 2015, 78, 2719–2725. [Google Scholar] [CrossRef]
- Hamada, Y.; Shioiri, T. Recent progress of the synthetic studies of biologically active marine cyclic peptides and depsipeptides. Chem. Rev. 2005, 105, 4441–4482. [Google Scholar] [CrossRef]
- Shinomiya, S.; Iwasaki, A.; Ohno, O.; Suenaga, K.J.P. Total synthesis and stereochemical determination of yoshinone A. Phytochemistry 2016, 132, 109–114. [Google Scholar] [CrossRef]
- Ogawa, H.; Iwasaki, A.; Sumimoto, S.; Iwatsuki, M.; Ishiyama, A.; Hokari, R.; Otoguro, K.; Ōmura, S.; Suenaga, K. Isolation and total synthesis of hoshinolactam, an antitrypanosomal lactam from a marine cyanobacterium. Org. Lett. 2017, 19, 890–893. [Google Scholar] [CrossRef]
- Iwasaki, A.; Teruya, T.; Suenaga, K. Isolation and structure of koshikalide, a 14-membered macrolide from the marine cyanobacterium Lyngbya sp. Tetrahedron Lett. 2010, 51, 959–960. [Google Scholar] [CrossRef]
- Chen, J.; Forsyth, C.J. Total synthesis of the marine cyanobacterial cyclodepsipeptide apratoxin A. Proc. Natl. Acad. Sci. USA 2004, 101, 12067–12072. [Google Scholar] [CrossRef] [Green Version]
- Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J.; Corbett, T.H. Total structure determination of Apratoxin A, a potent novel cytotoxin from the marine Cyanobacterium Lyngbya majuscula. J. Am. Chem. Soc. 2001, 123, 5418–5423. [Google Scholar] [CrossRef]
- Luo, D.; Chen, Q.-Y.; Luesch, H. Total synthesis of the potent marine-derived elastase inhibitor lyngbyastatin 7 and in vitro biological evaluation in model systems for pulmonary diseases. J. Org. Chem. 2015, 81, 532–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taori, K.; Matthew, S.; Rocca, J.R.; Paul, V.J.; Luesch, H.J. Lyngbyastatins 5–7, potent elastase inhibitors from Floridian marine cyanobacteria, Lyngbya spp. J. Nat. Prod. 2007, 70, 1593–1600. [Google Scholar] [CrossRef] [PubMed]
- Fuwa, H.; Okuaki, Y.; Yamagata, N.; Sasaki, M. Total synthesis, stereochemical reassignment, and biological evaluation of (−)-Lyngbyaloside B. Angew. Chem. 2015, 127, 882–887. [Google Scholar] [CrossRef]
- Luesch, H.; Yoshida, W.Y.; Harrigan, G.G.; Doom, J.P.; Moore, R.E.; Paul, V.J. Lyngbyaloside B, a new glycoside macrolide from a Palauan marine cyanobacterium, Lyngbya sp. J. Nat. Prod. 2002, 65, 1945–1948. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, A.; Iwasaki, A.; Suenaga, K. Total synthesis and stereochemical reassignment of maedamide. Tetrahedron Lett. 2015, 56, 4947–4949. [Google Scholar] [CrossRef]
- Iwasaki, A.; Ohno, O.; Sumimoto, S.; Ogawa, H.; Nguyen, K.A.; Suenaga, K. Jahanyne, an apoptosis-inducing lipopeptide from the marine cyanobacterium Lyngbya sp. Org. Lett. 2015, 17, 652–655. [Google Scholar] [CrossRef]
- Iwasaki, A.; Fujimura, H.; Okamoto, S.; Kudo, T.; Hoshina, S.; Sumimoto, S.; Teruya, T.; Suenaga, K. Isolation of jahanene and jahanane, and total synthesis of the jahanyne family. J. Org. Chem. 2018, 83, 9592–9603. [Google Scholar] [CrossRef]
- Inuzuka, T.; Yamamoto, K.; Iwasaki, A.; Ohno, O.; Suenaga, K.; Kawazoe, Y.; Uemura, D. An inhibitor of the adipogenic differentiation of 3T3-L1 cells, yoshinone A, and its analogs, isolated from the marine cyanobacterium Leptolyngbya sp. Tetrahedron Lett. 2014, 55, 6711–6714. [Google Scholar] [CrossRef]
- Cui, J.; Morita, M.; Ohno, O.; Kimura, T.; Teruya, T.; Watanabe, T.; Suenaga, K.; Shibasaki, M. Leptolyngbyolides, cytotoxic macrolides from the marine cyanobacterium Leptolyngbya sp.: Isolation, biological activity, and catalytic asymmetric total synthesis. Chem. A Eur. J. 2017, 23, 8500–8509. [Google Scholar] [CrossRef]
- Dai, L.; Chen, B.; Lei, H.; Wang, Z.; Liu, Y.; Xu, Z.; Ye, T. Total synthesis and stereochemical revision of lagunamide A. Chem. Commun. 2012, 48, 8697–8699. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, A.; Puddick, J.; Prinsep, M.R.; Rottmann, M.; Tan, L.T. Lagunamides A and B: Cytotoxic and antimalarial cyclodepsipeptides from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2010, 73, 1810–1814. [Google Scholar] [CrossRef]
- White, J.D.; Xu, Q.; Lee, C.-S.; Valeriote, F.A. Total synthesis and biological evaluation of (+)-kalkitoxin, a cytotoxic metabolite of the cyanobacterium Lyngbya majuscula. Org. Biomol. Chem. 2004, 2, 2092–2102. [Google Scholar] [CrossRef]
- Li, W.I.; Berman, F.W.; Okino, T.; Yokokawa, F.; Shioiri, T.; Gerwick, W.H.; Murray, T.F. Antillatoxin is a marine cyanobacterial toxin that potently activates voltage-gated sodium channels. Proc. Natl. Acad. Sci. USA 2001, 98, 7599–7604. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, V.-A.; Willis, C.L.; Gerwick, W.H. Synthesis of the marine natural product barbamide. Chem. Commun. 2001, 1934–1935. [Google Scholar] [CrossRef]
- Orjala, J.; Gerwick, W.H. Barbamide, a chlorinated metabolite with molluscicidal activity from the Caribbean cyanobacterium Lyngbya majuscula. J. Nat. Prod. 1996, 59, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Pirovani, R.; Brito, G.; Barcelos, R.; Pilli, R. Enantioselective Total Synthesis of (+)-Lyngbyabellin M. Mar. Drugs 2015, 13, 3309–3324. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.; Mevers, E.; Byrum, T.; Valeriote, F.A.; Gerwick, W.H. Lyngbyabellins K–N from two Palmyra atoll collections of the marine cyanobacterium Moorea bouillonii. Eur. J. Org. Chem. 2012, 2012, 5141–5150. [Google Scholar] [CrossRef] [PubMed]
- Ojima, D.; Iwasaki, A.; Suenaga, K. Total synthesis of kanamienamide and clarification of biological activity. J. Org. Chem. 2017, 82, 12503–12510. [Google Scholar] [CrossRef] [PubMed]
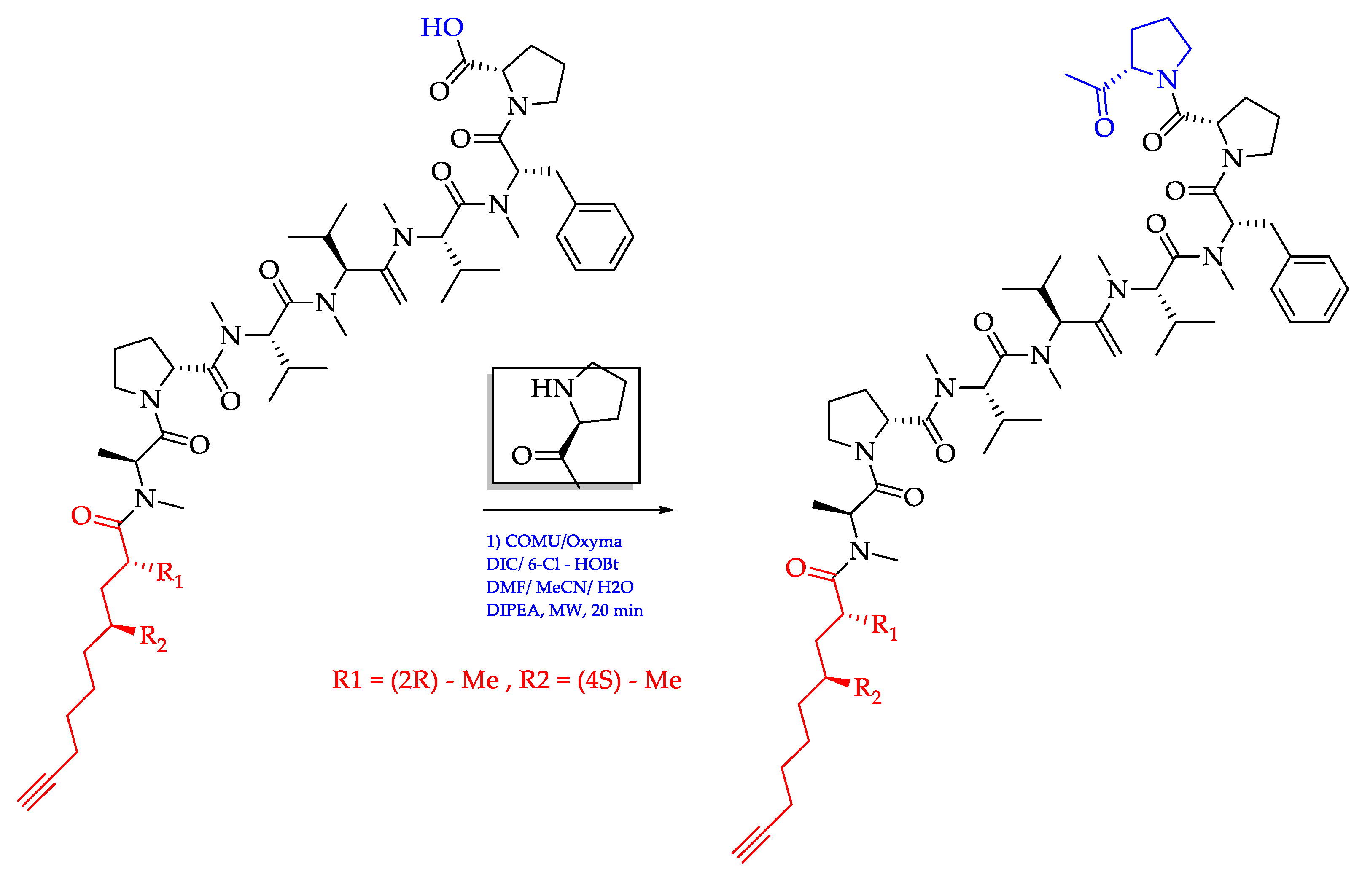
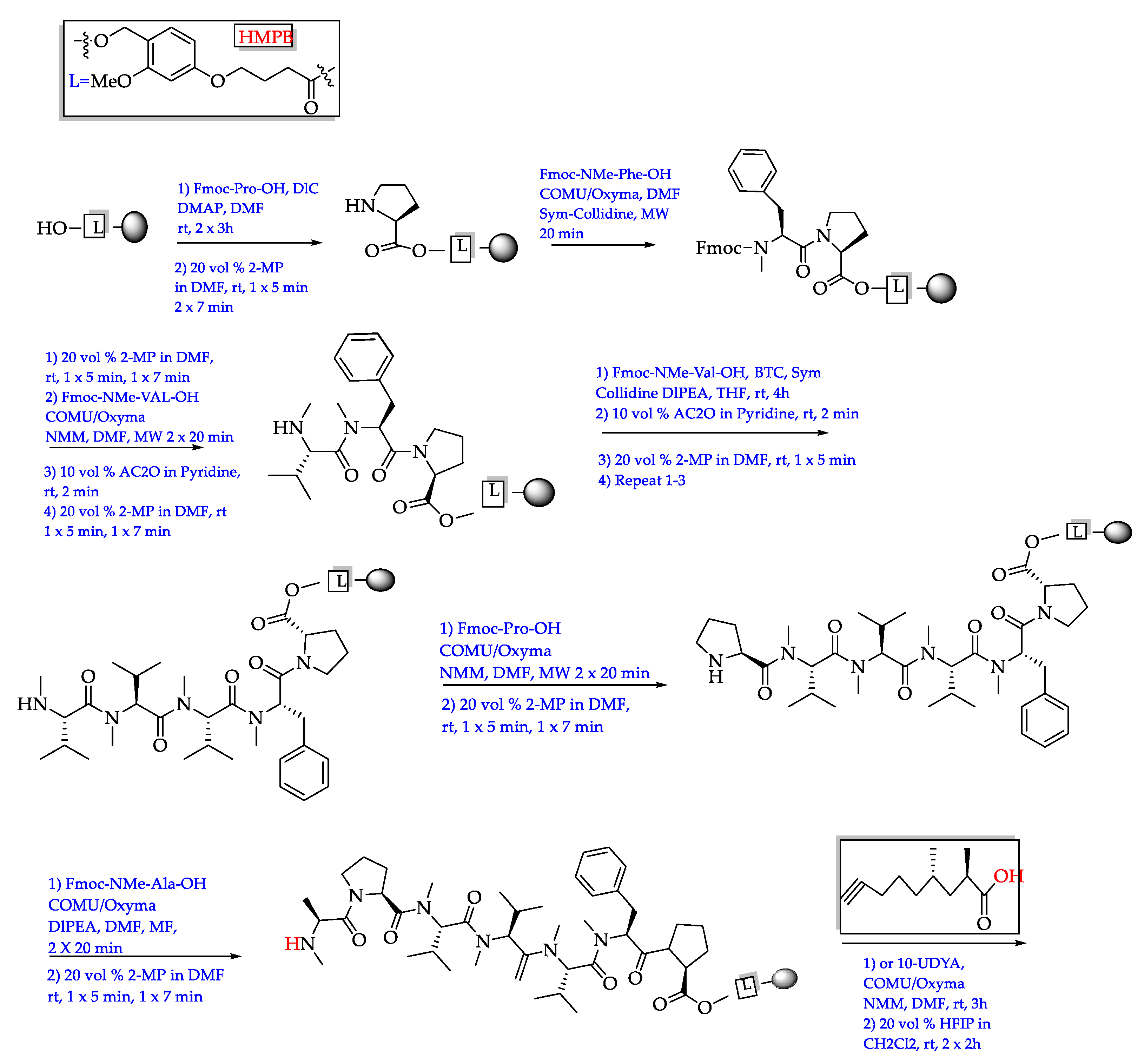
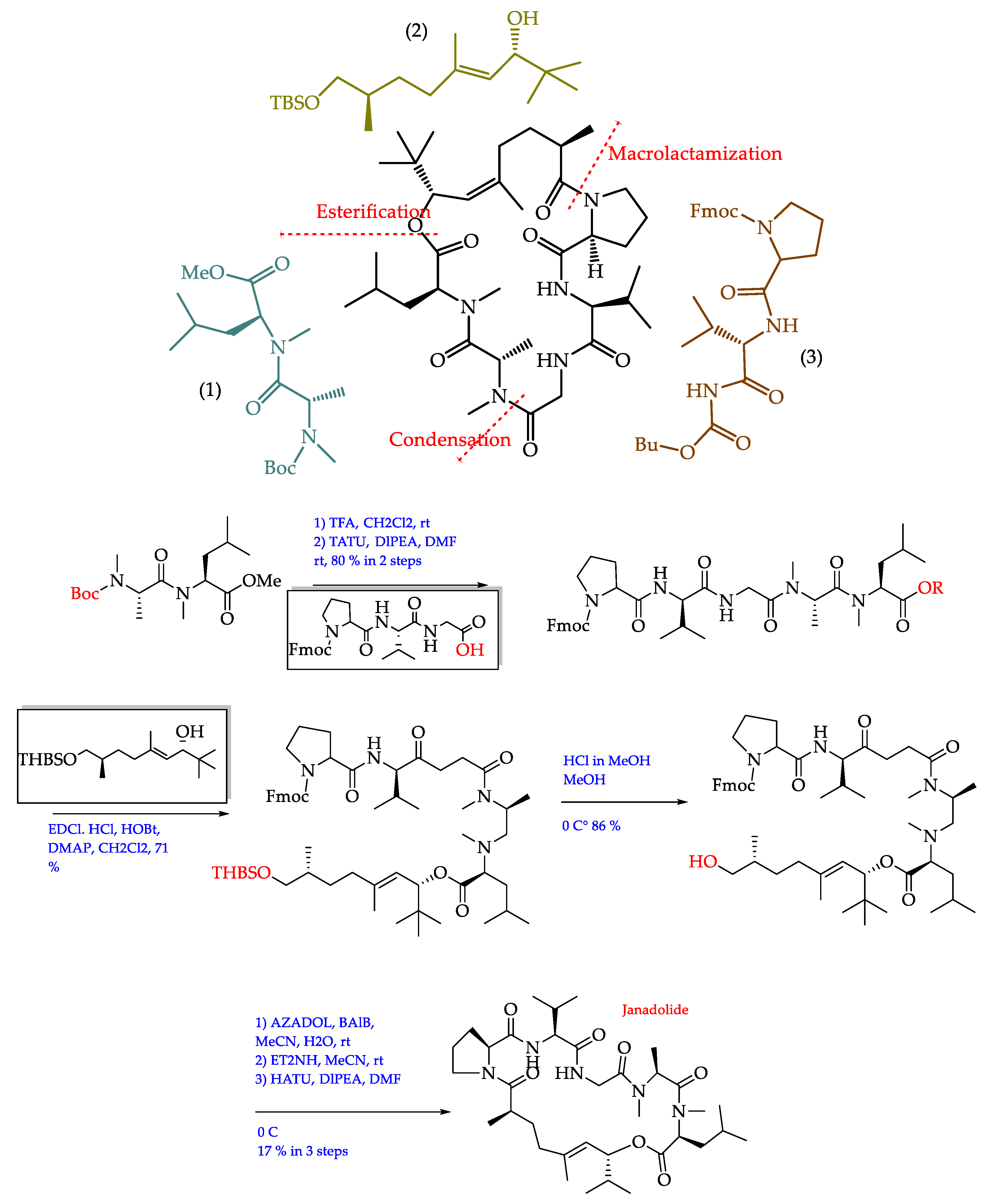
- Ogawa, H.; Iwasaki, A.; Sumimoto, S.; Kanamori, Y.; Ohno, O.; Iwatsuki, M.; Ishiyama, A.; Hokari, R.; Otoguro, K.; Ōmura, S.; et al. Janadolide, a cyclic polyketide–peptide hybrid possessing a tert-butyl group from an Okeania sp. marine cyanobacterium. J. Nat. Prod. 2016, 79, 1862–1866. [Google Scholar] [CrossRef]
- Sueyoshi, K.; Kaneda, M.; Sumimoto, S.; Oishi, S.; Fujii, N.; Suenaga, K.; Teruya, T.J.T. Odoamide, a cytotoxic cyclodepsipeptide from the marine cyanobacterium Okeania sp. Tetrahedron 2016, 72, 5472–5478. [Google Scholar] [CrossRef]
- Sun, T.; Zhang, W.; Zong, C.; Wang, P.; Li, Y. Total synthesis and stereochemical reassignment of tasiamide B. J. Peptide Sci. 2010, 16, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Gunasekera, S.P.; Li, Y.; Ratnayake, R.; Luo, D.; Lo, J.; Reibenspies, J.H.; Xu, Z.; Clare-Salzler, M.J.; Ye, T.; Paul, V.J. Discovery, total synthesis and key structural elements for the immunosuppressive activity of cocosolide, a symmetrical glycosylated macrolide dimer from marine cyanobacteria. Chemistry 2016, 22, 8158–8166. [Google Scholar] [CrossRef] [Green Version]
- Namikoshi, M.; Murakami, T.; Fujiwara, T.; Nagai, H.; Niki, T.; Harigaya, E.; Watanabe, M.F.; Oda, T.; Yamada, J.; Tsujimura, S. Biosynthesis and transformation of homoanatoxin-a in the cyanobacterium Raphidiopsis mediterranea Skuja and structures of three new homologues. Chem. Res. Toxicol. 2004, 17, 1692–1696. [Google Scholar] [CrossRef]
- Carneiro, V.M.; Avila, C.M.; Balunas, M.J.; Gerwick, W.H.; Pilli, R.A. Coibacins A and B: Total synthesis and stereochemical revision. J. Org. Chem. 2014, 79, 630–642. [Google Scholar] [CrossRef] [Green Version]
- Balunas, M.J.; Grosso, M.F.; Villa, F.A.; Engene, N.; McPhail, K.L.; Tidgewell, K.; Pineda, L.M.; Gerwick, L.; Spadafora, C.; Kyle, D.E. Coibacins A–D, antileishmanial marine cyanobacterial polyketides with intriguing biosynthetic origins. J. Nat. Prod. 2012, 14, 3878–3881. [Google Scholar] [CrossRef] [PubMed]
- Ojima, D.; Yasui, A.; Tohyama, K.; Tokuzumi, K.; Toriihara, E.; Ito, K.; Iwasaki, A.; Tomura, T.; Ojika, M.; Suenaga, K. Total Synthesis of Miuraenamides A and D. J. Org. Chem. 2016, 81, 9886–9894. [Google Scholar] [CrossRef]
- Wang, D.; Song, S.; Tian, Y.; Xu, Y.; Miao, Z.; Zhang, A. Total synthesis of the marine cyclic depsipeptide viequeamide A. J. Nat. Prod. 2013, 76, 974–978. [Google Scholar] [CrossRef] [PubMed]
- Boudreau, P.D.; Byrum, T.; Liu, W.-T.; Dorrestein, P.C.; Gerwick, W.H. Viequeamide A, a cytotoxic member of the kulolide superfamily of cyclic depsipeptides from a marine button cyanobacterium. J. Nat. Prod. 2012, 75, 1560–1570. [Google Scholar] [CrossRef] [Green Version]
- Lemmens-Gruber, R.; Kamyar, M.R.; Dornetshuber, R. Cyclodepsipeptides-potential drugs and lead compounds in the drug development process. Curr. Med. Chem. 2009, 16, 1122–1137. [Google Scholar] [CrossRef]
- Kitagaki, J.; Shi, G.; Miyauchi, S.; Murakami, S.; Yang, Y. Cyclic depsipeptides as potential cancer therapeutics. Anti Cancer Drugs 2015, 26, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Siow, A.; Opiyo, G.; Kavianinia, I.; Li, F.F.; Furkert, D.P.; Harris, P.W.R.; Brimble, M.A. Total synthesis of the highly N-methylated acetylene-containing anticancer peptide jahanyne. Org. Lett. 2018, 20, 788–791. [Google Scholar] [CrossRef] [PubMed]
- Kunifuda, K.; Iwasaki, A.; Nagamoto, M.; Suenaga, K. Total synthesis and absolute configuration of koshikalide. Tetrahedron Lett. 2016, 57, 3121–3123. [Google Scholar] [CrossRef]
- Shen, B. Polyketide biosynthesis beyond the type I, II and III polyketide synthase paradigms. Curr. Opin. Chem. Biol. 2003, 7, 285–295. [Google Scholar] [CrossRef]
- Staunton, J.; Weissman, K.J. Polyketide biosynthesis: A millennium review. Nat. Prod. Rep. 2001, 18, 380–416. [Google Scholar] [CrossRef] [PubMed]
- Ojima, D.; Mine, H.; Iwasaki, A.; Suenaga, K. Total synthesis of janadolide. Tetrahedron Lett. 2018, 59, 1360–1362. [Google Scholar] [CrossRef]
- Iwasaki, A.; Ohno, O.; Sumimoto, S.; Suda, S.; Suenaga, K. Kurahyne, an acetylene-containing lipopeptide from a marine cyanobacterial assemblage of Lyngbya sp. RSC Adv. 2014, 4, 12840–12843. [Google Scholar] [CrossRef] [Green Version]
- Sansone, C.; Brunet, C.; Noonan, D.M.; Albini, A. Marine algal antioxidants as potential vectors for controlling viral diseases. Antioxidants 2020, 9, 392. [Google Scholar] [CrossRef] [PubMed]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name (No.)/Chemical Class | Cyanobacteria Species/Source | Type of Activity | Clinical Status/Study Type | References |
|---|---|---|---|---|
| Dolastatin 10 (44)/Depsipeptides | Symploca sp. | Sarcoma, Leukemia, Lymphoma, Liver Cancer, and Kidney Cancer, among others. | Drug Investigational | [92,93,94,95,96,97,98,99,100] |
| Leukemia Lymphoma | Drug Intervention Phase II | |||
| Prostate Cancer | Drug Intervention Phase II | |||
| Kidney Cancer | Drug Intervention Phase II | |||
| Extrahepatic Bile Duct Cancer Gallbladder Cancer Liver Cancer | Drug Intervention Phase II | |||
| Ovarian Cancer Sarcoma | Drug Intervention Phase I | |||
| Leukemia Myelodysplastic Syndromes | Drug Intervention Phase I | |||
| Pancreatic Cancer | Drug Phase II Intervention | |||
| Cemadotin (47), Tasidotin (48) and Synthadotin (49) (Derived from dolastatin 15 (45)/Depsipeptide | Dolabella auricularia and cyanobacteria Symploca (later) | Melanoma | Drug: ILX651 Intervention Phase II | |
| Hormone-refractory Prostate Cancer | [101,102,103,104] | |||
| Non-Small-Cell Lung Carcinoma | ||||
| Cryptophycin 52 (43)/(Synthetic analog of cryptophycin 1 (5) Depsipeptides | Nostoc sp., terrestrial cyanobacteria | Schizophrenia | Cognitive remediation therapy, Intervention Not Applicable Phase | [101,105,106] |
| Hypertension Metabolic Disorder | Drug: losartan potassium (+) hydrochlorothiazide, Intervention Phase III | |||
| Toyocamycin (52)/Pyrrolopyrimidine nucleoside | Streptomyces toyocaensis Cyanobacteria | Non-Small-Cell Lung Carcinoma | Drug Experimental | [101,107] |
| Phytoalexin (53)/Polysaccharides | Scytonema ocellatum | Type 2 Diabetes (RED) | Drug Phase I Intervention | [108,109] |
| Soblidotin (46)/(Synthetic analog of dolastatin 10) Depsipeptides | Sarcoma | Drug Intervention Phase II | [101,110] | |
| Lung Cancer | ||||
| Phycocyanin (54)/A pigment-protein complex | Spirulina | Chronic Periodontitis | Drug: Spirulina capsules, Intervention Phase IV | [111,112,113] |
| Metabolic Syndrome | Dietary Supplement: Spirulysat® Dietary Supplement: Placebo Intervention Not Applicable Phase | |||
| Anatoxins-a (51)/ Peptides | Anabeana circinalis | Amyotrophic Lateral Sclerosis | Recruiting Patient Registry Intervention | [114,115] |
| Bacteriocins/Peptides | 43 different cyanobacteria viz., Prochlorcoccus marinus, Synechococcus sp., Cyanothece sp., Microcystis aeruginosa, Synechocystis, Arthospira, Nostoc, Anabaena, Nodularia | Ventilator Associated Pneumonia | Lactobacillus bacteria Intervention | [116,117,118,119,120] |
| Colic, Infantile Probiotic Gut Microbiome Bifidobacterium Breve | Drug Intervention Phase IV | |||
| Healthy | Plantaricin A—rejuvenating cream, antioxidant serum, rejuvenating serum Intervention Phase III | |||
| White Spot Lesion of Tooth Long Term Adverse Effects Caries, Dental Orthodontic Appliance Complication | Drug: Probiotic Toothpaste Drug: Dr. Reddy’s Clohex Other: Control Group Intervention Phase I and II | |||
| Curacin (50)/Lipopeptides | Lyngbya majuscule | In vivo animal trails. Preclinical Phase (but it served as a lead compound) | [91] | |
| Cyanovirin-N (40) (CVN)/A protein | Nostoc ellipsosporum | Inhibiting HIV cell entry in a highly specific manner. | Preclinical Phase | [121] |
| Marine Source | Compound Name/Class | Region/Year | Biological Activity | References |
|---|---|---|---|---|
| Marine cyanobacterium | Hoshinolactam (58) | The coast near Hoshino, Okinawa/2017 | Antitrypanosomal activity, IC50 = 3.9 (Syn.), 6.1 (Nat.) nM. Cytotoxicity against MRC-5 cells IC50 > 25 μM (Syn. and Nat.) PC = pentamidinea NC = not (in vitro) | [189] |
| Lyngbya sp. | Koshikalide (59) | Koshika, Shima City, Mie prefecture/2010 | Cytotoxicity against HeLa S3 cells, IC50 = 42 µg/mL. PC = not NC = not (in vitro) | [190] |
| Lyngbya majuscule. | Apratoxin A (22)/Cyclodepsipeptide | Finger’s Reef, Apra Harbor, Guam/2001 | Cytotoxicity against KB (IC50 = 0.52 nM) and LoVo cancer cells (IC50 = 0.36 nM). (in vitro) Against a colon tumor and ineffective against a mammary tumor. (in vivo) | [191,192] |
| Lyngbya sp. & Lyngbya confervoides./ | Lyngbyastatin 7 (60)/Lariat-type cyclic depsipeptide | Mangrove channel, Kemp Channel, at the northern end of Summerland Key, Florida Keys/2005 | Blocking elastase activity, IC50 = 70 nM, antiproliferation and abrogating the elastase-triggered induction of proinflammatory cytokine expression. PC = sivelestat, or (DMSO) NC = NR (in vivo) | [193,194] |
| Lyngbya bouillonii | (−)-Lyngbyaloside B (61)/Glycoside macrolide | Ulong Channel, Palau/2000 | Cytotoxicity against KB cells, IC50 = 4.3 µM and LoVo cells, IC50 = 15 µM. | [195,196] |
| Lyngbya sp. | Maedamide (62)/Acyclic peptide | Kuraha, Okinawa/2014 | Inhibitory activity against chymotrypsin, IC50 = 45 μM, HeLa and HL60 cells, IC50 = 4.2 and 2.2 μM. | [197] |
| Lyngbya sp. | Jahanyne (63)/Lipopeptides | The coast near Jahana, Okinawa, Japan/2015 | Cytotoxicity against HeLa cells and HL60 cells, IC50 = 1.8 μM and 0.63 μM (et al., 2015) natural jahanyne, IC50 = (22 ± 2, 4.6 ± 1.2 μM) and synthetic (21 ± 2, 8.3 ± 2.3 μM). | [198,199] |
| Leptolyngbya sp. | Yoshinone A (64) | Ishigaki island, Okinawa, Japan/2014 | Antiobesity activity (in vivo) in mice, (Inhibited differentiation of 3T3-L1 cells into adipocytes, EC50 = 420 nM) and toxicity against Saccharomyces cerevisiae ABC16-Monster, (IC50 = 63.8 µM). | [200] |
| Leptolyngbyolide C (65)/Macrolide | On the coast of Itoman City, Okinawa, Japan/2007 | Growth-inhibitory activity against HeLa S3 cells, (IC50 = 0.64 µg mL−1) and depolymerization of F-actin (EC50 = 26.9 µg mL−1). (in vitro) | [201] | |
| Lyngbya majuscula | Lagunamide A (66)/Cyclodepsipeptide | Western lagoon of Pulau Hantu Besar, Singapore/June 2007 | Antimalarial activity against Plasmodium falciparum, IC50 = 0.19 and cytotoxic activity against P388 murine leukemia cell lines, IC50 = 6.4 nM, and moderate antiswarming activities against Pseudomonas aeruginosa PA01. PC: MeOH-treated plate | [202,203] |
| Lyngbya majuscula | (−)-kalkitoxin (67) | Curaçao/2004 | Cytotoxicity against the human colon cell line HCT-116, IC50 = 1.0 × 10−3 μg mL−1, inhibited hypoxia-induced activation of HIF-1 in T47D breast tumor cells (IC50 = 5.6 nM) | [204] |
| Lyngbya majuscula | Antillatoxin (41)/Cyclic lipodepsipeptide | Curacüao/2005 | Strong ichthyotoxicity and neurotoxicity (EC50 = 20.1 ± 6.4 nM). | [187,205] |
| Lyngbya majuscula & Schizothrix sp. | Somamide A (68)/Macrocyclic depsipeptide | Fijian Island/2005 | [187] | |
| Lyngbya majuscula | Barbamide (69)/Lipopeptide | Curacüao/1996 | Potent molluscicidal activity against Biomphalaria glabrata, LC100 = 100 µg/mL | [206,207] |
| Moorea bouillonii | (+)-Lyngbyabellin M (70)/Lipopeptide | North lagoon at Strawn Island, Palmyra Atoll, USA/August 2009 | Not reported | [208,209] |
| Kanamienamide (71) | The shore of Kanami, Kagoshima, Japan/2016 | Growth-inhibitory activity. As a necrosislike cell death inducer. | [210] | |
| Okeania sp. | Janadolide (72)/Cyclic polyketide- peptide | Bise, Okinawa Prefecture, Japan/2016 | Antitrypanosomal activity without cytotoxicity against human cells (IC50 47 nM) | [211] |
| Kurahyne (73) (N-Me) Kurahyne B (74) (N-H) | The coast near Jahana, Okinawa/March 2013 | Growth-inhibitory activity (Inhibited the growth of both HeLa and HL60 cells, IC50 = 8.1 and 9.0 μM) PC = Adriamycin | [186] | |
| Odoamide (75)/Cyclodepsipeptide | Odo, Okinawa Prefecture, Japan/May 2009 | (in vitro) Cytotoxicity against HeLa S3 cells, IC50 = 26.3 nM. Toxicity against brine shrimp (Artemia), LD50 = 1.2 µM. | [212] | |
| Symploca sp. | Tasiamide B (76)/Acyclic peptide | Micronesia by Moore et al., 2003 | Cytotoxic against KB cells, IC50 = 0.8 µM | [213] |
| Cocosolide (77)/Glycosylated macrolide | Cocos Lagoon and Tanguisson reef flat, Guam/2016 | Inhibited IL-2 production in both T-cell receptors also suppressed the proliferation of anti-CD3-stimulated T-cells in a dose-dependent manner. (IC50 > 50 mm). | [214] | |
| Oscillatoria Formosa | Homoanatoxin-a (78) | Inniscarra reservoir, County Cork, Ireland/2004 | Cytotoxic activity LD50’s in mice of 200–250 µg/kg. | [215] |
| Oscillatoria sp. | Coibacin A (79)/Unsaturated polyketide lactone | Panamanian/2012 | Antileishmanial activity against axenic amastigotes of Leishmania donovani (IC50 = 2.4 μM). Cytotoxicity against NCI-H460 cells (IC50 = 31.5 μM). Antiinflammatory activity by cell-based nitric oxide (NO) (IC50 = 20 μM). | [216,217] |
| Coibacin B (80)/Unsaturated polyketide lactone | As a leishmanicidal drug (IC50 = 7.2 μM); cytotoxicity against human cancer lung cell lines (NCI-H460), IC50 = 17.0 μM. Active coibacin representative (IC50 = 5 μM). | |||
| Paraliomixa miuraensis | Miuraenamide A (81) (R1 = Ph, R2 = O Me) Miuraenamide D (82) (R1 = O Me, R2= Ph)/Cyclodepsipeptides | The seashore on Miura Peninsula in Kanagawa, Japan by Ojika et al., 2006 | Cytotoxicity against HeLa cells, IC50 = A (0.031), D (0.021) μM. Against HeLa-S3 cell line, IC50 = A (0.38), D (1.32) μM. antiphytophthora activity 3, 30 ng/disk | [218] |
| Rivularia sp. “button” Marine cyanobacterium | Viequeamide A (83)/Cyclic depsipeptide | Near the island of Vieques, Puerto Rico/2012 | Highly toxic against H460 human lung cancer cell lines, IC50 = 60 nm. PC = paclitaxel (3.2 nM) and etoposide (63.1 nM) | [219,220] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalifa, S.A.M.; Shedid, E.S.; Saied, E.M.; Jassbi, A.R.; Jamebozorgi, F.H.; Rateb, M.E.; Du, M.; Abdel-Daim, M.M.; Kai, G.-Y.; Al-Hammady, M.A.M.; et al. Cyanobacteria—From the Oceans to the Potential Biotechnological and Biomedical Applications. Mar. Drugs 2021, 19, 241. https://doi.org/10.3390/md19050241
Khalifa SAM, Shedid ES, Saied EM, Jassbi AR, Jamebozorgi FH, Rateb ME, Du M, Abdel-Daim MM, Kai G-Y, Al-Hammady MAM, et al. Cyanobacteria—From the Oceans to the Potential Biotechnological and Biomedical Applications. Marine Drugs. 2021; 19(5):241. https://doi.org/10.3390/md19050241
Chicago/Turabian StyleKhalifa, Shaden A. M., Eslam S. Shedid, Essa M. Saied, Amir Reza Jassbi, Fatemeh H. Jamebozorgi, Mostafa E. Rateb, Ming Du, Mohamed M. Abdel-Daim, Guo-Yin Kai, Montaser A. M. Al-Hammady, and et al. 2021. "Cyanobacteria—From the Oceans to the Potential Biotechnological and Biomedical Applications" Marine Drugs 19, no. 5: 241. https://doi.org/10.3390/md19050241