
An Automata-Based Cardiac Electrophysiology Simulator to Assess Arrhythmia Inducibility

 , , , , , , , and
, , , , , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Biophysical Modeling of Cardiac Cells
2.2. Biophysical Modeling of Cardiac Tissue
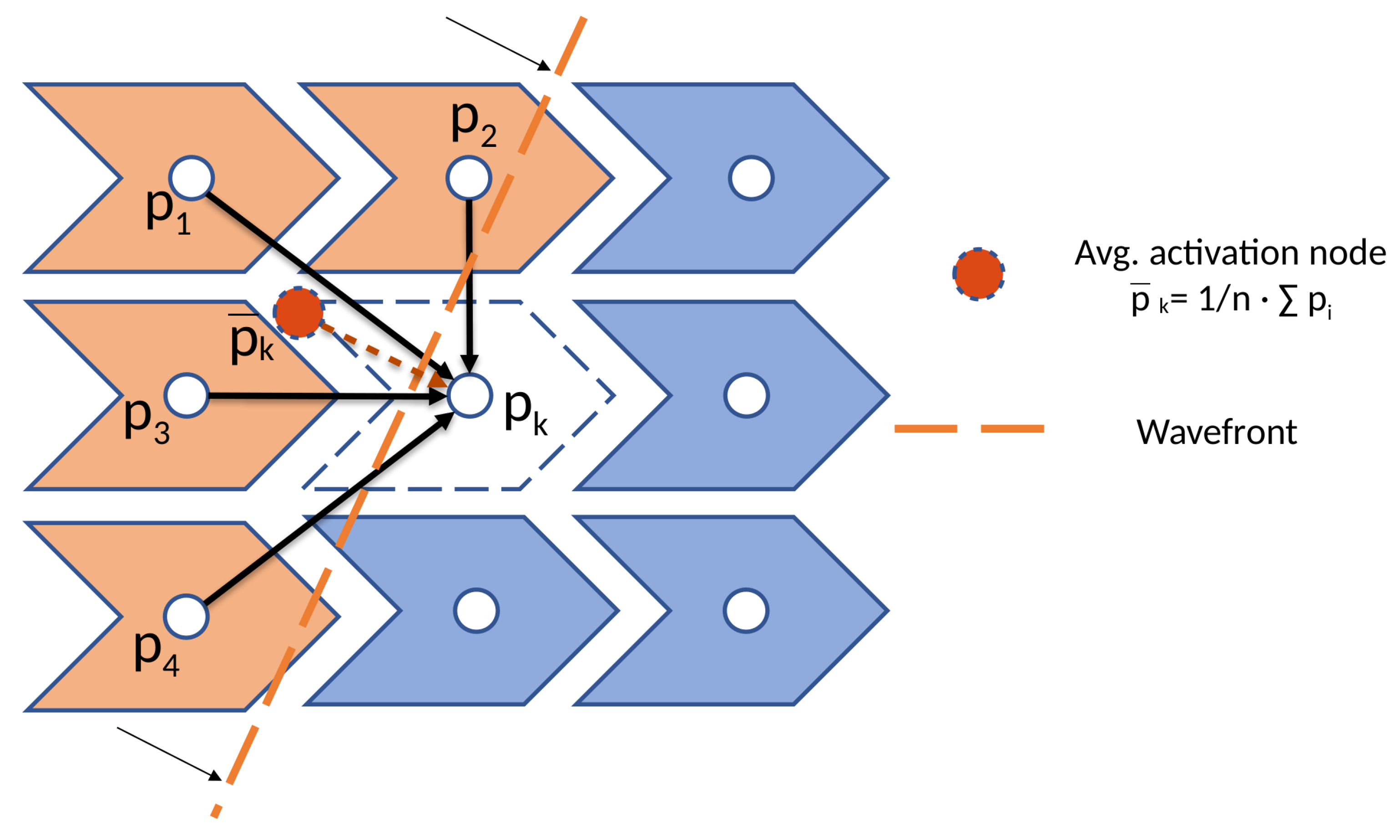
2.3. Cellular Automaton Electrophysiology Model
2.3.1. Cell Activation Model
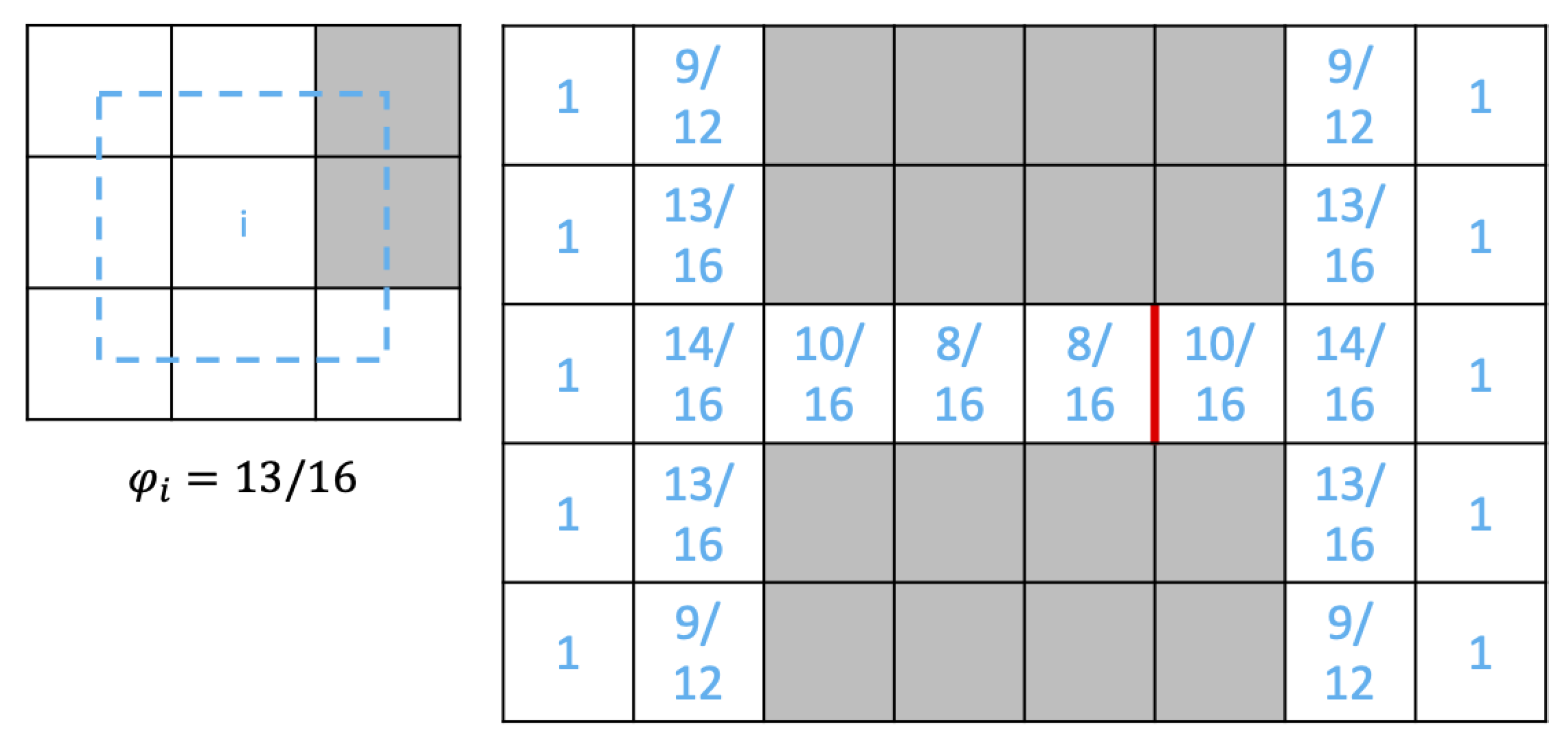
2.3.2. Tissue Modeling
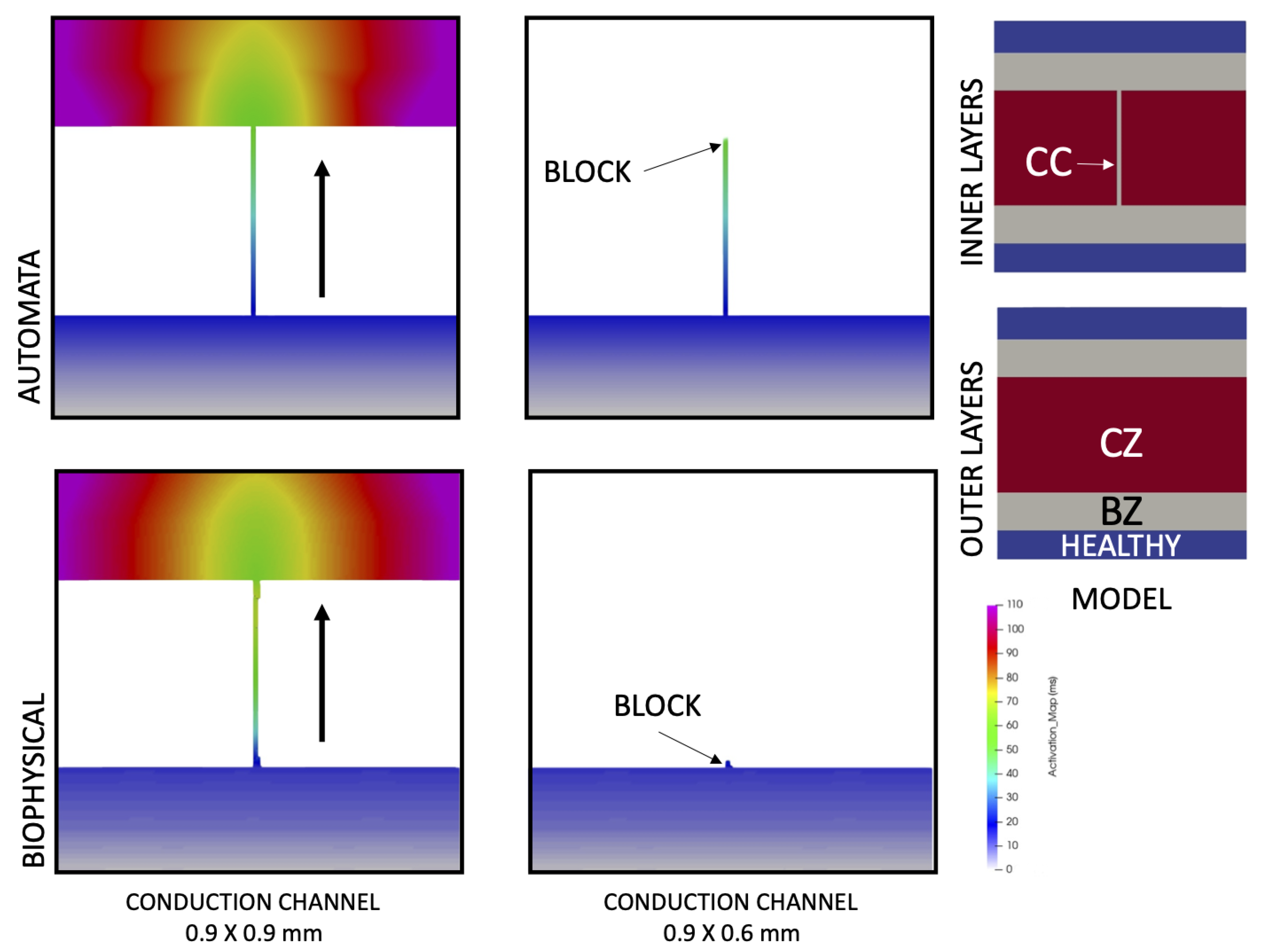
2.3.3. Safety Factor
| Algorithm 1 Activation propagation method. The reader can refer to the text for details on the notation. |
|
2.4. Geometry of the 3D Simulation Domains
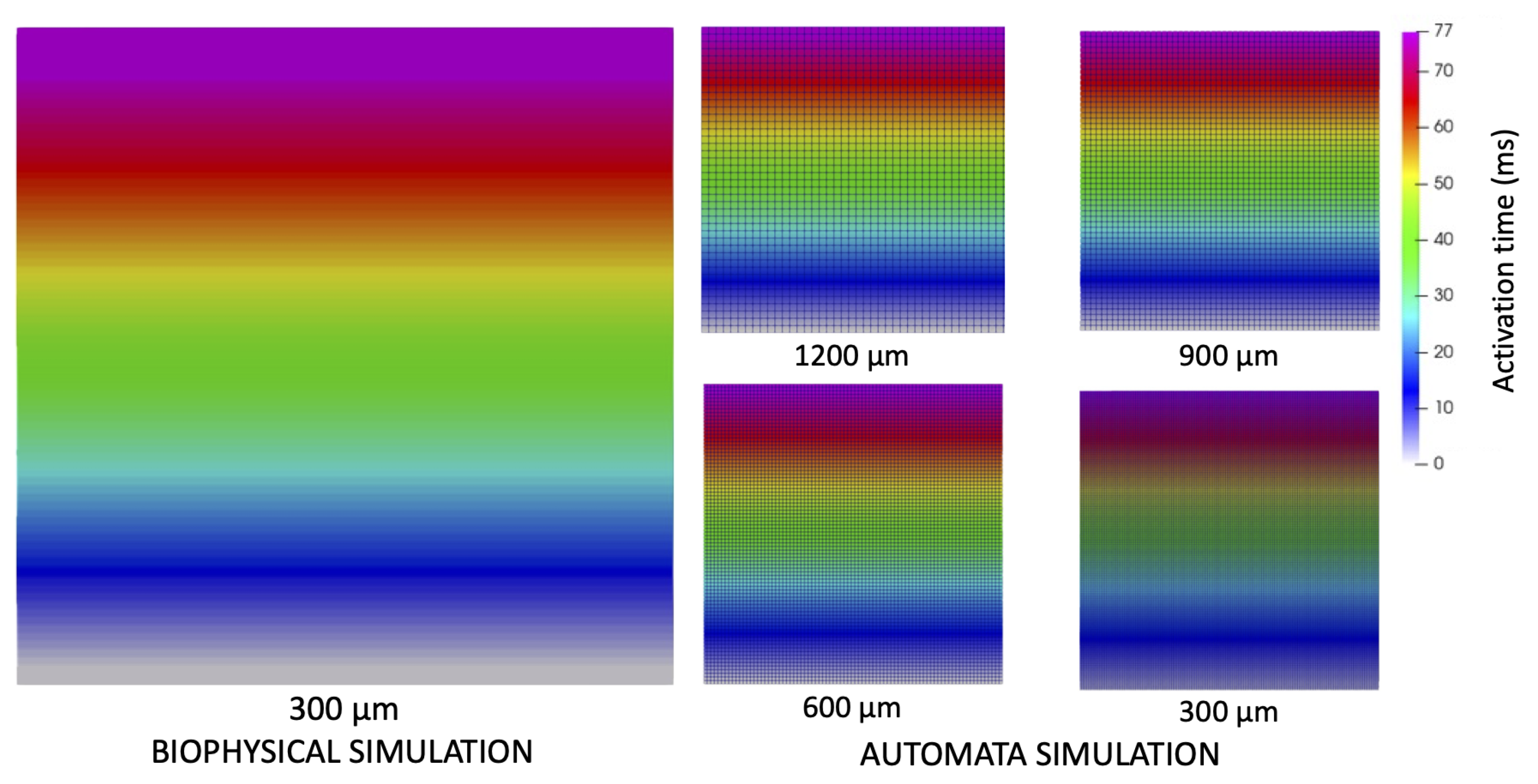
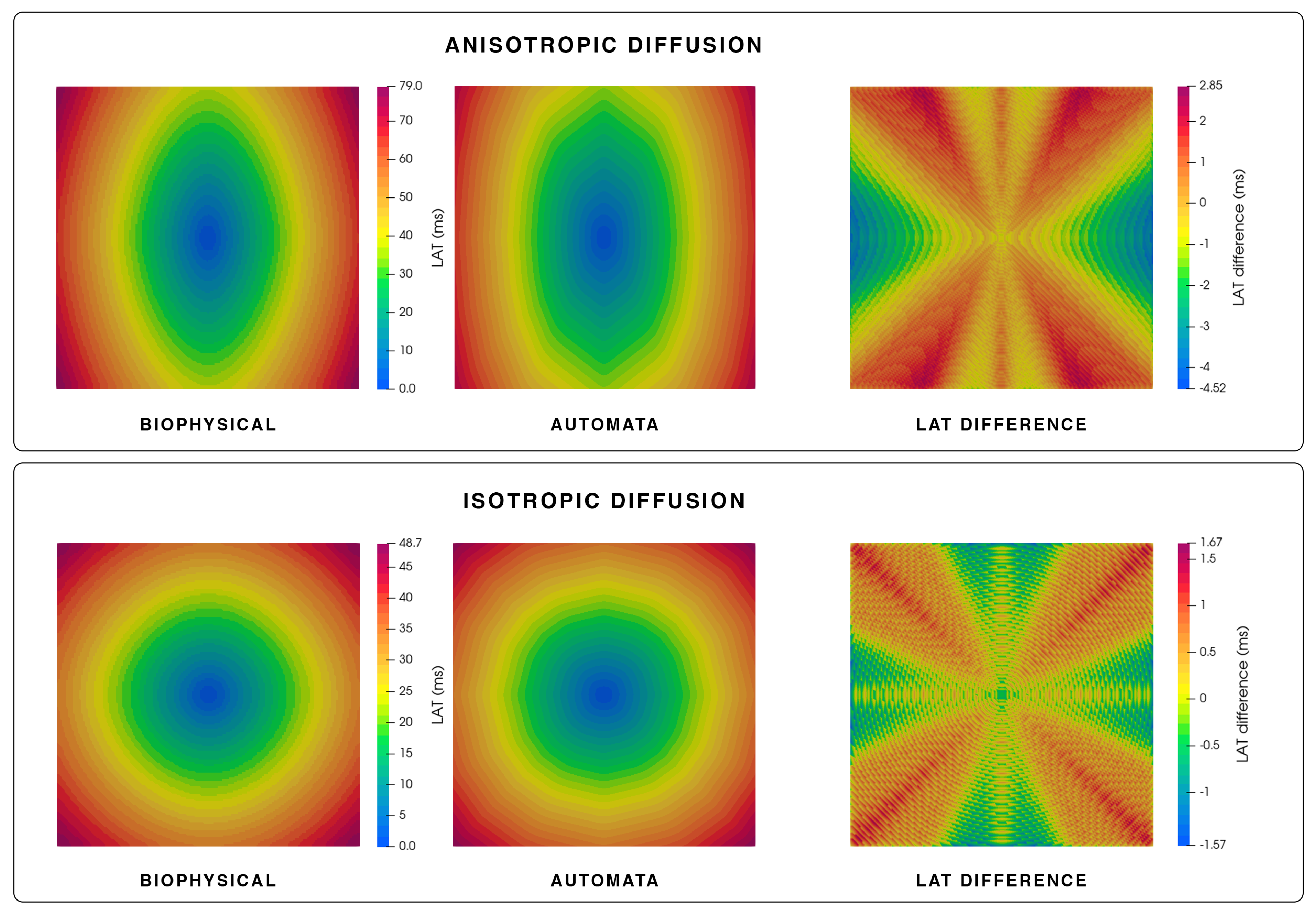
2.5. Biophysical Simulations in a Slab of Tissue
3. Results
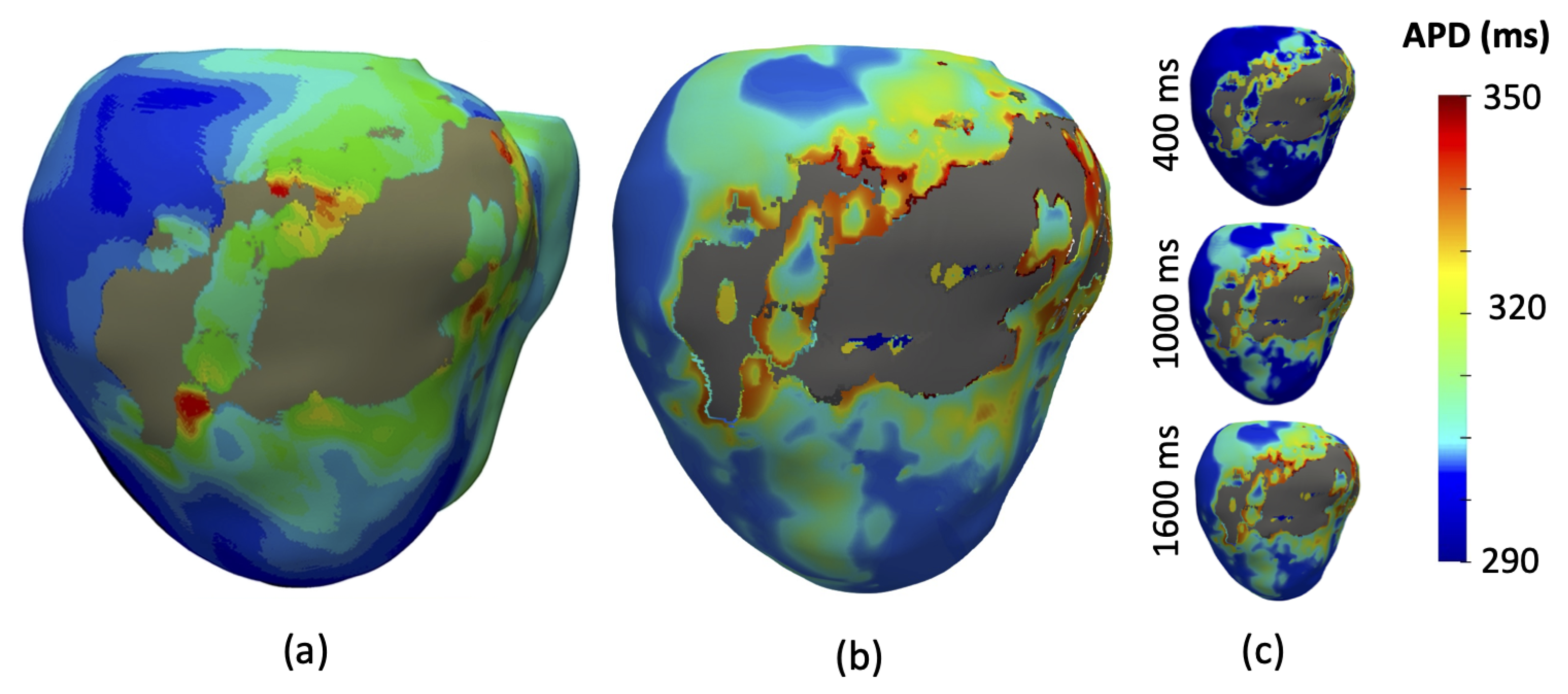
3.1. Simulation of Electrical Propagation
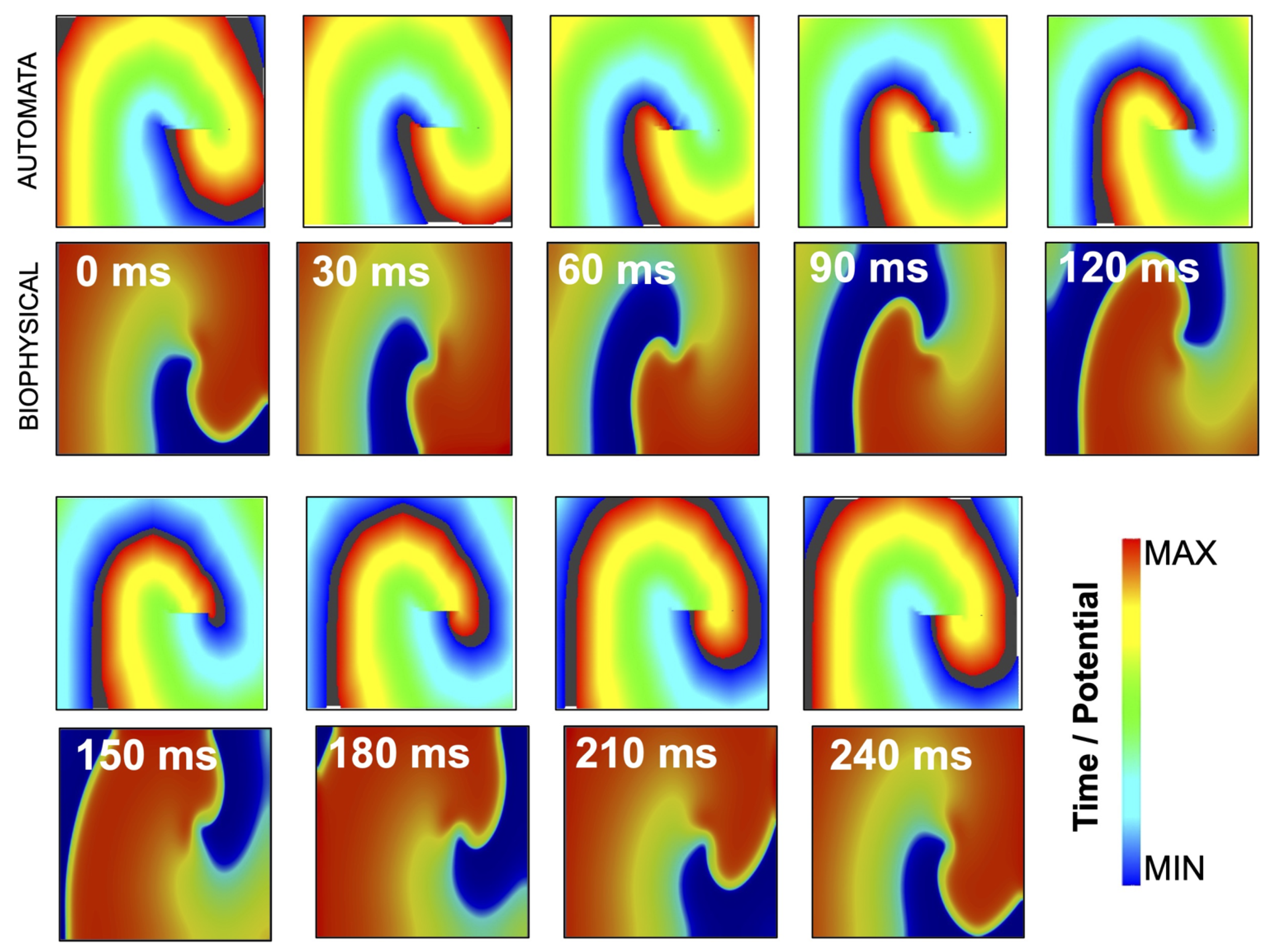
3.2. Simulation of Rotor Dynamics
3.3. Simulation of Safety Factor
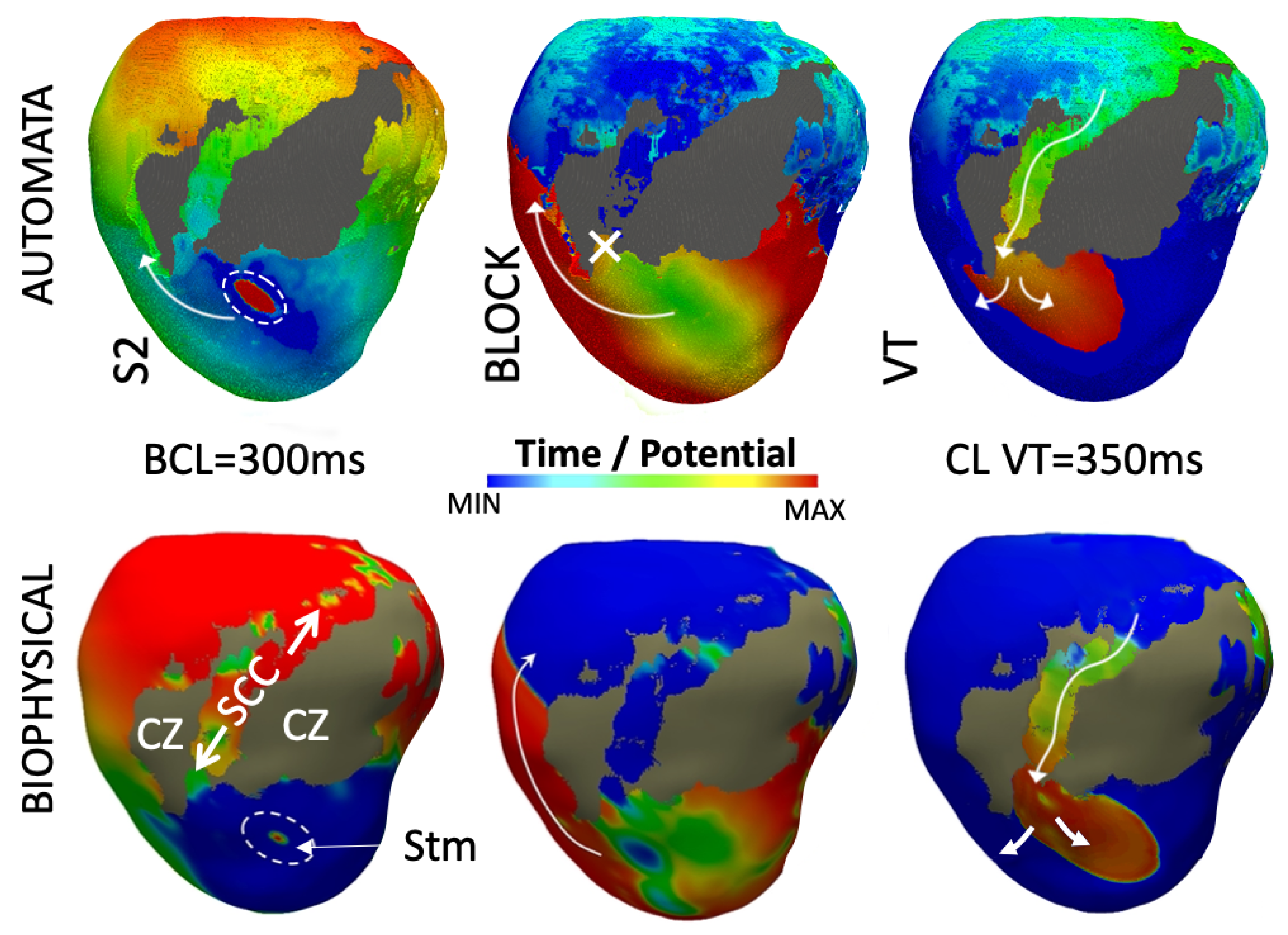
3.4. Simulation of Arrhythmia Dynamics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| APD(R) | Action potential duration (restitution) |
| (B)CL | (Basic) cycle length |
| BZ | Border zone |
| CA | Cellular automaton |
| CV(R) | Conduction velocity (restitution) |
| CZ | Core zone |
| DI | Diastolic interval |
| RFA | Radio-frequency ablation |
| SCC | Slow conduction channel |
| SCD | Sudden cardiac death |
| SF | Safety factor |
| VT | Ventricular tachycardia |
References
- Rudy, Y. From genome to physiome: Integrative models of cardiac excitation. Ann. Biomed. Eng. 2000, 28, 945–950. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Perez, A.; Sebastian, R.; Ferrero, J.M. Three-dimensional cardiac computational modelling: Methods, features and applications. Biomed. Eng. Online 2015, 14, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niederer, S.A.; Lumens, J.; Trayanova, N.A. Computational models in cardiology. Nat. Rev. Cardiol. 2019, 16, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Pollard, A.E.; Hooke, N.; Henriquez, C.S. Cardiac propagation simulation. Crit. Rev. Biomed. Eng. 1992, 20, 171–210. [Google Scholar] [PubMed]
- Lopez-Perez, A.; Sebastian, R.; Izquierdo, M.; Ruiz, R.; Bishop, M.; Ferrero, J.M. Personalized Cardiac Computational Models: From Clinical Data to Simulation of Infarct-Related Ventricular Tachycardia. Front. Physiol. 2019, 10, 580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidenreich, E.A.; Ferrero, J.M.; Doblaré, M.; Rodríguez, J.F. Adaptive macro finite elements for the numerical solution of monodomain equations in cardiac electrophysiology. Ann. Biomed. Eng. 2010, 38, 2331–2345. [Google Scholar] [CrossRef] [PubMed]
- Bernabeu, M.O.; Bordas, R.; Pathmanathan, P.; Pitt-Francis, J.; Cooper, J.; Garny, A.; Gavaghan, D.J.; Rodriguez, B.; Southern, J.A.; Whiteley, J.P. CHASTE: Incorporating a novel multi-scale spatial and temporal algorithm into a large-scale open source library. Philos. Trans. A Math. Phys. Eng. Sci. 2009, 367, 1907–1930. [Google Scholar] [CrossRef]
- Plank, G.; Loewe, A.; Neic, A.; Augustin, C.; Huang, Y.L.; Gsell, M.A.F.; Karabelas, E.; Nothstein, M.; Prassl, A.J.; Sánchez, J.; et al. The openCARP simulation environment for cardiac electrophysiology. Comput. Methods Programs Biomed. 2021, 208, 106223. [Google Scholar] [CrossRef]
- Talbot, H.; Marchesseau, S.; Duriez, C.; Sermesant, M.; Cotin, S.; Delingette, H. Towards an interactive electromechanical model of the heart. Interface Focus 2013, 3, 20120091. [Google Scholar] [CrossRef] [Green Version]
- Trayanova, N.A.; Pashakhanloo, F.; Wu, K.C.; Halperin, H.R. Imaging-Based Simulations for Predicting Sudden Death and Guiding Ventricular Tachycardia Ablation. Circ. Arrhythm. Electrophysiol. 2017, 10, e004743. [Google Scholar] [CrossRef]
- Trayanova, N.A.; Doshi, A.N.; Prakosa, A. How personalized heart modeling can help treatment of lethal arrhythmias: A focus on ventricular tachycardia ablation strategies in post-infarction patients. Wiley Interdiscip. Rev. Syst. Biol. Med. 2020, 12, e1477. [Google Scholar] [CrossRef] [PubMed]
- Mendonca Costa, C.; Gemmell, P.; Elliott, M.K.; Whitaker, J.; Campos, F.O.; Strocchi, M.; Neic, A.; Gillette, K.; Vigmond, E.; Plank, G.; et al. Determining anatomical and electrophysiological detail requirements for computational ventricular models of porcine myocardial infarction. Comput. Biol. Med. 2021, 141, 105061. [Google Scholar] [CrossRef] [PubMed]
- Relan, J.; Chinchapatnam, P.; Sermesant, M.; Rhode, K.; Ginks, M.; Delingette, H.; Rinaldi, C.A.; Razavi, R.; Ayache, N. Coupled personalization of cardiac electrophysiology models for prediction of ischaemic ventricular tachycardia. Interface Focus 2011, 1, 396–407. [Google Scholar] [CrossRef]
- Pathmanathan, P.; Gray, R.A. Verification of computational models of cardiac electro-physiology. Int. J. Numer. Method Biomed. Eng. 2014, 30, 525–544. [Google Scholar] [CrossRef] [PubMed]
- Arevalo, H.J.; Vadakkumpadan, F.; Guallar, E.; Jebb, A.; Malamas, P.; Wu, K.C.; Trayanova, N.A. Arrhythmia risk stratification of patients after myocardial infarction using personalized heart models. Nat. Commun. 2016, 7, 11437. [Google Scholar] [CrossRef] [PubMed]
- McDowell, K.S.; Arevalo, H.J.; Maleckar, M.M.; Trayanova, N.A. Susceptibility to arrhythmia in the infarcted heart depends on myofibroblast density. Biophys. J. 2011, 101, 1307–1315. [Google Scholar] [CrossRef] [Green Version]
- Corral-Acero, J.; Margara, F.; Marciniak, M.; Rodero, C.; Loncaric, F.; Feng, Y.; Gilbert, A.; Fernandes, J.F.; Bukhari, H.A.; Wajdan, A.; et al. The ‘Digital Twin’ to enable the vision of precision cardiology. Eur. Heart J. 2020, 41, 4556–4564. [Google Scholar] [CrossRef]
- Chen, Z.; Cabrera-Lozoya, R.; Relan, J.; Sohal, M.; Shetty, A.; Karim, R.; Delingette, H.; Gill, J.; Rhode, K.; Ayache, N.; et al. Biophysical Modeling Predicts Ventricular Tachycardia Inducibility and Circuit Morphology: A Combined Clinical Validation and Computer Modeling Approach. J. Cardiovasc. Electrophysiol. 2016, 27, 851–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, C.C.; Schaeffer, D.G. A two-current model for the dynamics of cardiac membrane. Bull. Math. Biol. 2003, 65, 767–793. [Google Scholar] [CrossRef] [Green Version]
- Hodgkin, A.L.; Huxley, A.F. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952, 117, 500–544. [Google Scholar] [CrossRef]
- ten Tusscher, K.H.W.J.; Noble, D.; Noble, P.J.; Panfilov, A.V. A model for human ventricular tissue. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, H1573–H1589. [Google Scholar] [CrossRef] [PubMed]
- Grandi, E.; Pasqualini, F.S.; Bers, D.M. A novel computational model of the human ventricular action potential and Ca transient. J. Mol. Cell. Cardiol. 2010, 48, 112–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hara, T.; Virág, L.; Varró, A.; Rudy, Y. Simulation of the undiseased human cardiac ventricular action potential: Model formulation and experimental validation. PLoS Comput. Biol. 2011, 7, e1002061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomek, J.; Bueno-Orovio, A.; Passini, E.; Zhou, X.; Minchole, A.; Britton, O.; Bartolucci, C.; Severi, S.; Shrier, A.; Virag, L.; et al. Development, calibration, and validation of a novel human ventricular myocyte model in health, disease, and drug block. eLife 2019, 8, e48890. [Google Scholar] [CrossRef] [PubMed]
- Antzelevitch, C.; Shimizu, W.; Yan, G.X.; Sicouri, S.; Weissenburger, J.; Nesterenko, V.V.; Burashnikov, A.; Di Diego, J.; Saffitz, J.; Thomas, G.P. The M cell: Its contribution to the ECG and to normal and abnormal electrical function of the heart. J. Cardiovasc. Electrophysiol. 1999, 10, 1124–1152. [Google Scholar] [CrossRef]
- Boyett, M.R.; Jewell, B.R. A study of the factors responsible for rate-dependent shortening of the action potential in mammalian ventricular muscle. J. Physiol. 1978, 285, 359–380. [Google Scholar] [CrossRef] [PubMed]
- Simurda, J.; Simurdová, M.; Pásek, M.; Bravený, P. Quantitative analysis of cardiac electrical restitution. Eur. Biophys. J. 2001, 30, 500–514. [Google Scholar] [CrossRef]
- Coveney, S.; Corrado, C.; Oakley, J.E.; Wilkinson, R.D.; Niederer, S.A.; Clayton, R.H. Bayesian Calibration of Electrophysiology Models Using Restitution Curve Emulators. Front. Physiol. 2021, 12, 693015. [Google Scholar] [CrossRef]
- Cao, J.M.; Qu, Z.; Kim, Y.H.; Wu, T.J.; Garfinkel, A.; Weiss, J.N.; Karagueuzian, H.S.; Chen, P.S. Spatiotemporal heterogeneity in the induction of ventricular fibrillation by rapid pacing: Importance of cardiac restitution properties. Circ. Res. 1999, 84, 1318–1331. [Google Scholar] [CrossRef] [Green Version]
- Dvir, H.; Zlochiver, S. Stochastic cardiac pacing increases ventricular electrical stability—A computational study. Biophys. J. 2013, 105, 533–542. [Google Scholar] [CrossRef] [Green Version]
- Tusscher, K.H.T.; Panfilov, A.V. Cell model for efficient simulation of wave propagation in human ventricular tissue under normal and pathological conditions. Phys. Med. Biol. 2006, 51, 6141–6156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geselowitz, D.B.; Miller, W.T., 3rd. A bidomain model for anisotropic cardiac muscle. Ann. Biomed. Eng. 1983, 11, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Sethian, J.A. Fast Marching Methods. SIAM Rev. 1999, 41, 199–235. [Google Scholar] [CrossRef]
- Bueno-Orovio, A.; Cherry, E.M.; Fenton, F.H. Minimal model for human ventricular action potentials in tissue. J. Theor. Biol. 2008, 253, 544–560. [Google Scholar] [CrossRef] [PubMed]
- Boyle, P.M.; Vigmond, E.J. An intuitive safety factor for cardiac propagation. Biophys. J. 2010, 98, L57–L59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyle, P.M.; Franceschi, W.H.; Constantin, M.; Hawks, C.; Desplantez, T.; Trayanova, N.A.; Vigmond, E.J. New insights on the cardiac safety factor: Unraveling the relationship between conduction velocity and robustness of propagation. J. Mol. Cell. Cardiol. 2019, 128, 117–128. [Google Scholar] [CrossRef]
- Godoy, E.J.; Lozano, M.; García-Fernández, I.; Ferrer-Albero, A.; MacLeod, R.; Saiz, J.; Sebastian, R. Atrial fibrosis hampers non-invasive localization of atrial ectopic foci from multi-electrode signals: A 3d simulation study. Front. Physiol. 2018, 9, 404. [Google Scholar] [CrossRef] [Green Version]
- Gouvêa de Barros, B.; Sachetto Oliveira, R.; Meira, W., Jr.; Lobosco, M.; Weber dos Santos, R. Simulations of complex and microscopic models of cardiac electrophysiology powered by multi-GPU platforms. Comput. Math. Methods Med. 2012, 2012, 824569. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.C.; DeMarco, K.R.; Aghasafari, P.; Jeng, M.T.; Dawson, J.R.; Bekker, S.; Noskov, S.Y.; Yarov-Yarovoy, V.; Vorobyov, I.; Clancy, C.E. A computational pipeline to predict cardiotoxicity: From the atom to the rhythm. Circ. Res. 2020, 126, 947–964. [Google Scholar] [CrossRef]
- Maleckar, M.M.; Myklebust, L.; Uv, J.; Florvaag, P.M.; Strøm, V.; Glinge, C.; Jabbari, R.; Vejlstrup, N.; Engstrøm, T.; Ahtarovski, K.; et al. Combined In-silico and Machine Learning Approaches Toward Predicting Arrhythmic Risk in Post-infarction Patients. Front. Physiol. 2021, 12, 745349. [Google Scholar] [CrossRef]
- Zhou, S.; AbdelWahab, A.; Sapp, J.L.; Sung, E.; Aronis, K.N.; Warren, J.W.; MacInnis, P.J.; Shah, R.; Horáček, B.M.; Berger, R.; et al. Prospective Multicenter Assessment of a New Intraprocedural Automated System for Localizing Idiopathic Ventricular Arrhythmia Origins. JACC Clin. Electrophysiol. 2021, 7, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; AbdelWahab, A.; Horáček, B.M.; MacInnis, P.J.; Warren, J.W.; Davis, J.S.; Elsokkari, I.; Lee, D.C.; MacIntyre, C.J.; Parkash, R.; et al. Prospective Assessment of an Automated Intraprocedural 12-Lead ECG-Based System for Localization of Early Left Ventricular Activation. Circ. Arrhythm. Electrophysiol. 2020, 13, e008262. [Google Scholar] [CrossRef] [PubMed]
- Vigmond, E.J.; Hughes, M.; Plank, G.; Leon, L.J. Computational tools for modeling electrical activity in cardiac tissue. J. Electrocardiol. 2003, 36 (Suppl. 1), 69–74. [Google Scholar] [CrossRef] [PubMed]
- Bradley, C.; Bowery, A.; Britten, R.; Budelmann, V.; Camara, O.; Christie, R.; Cookson, A.; Frangi, A.F.; Gamage, T.B.; Heidlauf, T.; et al. OpenCMISS: A multi-physics & multi-scale computational infrastructure for the VPH/Physiome project. Prog. Biophys. Mol. Biol. 2011, 107, 32–47. [Google Scholar] [CrossRef] [PubMed]
- Niederer, S.A.; Kerfoot, E.; Benson, A.P.; Bernabeu, M.O.; Bernus, O.; Bradley, C.; Cherry, E.M.; Clayton, R.; Fenton, F.H.; Garny, A.; et al. Verification of cardiac tissue electrophysiology simulators using an N-version benchmark. Philos. Trans. A Math. Phys. Eng. Sci. 2011, 369, 4331–4351. [Google Scholar] [CrossRef] [Green Version]
- Adabag, A.S.; Therneau, T.M.; Gersh, B.J.; Weston, S.A.; Roger, V.L. Sudden death after myocardial infarction. JAMA 2008, 300, 2022–2029. [Google Scholar] [CrossRef] [Green Version]
- Fern’andez-Armenta, J.; Berruezo, A.; Andreu, D.; Camara, O.; Silva, E.; Serra, L.; Barbarito, V.; Carotenutto, L.; Evertz, R.; Ortiz-P’erez, J.T.; et al. Three-dimensional architecture of scar and conducting channels based on high resolution ce-CMR: Insights for ventricular tachycardia ablation. Circ. Arrhythm. Electrophysiol. 2013, 6, 528–537. [Google Scholar] [CrossRef] [Green Version]
- Soto-Iglesias, D.; Penela, D.; J’auregui, B.; Acosta, J.; Fern’andez-Armenta, J.; Linhart, M.; Zucchelli, G.; Syrovnev, V.; Zaraket, F.; Ter’es, C.; et al. Cardiac Magnetic Resonance-Guided Ventricular Tachycardia Substrate Ablation. JACC Clin. Electrophysiol. 2020, 6, 436–447. [Google Scholar] [CrossRef]
- Arevalo, H.; Plank, G.; Helm, P.; Halperin, H.; Trayanova, N. Tachycardia in post-infarction hearts: Insights from 3D image-based ventricular models. PLoS ONE 2013, 8, e68872. [Google Scholar] [CrossRef] [Green Version]
- Deng, D.; Prakosa, A.; Shade, J.; Nikolov, P.; Trayanova, N.A. Characterizing Conduction Channels in Postinfarction Patients Using a Personalized Virtual Heart. Biophys. J. 2019, 117, 2287–2294. [Google Scholar] [CrossRef]
- Prakosa, A.; Arevalo, H.J.; Deng, D.; Boyle, P.M.; Nikolov, P.P.; Ashikaga, H.; Blauer, J.J.E.; Ghafoori, E.; Park, C.J.; Blake, R.C., 3rd; et al. Personalized virtual-heart technology for guiding the ablation of infarct-related ventricular tachycardia. Nat. Biomed. Eng. 2018, 2, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Vigmond, E.J.; Boyle, P.M.; Leon, L.; Plank, G. Near-real-time simulations of biolelectric activity in small mammalian hearts using graphical processing units. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2009, 2009, 3290–3293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigueras, G.; Roy, I.; Cookson, A.; Lee, J.; Smith, N.; Nordsletten, D. Toward GPGPU accelerated human electromechanical cardiac simulations. Int. J. Numer. Method Biomed. Eng. 2014, 30, 117–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachetto Oliveira, R.; Martins Rocha, B.; Burgarelli, D.; Meira, W., Jr.; Constantinides, C.; Weber Dos Santos, R. Performance evaluation of GPU parallelization, space-time adaptive algorithms, and their combination for simulating cardiac electrophysiology. Int. J. Numer. Method Biomed. Eng. 2018, 34, e2913. [Google Scholar] [CrossRef]
- Garcia-Molla, V.M.; Liberos, A.; Vidal, A.; Guillem, M.S.; Millet, J.; Gonzalez, A.; Martinez-Zaldivar, F.J.; Climent, A.M. Adaptive step ODE algorithms for the 3D simulation of electric heart activity with graphics processing units. Comput. Biol. Med. 2014, 44, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Pashaei, A.; Romero, D.; Sebastian, R.; Camara, O.; Frangi, A.F. Fast multiscale modeling of cardiac electrophysiology including Purkinje system. IEEE Trans. Biomed. Eng. 2011, 58, 2956–2960. [Google Scholar] [CrossRef]
- Cedilnik, N.; Duchateau, J.; Dubois, R.; Sacher, F.; Jaïs, P.; Cochet, H.; Sermesant, M. Fast personalized electrophysiological models from computed tomography images for ventricular tachycardia ablation planning. Europace 2018, 20, iii94–iii101. [Google Scholar] [CrossRef] [PubMed]
- Quaglino, A.; Pezzuto, S.; Koutsourelakis, P.S.; Auricchio, A.; Krause, R. Fast uncertainty quantification of activation sequences in patient-specific cardiac electrophysiology meeting clinical time constraints. Int. J. Numer. Method Biomed. Eng. 2018, 34, e2985. [Google Scholar] [CrossRef]
- Sermesant, M.; Konukoglu, E.; Delingette, H.; Coudiere, Y.; Chinchapatnam, P.; Rhode, K.S.; Razavi, R.; Ayache, N. An anisotropic multi-front fast marching method for real-time simulation of cardiac electrophysiology. In Proceedings of the International Conference on Functional Imaging and Modeling of the Heart, Salt Lake City, UT, USA, 7–9 June 2007; pp. 160–169. [Google Scholar]
- Loewe, A.; Poremba, E.; Oesterlein, T.; Luik, A.; Schmitt, C.; Seemann, G.; Dössel, O. Patient-Specific Identification of Atrial Flutter Vulnerability-A Computational Approach to Reveal Latent Reentry Pathways. Front. Physiol. 2018, 9, 1910. [Google Scholar] [CrossRef] [Green Version]
- Neic, A.; Campos, F.O.; Prassl, A.J.; Niederer, S.A.; Bishop, M.J.; Vigmond, E.J.; Plank, G. Efficient computation of electrograms and ECGs in human whole heart simulations using a reaction-eikonal model. J. Comput. Phys. 2017, 346, 191–211. [Google Scholar] [CrossRef]
- Saxberg, B.E.; Cohen, R.J. Cellular automata models for reentrant arrhythmias. J. Electrocardiol. 1990, 23, 95. [Google Scholar] [CrossRef]
- Siregar, P.; Sinteff, J.P.; Julen, N.; Le Beux, P. An interactive 3D anisotropic cellular automata model of the heart. Comput. Biomed. Res. 1998, 31, 323–347. [Google Scholar] [CrossRef] [PubMed]
- Werner, C.; Sachse, F.; Dössel, O. Electrical excitation propagation in the human heart. Int. J. Bioelectromagn. 2000, 2, 96–117. [Google Scholar]
- Zhu, H.; Sun, Y.; Rajagopal, G.; Mondry, A.; Dhar, P. Facilitating arrhythmia simulation: The method of quantitative cellular automata modeling and parallel running. Biomed. Eng. Online 2004, 3, 29. [Google Scholar] [CrossRef] [Green Version]
- Sabzpoushan, S.H.; Pourhasanzade, F. A Cellular Automata-based Model for Simulating Restitution Property in a Single Heart Cell. J. Med. Signals Sens. 2011, 1, 19–23. [Google Scholar]
- Corrado, C.; Zemzemi, N. A conduction velocity adapted eikonal model for electrophysiology problems with re-excitability evaluation. Med. Image Anal. 2018, 43, 186–197. [Google Scholar] [CrossRef] [Green Version]
- Ai, W.; Patel, N.D.; Roop, P.S.; Malik, A.; Trew, M.L. Cardiac Electrical Modeling for Closed-Loop Validation of Implantable Devices. IEEE Trans. Biomed. Eng. 2020, 67, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Bueno-Orovio, A.; Hanson, B.M.; Gill, J.S.; Taggart, P.; Rodriguez, B. In vivo human left-to-right ventricular differences in rate adaptation transiently increase pro-arrhythmic risk following rate acceleration. PLoS ONE 2012, 7, e52234. [Google Scholar] [CrossRef] [Green Version]
- Campos, R.S.; Silva, J.G.R.; Barbosa, H.J.C.; Santos, R.W.d. Electrotonic Effect on Action Potential Dispersion with Cellular Automata. In Proceedings of the International Conference on Computational Science and Its Applications, Cagliari, Italy, 1–4 July 2020; pp. 205–215. [Google Scholar]
- Dux-Santoy, L.; Sebastian, R.; Felix-Rodriguez, J.; Ferrero, J.M.; Saiz, J. Interaction of specialized cardiac conduction system with antiarrhythmic drugs: A simulation study. IEEE Trans. Biomed. Eng. 2011, 58, 3475–3478. [Google Scholar] [CrossRef]
- Ciaccio, E.J.; Coromilas, J.; Wit, A.L.; Peters, N.S.; Garan, H. Source-Sink Mismatch Causing Functional Conduction Block in Re-Entrant Ventricular Tachycardia. JACC Clin. Electrophysiol. 2018, 4, 1–16. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serra, D.; Romero, P.; Garcia-Fernandez, I.; Lozano, M.; Liberos, A.; Rodrigo, M.; Bueno-Orovio, A.; Berruezo, A.; Sebastian, R. An Automata-Based Cardiac Electrophysiology Simulator to Assess Arrhythmia Inducibility. Mathematics 2022, 10, 1293. https://doi.org/10.3390/math10081293
Serra D, Romero P, Garcia-Fernandez I, Lozano M, Liberos A, Rodrigo M, Bueno-Orovio A, Berruezo A, Sebastian R. An Automata-Based Cardiac Electrophysiology Simulator to Assess Arrhythmia Inducibility. Mathematics. 2022; 10(8):1293. https://doi.org/10.3390/math10081293
Chicago/Turabian StyleSerra, Dolors, Pau Romero, Ignacio Garcia-Fernandez, Miguel Lozano, Alejandro Liberos, Miguel Rodrigo, Alfonso Bueno-Orovio, Antonio Berruezo, and Rafael Sebastian. 2022. "An Automata-Based Cardiac Electrophysiology Simulator to Assess Arrhythmia Inducibility" Mathematics 10, no. 8: 1293. https://doi.org/10.3390/math10081293