Release of the Non-Steroidal Anti-Inflammatory Drug Flufenamic Acid by Multiparticulate Delivery Systems Promotes Adipogenic Differentiation of Adipose-Derived Stem Cells

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
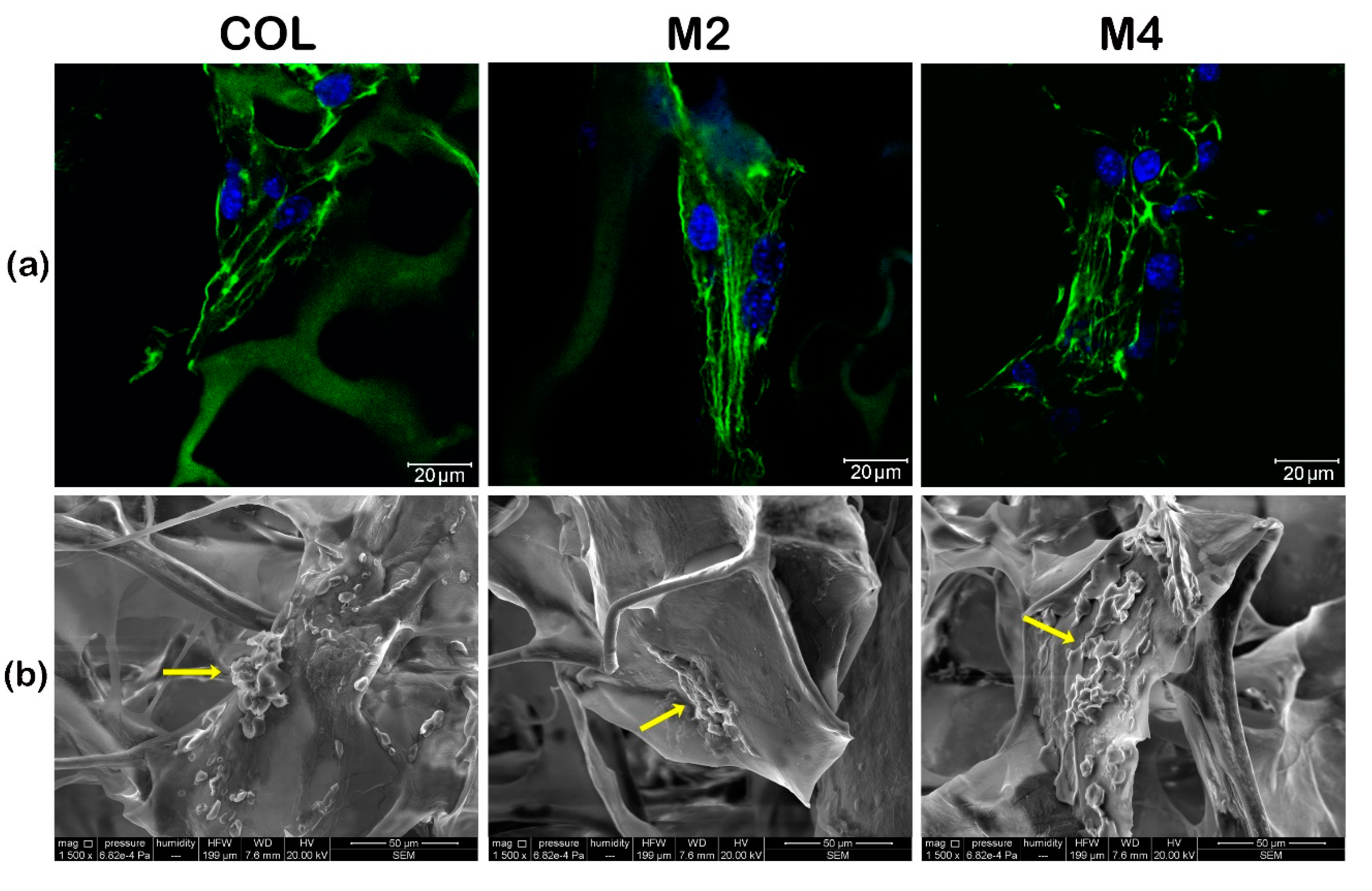
3.1. Assessment of the Tridimensional Cell-Scaffold Constructs Formation
3.2. Evaluation of Flufenamic Acid Effect on Adipogenic Differentiation of Human Adipose-Derived Stem Cells
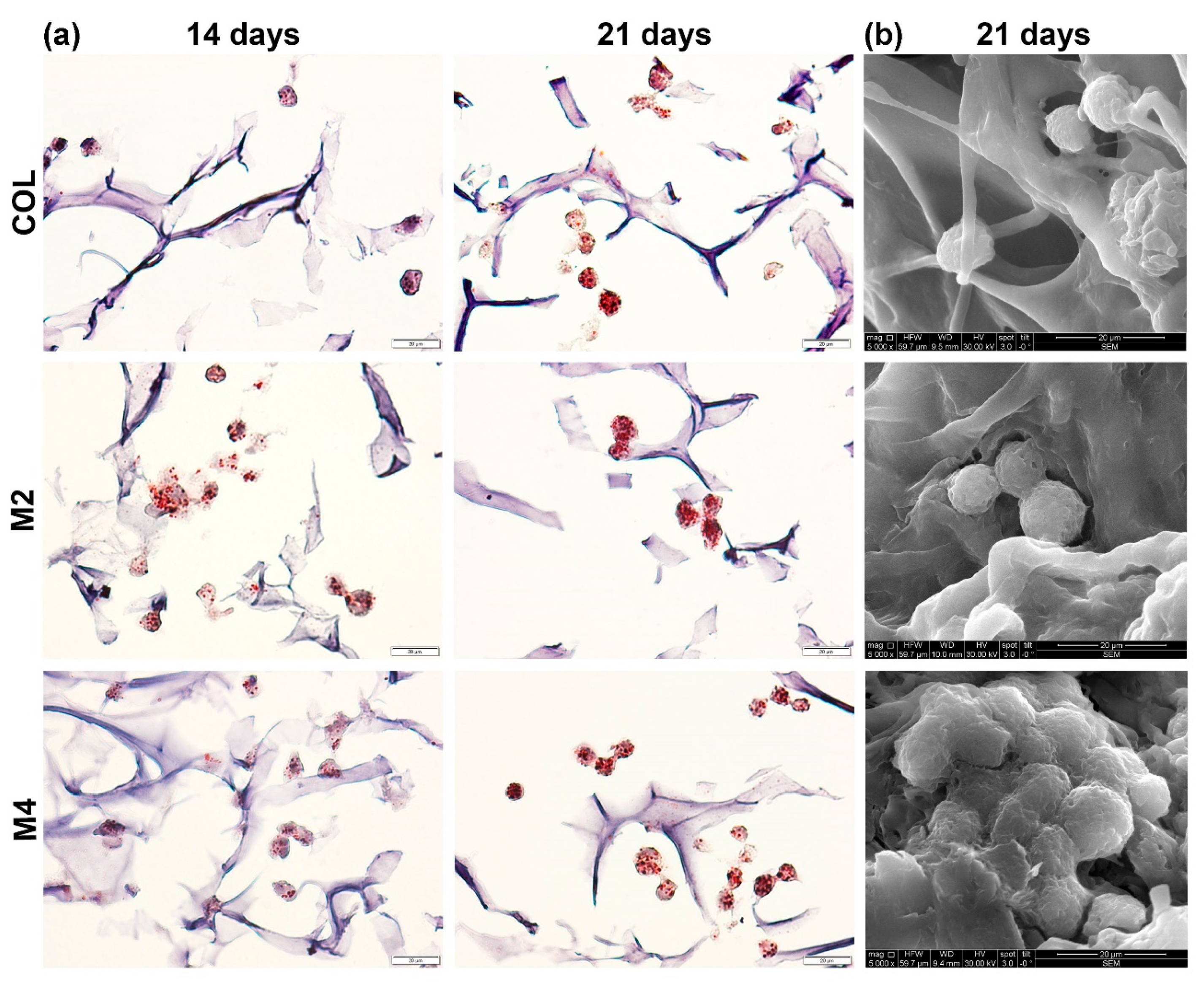
3.2.1. Evaluation of Intracellular Lipid Accumulation
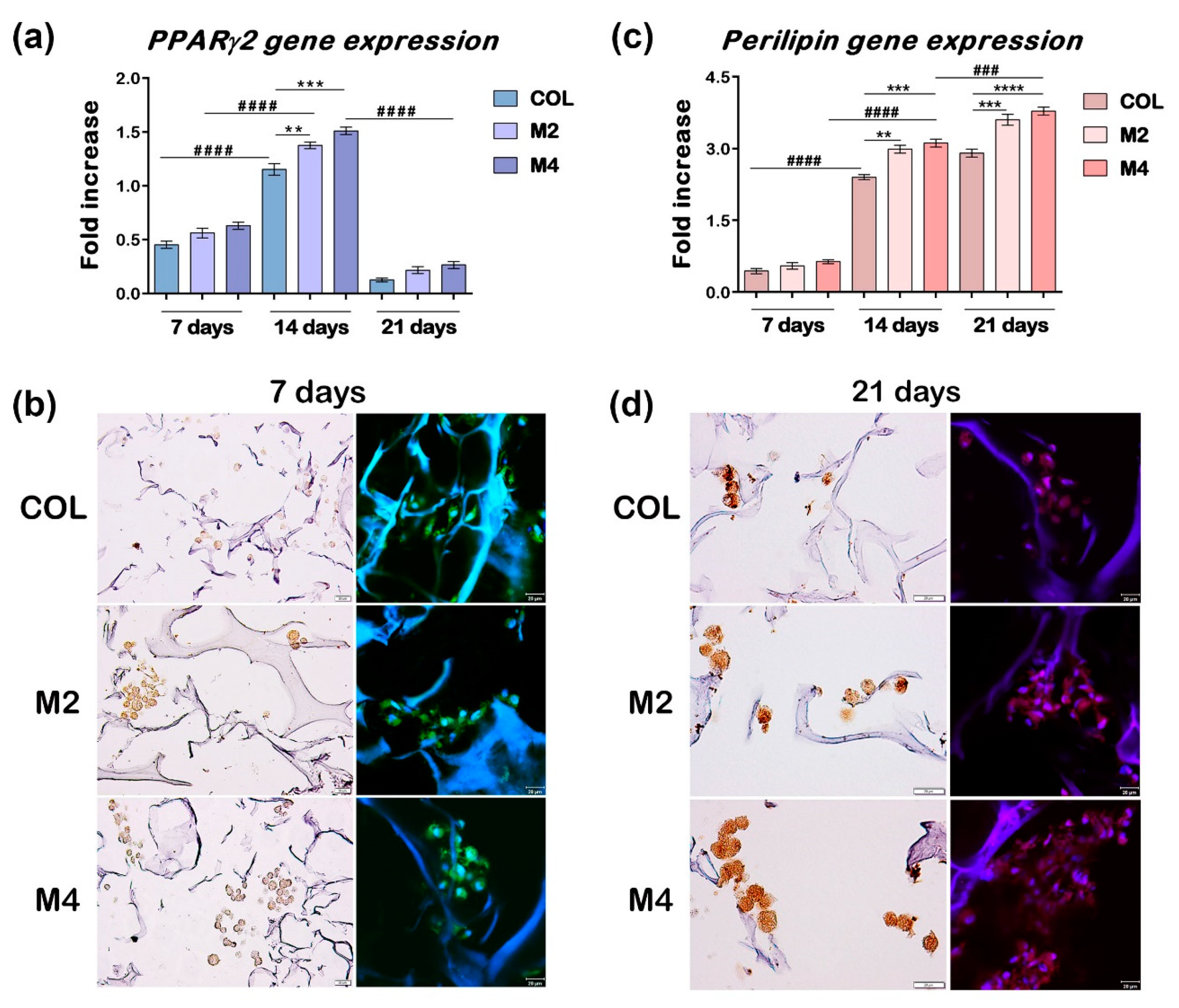
3.2.2. Gene Expression Evaluation of Early and Late Markers of Adipogenesis
3.2.3. Protein Expression Evaluation of Early and Late Markers of Adipogenesis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Langer, R.; Vacanti, J.P. Tissue engineering. Science 1993, 260, 920–926. [Google Scholar] [CrossRef] [Green Version]
- Olson, J.L.; Atala, A.; Yoo, J.J. Tissue engineering: Current strategies and future directions. Chonnam. Med. J. 2011, 47, 1–13. [Google Scholar] [CrossRef]
- Vacanti, J.P.; Vacanti, C.A. The history and scope of tissue engineering. In Principles of Tissue Engineering, 4th ed.; Lanza, R., Langer, R., Vacanti, J., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 3–8. [Google Scholar]
- Dinescu, S.; Hermenean, A.; Costache, M. Human adipose-derived stem cells for tissue engineering approaches: Current challenges and perspectives. In Stem Cells in Clinical Practice and Tissue Engineering; Sharma, R., Ed.; Intech Open: London, UK, 2017; pp. 293–314. [Google Scholar]
- Bacakova, L.; Zarubova, J.; Travnickova, M.; Musilkova, J.; Pajorova, J.; Slepicka, P.; Slepickova Kasalkova, N.; Svorcik, V.; Kolska, Z.; Motarjemi, H.; et al. Stem cells: Their source, potency and use in regenerative therapies with focus on adipose-derived stem cells—A review. Biotechnol. Adv. 2018, 36, 1111–1126. [Google Scholar] [CrossRef]
- Stromps, J.P.; Paul, N.E.; Rath, B.; Nourbakhsh, M.; Bernhagen, J.; Pallua, N. Chondrogenic differentiation of human adipose-derived stem cells: A new path in articular cartilage defect management? Biomed. Res. Int. 2014, 2014, 740926. [Google Scholar] [CrossRef]
- Dinescu, S.; Galateanu, B.; Radu, E.; Hermenean, A.; Lungu, A.; Stancu, I.C.; Jianu, D.; Tumbar, T.; Costache, M. A 3D porous gelatin-alginate-based-ipn acts as an efficient promoter of chondrogenesis from human adipose-derived stem cells. Stem. Cells Int. 2015, 2015, 1–17. [Google Scholar] [CrossRef]
- Han, T.T.; Toutounji, S.; Amsden, B.G.; Flynn, L.E. Adipose-derived stromal cells mediate in vivo adipogenesis, angiogenesis and inflammation in decellularized adipose tissue bioscaffolds. Biomaterials 2015, 72, 125–137. [Google Scholar] [CrossRef]
- Ignat, S.R.; Lazăr, A.D.; Şelaru, A.; Samoilă, I.; Vlăsceanu, G.M.; Ioniţă, M.; Radu, E.; Dinescu, S.; Costache, M. Versatile biomaterial platform enriched with graphene oxide and carbon nanotubes for multiple tissue engineering applications. Int. J. Mol. Sci. 2019, 20, 3868. [Google Scholar] [CrossRef] [Green Version]
- Storti, G.; Scioli, M.G.; Kim, B.S.; Orlandi, A.; Cervelli, V. Adipose-derived stem cells in bone tissue engineering: Useful tools with new applications. Stem. Cells Int. 2019, 2019, 3673857. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Guo, X.; Zhao, S.; Jin, Y.; Zhou, F.; Yuan, P.; Cao, L.; Wang, J.; Qiu, Y.; Sun, C.; et al. Differentiation of human adipose-derived stem cells into neuron/motoneuron-like cells for cell replacement therapy of spinal cord injury. Cell Death Dis. 2019, 10, 597. [Google Scholar] [CrossRef]
- Chang, Q.; Lu, F. A novel strategy for creating a large amount of engineered fat tissue with an axial vascular pedicle and a prefabricated scaffold. Med. Hypotheses 2012, 79, 267–270. [Google Scholar] [CrossRef]
- Herreros, M.D.; Garcia-Arranz, M.; Guadalajara, H.; De-La-Quintana, P.; Garcia-Olmo, D. Autologous expanded adipose-derived stem cells for the treatment of complex cryptoglandular perianal fistulas: A phase III randomized clinical trial (FATT 1: Fistula advanced therapy trial 1) and long-term evaluation. Dis. Colon. Rectum 2012, 55, 762–772. [Google Scholar] [CrossRef]
- Sanz-Baro, R.; Garcia-Arranz, M.; Guadalajara, H.; de la Quintana, P.; Herreros, M.D.; Garcia-Olmo, D. First-in-human case study: Pregnancy in women with Crohn’s perianal fistula treated with adipose-derived stem cells: A safety study. Stem. Cells Transl. Med. 2015, 4, 598–602. [Google Scholar] [CrossRef]
- Díaz-Agero Álvarez, P.J.; Bellido-Reyes, Y.A.; Sánchez-Girón, J.G.; García-Olmo, D.; García-Arranz, M. Novel bronchoscopic treatment for bronchopleural fistula using adipose-derived stromal cells. Cytotherapy 2016, 18, 36–40. [Google Scholar] [CrossRef]
- Flynn, L.; Woodhouse, K.A. Adipose tissue engineering with cells in engineered matrices. Organogenesis 2008, 4, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Lazar, A.D.; Dinescu, S.; Costache, M. Adipose tissue engineering and adipogenesis—A review. Rev. Biol. Biomed. Sci. 2018, 1, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Lefterova, M.I.; Lazar, M.A. New developments in adipogenesis. Trends Endocrinol. Metab. 2009, 20, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Lefterova, M.I.; Haakonsson, A.K.; Lazar, M.A.; Mandrup, S. PPARγ and the global map of adipogenesis and beyond. Trends Endocrinol. Metab. 2014, 25, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Farmer, S. Transcriptional control of adipocyte formation. Cell Metab. 2006, 4, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Arimura, N.; Horiba, T.; Imagawa, M.; Shimizu, M.; Sato, R. The peroxisome proliferator-activated receptor gamma regulates expression of the perilipin gene in adipocytes. J. Biol. Chem. 2004, 279, 10070–10076. [Google Scholar] [CrossRef] [Green Version]
- Brasaemle, D.L. Thematic review series: Adipocyte biology. The perilipin family of structural lipid droplet proteins: Stabilization of lipid droplets and control of lipolysis. J. Lipid. Res. 2007, 48, 2547–2559. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Engelmayr, G.; Auguste, D.T.; da Silva Ferreira, L.; Karp, J.M.; Saigal, R.; Langer, R. 3D Scaffolds. In Principles of Tissue Engineering, 4th ed.; Lanza, R., Langer, R., Vacanti, J., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 475–494. [Google Scholar]
- Choi, J.H.; Gimble, J.M.; Lee, K.; Marra, K.G.; Rubin, J.P.; Yoo, J.J.; Vunjak-Novakovic, G.; Kaplan, D.L. Adipose tissue engineering for soft tissue regeneration. Tissue Eng. Part B Rev. 2010, 16, 413–426. [Google Scholar] [CrossRef]
- He, Y.; Feng Lu, F. Development of synthetic and natural materials for tissue engineering applications using adipose stem cells. Stem. Cells Int. 2016, 2016, 5786257. [Google Scholar] [CrossRef] [Green Version]
- Rubin, J.P.; Bennett, J.M.; Doctor, J.S.; Tebbets, B.M.; Marra, K.G. Collagenous microbeads as a scaffold for tissue engineering with adipose-derived stem cells. Plast. Reconstr. Surg. 2007, 120, 414–424. [Google Scholar] [CrossRef]
- Ferraro, G.A.; De Francesco, F.; Nicoletti, G.; Paino, F.; Desiderio, V.; Tirino, V.; D’Andrea, F. Human adipose CD34+ CD90+ stem cells and collagen scaffold constructs grafted in vivo fabricate loose connective and adipose tissues. J. Cell Biochem. 2013, 114, 1039–1049. [Google Scholar] [CrossRef]
- Dinescu, S.; Albu Kaya, M.; Chitoiu, L.; Ignat, S.; Kaya, D.A.; Costache, M. Collagen-based hydrogels and their applications for tissue engineering and regenerative medicine. In Cellulose-Based Superabsorbent Hydrogels. Polymers and Polymeric Composites: A Reference Series; Mondal, M., Ed.; Springer: Cham, Switzerland, 2018; pp. 1–21. [Google Scholar]
- Ghica, M.V.; Kaya, M.G.A.; Dinu-Pîrvu, C.-E.; Lupuleasa, D.; Udeanu, D.I. Development, optimization and in vitro/in vivo characterization of collagen-dextran spongious wound dressings loaded with flufenamic acid. Molecules 2017, 22, 1552. [Google Scholar] [CrossRef]
- Vashi, A.V.; Abberton, K.M.; Thomas, G.P.; Morrison, W.A.; O’Connor, A.J.; Cooper-White, J.J.; Thompson, E.W. Adipose tissue engineering based on the controlled release of fibroblast growth factor-2 in a collagen matrix. Tissue Eng. 2006, 12, 3035–3043. [Google Scholar] [CrossRef] [Green Version]
- Kimura, Y.; Tsuji, W.; Yamashiro, H.; Toi, M.; Inamoto, T.; Tabata, Y. In situ adipogenesis in fat tissue augmented by collagen scaffold with gelatin microspheres containing basic fibroblast growth factor. J. Tissue Eng. Regen. Med. 2010, 4, 55–61. [Google Scholar] [CrossRef]
- Smith, W.L.; Meade, E.A.; DeWitt, D.L. Interactions of PGH synthase isozymes-1 and -2 with NSAIDs. Ann. N. Y. Acad. Sci. 1994, 744, 50–57. [Google Scholar] [CrossRef]
- Lehmann, J.M.; Lenhard, J.M.; Oliver, B.B.; Ringold, G.M.; Kliewer, S.A. Peroxisome proliferator-activated receptors α and γ are activated by indomethacin and other non-steroidal anti-inflammatory drugs. J. Biol. Chem. 1997, 272, 3406–3410. [Google Scholar] [CrossRef] [Green Version]
- Puhl, A.C.; Milton, F.A.; Cvoro, A.; Sieglaff, D.H.; Campos, J.C.L.; Bernardes, A.; Filgueira, C.S.; Lindemann, J.L.; Deng, T.; Neves, F.A.R.; et al. Mechanisms of peroxisome proliferator activated receptor γ regulation by non-steroidal anti-inflammatory drugs. Nucl. Recept. Signal 2015, 13, e004. [Google Scholar] [CrossRef] [Green Version]
- Styner, M.; Sen, B.; Xie, Z.; Case, N.; Rubin, J. Indomethacin promotes adipogenesis of mesenchymal stem cells through a cyclooxygenase independent mechanism. J. Cell Biochem. 2010, 111, 1042–1050. [Google Scholar] [CrossRef] [Green Version]
- Farmer, S.R. Regulation of PPARγ activity during adipogenesis. Int. J. Obes. 2005, 29, S13–S16. [Google Scholar] [CrossRef] [Green Version]
- Dinescu, S.; Ignat, S.; Lazar, A.D.; Marin, S.; Danilă, E.; Marin, M.M.; Udeanu, D.I.; Ghica, M.V.; Albu-Kaya, M.G.; Costache, M. Efficiency of multiparticulate delivery systems loaded with flufenamic acid designed for burn wound healing applications. J. Immunol. Res. 2019, 2019, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Gimble, J.M.; Katz, A.J.; Bunnell, B.A. Adipose-derived stem cells for regenerative medicine. Circ. Res. 2007, 100, 1249–1260. [Google Scholar] [CrossRef]
- Galateanu, B.; Dinescu, S.; Cimpean, A.; Dinischiotu, A.; Costache, M. Modulation of adipogenic conditions for prospective use of hADSCs in adipose tissue engineering. Int. J. Mol. Sci. 2012, 13, 15881–15900. [Google Scholar] [CrossRef] [Green Version]
- Rao, P.; Knaus, E.E. Evolution of nonsteroidal anti-inflammatory drugs (NSAIDs): Cyclooxygenase (COX) inhibition and beyond. J. Pharm. Sci. 2008, 11, 81s–110s. [Google Scholar] [CrossRef] [Green Version]
- Jaradat, M.S.; Wongsud, B.; Phornchirasilp, S.; Rangwala, S.M.; Shams, G.; Sutton, M.; Romstedt, K.J.; Noonan, D.J.; Feller, D.R. Activation of peroxisome proliferator-activated receptor isoforms and inhibition of prostaglandin H(2) synthases by ibuprofen, naproxen, and indomethacin. Biochem. Pharm. 2001, 62, 1587–1595. [Google Scholar] [CrossRef]
- Díaz-González, F.; Sánchez-Madrid, F. NSAIDs: Learning new tricks from old drugs. Eur. J. Immunol. 2015, 45, 679–686. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, Z.; Liu, H.; Zhu, Y.; Xia, D.; Wang, S.; Gu, R.; Wu, W.; Zhang, P.; Liu, Y.; et al. Low concentration flufenamic acid enhances osteogenic differentiation of mesenchymal stem cells and suppresses bone loss by inhibition of the NF-κB signaling pathway. Stem. Cell Res. 2019, 10, 213. [Google Scholar] [CrossRef]
- Kagan, G.; Huddlestone, L.; Wolstencroft, P. Flufenamic acid and placebo compared in rheumatoid arthritis and osteoarthritis. J. Int. Med. Res. 1981, 9, 253–256. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Systems | Composite Gel % | Microcapsules with FA % | |||
|---|---|---|---|---|---|
| Collagen | Dextran 1 | Glutaraldehyde 1 | FA 1 | ||
| COL | 0.8 | 1.2 | 0.006 | - | - |
| M2 | 0.8 | 1.2 | 0.006 | - | 30 |
| M4 | 0.8 | 1.2 | 0.006 | 0.5 | 30 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| PPARγ2 | 5’-TTACACAATGCTGGCCTCCTT-3’ | 5’-AGGCTTTCGCAGGCTCTTTAG-3’ |
| perilipin | 5’-ATGCTTCCAGAAGACCTACA-3’ | 5’-CAGCTCAGAAGCAATCTTTT-3’ |
| TBP | 5’-AGGCATCTGTCTTTGCACAC-3’ | 5’-GGGTCAGTCCAGTGCCATAA-3’ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazăr, A.D.; Dinescu, S.; Albu-Kaya, M.G.; Gharbia, S.; Hermenean, A.; Costache, M. Release of the Non-Steroidal Anti-Inflammatory Drug Flufenamic Acid by Multiparticulate Delivery Systems Promotes Adipogenic Differentiation of Adipose-Derived Stem Cells. Materials 2020, 13, 1550. https://doi.org/10.3390/ma13071550
Lazăr AD, Dinescu S, Albu-Kaya MG, Gharbia S, Hermenean A, Costache M. Release of the Non-Steroidal Anti-Inflammatory Drug Flufenamic Acid by Multiparticulate Delivery Systems Promotes Adipogenic Differentiation of Adipose-Derived Stem Cells. Materials. 2020; 13(7):1550. https://doi.org/10.3390/ma13071550
Chicago/Turabian StyleLazăr, Andreea D., Sorina Dinescu, Mădălina G. Albu-Kaya, Sami Gharbia, Anca Hermenean, and Marieta Costache. 2020. "Release of the Non-Steroidal Anti-Inflammatory Drug Flufenamic Acid by Multiparticulate Delivery Systems Promotes Adipogenic Differentiation of Adipose-Derived Stem Cells" Materials 13, no. 7: 1550. https://doi.org/10.3390/ma13071550