
Curcumin and Osteosarcoma: Can Invertible Polymeric Micelles Help?


Abstract
:1. Introduction
1.1. Curcumin and Its Molecular Targets
1.2. Curcumin-Induced Apoptosis
1.3. Notch-1 Signaling Mechanism
1.4. Effects of Curcumin on Normal Cells
2. Polymeric Micelles as a Platform for Drug Delivery
3. Invertible Polymer Micelles as a Safe Way to Treat Osteosarcoma
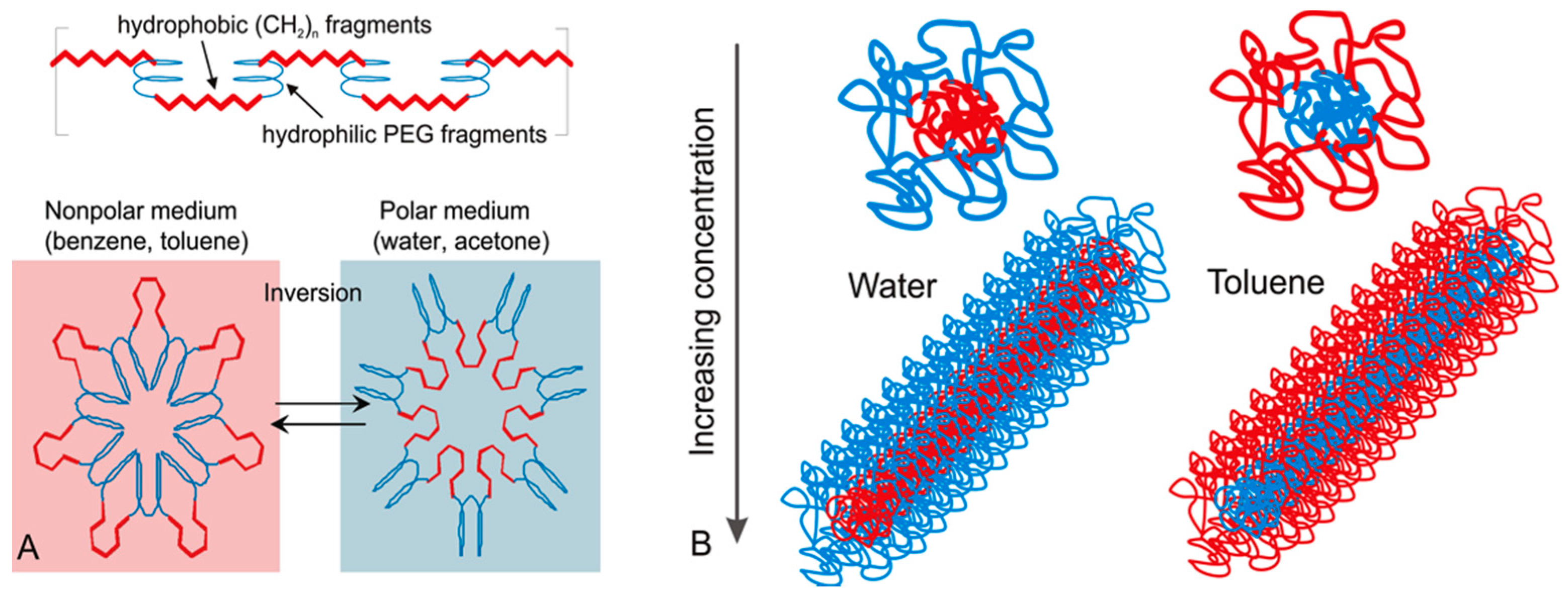
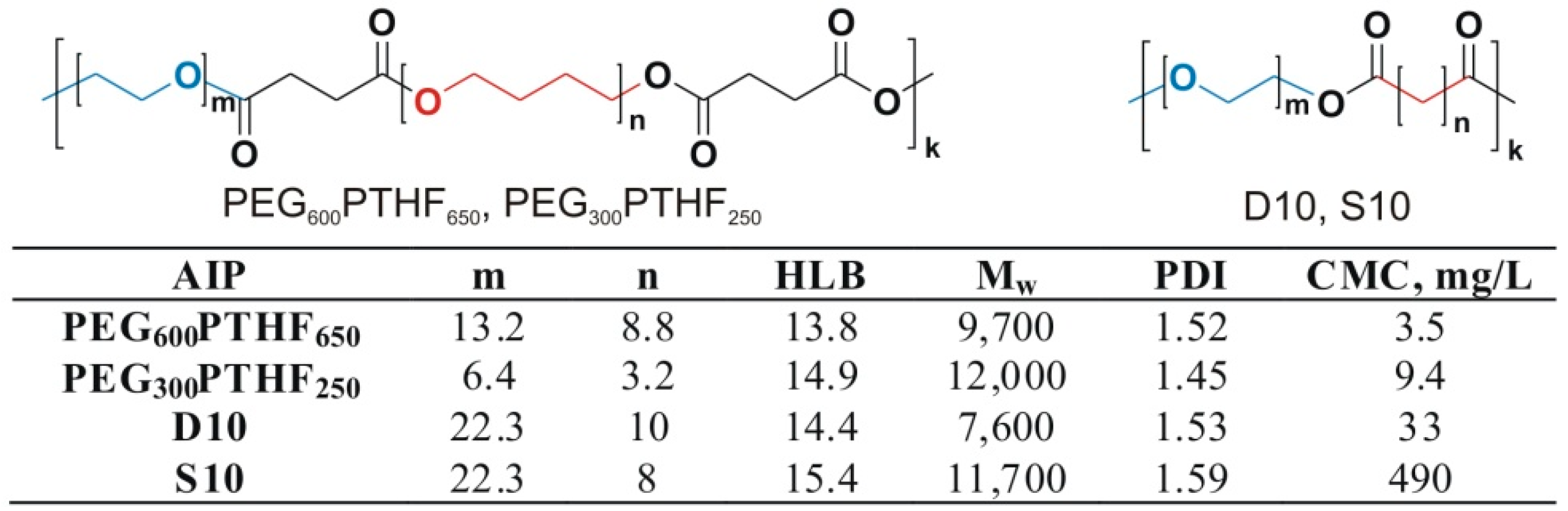
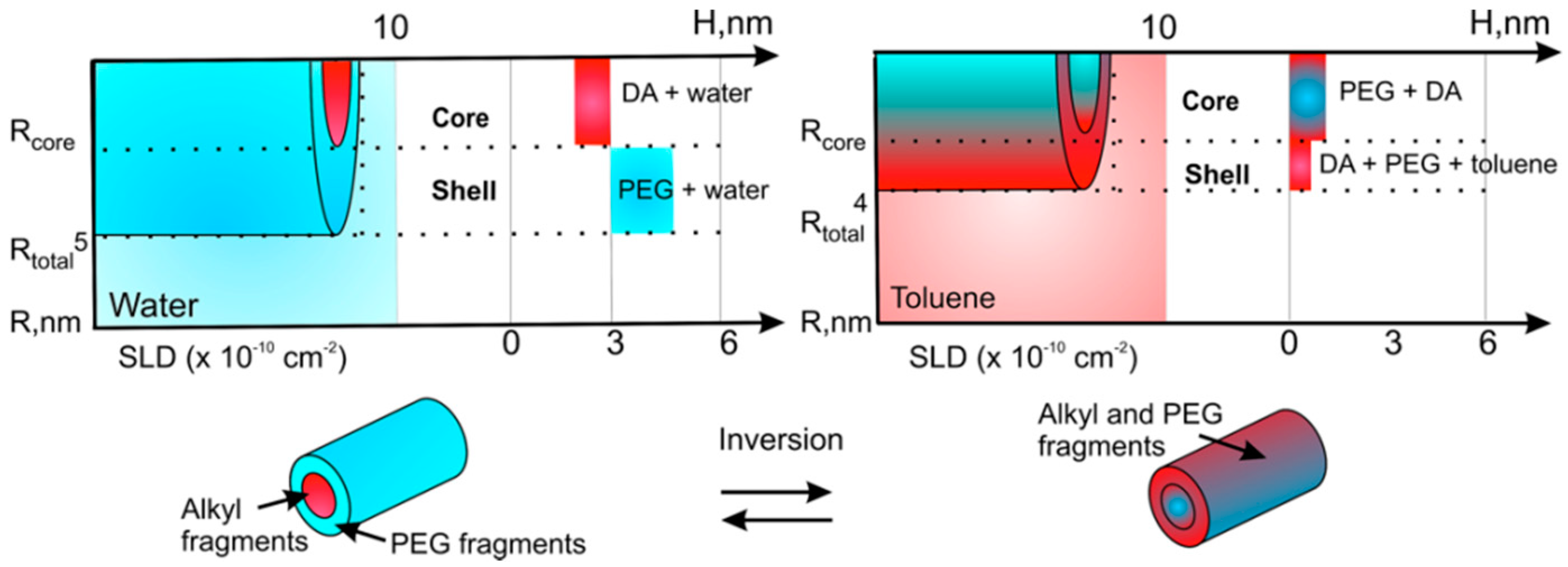
3.1. Self-Assembly of Invertible Macromolecules
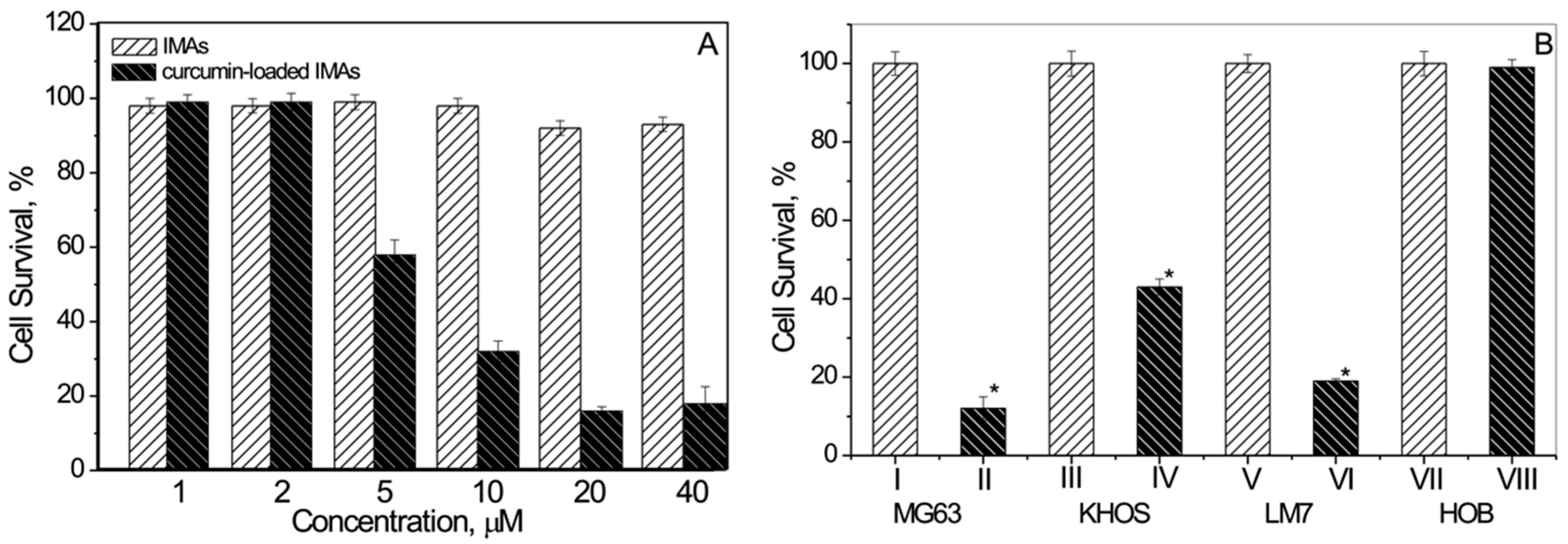
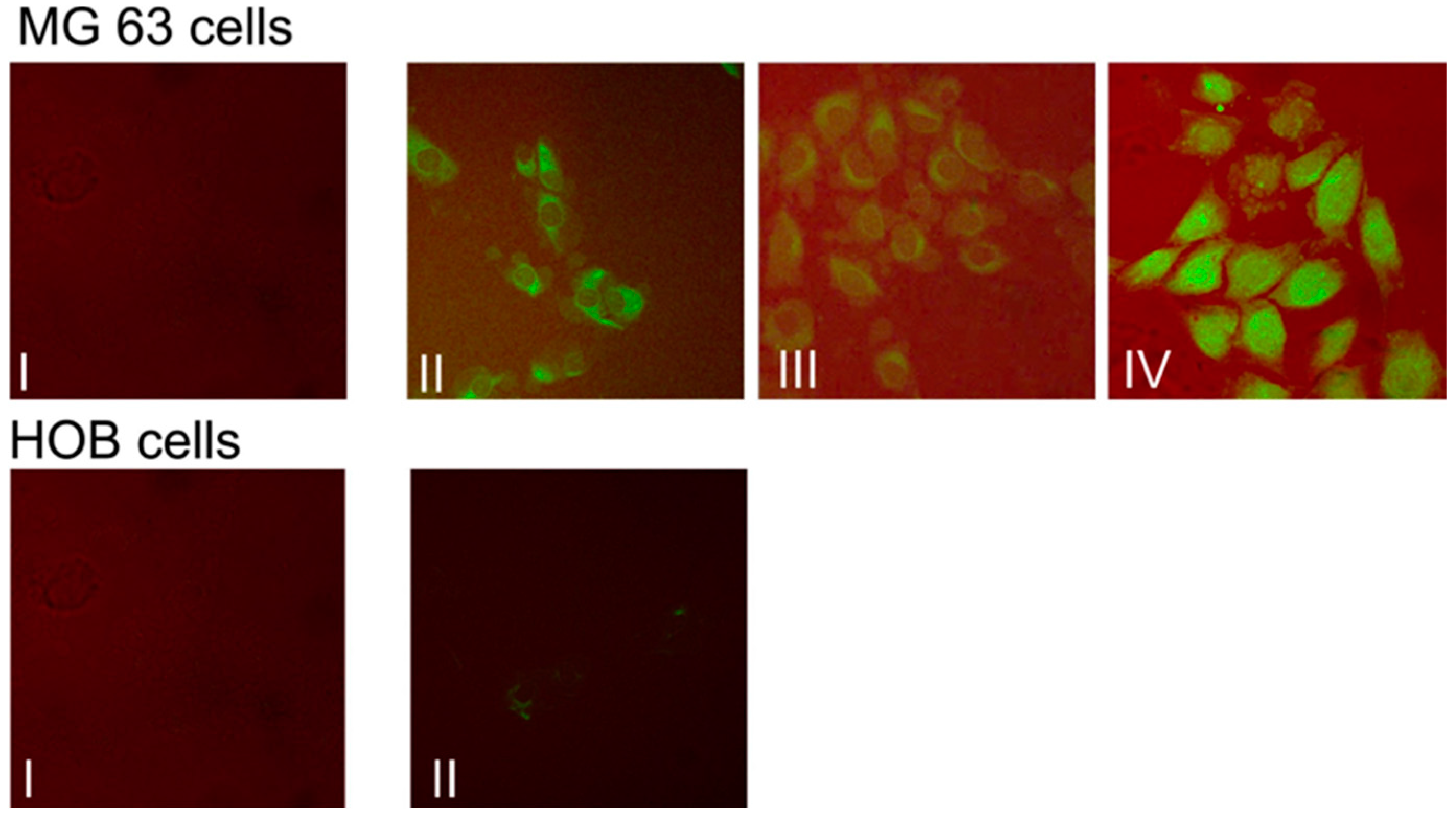
3.2. Micellar Assembly Mediated Curcumin Delivery to Osteosarcoma Cells
4. Conclusions
Author Contributions
Conflicts of Interest
References
- Arndt, C.A.; Crist, W.M. Common musculoskeletal tumors of childhood and adolescence. N. Engl. J. Med. 1999, 341, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Hayden, J.B.; Hoang, B.H. Osteosarcoma: Basic science and clinical implications. Orthop. Clin. N. Am. 2006, 37, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kansara, M.; Thomas, D.M. Molecular pathogenesis of osteosarcoma. DNA Cell Biol. 2004, 26, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Marina, N.; Gebhardt, M.; Teot, L.; Gorlick, R. Biology and therapeutic advances for pediatric osteosarcoma. Oncologist 2004, 9, 422–441. [Google Scholar] [CrossRef] [PubMed]
- Bielack, S.S.; Kempf-Bielack, B.; Delling, G.; Exner, G.U.; Flege, S.; Helmke, K.; Kotz, R.; Salzer-Kuntschik, M.; Werner, M.; Winkelmann, W.; et al. Prognostic factors in high-grade osteosarcoma of the extremities or trunk: An analysis of 1702 patients treated on neoadjuvant cooperative osteosarcoma study group protocols. J. Clin. Oncol. 2002, 20, 776–790. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, R.; Cheung, N.K.; Bowman, L.; Castle, V.; Hoffer, F.; Kapoor, N.; Kletzel, M.; Lindsley, K.; Shamberger, R.; Tubergen, D. NCCN pediatric neuroblastoma practice guidelines. The National Comprehensive Cancer Network. Oncology (Williston Park) 1996, 10, 1813–1822. [Google Scholar] [PubMed]
- Ward, W.G.; Mikaelian, K.; Dorey, F.; Mirra, J.M.; Sassoon, A.; Holmes, E.C.; Eilber, F.R.; Eckardt, J.J. Pulmonary metastases of stage IIB extremity osteosarcoma and subsequent pulmonary metastases. J. Clin. Oncol. 1994, 12, 1849–1858. [Google Scholar] [PubMed]
- Arndt, C.A.; Rose, P.S.; Folpe, A.L.; Laack, N.N. Common musculoskeletal tumors of childhood and adolescence. Mayo Clin. Proc. 2012, 87, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, Y.; Tanaka, K. The activity of the Bone and Soft Tissue Tumor Study Group of the Japan Clinical Oncology Group. Jpn. J. Clin. Oncol. 2012, 42, 467–470. [Google Scholar] [CrossRef] [PubMed]
- Baum, E.S.; Gaynon, P.; Greenberg, L.; Krivit, W.; Hammond, D. Phase II trail cisplatin in refractory childhood cancer: Children’s cancer study group report. Cancer Treat. Rep. 1981, 65, 815–822. [Google Scholar] [PubMed]
- Smith, M.A.; Ungerleider, R.S.; Horowitz, M.E.; Simon, R. Influence of doxorubicin dose intensity on response and outcome for patients with osteogenic sarcoma and Ewing's sarcoma. J. Nat. Cancer Inst. 1991, 83, 1460–1470. [Google Scholar] [CrossRef] [PubMed]
- Meistrich, M.L.; Chawla, S.P.; da Cunha, M.F.; Johnson, S.L.; Plager, C.; Papadopoulos, N.E.; Lipshultz, L.I.; Benjamin, R.S. Recovery of sperm production after chemotherapy for osteosarcoma. Cancer 1989, 63, 2115–2123. [Google Scholar] [CrossRef]
- Goorin, A.M.; Shuster, J.J.; Baker, A.; Horowitz, M.E.; Meyer, W.H.; Link, M.P. Changing pattern of pulmonary metastases with adjuvant chemotherapy in patients with osteosarcoma: Results from the multiinstitutional osteosarcoma study. J. Clin. Oncol. 1991, 9, 600–605. [Google Scholar] [PubMed]
- Kunchandy, E.; Rao, M.N.A. Oxygen radical scavenging activity of curcumin. Int. J. Pharm. 1990, 58, 237–240. [Google Scholar] [CrossRef]
- Chan, M.M.; Huang, H.I.; Fenton, M.R.; Fong, D. In vivo inhibition of nitric oxide synthase gene expression by curcumin, a cancer preventive natural product with anti-inflammatory properties. Biochem. Pharmacol. 1998, 55, 1955–1962. [Google Scholar] [CrossRef]
- Gota, V.S.; Maru, G.B.; Soni, T.G.; Gandhi, T.R.; Kochar, N.; Agarwal, M.G. Safety and pharmacokinetics of a solid lipid curcumin particle formulation in osteosarcoma patients and healthy volunteers. J. Agric. Food Chem. 2010, 58, 2095–2099. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Xu, H.G.; Shen, J.N.; Chen, X.W.; Wang, H.; Zhou, J.G. Apoptotic effects of curcumin on human osteosarcoma U2OS cells. Orthop. Surg. 2009, 1, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, J.; Ma, D.; Zhang, L.; Si, M.; Yin, H.; Li, J. Curcumin inhibits proliferation and invasion of osteosarcoma cells through inactivation of Notch-1 signaling. FEBS J. 2012, 279, 2247–2259. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Tremblaym, P.; Mahngar, K.; Collins, J.; Hudlicky, T.; Pandey, S. Selective cytotoxicity against human osteosarcoma cells by a novel synthetic C-1 analogue of 7-deoxypancratistatin is potentiated by curcumin. PLoS ONE 2001, 6, e28780. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Eifes, S.; Dicato, M.; Aggarwal, B.B.; Diederich, M. Modulation of anti-apoptotic and survival pathways by curcumin as a strategy to induce apoptosis in cancer cells. Biochem. Pharmacol. 2008, 76, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Strimpakos, A.S.; Sharma, R.A. Curcumin: Preventive and therapeutic properties in laboratory studies and clinical trials. Antioxid. Redox Signal. 2008, 10, 511–545. [Google Scholar] [CrossRef] [PubMed]
- Hatcher, H.; Planalp, R.; Cho, J.; Torti, F.M.; Torti, S.V. Curcumin: From ancient medicine to current clinical trials. Cell. Mol. Life Sci. 2008, 65, 1631–1652. [Google Scholar] [CrossRef] [PubMed]
- Sikora, E.; Bielak-Zmijewska, A.; Magalska, A.; Piwocka, K.; Mosieniak, G.; Kalinowska, M.; Widlak, P.; Cymerman, I.A.; Bujnicki, J.M. Curcumin induces caspase-3-dependent apoptotic pathway but inhibits DNA fragmentation factor 40/caspase-activated DNase endonuclease in human Jurkat cells. Mol. Cancer Ther. 2006, 5, 927–934. [Google Scholar] [CrossRef] [PubMed]
- Magalska, A.; Brzezinska, A.; Bielak-Zmijewska, A.; Piwocka, K.; Mosieniak, G.; Sikora, E. Curcumin induces cell death without oligonucleosomal DNA fragmentation in quiescent and proliferating human CD8+ cells. Acta Biochim. Pol. 2006, 53, 531–538. [Google Scholar] [PubMed]
- Mosieniak, G.; Sliwinska, M.; Piwocka, K.; Sikora, E. Curcumin abolishes apoptosis resistance of calcitriol-differentiated HL-60 cells. FEBS Lett. 2006, 580, 4653–4660. [Google Scholar] [CrossRef] [PubMed]
- Wolanin, K.; Magalska, A.; Mosieniak, G.; Klinger, R.; McKenna, S.; Vejda, S.; Sikora, E.; Piwocka, K. Curcumin affects components of the chromosomal passenger complex and induces mitotic catastrophe in apoptosis-resistant Bcr-Abl-expressing cells. Mol. Cancer Res. 2006, 4, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Bielak-Zmijewska, A.; Sikora-Polaczek, M.; Nieznanski, K.; Mosieniak, G.; Kolano, A.; Maleszewski, M.; Styrna, J.; Sikora, E. Curcumin disrupts meiotic and mitotic divisions via spindle impairment and inhibition of CDK1 activity. Cell Prolif. 2010, 43, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, J.; Prasad, S.; Aggarwal, B.B. Curcumin and cancer cells: How many ways can curry kill tumor cells selectively? AAPS J. 2009, 11, 495–510. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Sundaram, C.; Jhuran, S.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin and cancer: An “old-age” disease with an “age-old” solution. Cancer Lett. 2008, 67, 133–164. [Google Scholar] [CrossRef] [PubMed]
- Lan, L.; Fadi, S.B.; Razelle, K. Liposome-encapsulated curcumin. Cancer 2005, 104, 1322–1331. [Google Scholar]
- Kunwar, A.; Barik, A.; Pandey, R.; Priyadarsini, K.I. Transport of liposomal and albumin loaded curcumin to living cells: An absorption and fluorescence spectroscopic study. Biochim. Biophys. Acta 2006, 1760, 1513–1520. [Google Scholar] [CrossRef] [PubMed]
- Patel, B.B.; Majumdar, A.P. Synergistic role of curcumin with current therapeutics in colorectal cancer: Minireview. Nutr. Cancer 2009, 61, 842–846. [Google Scholar] [CrossRef] [PubMed]
- Chakravarti, N.; Kadara, H.; Yoon, D.J.; Shay, J.W.; Myers, J.N.; Lotan, D.; Sonenberg, N.; Lotan, R. Differential inhibition of protein translation machinery by curcumin in normal, immortalized, and malignant oral epithelial cells. Cancer Prev. Res. 2010, 3, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, Y.; Banerjee, S.; Li, Y.; Sarkar, F.H. Notch-1 down-regulation by curcumin is associated with the inhibition of cell growth and the induction of apoptosis in pancreatic cancer cells. Cancer 2006, 106, 2503–2513. [Google Scholar] [CrossRef] [PubMed]
- Walters, D.K.; Muff, R.; Langsam, B.; Born, W.; Fuchs, B. Cytotoxic effects of curcumin on osteosarcoma cell lines. Investig. New Drugs 2008, 26, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Hanif, R.; Qiao, L.; Shiff, S.J.; Rigas, B. Curcumin, a natural plant phenolic food additive, inhibits cell proliferation and induces cell cycle changes in colon adenocarcinoma cell lines by a prostaglandin-independent pathway. J. Lab. Clin. Med. 1997, 130, 576–584. [Google Scholar] [CrossRef]
- Dorai, T.; Cao, Y.C.; Dorai, B.; Buttyan, R.; Katz, A.E. Therapeutic potential of curcumin in human prostate cancer. III. Curcumin inhibits proliferation, induces apoptosis, and inhibits angiogenesis of LNCaP prostate cancer cells in vivo. Prostate 2001, 47, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.T.; Lou, Y.R.; Xie, J.G.; Ma, W.; Lu, Y.P.; Yen, P.; Zhu, B.T.; Newmark, H.; Ho, C.T. Effect of dietary curcumin and dibenzoylmethane on formation of 7,12-dimethylbenz[a]anthracene-induced mammary tumors and lymphomas/leukemias in Sencar mice. Carcinogenesis 1998, 19, 1697–1700. [Google Scholar] [CrossRef] [PubMed]
- Fossey, S.L.; Bear, M.D.; Lin, J.; Li, C.; Schwartz, E.B.; Li, P.K.; Fuchs, J.R.; Fenger, J.; Kisseberth, W.C.; London, C.A. The novel curcumin analog FLLL32 decreases STAT3 DNA binding activity and expression, and induces apoptosis in osteosarcoma cell lines. BMC Cancer 2011, 11, 112. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Shen, G.; Khor, T.O.; Kim, J.H.; Kong, A.N. Curcumin inhibits Akt/mammalian target of rapamycin signaling through protein phosphatase-dependent mechanism. Mol. Cancer Ther. 2008, 7, 2609–2620. [Google Scholar] [CrossRef] [PubMed]
- Kuo, M.L.; Huang, T.S.; Lin, J.K. Curcumin, an antioxidant and anti-tumor promoter, induces apoptosis in human leukemia cells. Biochim. Biophys. Acta 1996, 1317, 95–100. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, Z.S.; Zhang, Y.L.; Zhou, D.H. Curcumin inhibits cell proliferation by interfering with the cell cycle and inducing apoptosis in colon carcinoma cells. Anticancer Res. 1999, 19, 3675–3680. [Google Scholar] [PubMed]
- Mehta, K.; Pantazis, P.; McQueen, T.; Aggarwal, D.H. Antiproliferative effects of curcumin (diferuloylmethane) against human breast tumor cell lines. Anticancer Drugs 1997, 8, 470–481. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.A.; Gescher, A.J.; Steward, W. Curcumin: The story so far. Eur. J. Cancer 2005, 41, 1955–1968. [Google Scholar] [CrossRef] [PubMed]
- Er, E.; Oliver, L.; Cartron, P.-F.; Juin, P.; Manon, S.; Vallette, F.M. Mitochondria as the target of the pro-apoptotic protein Bax. Biochim. Biophys. Acta 2006, 1757, 1301–1311. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.M.; Ranganathan, V.; Farnsworth, M.L.; Kavallaris, M.; Lock, R.B. Bcl-2 inhibits Bax translocation from cytosol to mitochondria during drug-induced apoptosis of human tumor cells. Cell Death Differ. 2000, 7, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Narita, M.; Shimizu, S.; Ito, T.; Chittenden, T.; Lutz, R.J.; Matsuda, H.; Tsujimoto, Y. Bax interacts with the permeability transition pore to induce permeability transition and cytochrome c release in isolated mitochondria. Proc. Natl. Acad. Sci. USA 1998, 95, 14681–14686. [Google Scholar] [CrossRef] [PubMed]
- Shankar, S.; Srivastava, R.K. Involvement of Bcl-2 family members, phosphatidylinositol 3′-kinase/AKT and mitochondrial p53 in curcumin (diferuloylmethane)-induced apoptosis in prostate cancer. Int. J. Oncol. 2007, 30, 905–918. [Google Scholar] [PubMed]
- Mukherjee, N.C.S.; Ghosh, U.; Bhattacharyya, N.P.; Bhattacharya, R.K.; Dey, S.; Roy, M. Curcumin-induced apoptosis in human leukemia cell HL-60 is associated with inhibition of telomerase activity. Mol. Cell Biochem. 2007, 297, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Anto, R.J.; Mukhopadhyay, A.; Denning, K.; Aggarwal, B.B. Curcumin (diferuloylmethane) induces apoptosis through activation of caspase-8, BID cleavage and cytochrome c release: Its suppression by ectopic expression of Bcl-2 and Bcl-xl. Carcinogenesis 2002, 23, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Shankar, S.; Srivastava, R.K. Bax and Bak genes are essential for maximum apoptotic response by curcumin, a polyphenolic compound and cancer chemopreventive agent derived from turmeric, Curcuma longa. Carcinogenesis 2007, 28, 1277–1286. [Google Scholar] [CrossRef] [PubMed]
- Li, J.L.; Harris, A.L. Notch signaling from tumor cells: A new mechanism of angiogenesis. Cancer Cell 2005, 8, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.S.; Lee, M.K.; Kim, J.H. Curcumin induces cell cycle arrest and apoptosis in human osteosarcoma (HOS) cells. Anticancer Res. 2009, 29, 5039–5044. [Google Scholar] [PubMed]
- Leow, P.C.; Tian, Q.; Ong, Z.Y.; Yang, Z.; Ee, P.L. Antitumor activity of natural compounds, curcumin and PKF118–310, as Wnt/beta-catenin antagonists against human osteosarcoma cells. Investig. New Drugs 2010, 28, 766–782. [Google Scholar] [CrossRef] [PubMed]
- John, A.; Tuszynski, G. The role of matrix metalloproteinases in tumor angiogenesis and tumor metastasis. Pathol. Oncol. Res. 2001, 7, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Stetler-Stevenson, W.G. The role of matrix metalloproteinases in tumor invasion, metastasis, and angiogenesis. Surg. Oncol. Clin. N. Am. 2001, 10, 383–392. [Google Scholar] [PubMed]
- Wang, Z.; Banerjee, S.; Li, Y.; Rahman, K.M.; Zhang, Y.; Sarkar, F.H. Down-regulation of notch-1 inhibits invasion by inactivation of nuclear factor-kappaB, vascular endothelial growth factor, and matrix metalloproteinase-9 in pancreatic cancer cells. Cancer Res. 2006, 66, 2778–2784. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fu, L.; Gu, F.; Ma, Y. Notch1 is involved in migration and invasion of human breast cancer cells. Oncol. Rep. 2011, 26, 1295–1303. [Google Scholar] [PubMed]
- Yu, B.; Wei, J.; Qian, X.; Lei, D.; Ma, Q.; Liu, Y. Notch1 signaling pathway participates in cancer invasion by regulating MMPs in lingual squamous cell carcinoma. Oncol. Rep. 2012, 27, 547–552. [Google Scholar] [PubMed]
- Jaruga, E.; Bielak-Zmijewska, A.; Sikora, E.; Skierski, J.; Radziszewska, E.; Piwocka, K.; Bartosz, G. Glutathione-independent mechanism of apoptosis inhibition by curcumin in rat thymocytes. Biochem. Pharmacol. 1998, 56, 961–965. [Google Scholar] [CrossRef]
- Syng-Ai, C.; Kumari, A.L.; Khar, A. Effect of curcumin on normal and tumor cells: Role of glutathione and bcl-2. Mol. Cancer Ther. 2004, 3, 1101–1108. [Google Scholar] [PubMed]
- Uddin, S.; Hussain, A.R.; Manogaran, P.S.; Al-Hussein, K.; Platanias, L.C.; Gutierrez, M.I.; Bhatia, K.G. Curcumin suppresses growth and induces apoptosis in primary effusion lymphoma. Oncogene 2005, 24, 7022–7030. [Google Scholar] [CrossRef] [PubMed]
- Karunagaran, D.; Rashmi, R.; Kumar, T.R. Induction of apoptosis by curcumin and its implications for cancer therapy. Curr. Cancer Drug Targets 2005, 5, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Watson, J.L.; Hill, R.; Lee, P.W.; Giacomantonio, C.A.; Hoskin, D.W. Curcumin induces apoptosis in HCT-116 human colon cancer cells in a p21-independent manner. Exp. Mol. Pathol. 2008, 84, 230–233. [Google Scholar] [CrossRef] [PubMed]
- Ghoneim, A.I. Effects of curcumin on ethanol-induced hepatocyte necrosis and apoptosis: Implication of lipid peroxidation and cytochrome c. Naunyn. Schmiedebergs Arch. Pharmacol. 2009, 379, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Cucuzza, L.S.; Motta, M.; Miretti, S.; Accornero, P.; Baratta, M. Curcuminoid-phospholipid complex induces apoptosis in mammary epithelial cells by STAT-3 signaling. Exp. Mol. Med. 2008, 40, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Kunwar, A.; Barik, A.; Mishra, B.; Rathinasamy, K.; Pandey, R.; Priyadarsini, K.I. Quantitative cellular uptake, localization and cytotoxicity of curcumin in normal and tumor cells. Biochim. Biophys. Acta 2008, 1780, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Shishodia, S.; Amin, H.M.; Lai, R.; Aggarwal, B.B. Curcumin (diferuloylmethane) inhibits constitutive NF-kappaB activation, induces G1/S arrest, suppresses proliferation, and induces apoptosis in mantle cell lymphoma. Biochem. Pharmacol. 2005, 70, 700–713. [Google Scholar] [CrossRef] [PubMed]
- Scharstuhl, A.; Mutsaers, H.A.; Pennings, S.W.; Szarek, W.A.; Russel, F.G.; Wagener, F.A. Curcumin-Induced Fibroblast Apoptosis and in vitro Wound Contraction Are Regulated by Antioxidants and Heme Oxygenase: Implications for Scar Formation. J. Cell. Mol. Med. 2009, 13, 712–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, W.H.; Wu, H.Y.; Chang, W.H. Dosage effects of curcumin on cell death types in a human osteoblast cell line. Food Chem. Toxicol. 2006, 44, 1362–1371. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.; Sun, L.; Webster, T.J. Short communication: Selective cytotoxicity of curcumin on osteosarcoma cells compared to healthy osteoblasts. Int. J. Nanomedicine 2014, 9, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, M.; Satoh, A.; Sakurai, Y.; Okano, T.; Matsumura, Y.; Kakizoe, T. Incorporation of water-insoluble anticancer drug into polymeric micelles and control of their particle size. J. Control. Release 1998, 55, 219–229. [Google Scholar] [CrossRef]
- Batrakova, E.V.; Dorodnych, T.Y.; Klinskii, E.Y.; Kliushnenkova, E.N.; Shemchukova, O.V.; Goncharova, O.N. Anthracycline antibiotics non-covalently incorporated into the block copolymer micelles: In vivo evaluation of anticancer activity. Br. J. Cancer 1996, 74, 1545–1552. [Google Scholar] [CrossRef] [PubMed]
- Kabanov, A.V.; Nazarova, I.R.; Astafieva, I.R.; Batrakova, E.V.; Alakhov, V.Y.; Yaroslavov, A.A. Micelle formation and solubilization of fluorescence probes in poly(oxyethylene-b-oxypropylene-b-oxyethylene) solutions. Macromolecules 1995, 28, 2303–2314. [Google Scholar] [CrossRef]
- Kabanov, A.V.; Vinogradov, S.V.; Suzdaltseva, U.G.; Alakhov, V.Y. Water-soluble block polycations as carriers for oligonucleotide delivery. Bioconj. Chem. 1995, 6, 639–643. [Google Scholar] [CrossRef]
- Alakhov, V.Y.; Kabanov, A.V. Block copolymeric biotransport carriers as versatile vehicles for drug delivery. Expert Opin. Investig. Drugs 1998, 7, 1453–1473. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Yokoyama, M.; Kataoka, K.; Okano, T.; Sakurai, Y.; Kawaguchi, T. Reduction of the side effects of an antitumor agent, KRN5500, by incorporation of the drug into polymeric micelles. Jpn. J. Cancer Res. 1999, 90, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Torchilin, V.P.; Trubetskoy, V.S. Biodistribution of surface-modified liposomes and particles. In Microparticulate Systems for the Delivery of Proteins and Vaccines; Cohen, S., Bernstein, H., Eds.; Marcel Dekker: New York, NY, USA, 1996; Volume 8, pp. 243–277. [Google Scholar]
- Trubetskoy, V.S.; Torchilin, V.P. Use of polyoxyethylene-lipid conjugates as long-circulating carriers for delivery of therapeutic and diagnostic agents. Adv. Drug Deliv. Rev. 1995, 16, 311–320. [Google Scholar] [CrossRef]
- Kwon, G.S.; Kataoka, K. Block-Copolymer Micelles as Long-Circulating Drug Vehicles. Adv. Drug Deliv. Rev. 1995, 16, 295–309. [Google Scholar] [CrossRef]
- Kwon, G.S.; Okano, T. Soluble self-assembled block copolymers for drug delivery. Pharm. Res. 1999, 16, 597–560. [Google Scholar] [CrossRef] [PubMed]
- Martin, A. Physical Pharmacy, 4th ed.; Williams and Wilkins: Baltimore, MA, USA, 1993; pp. 396–398. [Google Scholar]
- Gao, Z.; Eisenberg, A. A model of micellization for block copolymers in solutions. Macromolecules 1993, 26, 7353–7360. [Google Scholar] [CrossRef]
- Kakizawa, Y.; Kataoka, K. Block copolymer micelles for delivery of gene and related compounds. Adv. Drug Deliv. Rev. 2002, 54, 203–222. [Google Scholar] [CrossRef]
- Torchilin, V.P. Structure and design of polymeric surfactant-based drug delivery systems. J. Control. Release 2001, 73, 137–172. [Google Scholar] [CrossRef]
- Lin, S.Y.; Kawashima, Y. The influence of three poly(oxyethylene) poly(oxypropylene) surface-active block copolymers on the solubility behavior of indomethacin. Pharm. Acta Helv. 1985, 60, 339–344. [Google Scholar] [PubMed]
- Lin, S.Y.; Kawashima, Y. Pluronic surfactants affecting diazepam solubility, compatibility and adsorption from i.v. admixture solutions. J. Pharm. Sci. Technol. 1985, 41, 83–87. [Google Scholar]
- Yokoyama, M.; Okano, T.; Kataoka, K. Improved synthesis of adriamycin-conjugated poly(ethylene oxide)-poly(aspartic acid) block copolymer and formation of unimodal micellar structure with controlled amount of physically entrapped adriamycin. J. Control. Release 1994, 32, 269–277. [Google Scholar] [CrossRef]
- Yokoyama, M.; Fukushima, S.; Uehara, R.; Okamoto, K.; Kataoka, K.; Sakurai, Y. Characterization of physical entrapment and chemical conjugation of adriamycin in polymeric micelles and their design for in vivo delivery to a solid tumor. J. Control. Release 1998, 50, 79–92. [Google Scholar] [CrossRef]
- Kabanov, A.V.; Chekhonin, V.P.; Alakhov, V.Y.; Batrakova, E.V.; Lebedev, A.S.; Melik-Nubarov, N.S. The neuroleptic activity of haloperidol increases after its solubilization in surfactant micelles. FEBS Lett. 1989, 258, 343–345. [Google Scholar] [CrossRef]
- Hagan, S.A.; Coombes, A.G.A.; Garnett, M.C.; Dunn, S.E.; Davies, M.C.; Illum, L. Polylactide-poly(ethelene glycol) copolymers as drug delivery systems, 1. Characterization of water dispersible micelle-forming systems. Langmuir 1996, 12, 2153–2161. [Google Scholar] [CrossRef]
- Inoue, T.; Chen, G.; Nakamae, K.; Hoffman, A.S. An AB block copolymers of oligo(methyl methacrylate) and poly(acrylic acid) for micellar delivery of hydrophobic drugs. J. Control. Release 1998, 51, 221–229. [Google Scholar] [CrossRef]
- Zeng, J.G.; Shi, K.Y.; Zhang, Y.Y.; Sun, X.H.; Zhang, B.L. Construction and micellization of a noncovalent double hydrophilic block copolymer. Chem. Commun. 2008, 32, 3753–3755. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.W.; Zhuang, X.L.; He, C.L.; Chen, X.S.; Jing, X.B. Synthesis of Novel Thermo- and pH-Responsive Poly(l-lysine)-Based Copolymer and its Micellization in Water. Macromol. Rapid Commun. 2008, 29, 1810–1816. [Google Scholar] [CrossRef]
- Gillies, E.R.; Jonsson, T.B.; Frechet, J.M.J. Stimuli-responsive supramolecular assemblies of linear-dendritic copolymers. J. Am. Chem. Soc. 2004, 126, 11936–11943. [Google Scholar] [CrossRef] [PubMed]
- Washington, C. Drug release from microdisperse systems: A critical review. Int. J. Pharm. 2005, 58, 1–12. [Google Scholar] [CrossRef]
- Hubbell, J.A. Materials science: Enhancing drug function. Science 2003, 300, 596–596. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Lavender, C.A.; Woodcock, J.W.; Zhao, B. Multiple micellization and dissociation transitions of thermo- and light-sensitive poly(ethylene oxide)-b-poly(ethoxytri(ethylene glycol) acrylate-co-o-nitrobenzyl acrylate) in water. Macromolecules 2008, 41, 2632–2643. [Google Scholar] [CrossRef]
- Kwon, G.S.; Naito, M.; Yokoyama, M.; Okano, T.; Sakurai, Y.; Kataoka, K. Block copolymer micelles for drug delivery: Loading and release of doxorubicin. J. Control. Release 1997, 48, 195–201. [Google Scholar] [CrossRef]
- Harada, A.; Kataoka, K. Novel polyion complex micelles entrapping enzyme molecules in the core. Preparation of narrowly-distributed micelles from lysozyme and poly(ethylene glycol)-poly(aspartic acid) block copolymer in aqueous medium. Macromolecules 1998, 31, 288–294. [Google Scholar] [CrossRef]
- Katayose, S.; Kataoka, K. Remarkable increase in nuclease resistance of plasmid DNA through supramolecular assembly with poly(ethylene glycol)-poly(l-lysine) block copolymer. J. Pharm. Sci. 1998, 87, 160–163. [Google Scholar] [CrossRef] [PubMed]
- Kabanov, A.V.; Kabanov, V.A. Interpolyelectrolyte and block ionomer complexes for gene delivery: Physicochemical aspects. Adv. Drug Deliv. Rev. 1990, 30, 49–60. [Google Scholar] [CrossRef]
- Batrakova, E.V.; Li, S.; Miller, D.W.; Kabanov, A.V. Pluronic P85 increases permeability of a broad spectrum of drugs in polarized BBMEC and Caco-2 cell monolayers. Pharm. Res. 1999, 16, 1366–1372. [Google Scholar] [CrossRef] [PubMed]
- Kabanov, A.V.; Slepnev, V.I.; Kuznetsova, L.E.; Batrakova, E.V.; Alakhov, V.; Melik-Nubarov, N.S.; Sveshnikov, P.G.; Kabanov, V.A. Pluronic micelles as a tool for lowmolecular compound vector delivery into a cell: Effect of Staphylococcus aureus enterotoxin B on cell loading with micelle incorporated fluorescent dye. Biochem. Int. 1992, 26, 1035–1042. [Google Scholar] [PubMed]
- Kabanov, A.V.; Alakhov, V.Y. Pluronic block copolymers in drug delivery: From micellar nanocontainers to biological response modifiers. Crit. Rev. Ther. Drug Carrier Syst. 2002, 19, 1–72. [Google Scholar] [CrossRef] [PubMed]
- Batrakova, E.V.; Li, S.; Vinogradov, S.V.; Alakhov, V.Y.; Miller, D.W.; Kabanov, A.V. Mechanism of pluronic effect on P-glycoprotein efflux system in blood-brain barrier: Contributions of energy depletion and membrane fluidization. J. Pharmacol. Exp. Ther. 2001, 299, 483–493. [Google Scholar] [PubMed]
- Kabanov, A.V.; Batrakova, E.V.; Miller, D.W. Pluronic block copolymers as modulators of drug efflux transporter activity in the blood-brain barrier. Adv. Drug Delivery Rev. 2003, 55, 151–164. [Google Scholar] [CrossRef]
- Sahay, G.; Batrakova, E.V.; Kabanov, A.V. Different Internalization Pathways of Polymeric Micelles and Unimers and Their Effects on Vesicular Transport. Bioconj. Chem. 2008, 19, 2023–2029. [Google Scholar] [CrossRef] [PubMed]
- Kabanov, A.V. Polymer genomics: An insight into pharmacology and toxicology of nanomedicines. Adv. Drug Deliv. Rev. 2006, 58, 1597–1621. [Google Scholar] [CrossRef] [PubMed]
- Torchilin, V. Tumor delivery of macromolecular drugs based on the EPR effect. Adv. Drug Deliv. Rev. 2011, 63, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Cheng, R.; Deng, C.; Zhong, Z. Intercellular drug release nanosystems. Mater. Today 2012, 15, 436–442. [Google Scholar] [CrossRef]
- Osada, K.; Christie, R.J.; Kataoka, K. Polymeric micelles from poly(ethylene glycol)-poly(amino acid) block copolymer for drug and gene delivery. J. R. Soc. Interface R. Soc. 2009, 6, S325–S339. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Shen, Y.; Hou, J.; Lei, L.; Guo, S.; Qian, C. Polymeric micelles for parenteral delivery of curcumin: preparation, characterization and in vitro evaluation. Colloid Surf. A 2011, 390, 25–32. [Google Scholar] [CrossRef]
- Song, Z.; Feng, R.; Sun, M.; Guo, C.; Gao, Y.; Li, L.; Zhai, G. Curcumin-loaded PLGA-PEG-PLGA triblock copolymeric micelles: Preparation, pharmacokinetics and distribution in vivo. J. Colloid Interface Sci. 2011, 354, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Du, J.; Duan, Y.; Zang, Y.; Zhang, H.; Yang, C.; Cao, F.; Zhai, G. Curcumin loaded mixed micelles composed of pluronic P123 and F68: Preparation, optimization and in vitro characterization. Colloid Surf. B 2012, 97, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Samanta, S.; Roccatano, D. Interaction of curcumin with PEO-PPO-PEO triblock copolymers: A molecular dynamics study. J. Phys. Chem. B 2013, 117, 3250–3257. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Deng, S.; Wu, Q.; Xiang, M.; Wei, X.; Li, L.; Gao, X.; Wang, B.; Sun, L.; Chen, Y.; et al. Improving antiangiogenesis and anti-tumor activity of curcumin by biodegradable polymeric micelles. Biomaterials 2013, 34, 1413–1432. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Sun, L.; Wu, Q.; Guo, W.; Li, L.; Chen, Y.; Li, Y.; Gong, C.; Qian, Z.; Wei, Y. Curcumin loaded polymeric micelles inhibit breast tumor growth and spontaneous pulmonary metastasis. Int. J. Pharm. 2013, 443, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Haddadi, A.; Molavi, O.; Lavasanifar, A.; Lai, R.; Samuel, J. Micelles of poly(ethylene oxide)-b-poly(ε-caprolactone) as vehicles for the solubilization, stabilization, and controlled delivery of curcumin. J. Biomed. Mater. Res. A 2008, 86, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Soliman, G.M.; Sharma, A.; Cui, Y.; Sharma, R.; Kakkar, A.; Maysinger, D. Miktoarm Star Micelles Containing Curcumin Reduce Cell Viability of Sensitized Glioblastoma. J. Nanomed. Biother. Discov. 2014, 4, 124. [Google Scholar] [CrossRef]
- Yang, X.; Li, Z.; Wang, N.; Li, L.; Song, L.; He, T.; Sun, L.; Wang, Z.; Wu, Q.; Luo, N.; et al. Curcumin-Encapsulated Polymeric Micelles Suppress the Development of Colon Cancer in vitro and in vivo. Sci. Rep. 2015, 5, 10322. [Google Scholar] [CrossRef] [PubMed]
- Kohut, A.; Voronov, A. Hierarchical Micellar Structures from Amphiphilic Invertible Polyesters: 1H NMR Spectroscopic Study. Langmuir 2009, 25, 4356–4360. [Google Scholar] [CrossRef] [PubMed]
- Hevus, I.; Kohut, A.; Voronov, A. Interfacial micellar phase transfer using amphiphilic invertible polymers. Pol. Chem. 2011, 2, 2767–2770. [Google Scholar] [CrossRef]
- Hevus, I.; Modgil, A.; Daniels, J.; Kohut, A.; Sun, C.; Stafslien, S.; Voronov, A. Invertible Micellar Polymer Assemblies for Delivery of Poorly Water-Soluble Drugs. Biomacromolecules 2012, 13, 2537–2545. [Google Scholar] [CrossRef] [PubMed]
- Kudina, O.; Kohut, A.; Tarnavchyk, I.; Hevus, I.; Voronov, A. Solvent-Responsive Self-Assembly of Amphiphilic Invertible Polymers Determined with SANS. Langmuir 2014, 30, 3310–3318. [Google Scholar] [CrossRef] [PubMed]
- Kudina, O.; Shogren, K.L.; Gustafson, K.T.; Yaszemski, M.J.; Maran, A.; Voronov, A. Invertible micellar polymer nanoassemblies target bone tumor cells but not normal osteoblast cells. Future Sci. OA 2015, 1, 1–9. [Google Scholar] [CrossRef]
- Kohut, A.; Hevus, I.; Voronov, S.; Voronov, A. Invertible Polymers and their Applications. In Industrial Applications for Intelligent Polymers and Coatings; Hosseini, M., Makhlouf, A.S.H., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 399–416. [Google Scholar]
- Kohut, A.; Sieburg, L.; Vasylyev, S.; Kudina, O.; Hevus, I.; Stafslien, S.; Daniels, J.; Kislenko, V.; Voronov, A. Amphiphilic invertible polymers (AIPs): Micellization and self-assembly in aqueous solutions. In Amphiphiles: Molecular Assembly and Applications; Nagarajan, R., Ed.; Oxford University Press Inc.: Washington, DC, USA, 2011; pp. 205–224. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AIP | Loading, wt % | Size, nm (Blank) | Size, nm (Loaded) | ζ-Potential, mV (Blank) | ζ-Potential, mV (Loaded) |
|---|---|---|---|---|---|
| PEG600PTHF650 | 10.3 ± 0.4 | 12.0 ± 0.2 | 17.5 ± 2.6 | −24.2 ± 3.8 | −18.2 ± 0.6 |
| PEG300PTHF250 | 3.6 ± 0.3 | 12.4 ± 0.2 | 18.4 ±1.1 | −43.0 ± 2.3 | −42.6 ± 1.5 |
| D10 | 1.9 ± 0.3 | 6.3 ± 0.3 | 7.8 ± 0.6 | −10.3 ± 0.9 | −7.4 ± 0.3 |
| S10 | 0.14 ± 0.03 | 3.3 ± 0.5 | 3.6 ± 0.2 | −7.1 ± 0.3 | −6.7 ± 0.9 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maran, A.; Yaszemski, M.J.; Kohut, A.; Voronov, A. Curcumin and Osteosarcoma: Can Invertible Polymeric Micelles Help? Materials 2016, 9, 520. https://doi.org/10.3390/ma9070520
Maran A, Yaszemski MJ, Kohut A, Voronov A. Curcumin and Osteosarcoma: Can Invertible Polymeric Micelles Help? Materials. 2016; 9(7):520. https://doi.org/10.3390/ma9070520
Chicago/Turabian StyleMaran, Avudaiappan, Michael J. Yaszemski, Ananiy Kohut, and Andriy Voronov. 2016. "Curcumin and Osteosarcoma: Can Invertible Polymeric Micelles Help?" Materials 9, no. 7: 520. https://doi.org/10.3390/ma9070520