
New Probabilistic, Dynamic Multi-Method Ensembles for Optimization Based on the CRO-SL
 , , , , and
, , , , and
Abstract
:1. Introduction
1.1. Literature Review
1.2. Contribution and Structure
2. Methods
2.1. The CRO-SL: A Multi-Method-Ensemble Evolutionary Algorithm
2.1.1. Basic CRO
- 1.
- Initialization: A fraction of the total reef capacity is occupied with randomly generated corals. The reef position that each coral occupies is also randomly selected.
- 2.
- Evolution: Once the reef has been populated, the evolutionary process begins. This process is divided into five phases per generation:
- (a)
- Sexual reproduction: In this phase, new solutions (larvae set) are created from the ones belonging to the reef in order to compete for a place in the reef. Sexual reproduction can be performed in two ways: external and internal. A percentage () of the corals settled in the reef perform external reproduction (Broadcast spawning), and the rest of them reproduce themselves through internal sexual reproduction (brooding). These reproduction processes are performed as follows:
- i.
- Broadcast spawning: from the set of corals selected for external sexual reproduction (), new solutions (larvae) are generated and released.
- ii.
- Brooding: each one of the remaining corals () produces a larva by means of a small perturbation and releases it.
- (b)
- Larvae setting: In this step, all the larvae produced by broadcast spawning or brooding try to find a spot in the reef to grow up. A reef position is randomly chosen, and the larva will settle in that spot in only one of the following scenarios:
- i.
- The spot is empty.
- ii.
- The larva has a better health function value (fitness) than the coral currently occupying that spot.
Each larva can try to settle in the reef a maximum of three times. If the larva has not been able to settle down in the reef after that number of attempts, it is discarded. - (c)
- Asexual reproduction: In this phase (also called budding), a fraction of the corals with better fitness present in the reef duplicate themselves, and after a small number of mutations, are released. They will try to settle in the reef as in the previously described step.
- (d)
- Depredation: Finally, each coral belonging to the worst fraction can be predated (erased from the reef) with a low probability, .
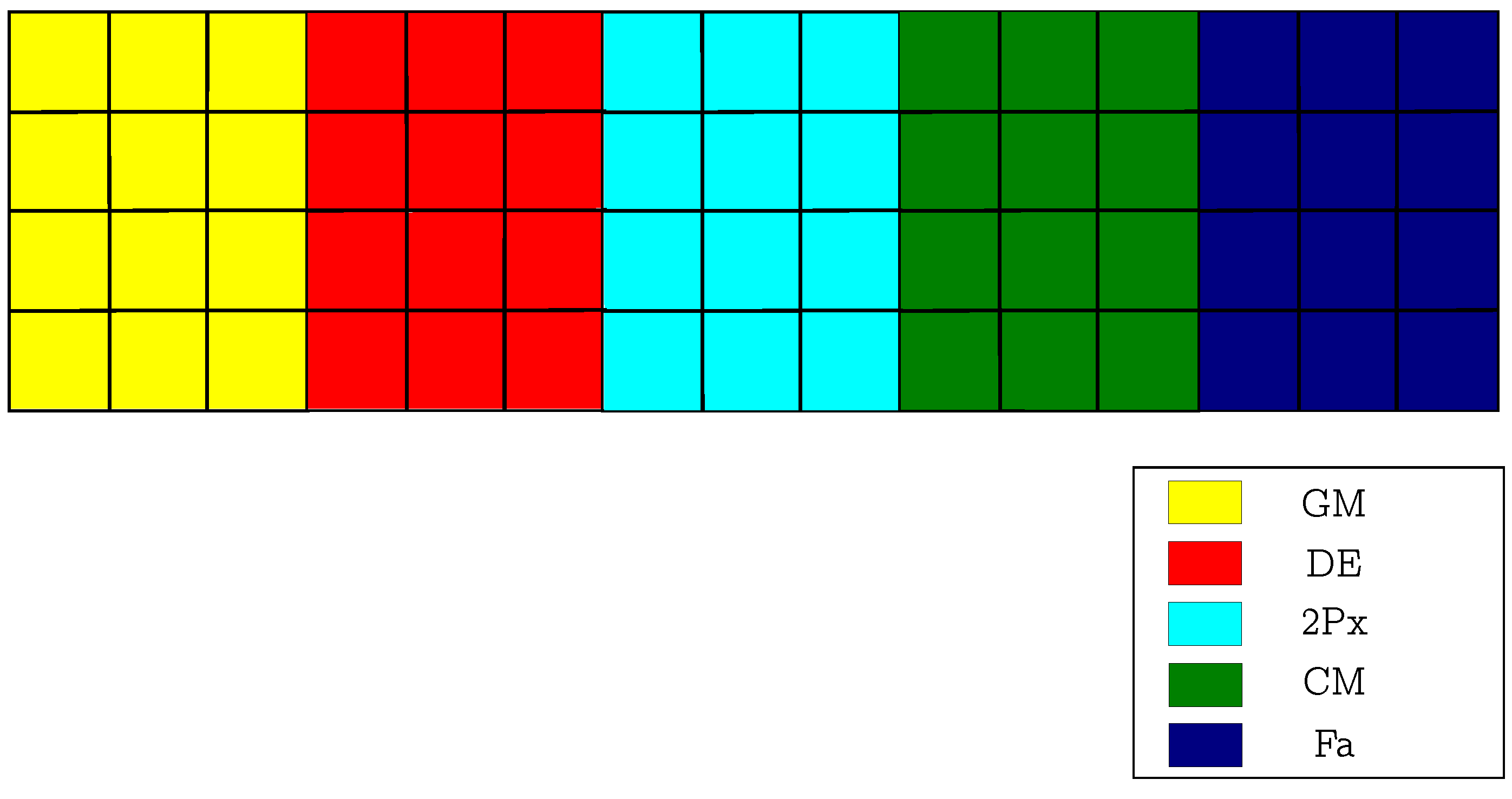
2.1.2. CRO with Substrate Layers (CRO-SL)
2.2. Substrate Layers Defined in the CRO-SL
- DE: The DE algorithm [15] is a stochastic population-based method specifically designed for global optimization problems [46]. In its more common form, DE maintains a population with individuals, where every individual within the population stands for a possible solution to the problem. Individuals are represented by a vector , where and g refers to the index of the generation. A normal DE cycle consists of three consecutive steps: mutation, crossover, and selection. We adapted the algorithm for the CRO-SL by considering only the mutation and crossover parts of the meta-heuristic. Thus, mutation is carried out to generate random perturbations on the population. For each individual, a mutant vector is generated. There are different approaches for DE mutation in the literature [15]. We describe here the procedure known as the “best mutation strategy” [47], which has been successfully applied in many optimization problems before. It attempts to mutate the best individual of the population, according to Equation (1), where denotes the mutated vector, i is the index of the vector, g stands for the generation index, are randomly created integers, denotes the best solution in the population, and F is the scaling factor in the interval . This mutation strategy uses the scaled difference between two randomly selected vectors to mutate the best individual in the population.A crossover procedure is then applied between the mutated vector created in the mutation stage and an individual randomly chosen from the population. The new solutions created are called trial vectors and denoted by for individual i at generation g. Every parameter in the trial vector is decided following Equation (2), where j represents the index of every parameter in a vector, is the probability of recombination, and denotes a randomly selected integer within to ensure that at least one parameter from the mutated vector enters the trial vector:
- Fa: The Fa is a kind of swarm intelligence algorithm based on the flashing patterns and behavior of fireflies in nature [48,49]. In this algorithm, the pattern movement of a firefly i attracted to another (brighter) firefly j is calculated as follows:where stands for the attractiveness at distance . The specific Fa mutation implemented in the CRO-SL is a modified version of the algorithm known as the neighborhood attraction firefly algorithm (NaFa) [50]. It has been implemented as follows: When a coral (solution) in the reef belongs to the Fa substrate, it is updated following Equation (3). All the parameters of the equation are tuned during the CRO-SL evolution. The corals in the Fa substrate consider as swarm a neighborhood among all other corals in the reef (not only the Fa substrate). Thus, the corals in the Fa substrate are updated taking into account some solutions from other substrates, since all the corals in the reef share the same objective function.
- 2Px: Classical 2-point crossover. The crossover operator is the most classical exploration mechanism in genetic and evolutionary algorithms [42,51]. It consists of coupling individuals at random, choosing two points for the crossover, and interchanging the genetic material between both points. In the classical version of the CRO-SL, one individual to be crossed is from the 2Px substrate, whereas the couple can be chosen from any part of the reef.
- BLX: BLX- crossover. This crossover operator [52] considers two real-encoded vectors, and , and generates two offspring, , , where is a randomly (uniformly) chosen number from the interval , where , , and .
- GM: Gaussian mutation with a value linearly decreasing during the run, from to , where is the domain search. Specifically, the Gaussian probability density function is:The reason for adapting the value of throughout the generations is to provide more mutations in the beginning of the optimization and fine tuning with smaller displacements nearing the end. The mutated larva is thus calculated as: , where is a random number following the Gaussian distribution.
- CM: Cauchy mutation. The one-dimensional Cauchy density function centered at the origin is defined by:where is a scale parameter [53]; in this case, . Note that the Cauchy probability distribution looks like the Gaussian distribution, but it approaches the axis so slowly that an expectation does not exist. As a result, the variance of the Cauchy distribution is infinite [53]. In this case, the mutated larva is calculated as: , where stands for a variance and is a random number following the Cauchy distribution.
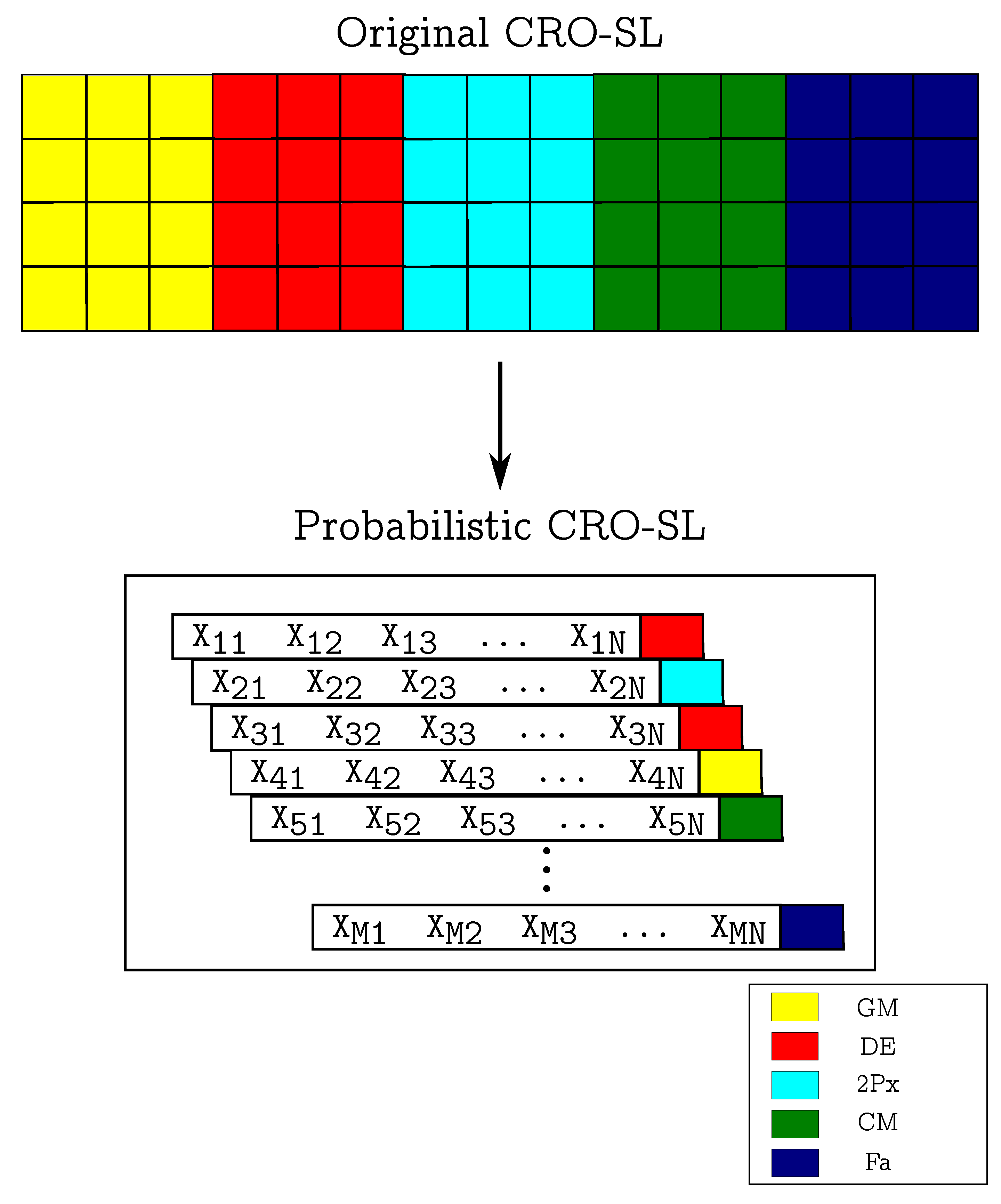
3. Proposed Probabilistic Dynamic Ensembles with the CRO-SL
3.1. Probabilistic CRO-SL Ensemble
| Algorithm 1 Probabilistic CRO-SL. |
|
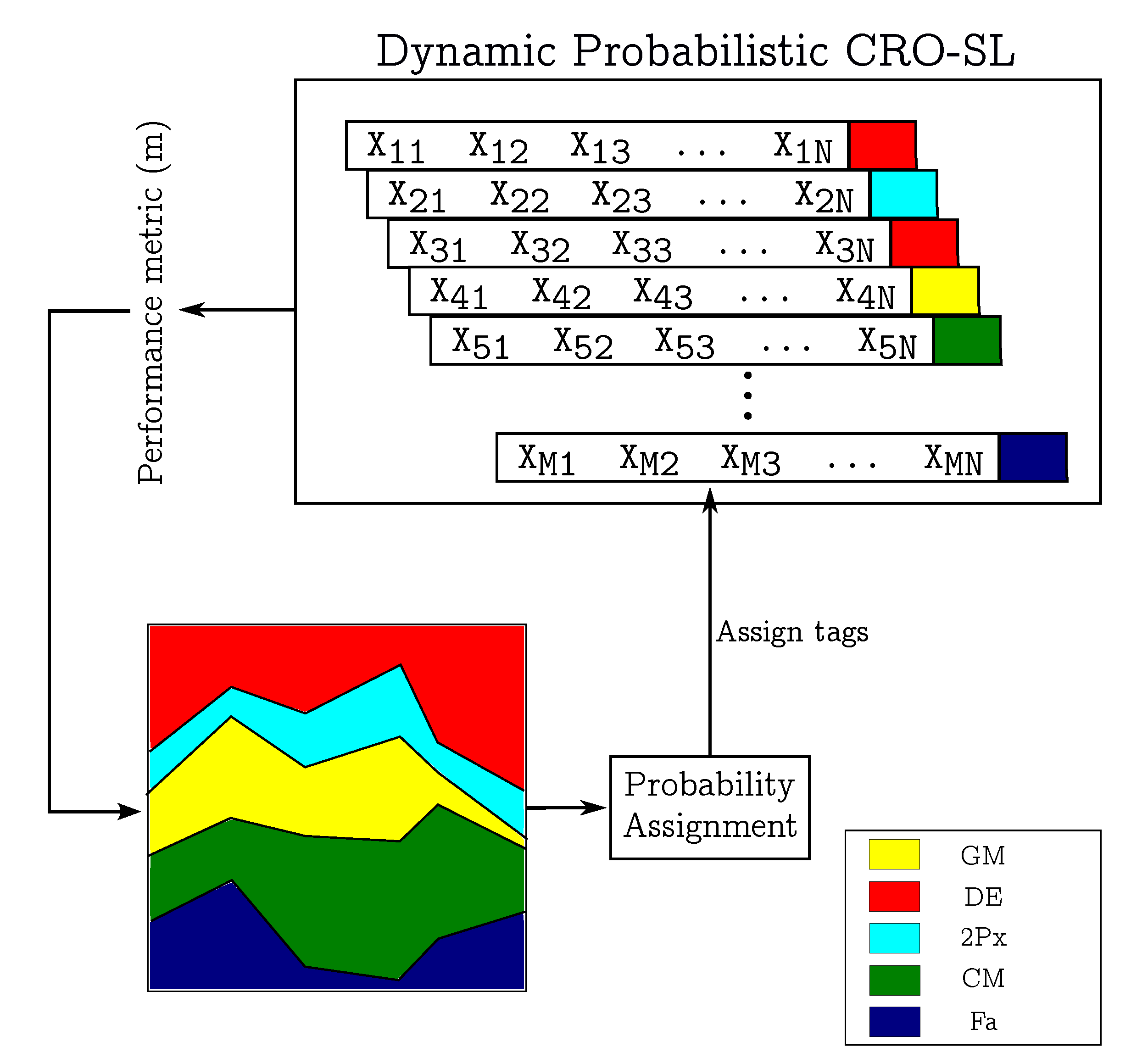
3.2. Dynamic Probabilistic CRO-SL Ensemble
- 1.
- Larval success rate metric. The first probability-assignment procedure depends on the rate of success of the larvae (new solutions) produced by the corals in each substrate. In other words, during the larvae setting phase, we keep track of the substrate (search method) from which each larva was produced, and we note the number of them that were successful in being inserting into the reef. The probability of each searching method in the next step is obtained as the rate of successes of the total number of generated larvae.
- 2.
- Raw fitness metric. The second probability-assignment procedure uses the fitness of the generated solutions; i.e., it considers the quality of individual solutions to obtain a metric for each substrate. In other words, if the operator applied generates good solutions, it will have a higher probability of being assigned to an individual in the next step. Note that there are different ways of implementing this metric: for example, we can take the average of the fitness levels of all the larvae produced, the best fitness across all of them, the worst one, etc.
- 3.
- Improvement of fitness. The last procedure for assigning the methods probabilities is a differential approach, based on the difference from the best fitness level obtained in the previous generation. It works very similarly to the previous strategy, giving higher values to those substrates that generated solutions with better fitness. This method also allows some variants, so we can take the average of the difference, the best value, or the worst value to assign the probability of the method being used in the next step.
| Algorithm 2 Dynamic probabilistic CRO-SL. |
|
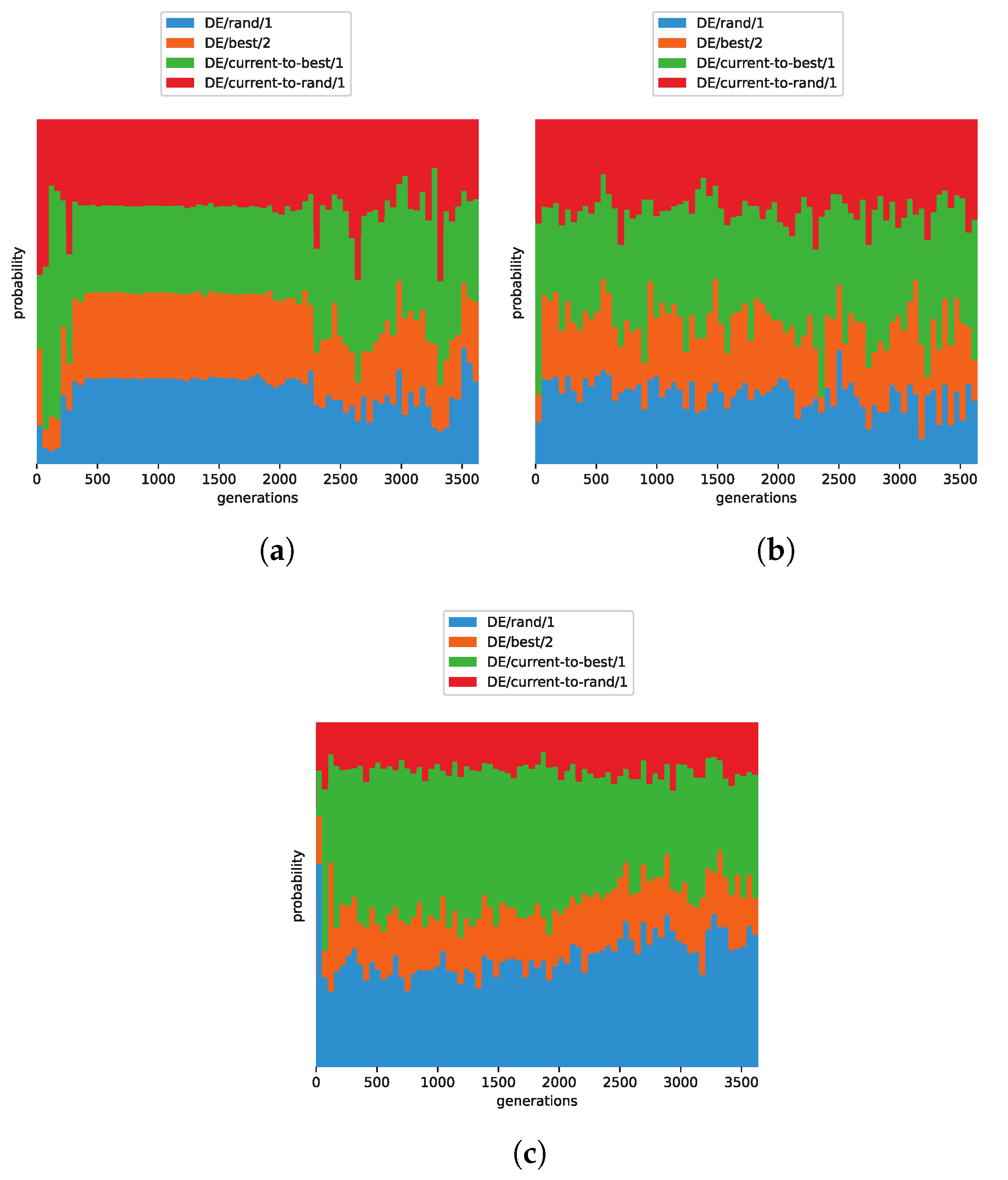
4. Experimental Results
4.1. Comparison in Benchmark Functions
- 1.
- DE/best/1
- 2.
- DE/best/2
- 3.
- DE/current-to-best/1
- 4.
- DE/current-to-pbest/1
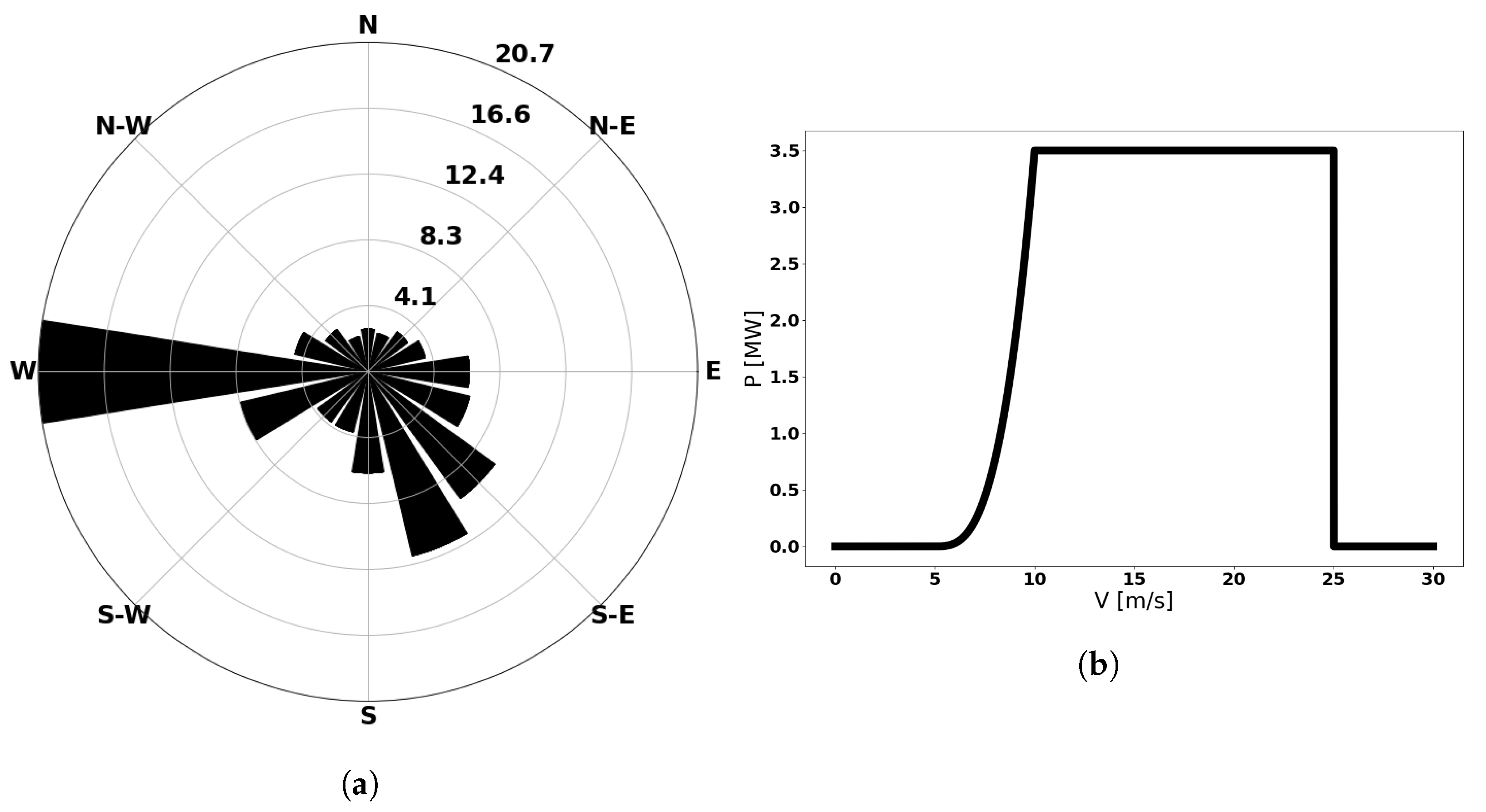
4.2. Comparison in a Real Problem—Wind-Turbine Assignment
Results
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
Appendix A.1. Benchmark Functions
- F1: Sphere.
- F2: High Condition Elliptic.
- F3: Bent Cigar.
- F4: Discus.
- F5: Rosenbrock.
- F6: Ackley.
- F7: Weierstrass (limited to 20 iterations).
- F8: Griewank.
- F9: Rastrigin.
- F10: Modified Schwefel.
- F11: Katsuura.
- F12: Happy Cat.
- F13: HGBat.
- F14: Griewank plus Rosenbrock.
- F15: Exp Shaffer F6.
- F16: Bukin F6.
- F17: Cola; d is a triangular matrix of size 10 × 10.
- F18: CrownedCross.
- F19: CrossLegTable.
- F20: Meyer. Regression with 3 parameters. Fit the model p to 16 observations. We are given a vector of predictors and a vector of targets:
- F21: Paviani.
- F22: SineEnvelope.
- F23: Trefethen.
- F24: Alpine F2.
- F25: BiggsExp F5.
References
- Wu, G.; Mallipeddi, R.; Suganthan, P.N. Ensemble strategies for population-based optimization algorithms–A survey. Swarm Evol. Comput. 2019, 44, 695–711. [Google Scholar] [CrossRef]
- Vrugt, J.A.; Robinson, B.A. Improved evolutionary optimization from genetically adaptive multimethod search. Proc. Natl. Acad. Sci. USA 2007, 104, 708–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vrugt, J.A.; Robinson, B.A.; Hyman, J.M. Self-adaptive multimethod search for global optimization in real-parameter spaces. IEEE Trans. Evol. Comput. 2008, 13, 243–259. [Google Scholar] [CrossRef]
- Mashwani, W.K.; Salhi, A. Multiobjective evolutionary algorithm based on multimethod with dynamic resources allocation. Appl. Soft Comput. 2016, 39, 292–309. [Google Scholar] [CrossRef]
- Xue, Y.; Zhong, S.; Zhuang, Y.; Xu, B. An ensemble algorithm with self-adaptive learning techniques for high-dimensional numerical optimization. Appl. Math. Comput. 2014, 231, 329–346. [Google Scholar] [CrossRef]
- Price, D.; Radaideh, M.I. Animorphic ensemble optimization: A large-scale island model. Neural Comput. Appl. 2022, 35, 3221–3243. [Google Scholar] [CrossRef]
- Peng, F.; Tang, K.; Chen, G.; Yao, X. Population-based algorithm portfolios for numerical optimization. IEEE Trans. Evol. Comput. 2010, 14, 782–800. [Google Scholar] [CrossRef]
- Drake, J.H.; Kheiri, A.; Özcan, E.; Burke, E.K. Recent advances in selection hyper-heuristics. Eur. J. Oper. Res. 2020, 285, 405–428. [Google Scholar] [CrossRef]
- Grobler, J.; Engelbrecht, A.P.; Kendall, G.; Yadavalli, V.S. Multi-method algorithms: Investigating the entity-to-algorithm allocation problem. In Proceedings of the 2013 IEEE Congress on Evolutionary Computation, Cancun, Mexico, 20–23 June 2013; pp. 570–577. [Google Scholar]
- Du, W.; Li, B. Multi-strategy ensemble particle swarm optimization for dynamic optimization. Inf. Sci. 2008, 178, 3096–3109. [Google Scholar] [CrossRef]
- Wang, H.; Wu, Z.; Rahnamayan, S.; Sun, H.; Liu, Y.; Pan, J.s. Multi-strategy ensemble artificial bee colony algorithm. Inf. Sci. 2014, 279, 587–603. [Google Scholar] [CrossRef]
- Xiong, G.; Shi, D.; Duan, X. Multi-strategy ensemble biogeography-based optimization for economic dispatch problems. Appl. Energy 2013, 111, 801–811. [Google Scholar] [CrossRef]
- Trivedi, A.; Srinivasan, D.; Sanyal, K.; Ghosh, A. A Survey of Multiobjective Evolutionary Algorithms Based on Decomposition. IEEE Trans. Evol. Comput. 2017, 21, 440–462. [Google Scholar] [CrossRef]
- Hamza, N.M.; Essam, D.L.; Sarker, R.A. Constraint consensus mutation-based differential evolution for constrained optimization. IEEE Trans. Evol. Comput. 2015, 20, 447–459. [Google Scholar] [CrossRef]
- Ahmad, M.F.; Isa, N.A.M.; Lim, W.H.; Ang, K.M. Differential evolution: A recent review based on state-of-the-art works. Alex. Eng. J. 2022, 61, 3831–3872. [Google Scholar] [CrossRef]
- Gong, W.; Fialho, Á.; Cai, Z.; Li, H. Adaptive strategy selection in differential evolution for numerical optimization: An empirical study. Inf. Sci. 2011, 181, 5364–5386. [Google Scholar] [CrossRef]
- Mallipeddi, R.; Suganthan, P.N.; Pan, Q.K.; Tasgetiren, M.F. Differential evolution algorithm with ensemble of parameters and mutation strategies. Appl. Soft Comput. 2011, 11, 1679–1696. [Google Scholar] [CrossRef]
- Awad, N.H.; Ali, M.Z.; Suganthan, P.N. Ensemble of parameters in a sinusoidal differential evolution with niching-based population reduction. Swarm Evol. Comput. 2018, 39, 141–156. [Google Scholar] [CrossRef]
- Tanabe, R.; Fukunaga, A.S. Improving the search performance of SHADE using linear population size reduction. In Proceedings of the 2014 IEEE Congress on Evolutionary Computation (CEC), Beijing, China, 6–11 July 2014; pp. 1658–1665. [Google Scholar]
- Ali, M.Z.; Awad, N.H.; Suganthan, P.N.; Reynolds, R.G. An adaptive multipopulation differential evolution with dynamic population reduction. IEEE Trans. Cybern. 2016, 47, 2768–2779. [Google Scholar] [CrossRef]
- Wu, G.; Mallipeddi, R.; Suganthan, P.N.; Wang, R.; Chen, H. Differential evolution with multi-population based ensemble of mutation strategies. Inf. Sci. 2016, 329, 329–345. [Google Scholar] [CrossRef]
- Wu, G.; Shen, X.; Li, H.; Chen, H.; Lin, A.; Suganthan, P.N. Ensemble of differential evolution variants. Inf. Sci. 2018, 423, 172–186. [Google Scholar] [CrossRef]
- Yao, J.; Chen, Z.; Liu, Z. Improved ensemble of differential evolution variants. PLoS ONE 2021, 16, e0256206. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Dai, G.; Wang, M.; Liao, Z.; Ma, K. A two-stage ensemble of differential evolution variants for numerical optimization. IEEE Access 2019, 7, 56504–56519. [Google Scholar] [CrossRef]
- Tanabe, R.; Fukunaga, A. Success-history based parameter adaptation for differential evolution. In Proceedings of the 2013 IEEE Congress on Evolutionary Computation, Cancun, Mexico, 20–23 June 2013; pp. 71–78. [Google Scholar]
- Wang, X.; Li, C.; Zhu, J.; Meng, Q. L-SHADE-E: Ensemble of two differential evolution algorithms originating from L-SHADE. Inf. Sci. 2021, 552, 201–219. [Google Scholar] [CrossRef]
- Salcedo-Sanz, S.; Muñoz-Bulnes, J.; Vermeij, M.J. New coral reefs-based approaches for the model type selection problem: A novel method to predict a nation’s future energy demand. Int. J. Bio-Inspired Comput. 2017, 10, 145–158. [Google Scholar] [CrossRef]
- Salcedo-Sanz, S.; Camacho-Gómez, C.; Molina, D.; Herrera, F. A coral reefs optimization algorithm with substrate layers and local search for large scale global optimization. In Proceedings of the 2016 IEEE Congress on Evolutionary Computation (CEC), Vancouver, BC, Canada, 24–29 July 2016; pp. 3574–3581. [Google Scholar]
- Salcedo-Sanz, S. A review on the coral reefs optimization algorithm: New development lines and current applications. Prog. Artif. Intell. 2017, 6, 1–15. [Google Scholar] [CrossRef]
- Salcedo-Sanz, S.; Camacho-Gómez, C.; Mallol-Poyato, R.; Jiménez-Fernández, S.; Del Ser, J. A novel Coral Reefs Optimization algorithm with substrate layers for optimal battery scheduling optimization in micro-grids. Soft Comput. 2016, 20, 4287–4300. [Google Scholar] [CrossRef]
- Jiménez-Fernández, S.; Camacho-Gómez, C.; Mallol-Poyato, R.; Fernández, J.C.; Del Ser, J.; Portilla-Figueras, A.; Salcedo-Sanz, S. Optimal microgrid topology design and siting of distributed generation sources using a multi-objective substrate layer coral reefs optimization algorithm. Sustainability 2019, 11, 169. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Aracil, J.; Casillas-Pérez, D.; Jiménez-Fernández, S.; Prieto-Godino, L.; Salcedo-Sanz, S. A versatile multi-method ensemble for wind farm layout optimization. J. Wind. Eng. Ind. Aerodyn. 2022, 225, 104991. [Google Scholar] [CrossRef]
- Salcedo-Sanz, S.; Camacho-Gómez, C.; Magdaleno, A.; Pereira, E.; Lorenzana, A. Structures vibration control via tuned mass dampers using a co-evolution coral reefs optimization algorithm. J. Sound Vib. 2017, 393, 62–75. [Google Scholar] [CrossRef]
- Camacho-Gómez, C.; Wang, X.; Pereira, E.; Díaz, I.; Salcedo-Sanz, S. Active vibration control design using the Coral Reefs Optimization with Substrate Layer algorithm. Eng. Struct. 2018, 157, 14–26. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Aracil, J.; Camacho-Gómez, C.; Hernández-Díaz, A.M.; Pereira, E.; Salcedo-Sanz, S. Submerged Arches Optimal Design With a Multi-Method Ensemble Meta-Heuristic Approach. IEEE Access 2020, 8, 215057–215072. [Google Scholar] [CrossRef]
- Hernández-Díaz, A.M.; Pérez-Aracil, J.; Casillas-Perez, D.; Pereira, E.; Salcedo-Sanz, S. Hybridizing machine learning with metaheuristics for preventing convergence failures in mechanical models based on compression field theories. Appl. Soft Comput. 2022, 130, 109654. [Google Scholar] [CrossRef]
- Pérez-Aracil, J.; Camacho-Gómez, C.; Pereira, E.; Vaziri, V.; Aphale, S.S.; Salcedo-Sanz, S. Eliminating Stick-Slip Vibrations in Drill-Strings with a Dual-Loop Control Strategy Optimised by the CRO-SL Algorithm. Mathematics 2021, 9, 1526. [Google Scholar] [CrossRef]
- Sánchez-Montero, R.; Camacho-Gómez, C.; López-Espí, P.L.; Salcedo-Sanz, S. Optimal design of a planar textile antenna for industrial scientific medical (ISM) 2.4 GHz wireless body area networks (WBAN) with the CRO-SL algorithm. Sensors 2018, 18, 1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho-Gómez, C.; Marsa-Maestre, I.; Gimenez-Guzman, J.M.; Salcedo-Sanz, S. A Coral Reefs Optimization algorithm with substrate layer for robust Wi-Fi channel assignment. Soft Comput. 2019, 23, 12621–12640. [Google Scholar] [CrossRef]
- Camacho-Gomez, C.; Sanchez-Montero, R.; Martínez-Villanueva, D.; López-Espí, P.L.; Salcedo-Sanz, S. Design of a Multi-Band Microstrip Textile Patch Antenna for LTE and 5G Services with the CRO-SL Ensemble. Appl. Sci. 2020, 10, 1168. [Google Scholar] [CrossRef] [Green Version]
- Salcedo-Sanz, S.; Del Ser, J.; Landa-Torres, I.; Gil-López, S.; Portilla-Figueras, J. The coral reefs optimization algorithm: A novel metaheuristic for efficiently solving optimization problems. Sci. World J. 2014, 2014, 739768. [Google Scholar] [CrossRef] [Green Version]
- Del Ser, J.; Osaba, E.; Molina, D.; Yang, X.S.; Salcedo-Sanz, S.; Camacho, D.; Das, S.; Suganthan, P.N.; Coello, C.A.C.; Herrera, F. Bio-inspired computation: Where we stand and what’s next. Swarm Evol. Comput. 2019, 48, 220–250. [Google Scholar] [CrossRef]
- Kirkpatrick, S.; Gelatt Jr, C.D.; Vecchi, M.P. Optimization by simulated annealing. Science 1983, 220, 671–680. [Google Scholar] [CrossRef]
- Salcedo-Sanz, S.; Pastor-Sánchez, A.; Del Ser, J.; Prieto, L.; Geem, Z.W. A coral reefs optimization algorithm with harmony search operators for accurate wind speed prediction. Renew. Energy 2015, 75, 93–101. [Google Scholar] [CrossRef]
- Ahmed, S.; Ghosh, K.K.; Garcia-Hernandez, L.; Abraham, A.; Sarkar, R. Improved coral reefs optimization with adaptive β-hill climbing for feature selection. Neural Comput. Appl. 2021, 33, 6467–6486. [Google Scholar] [CrossRef]
- Leon, M.; Xiong, N. Investigation of mutation strategies in differential evolution for solving global optimization problems. In Artificial Intelligence and Soft Computing, Proceedings of the 13th International Conference on Artificial Intelligence and Soft Computing, Zakopane, Poland, 1–5 June 2014; Rutkowski, L., Korytkowski, M., Scherer, R., Tadeusiewicz, R., Zadeh, L.A., Zurada, J.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 8467, pp. 372–383. [Google Scholar]
- Xu, H.; Wen, J. Differential evolution algorithm for the optimization of the vehicle routing problem in logistics. In Proceedings of the 2012 8th International Conference on Computational Intelligence and Security, Guangzhou, China, 17–18 November 2012; pp. 48–51. [Google Scholar]
- Yang, X.S. Firefly algorithms for multimodal optimization. In Stochastic Algorithms: Foundations and Applications, Proceedings of the International Symposium on Stochastic Algorithms, Sapporo, Japan, 26–28 October 2009; Watanabe, O., Zeugmann, T., Eds.; Lecture Notes in Computer Science; Springer: Berlin/Heidelberg, Germany, 2009; Volume 5792, pp. 169–178. [Google Scholar]
- Yang, X.S.; Slowik, A. Firefly algorithm. In Swarm Intelligence Algorithms; Springer: Berlin/Heidelberg, Germany, 2020; pp. 163–174. [Google Scholar]
- Wang, H.; Wang, W.; Zhou, X.; Sun, H.; Zhao, J.; Yu, X.; Cui, Z. Firefly algorithm with neighborhood attraction. Inf. Sci. 2017, 382, 374–387. [Google Scholar] [CrossRef]
- Eiben, A.E.; Smith, J.E. Introduction to Evolutionary Computing; Springer: Berlin/Heidelberg, Germany, 2003; Volume 53. [Google Scholar]
- Herrera, F.; Lozano, M.; Pérez, E.; Sánchez, A.M.; Villar, P. Multiple crossover per couple with selection of the two best offspring: An experimental study with the BLX-α crossover operator for real-coded genetic algorithms. In Advances in Artificial Intelligence, Proceedings of the 8th Ibero-American Conference on Artificial Intelligence, Seville, Spain, 12–15 November 2002; Garijo, F.J., Riquelme, J.C., Toro, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; Volume 2527, pp. 392–401. [Google Scholar]
- Yao, X.; Liu, Y.; Lin, G. Evolutionary programming made faster. IEEE Trans. Evol. Comput. 1999, 3, 82–102. [Google Scholar]
- Wang, D.; Tan, D.; Liu, L. Particle swarm optimization algorithm: An overview. Soft Comput. 2018, 22, 387–408. [Google Scholar] [CrossRef]
- Baker, N.F.; Stanley, A.P.; Thomas, J.J.; Ning, A.; Dykes, K. Best practices for wake model and optimization algorithm selection in wind farm layout optimization. In Proceedings of the AIAA Scitech 2019 Forum, San Diego, CA, USA, 7–11 January 2019; p. 0540. [Google Scholar]
- Bortolotti, P.; Dykes, K.; Merz, K.; Sethuraman, L.; Verelst, D.; Zahle, F.; IEA Wind Task 37 on Systems Engineering in Wind Energy. WP2—Reference Wind Turbines. 2019. Available online: https://www.nrel.gov/wind/assets/pdfs/se17-9-iea-wind-task-37-systems-engineering.pdf (accessed on 5 March 2023).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function | DPCRO-SL | PCRO-SL | CRO-SL | ||||||
|---|---|---|---|---|---|---|---|---|---|
| # | Best | Mean | Std | Best | Mean | Std | Best | Mean | Std |
| F1 | |||||||||
| F2 | |||||||||
| F3 | |||||||||
| F4 | |||||||||
| F5 | |||||||||
| F6 | |||||||||
| F7 | |||||||||
| F8 | |||||||||
| F9 | |||||||||
| F10 | |||||||||
| F11 | |||||||||
| F12 | |||||||||
| F13 | |||||||||
| F14 | |||||||||
| F15 | |||||||||
| F16 | |||||||||
| F17 | |||||||||
| F18 | |||||||||
| F19 | |||||||||
| F20 | |||||||||
| F21 | |||||||||
| F22 | |||||||||
| F23 | |||||||||
| F24 | |||||||||
| F25 | |||||||||
| Function | DPCRO-SL | PSO | LSHADE | ||||||
|---|---|---|---|---|---|---|---|---|---|
| # | Best | Mean | Std | Best | Mean | Std | Best | Mean | Std |
| F1 | |||||||||
| F2 | |||||||||
| F3 | |||||||||
| F4 | |||||||||
| F5 | |||||||||
| F6 | |||||||||
| F7 | |||||||||
| F8 | |||||||||
| F9 | |||||||||
| F10 | |||||||||
| F11 | |||||||||
| F12 | |||||||||
| F13 | |||||||||
| F14 | |||||||||
| F15 | |||||||||
| F16 | |||||||||
| F17 | |||||||||
| F18 | |||||||||
| F19 | |||||||||
| F20 | |||||||||
| F21 | |||||||||
| F22 | |||||||||
| F23 | |||||||||
| F24 | |||||||||
| F25 | |||||||||
| Parameter | Value | Units |
|---|---|---|
| Rotor Diameter | 130 | m |
| Turbine Rating | 3.35 | MW |
| Cut-In Wind Speed | 4 | m/s |
| Rated Wind Speed | 9.8 | m/s |
| Cut-Out Wind Speed | 25 | m/s |
| Rank | Algorithm | Grad. | AEP |
|---|---|---|---|
| 1 | DPCRO-SL | GF | |
| 2 | SNOPT+WEC | G | |
| 3 | fmincon | G | |
| 4 | SNOPT | G | |
| 5 | SNOPT | G | |
| 6 | PSQP | G | |
| 7 | Multistart Interior-Point | G | |
| 8 | Full Pseudo-Gradient Approach | GF | |
| 9 | Basic Genetic Algorithm | GF | |
| 10 | Simple Particle Swarm Optimization | GF | |
| 11 | Simple Pseudo-Gradient Approach | GF |
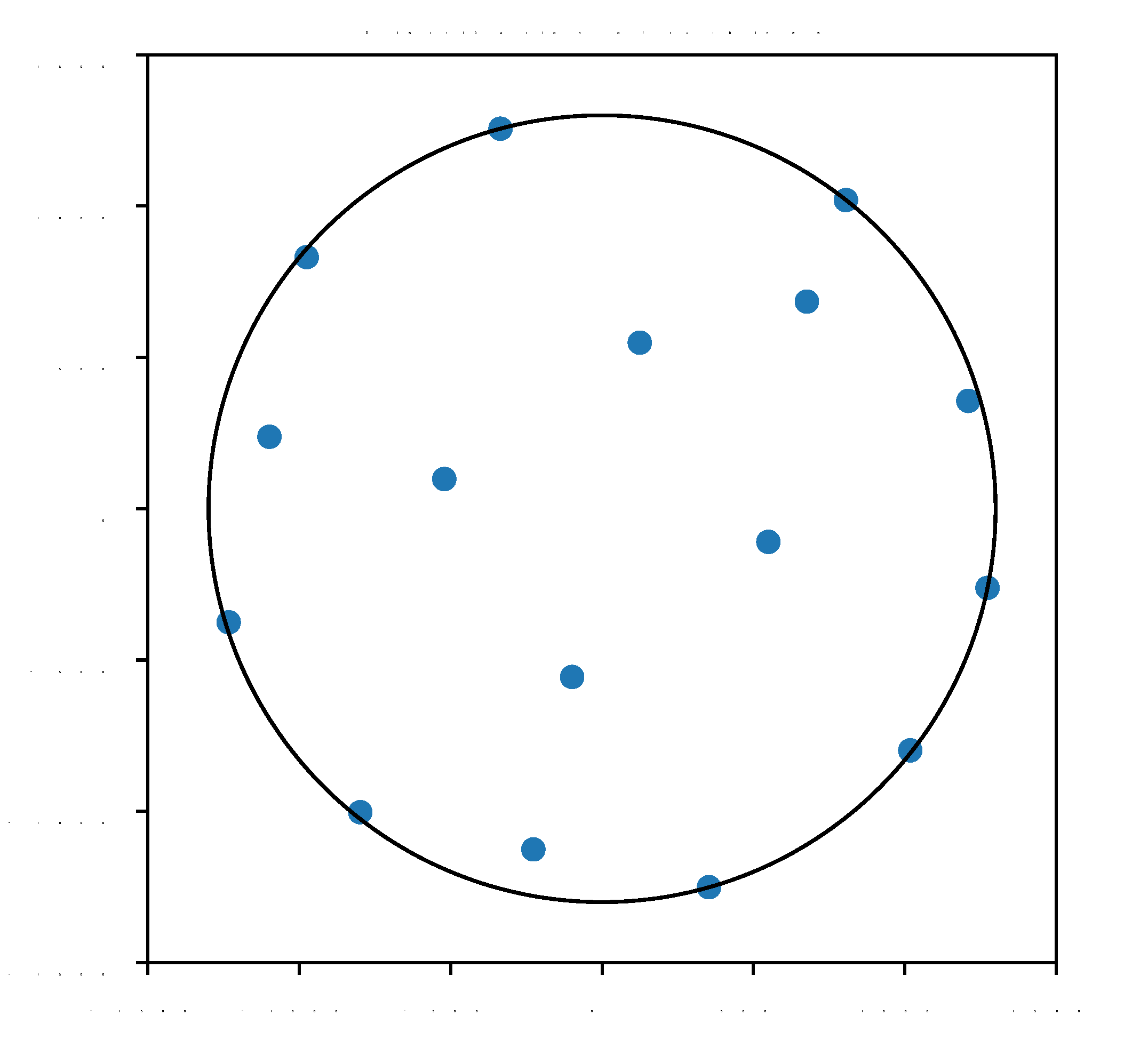
| i | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|
| x | −335.6 | 1273.3 | 1210.0 | −521.1 | -798.7 | −226.9 | 124.6 | 1018.1 |
| y | 1255.7 | −261.8 | 356.3 | 98.0 | −1003.0 | −1125.9 | 548.6 | −798.7 |
| i | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 |
| x | −1233.3 | −975.6 | 805.6 | 676.7 | −1098.8 | 549.4 | 353.1 | −98.7 |
| y | −375.5 | 831.4 | 1019.8 | 684.4 | 237.8 | −109.7 | −1250.9 | −556.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Aracil, J.; Camacho-Gómez, C.; Lorente-Ramos, E.; Marina, C.M.; Cornejo-Bueno, L.M.; Salcedo-Sanz, S. New Probabilistic, Dynamic Multi-Method Ensembles for Optimization Based on the CRO-SL. Mathematics 2023, 11, 1666. https://doi.org/10.3390/math11071666
Pérez-Aracil J, Camacho-Gómez C, Lorente-Ramos E, Marina CM, Cornejo-Bueno LM, Salcedo-Sanz S. New Probabilistic, Dynamic Multi-Method Ensembles for Optimization Based on the CRO-SL. Mathematics. 2023; 11(7):1666. https://doi.org/10.3390/math11071666
Chicago/Turabian StylePérez-Aracil, Jorge, Carlos Camacho-Gómez, Eugenio Lorente-Ramos, Cosmin M. Marina, Laura M. Cornejo-Bueno, and Sancho Salcedo-Sanz. 2023. "New Probabilistic, Dynamic Multi-Method Ensembles for Optimization Based on the CRO-SL" Mathematics 11, no. 7: 1666. https://doi.org/10.3390/math11071666