Ureolytic/Non-Ureolytic Bacteria Co-Cultured Self-Healing Agent for Cementitious Materials Crack Repair


Department of Civil and Environmental Engineering, Korea Advanced Institute of Science and Technology, 291 Daehak-ro, Yuseong-gu, Daejeon 34141, Korea
*
Author to whom correspondence should be addressed.
Materials 2018, 11(5), 782; https://doi.org/10.3390/ma11050782
Submission received: 18 March 2018
/
Revised: 21 April 2018
/
Accepted: 4 May 2018
/
Published: 11 May 2018
(This article belongs to the Section Advanced Composites)
Abstract
:The present study investigated the CaCO3 precipitation performance of ureolytic and non-ureolytic bacteria co-cultured as a self-healing agent for cementitious materials crack repair. Three different inoculum ratios of ureolytic Sporosarcina pasteurii and non-ureolytic Bacillus thuringiensis (10:0, 8:2, or 5:5) were used. The effect of coculturing ureolytic and non-ureolytic bacteria on microbial metabolism was investigated by measuring the rate of growth in urea-containing medium and the rate of NH4+ and CaCO3 production in urea–calcium lactate medium. The self-healing efficiency of co-cultured bacteria was examined by exposing cement mortar specimens with predefined cracks to media containing single urease-producing or co-cultured bacteria. The obtained results provide new findings, where CaCO3 precipitation is improved by co-culturing ureolytic and non-ureolytic bacteria, owing to the relatively faster growth rate of non-ureolytic bacteria. The crack filling rate correlated with the width of crack, in particular, specimens with a smaller crack width showed the faster filling effect, indicating that the crack width can be a dominant factor influencing the CaCO3 precipitation capacity of co-cultured bacteria.
1. Introduction
Concrete microcracks are a natural phenomenon and occur owing to many factors, such as changes in external force and drying shrinkage [1,2]. The formation of microcracks weakens the mechanical properties of concrete and makes the transport of moisture and chemicals through the matrix easier, accelerating concrete deterioration [3,4,5,6,7]. The durability performance of reinforced concrete structures is highly dictated by the ability to resist against such material transport. For example, sulfate ions penetrate through microcracks and cause internal pore expansion due to ettringite formation, known as sulfate attack [8,9], and chloride ions oxidize steel to accelerate the corrosion of steel bars embedded in concrete [10,11,12]. Repair work is mandatory to mitigate this problem and to extend the service life of concrete structures. While this often involves chemical repair materials (i.e., epoxy resins, polyesters, and polyurethane) [13,14], fundamental problems remain owing to difficulties in precisely locating microcracks, use of chemicals for healing microcracks being environmentally harmful, and the repair effect not being continuously maintained [15,16].
Against this backdrop, self-healing concrete has received significant attention over recent years owing to the potential of repair/maintenance-free concrete with the ability to heal microcracks, deemed as the main source of deterioration in concrete structures [17,18,19,20,21,22,23]. Bacteria incorporated in self-healing concrete act as healing agents, healing by precipitating CaCO3 at the cracked zone [3,4,17,18,19,22,24,25,26,27]. Self-healing concrete has a mechanism to heal cracks through forming CaCO3 by binding to CO32− formed by urea decomposition of bacteria and Ca2+ in concrete [28]. As a result, urea decomposition performance of bacteria is essential.
Accordingly, studies on self-healing concrete using bacteria preceded the employment of ureolytic bacteria as a self-healing organism [23]. For instance, Bang et al. reported that CaCO3 precipitated by decomposition of urea by ureolytic bacteria was effective to improve the strength and to repair microcracks of concrete [29]. A similar result was obtained by Ghosh et al., who reported an increase in the strength of mortar specimens when Shewanella species were used [30]. The increased strength of concrete induced by CaCO3 precipitation of Bacillus sphaericus, another known type of ureolytic bacteria, was also found to be accompanied by an enhancement in durability performance, compared with conventional treatment methods [31]. Self-healing induced by bacteria is not only limited to conventional concrete (i.e., those using Portland cement as a binder), but is also known to occur in fly ash-based concrete. This was intensively investigated by Chahal et al. [32], which found that compressive strength, water absorption and chloride permeability were affected by the added amount of S. pasteurii. Another aspect of self-healing concrete being widely studied concerns enhancing the survival rate of bacteria in the cement matrix, since it is directly linked to the self-healing capacity. One way to achieve this is to incorporate bacterial spores directly into concrete during the mixing process, along with a calcium source and urea [19]. In addition, microcapsulation of bacterial spores was found to be effective for ensuring the survival of the bacteria despite the internal pressure caused by concrete hardening when the bacteria spore is incorporated [19,22].
Despite a considerable number of studies investigating various means to improve the self-healing performance of bacteria, these studies used a single urease-producing bacterium, implying that there remains plenty of room for improvement with regards to microbial metabolism. In addition, in a self-healing concrete with bacteria, the limitations of healing effects with respect to crack width and crack generation time have been studied. In a self-healing concrete with bacteria, the limitations of healing effects with respect to crack width and crack generation time have been studied. Luo et al. reported that it is difficult to heal cracks of ≥0.8 mm and that it is hard to heal the crack as the crack width increases [25]. In addition, they showed that the crack healing effect is highly reduced when crack generation time is more than 60 days [25]. Considering these limitations on the crack healing performance with ureolytic bacteria, further studies are necessary to improve the crack healing rate. To this end, the present study investigated the self-healing agent using co-cultured bacteria as an effective means to improve microbial metabolism, thereby enhancing the CaCO3 precipitation efficiency.
2. Experimental Procedure
2.1. Bacterial Strains and Growth Medium
Sporosarcina pasteurii (ATCC 11859) and Bacillus thuringiensis (ATCC 10792) were the bacteria used in this study. S. pasteurii, a urease-producing organism, is known to survive in high alkaline environments of concrete [33], and decomposes urea (CH4N2O) into NH4+ and HCO3− to precipitate CaCO3 in the presence of Ca2+ [26]. B. thuringiensis, a non-urease producing organism, is a gram-positive bacteria that has a negative charge on the outer cell wall and can absorb Ca2+ [34]. However, this bacterial species cannot precipitate CaCO3 on its own because it does not decompose urea. For the preculturing of bacteria, S. pasteurii (obtained from the Korean Collection for Type Cultures, Jeongeup, South Korea) was added to 30 g/L Tryptic Soy Broth (Soybean–Casein Digest Medium) (TSB, Becton Dickinson, Franklin Lakes, NJ, USA) supplemented with 2% urea (333 mM) and cultured at 30 °C by shaking at 200 rpm in 650 mL cell culture flasks (SPL Life Science, Pocheon, South Korea) sealed with filter caps for sterile gas exchange. TSB was autoclaved at 121 °C and 1.5 psi for 20 min for sterilization, and the urea was filtered with a 0.22 μm pore size filter to prevent urea decomposition at high temperatures. B. thuringiensis (obtained from the Korea Collection for Type Cultures, Jeongeup, South Korea) was shake-cultured in 650 mL cell culture flasks (SPL Life Science, Pocheon, South Korea) sealed with filter caps for sterile gas exchange at 30 °C and 200 rpm in sterilized 25 g/L Miller’s Luria–Bertani broth (LB broth, Becton Dickinson, Franklin Lakes, NJ, USA).
2.2. Experimental Details
Three different inoculum ratios were employed to investigate the effect of co-culture. S. pasteurii alone was used as a control, and S. pasteurii and B. thuringiensis were inoculated at a ratio of 8:2 and 5:5, respectively. These ratios are hereinafter referred to as A, B, and C, respectively, for simplicity (Table 1). S. pasteurii and B. thuringiensis, having reached the exponential phase in the preculture, were inoculated at 0.5% in medium at A, B, and C ratios. Optical Density (OD) was measured using the Genesystm 30 Visible Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) at a wavelength of 600 nm to determine the growth pattern of the two species according to the inoculation ratios of A, B, and C. To observe the growth curve of bacteria at the tested inoculum ratios, the medium comprised 30 g/L TSB supplemented with 2% urea (333 mM) and was cultured in 650 mL cell culture flasks (SPL Life Science, Pocheon, South Korea) sealed with filter caps for sterile gas exchange at 30 °C with shaking at 200 rpm. Extraction of 1 mL of aqueous phase was performed every 4 h during 100 h across A, B, and C ratios. S. pasteurii and B. thuringiensis were incubated with Urea–TSB solid medium containing 3% TSB, 2% Urea, Urea agar base medium consisting of 1.8% Urea agar base and 1.8% agar for 1 day to confirm the antagonistic activity of the two bacteria. The crossing points of S. pasteurii and B. thuringiensis drawn in “M” and “=” shapes on the same medium were analyzed to investigate the effect of each bacteria on the growth of other bacteria.
The urea–calcium lactate medium was prepared for production of CaCO3. This medium contained 6 g/L Nutrient Broth (NB, Becton Dickinson, Franklin Lakes, NJ, USA) for the inclusion of various basic nutrient minerals, 333 mM urea (2%) for HCO3− production, and 91.7 mM calcium lactate for the supply of Ca2+. NB was autoclaved, and the urea and calcium lactate were sterilized using a 0.22 μm pore size filter. The inoculum of S. pasteurii and B. thuringiensis was added to urea–calcium lactate medium in 650 mL cell culture flasks (SPL Life Science, Pocheon, South Korea) sealed with filter caps for sterile gas exchange at ratios of A, B, and C and was incubated at 30 °C by shaking at 200 rpm.
To compare the ureolytic activity and the degree of Ca2+ consumption in CaCO3 production, the concentration of NH4+ produced by the urease expressed by S. pasteurii and the concentration of Ca2+ used in the production of calcium carbonate were measured. The concentration of NH4+ and Ca2+ dissolved in the aqueous phase in the culture solutions was measured using Metrohm 930 Compact IC Flex (Metrohm, Riverview, Herisau, Switzeland) and a cation exchange column. Five milliliters of the aqueous phase were extracted at an interval of 11 h for each of the tested ratios. Sampling was conducted until no further change in the NH4+ and Ca2+ concentration was observed. The supernatants of the extracted samples were obtained using a High Speed Refrigerated Centrifuge SUPRA 22K (Hanil Science Medical, Daejeon, South Korea) at 4800× g, 4 °C and then the NH4+ and Ca2+ concentration was analyzed.
Changes in the pH of the culture medium due to NH4+ production from urea hydrolysis in S. pasteurii were measured using a SevenGo Portable pH meter (Mettler Tolredo, Columbus, OH, USA). Four milliliters of aqueous phase were extracted at an interval of 4 h for A, B, and C ratios in urea–calcium lactate medium. The initial pH of each culture was kept identical at 7.5.
The charge of microbial cell surface upon precipitation of calcium carbonate was investigated by zeta potential measurement. The bacterial species were incubated in urea–lactate medium at a co-culture ratio of A, B, and C, and 5 mL of aqueous phase was extracted after 11 h, 22 h, and 33 h. Each sample was diluted 10-fold and a well dispersed sample was measured on a Nano ZSP/ZEN5602 (Malvern Instrument, Malvern, UK).
The amount of CaCO3 produced with the different S. pasteurii: B. thuringiensis ratios was measured to compare the difference in production for different ratios. CaCO3 was recovered and quantified after shake-incubating in urea–calcium lactate medium at 30 °C and 200 rpm for 33 h. For CaCO3 quantitation, the culture broth was collected on Whatman filter paper No. 2. The remaining cells and culture medium were removed by washing the filter paper with distilled water, followed by drying at 105 °C for 2 h before measuring the weight.
To compare the self-filling capacity of ratios A, B, and C, mortar specimens were prepared. Mortar specimens were fabricated with a water-to-cement ratio of 0.4 using a 5 × 5 × 5 cm3 mold. The mortar specimens were sealed with a plastic wrap after casting and were cured at room temperature for the initial 24 h to prevent water evaporation during setting, then cracks were introduced in the mortar specimens. Both the mortar specimens with predefined cracks and the culture container were sterilized by spraying with 70% ethanol. The sterile urea–calcium lactate medium for producing CaCO3 was placed in a culture vessel together with the mortar specimen, and the two species that reached the exponential phase were exposed to the culture vessel at ratios A, B, and C. The self-filling of cracks was observed at 500 magnification on a Digibird USB microscope ORT-500 (Digibird, Goyang, Korea) after 1 and 3 days of incubation at 30 °C. For comparing the effect of crack filling by co-culture with that of control, a bacteria-free urea–calcium lactate medium was prepared to cure mortar specimens as control. The other experimental procedures are the same except for the medium conditions.
The morphology of the co-cultured species and the precipitate formed around the bacteria were observed using a scanning electron microscope JSM-6300 (SEM, JEOL LTD, Akishima, Tokyo, Japan). Three samples with different co-culture ratios of the two species were shake-incubated in urea–calcium lactate medium for 33 h at 30 °C and 200 rpm. The precipitated CaCO3 was filtered through Whatman filter paper No. 2, collected, and dried at 105 °C in an oven to prepare powder samples for SEM (JEOL LTD, Akishima, Tokyo, Japan) and XRD (Rigaku, Tokyo, Japan) analyses. The XRD analysis was conducted using Rigaku D/MAX-2500 (Rigaku, Tokyo, Japan) with Cu-Kα radiation at a scan range of 5°–65° 2θ and a scan speed of 0.5°/min.
3. Results
3.1. Culture Growth
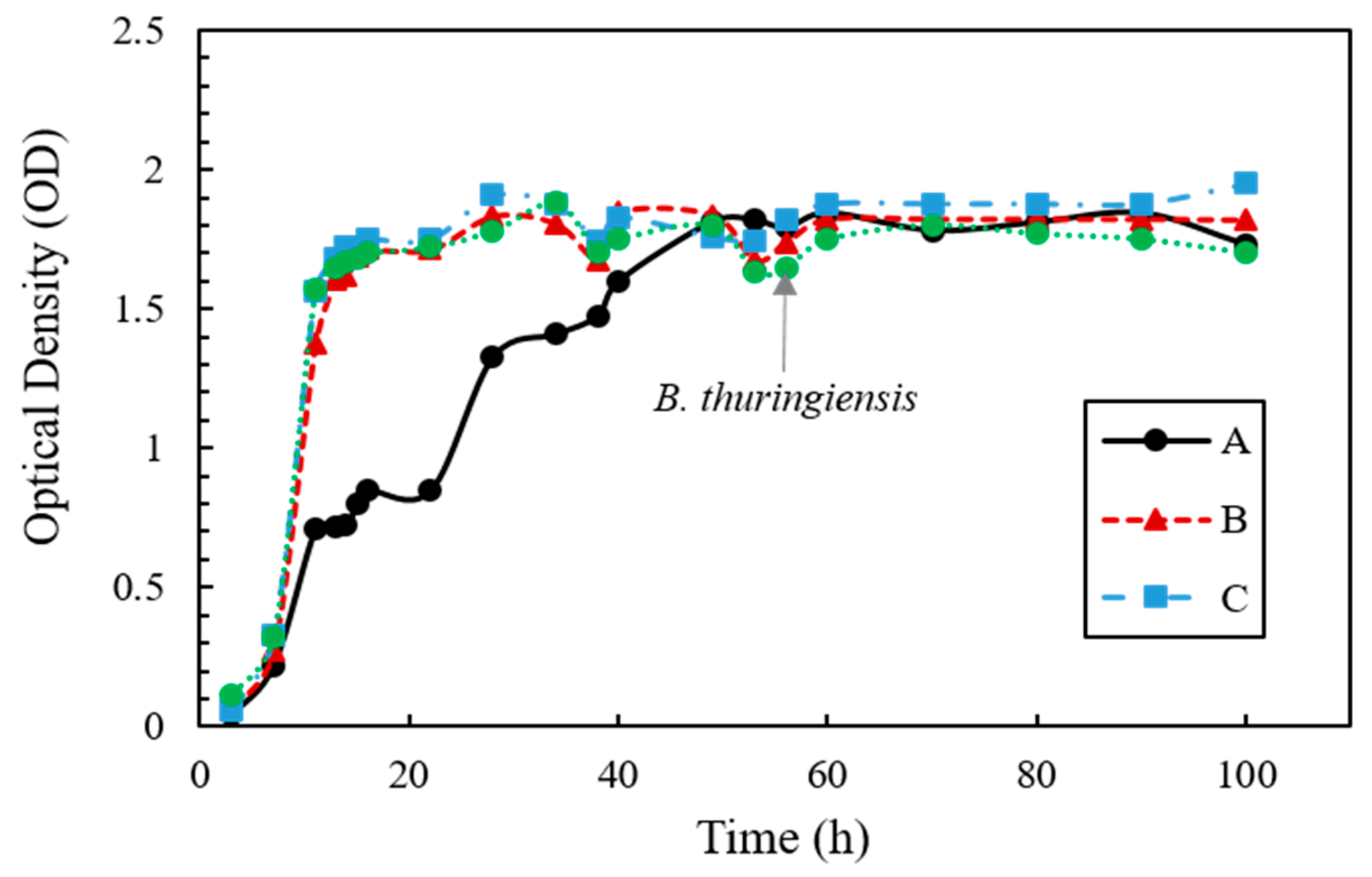
The growth rate of the bacteria in urea–TSB medium with inoculum ratios of A, B, and C are shown in Figure 1. The growth for A, where S. pasteurii was cultured alone, showed an exponential phase between 7 and 40 h of incubation. For B and C, where S. pasteurii and B. thuringiensis were co-cultured at 8:2 and 5:5, respectively, the OD was 1.37 and 1.56 at 11 h, respectively, corresponding to a value two-fold higher than for A (0.71). The OD of C was slightly higher than B at almost all sampling points, and B and C reached their peak OD (1.83 and 1.90 respectively) at 30 h. B. thuringiensis showed rapid growth until the first 10 h, followed by S. pasteurii, and the OD increased at 30 h in B and C. The bacterial growth for all of the tested co-culture ratios (A, B, and C) reached the stationary phase after 40 h, showing similar OD values from this point onward.
Urea–TSB and Urea Agar base culture flasks cultured with B. thurigiensis and S. pasteurii are shown in Figure 2. Inhibition zones which occurs when antagonistic activity [35] is progressed were not found in both TSB-Urea and Urea agar base medium. It can be inferred that B. thuringiensis and S. pasteurii did not inhibit growth with each other. In addition, district red colors were observed by urea decomposition in Urea agar base medium. Since these red colors were observed in the co-cultured contacts of both bacteria (Figure 2), it can be said that non-ureolytic bacteria, B. thuringiensis, did not inhibit the ureolytic activity of S. pasteurii.
3.2. Dissolved NH4+ and Ca2+ Concentration
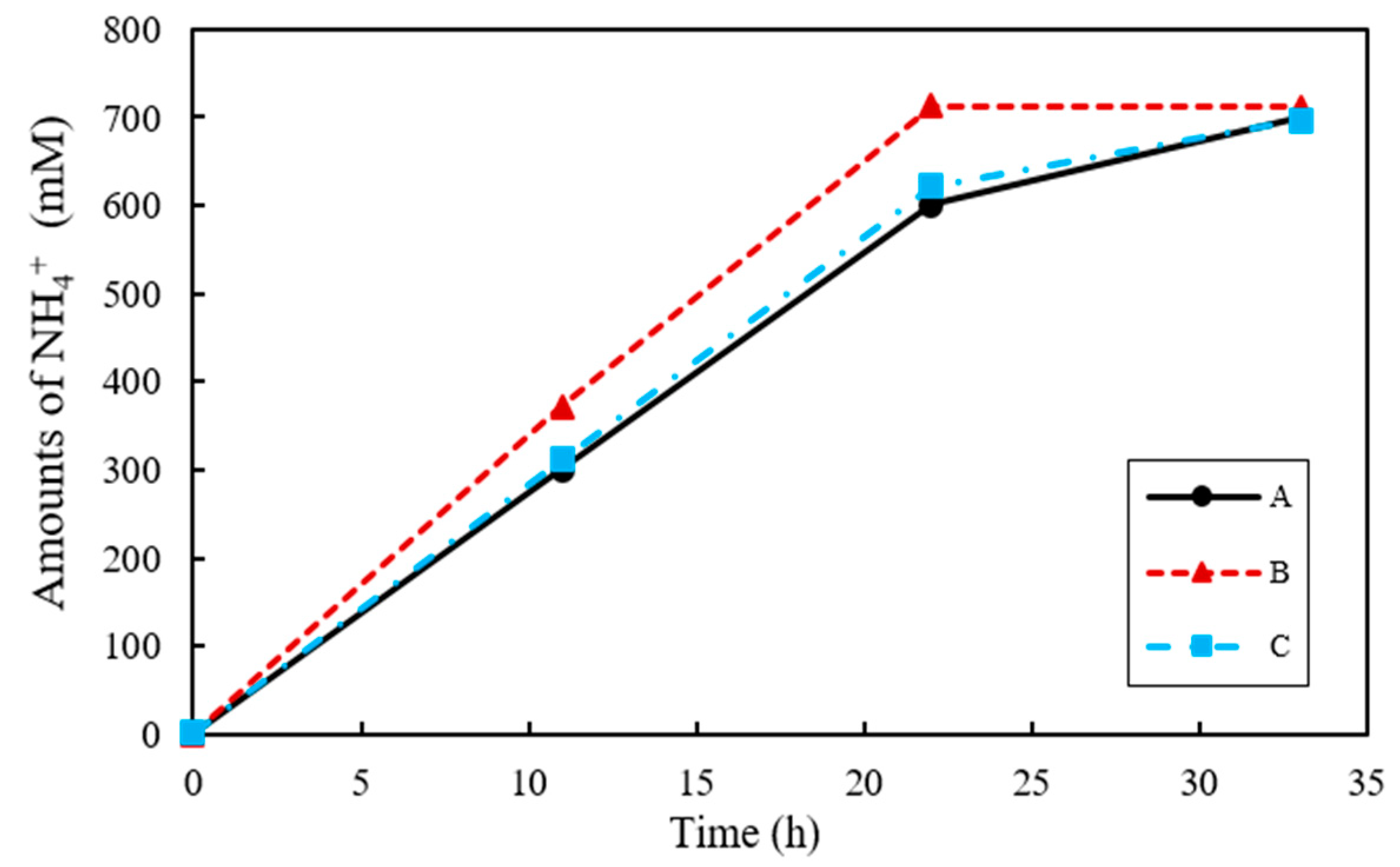
Urease is a common enzyme expressed by microorganisms, where 1 mol of urea (NH4CO3) is hydrated by urease to generate 2 mol of NH4+, 1 mol of HCO3−, and 2 mol of OH− [36]. As a result, the pH of culture media increases, with HCO3− converted to CO32− that then binds Ca2+ in the surroundings, and as a result CaCO3 is produced [37]. The urea degradation performance of the ureolytic bacteria S. pasteurii can be determined by measuring the concentration of NH4+ produced in the culture medium. The change in NH4+ concentration in each sample is shown in Figure 3. The NH4+ concentration for co-culture ratios A, B, and C reached 698.2, 711.1 and 696.3 mM at 33 h, respectively. However, the rate at which NH4+ was produced at 11 h was the highest for B at 371.4 mM, while those for A and C were 300.0 mM and 311.2 mM, respectively. Given that 1 mol of urea is decomposed into 2 mol of NH4+, 666 mM NH4+ was expected to be produced from 333 mM urea. Nevertheless, the amount of NH4+ produced was in fact greater than predicted at 104.83%, 106.77%, and 104.59% for A, B, and C, respectively, in relation to the amount of nitrogen initially supplied by the urea.
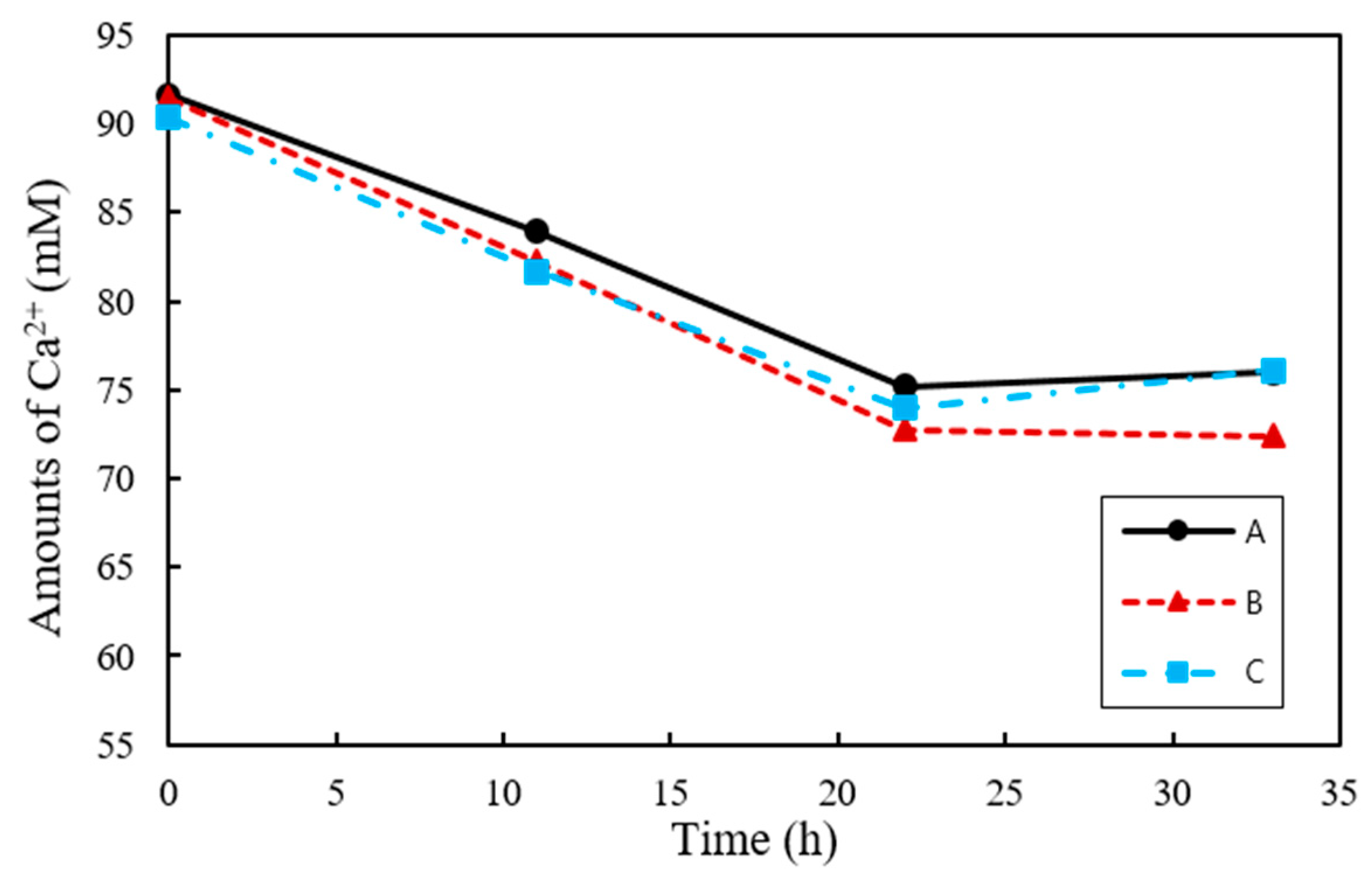
As urea is degraded by urease, CO32− is combined with Ca2+ in the culture solution to form CaCO3. Therefore, it is possible to deduce the amount of Ca2+ used in the production of CaCO3 by measuring the concentration of the remaining Ca2+. At 33 h the Ca2+ consumption was highest for B at 19.12 mM, followed by A and C at 16.44 and 14.25 mM, respectively (Figure 4). The Ca2+ concentrations showed no further change after 22 h and were not entirely exhausted in all samples.
3.3. CaCO3 Precipitation, Ph Change and Zeta Potential
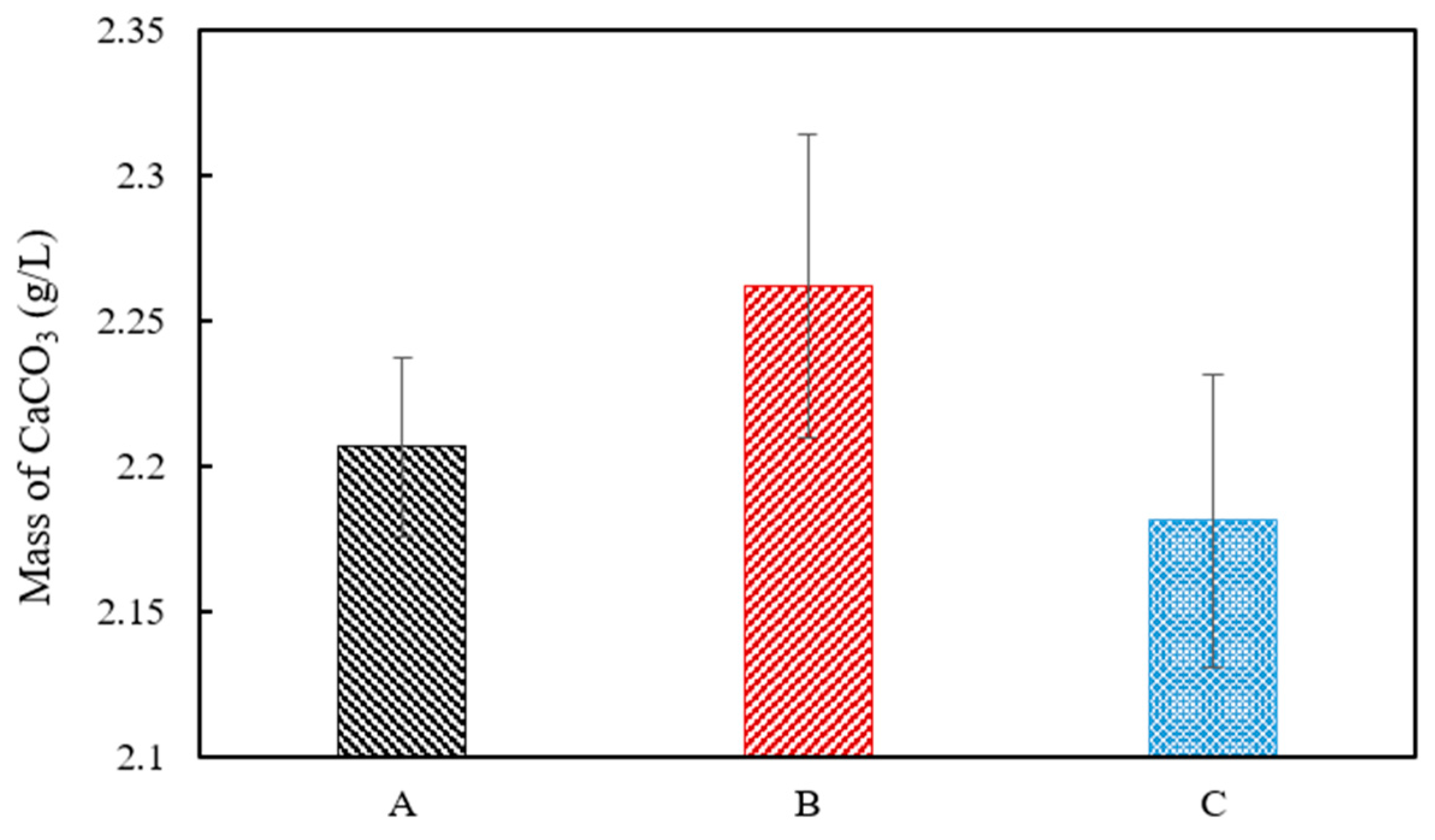
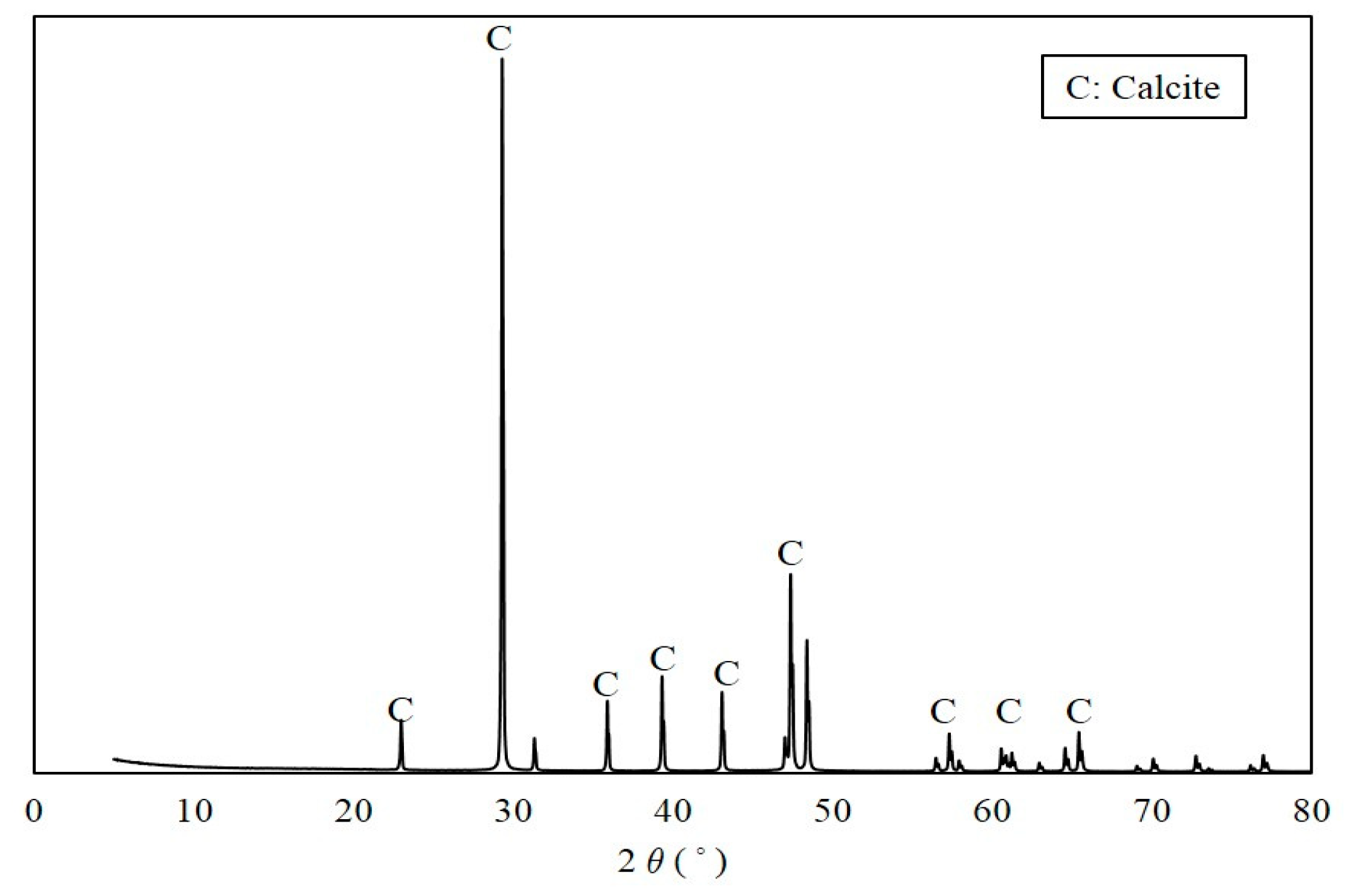
A white precipitate that formed in the urea–calcium lactate medium after culturing for 33 h was retrieved, and its weight was measured (Figure 5). The data values and error bars represent the averages and standard derivations of triplicate experiments. The weight of CaCO3 precipitated was the highest from ratio B cultures with S. pasteurii and B. thuringiensis at ratio of 8:2 (2.26 ± 0.05). The weight of recovered CaCO3 from ratio A and C cultures was 2.20 ± 0.03 and 2.18 ± 0.05, respectively. The weight of CaCO3 precipitated from ratio B cultures with a S. pasteurii:B. thuringiensis ratio of 8:2 was found to have the greatest weight. This is in close agreement with the Ca2+ consumption provided in Figure 4 at 33 h, showing that Ca2+ consumption was highest for B at 19.03 mM, followed by A and C at 15.65 and 14.25 mM, respectively. The XRD pattern of the white precipitate shown in Figure 6 confirms the presence of calcite (CaCO3, PDF# 01-072-1937), with noticeable peak intensity at 28°–30° 2θ.
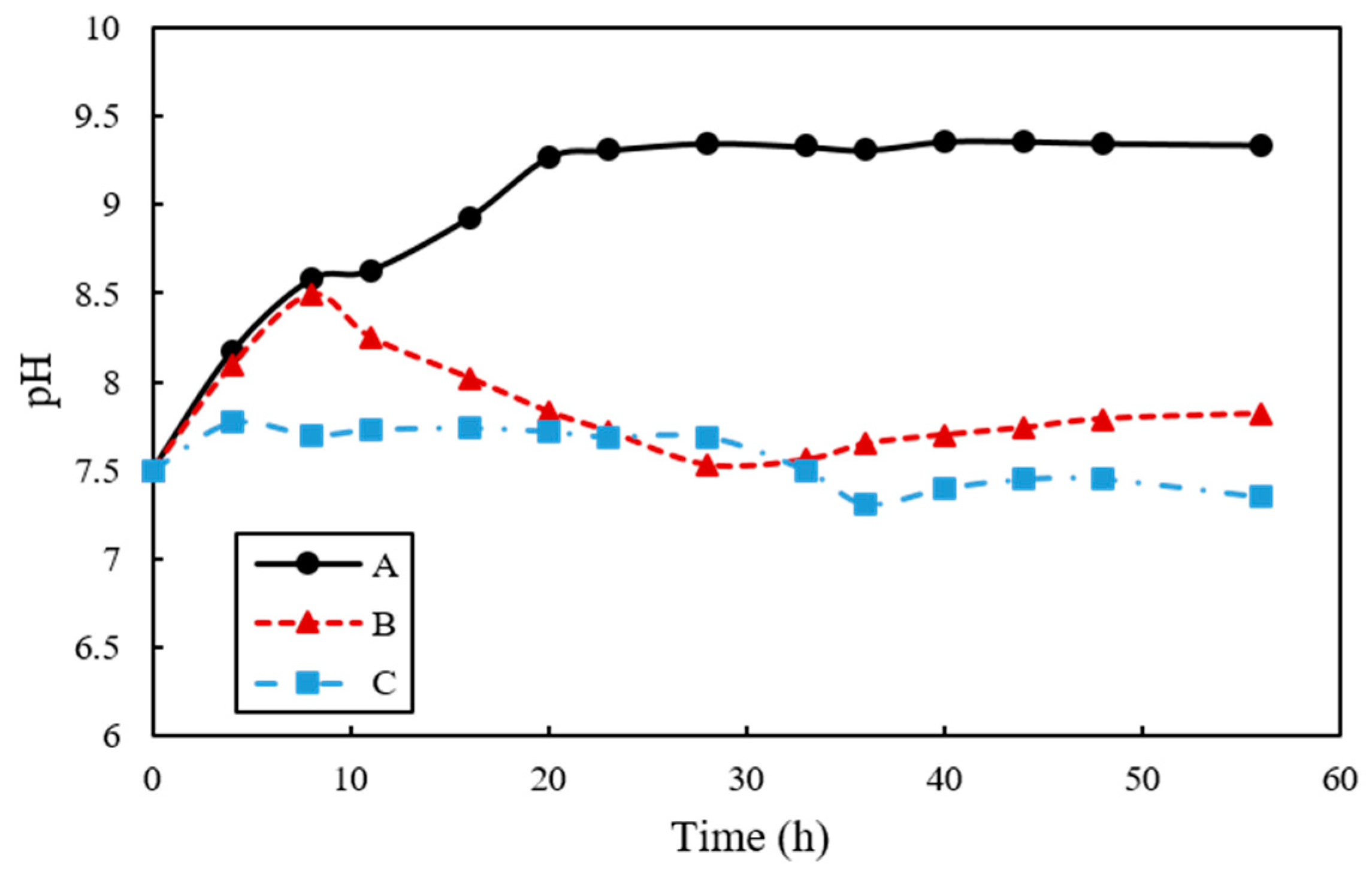
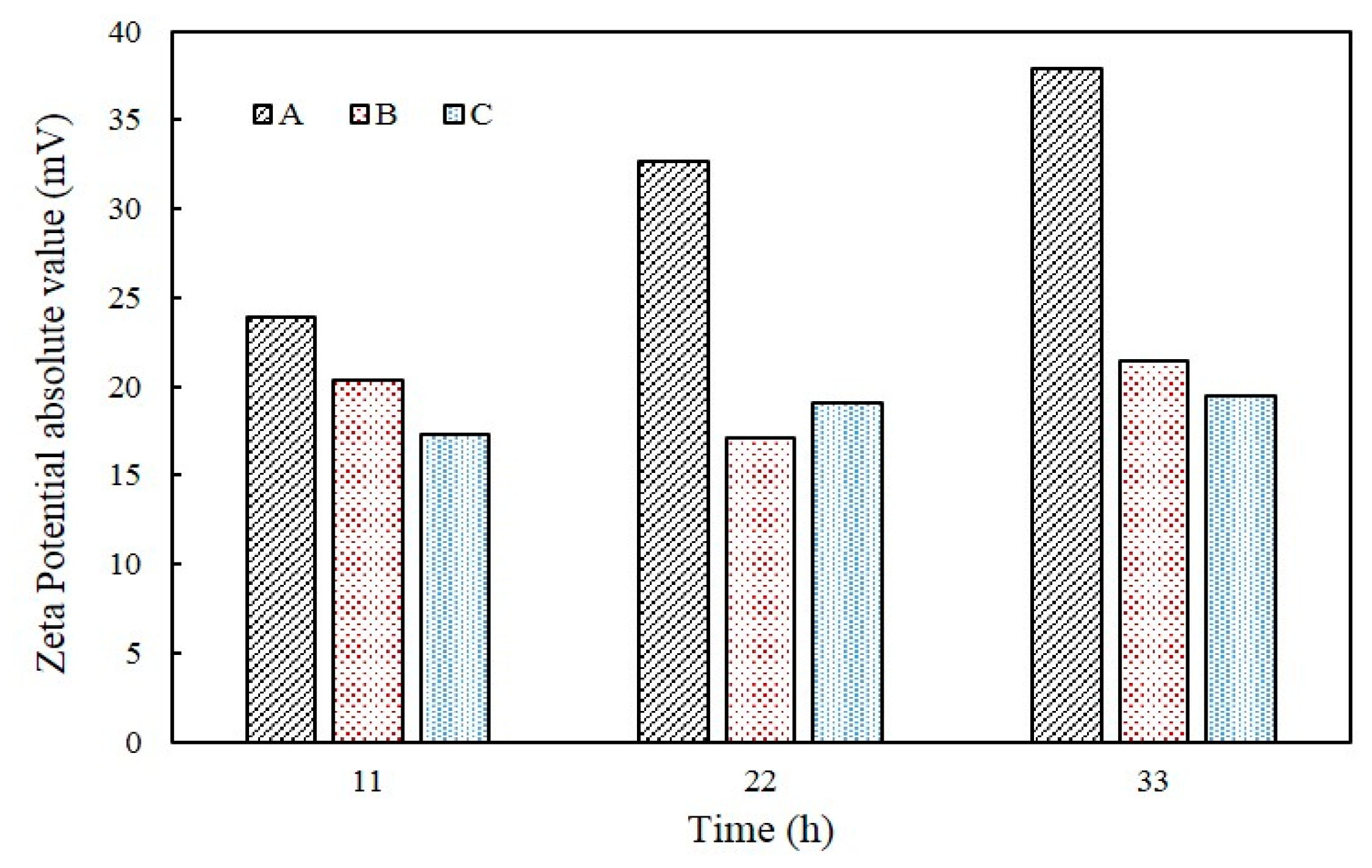
The evolution of pH in A, B, and C samples are shown in Figure 7. The pH in A tended to increase from 7.5 to 9.31 until 24 h. On the other hand, for B samples the pH increased to 8.49 until 8 h and then decreased to 7.56 after 35 h. Lastly, C showed almost no change in pH and was maintained at approximately 7.6. The zeta potential absolute value of A increased, but B and C tended to be maintained. The value of pH of A increased from 8.63 to 9.31 and the zeta potentials ranged from −23.9 mV to −37.9 mV between 11 h and 33 h. The value of zeta potential of B is −20.4 mV, −17.1 mV, and −21.4 mV, respectively, in pH 8.25 (11 h), 7.72 pH (22 h), and 7.65 pH (33 h) and that of C showed continuous value around −19 mV from 11 h to 33 h (Figure 8).
3.4. Effect of CaCO3 Precipitation in Crack on Cementitious Materials
The CaCO3 precipitation process in crack for the mortar specimens with predefined cracks in A, B, and C cultures is shown in Figure 10. It was observed that not only the composition ratio of the culture medium but also the shape of the cracks had a significant influence on the filling rate. For culture A-1 with a crack width of 0.12 mm, a CaCO3 precipitate was observed after three days of incubation. On the other hand, A-2 with a crack width of 0.04 mm showed the formation of CaCO3 and the crack filling even as early as one day of incubation. Similarly, B-1 showed complete crack filling performance after one day of incubation, and the dried bacteria and culture solution appeared prominently around the crack after three days of incubation. Meanwhile, the 0.19 mm wide crack combined with the C culture had the slowest rate of crack filling, showing scarce distribution of CaCO3 precipitates after three days of incubation. However, in the case of the C-2 mortar specimen with a 0.1 mm crack, filling of the crack had occurred due to deposition of CaCO3 after one day of incubation, with complete filling of the crack observed after three days of incubation. On the other hand, cracks in mortar specimens that was cured in bacteria-free urea–calcium lactate medium as control did not heal after one and three days.
4. Discussion
4.1. Microbial Metabolism of Co-Cultured Bacteria
As self-healing of concrete using bacteria relies on the formation of CaCO3 upon decomposition of urea by bacteria, it is essential to incorporate urea-decomposing bacteria. In addition, NH4+ and OH− formed through urea decomposition of bacteria induced an increase in the pH by producing CO2 and created an environment favorable for precipitation of CaCO3 [38]. Therefore, most of the studies on self-healing concrete using bacteria use urea-decomposing bacteria. In contrast, little attention has been given to the self-healing potential of non-ureolytic bacteria and its application.
Obst et al. (2006) studied that the reduction of net negative electric potential of bacteria cell surface is due to the adsorption of Ca2+ in culture solution on the cell surface, suggesting that the cell surface of the bacteria provides a nucleation site for the CaCO3 precipitation [39]. In this study, the amount of Ca2+ dissolved in the culture medium was the smallest in B, while A and C showed a similar value. This result is consistent with B showing the highest amount of CaCO3 production, as indicated by weight measurements of the precipitated CaCO3. Both results show that cocultivation of S. pasteurii and B. thuringiensis has a synergetic effect on CaCO3 precipitation, attributed to the presence of more nucleation sites. High cell density during the hydrolysis of urea was effective in increasing the diversity and size of CaCO3 crystals.
As S. pasteurii is a gram-positive bacteria, the surface of the cell wall is negatively charged by teichoic acid bound with phosphate, and Ca2+ in the culture solution is attracted to saturation around the cell wall [40]. Therefore, CaCO3 is produced as the Ca2+ present around bacteria reacts with CO32− formed upon urea degradation by the release of urease into the culture medium by S. pasteurii. However, as CaCO3 is formed around the cell membrane, the surface of S. pasteurii is saturated owing to continuous CaCO3 precipitation, effectively making it difficult for Ca2+ and urea to reach the surface of the bacteria. Consequently, the bacterial CaCO3 production activity is lowered. B. thuringiensis with a relatively faster growth rate may possess an enhanced binding capacity for Ca2+ and CO32− by providing additional nucleation sites outside of those provided by S. pasteurii embedded in CaCO3.
When B. thuringiensis and S. pasteurii are co-cultured, the concentrations of NH4+ and Ca2+ for CaCO3 precipitation were expected to vary according to urea degradation performance of bacteria. Since S. pasteurii was the only species capable of decomposing urea in this study, it was anticipated that the cultures that solely consist of S. pasteurii would display the highest concentration of NH4+ among the samples. However, the highest amount of NH4+ was observed in the B cultures at all sampling points during the measurement of NH4+ concentration. As shown in the OD results, B. thuringiensis had a relatively faster growth rate in comparison with S. pasteurii, more rapidly decomposing NB and thereby releasing additional NH4+. Consequently, the highest concentration of NH4+ was achieved in B cultures, where the amount of NH4+ was further increased by decomposition of NB as well as urea.
S. pasteurii and B. thuringiensis were inoculated into the culture medium at a ratio of A, B, and C, but the ratio of the two species in a culture medium after incubation is different from the initial inoculation ratio because of the different growth rates of each bacterium. However, rapid and high production of CaCO3 is linked to the initial inoculation ratio, and it may be reasonable to assume that the change in the ratio of two bacteria after co-culturing does not affect the outcome of the rapid effect of crack healing. In Figure 1, the optical density value of sample A cultured with only S. pasteurii was twice as low as that of B. thuringiensis-cultured sample. However, although a difference was observed in NH4+ production between each sample inoculated at ratios of A, B, and C, there was no significant difference in NH4+ production in each sample with respect to the difference in OD value. In addition, despite of the slow growth rate, S. pasteurii provided sufficient CO32− by degrading urea, and the nucleation site increased owing to the rapid growth of B. thuringiensis. Therefore, ratio B culture produced higher amount of CaCO3 in a short time (Figure 3 and Figure 4). Hence, to increase the effect of crack healing, co-cultivation of S. pasteurii and B. thuringiensis at a ratio of 8:2 is more effective.
The evolution of pH was also observed to be different according to the samples used. For instance, the pH in A and B was increased to 8.5 within 10 h of incubation, probably due to the increased concentrations of NH4+ and OH− that were produced by the hydrolysis of urea. However, the pH in B showed a sharp decrease after 10 h, presumably attributed to CaCO3 precipitation, where by water is released. Despite the decomposition of urea, the facultative anaerobic organism B. thuringiensis reduces culture medium pH due to the respiration process when oxygen is present, hence, the pH remained stationary and relatively unchanged in B and C that contained B. thuringiensis with a higher growth rate than S. pasteurii. It is interesting to note that the pH in C was constant from the beginning; even though the pH can be increased by increased OH− concentration from the ureolytic process, this can be compensated by the breathing of CO2 and production of CaCO3 by B. thuringiensis. These results are also closely related to the zeta potential change of each sample. This implied that increasing degradation of urea had an effect on increasing pH, indicating that increasing the degree of dispersion result in increasing zeta potential absolute value.
4.2. Morphology of CaCO3 Precipitated by Co-Cultured Bacteria
SEM results showed various crystal phases that formed around the nucleation site of the bacteria. In addition, the crystal phases of calcite with a hexagonal shape and other phases with spherical shape were observed. It should be noted that the size and crystallinity of the crystal phase are affected by the concentration of urea and CaCO3 in the culture medium; as the concentration of urea and calcium increases, the crystal size and crystallinity becomes lower [40]. This led to the formation of calcite, as well as a variety of other phases that were amorphous in nature.
4.3. Crack-Filling Efficiency of Co-Cultured Bacteria
Numerous studies have been performed to determine the performance of self-healing concrete with respect to the healing time and crack width variable, and it has been shown that the healing rate reduces as crack width and healing time increase. In addition, because crack healing performance is influenced by the change in bacterial species and healing environment conditions, crack width and healing time should be investigated in crack healing by co-culturing, as reported in this study. The crack filling process of mortar specimens with predefined cracks via CaCO3 precipitation in culture media showed different rates of filling. The smaller the width of the crack, the faster the observed crack filling was. Similarly, crack filling was less apparent even after three days of exposure to the culture medium, when the width of the crack was sufficiently large. The results indicate that the crack width is an important factor in crack filling via CaCO3 precipitation. A similar result was found in a previous study that reported that the maximum crack filling capacity achieved in a mortar specimen exposed to medium containing 5% B. sphaericus spores was in the range of 330–400 nm [22].
In this study, CaCO3 precipitation and the effect of self-filling in the mortar by the coculturing of ureolytic and non-ureolytic bacteria were investigated. As a result, it might be assumed that the increase of the quantity of CaCO3 precipitation and the crack filling effect can be improved by co-culture. However, in this experiment, although the possibility of co-culture to increase precipitation of CaCO3 was investigated, incorporating bacteria spore and urea in the cementitious materials for the self-healing concrete production was not achieved. Thus, further work should therefore include to apply spore cultures as well as bacterial co-culture to the exploration of potential applications.
5. Conclusions
The present study investigated the effect of coculturing ureolytic and non-ureolytic bacteria on the self-filling of cement mortar. To this aim, urea–calcium lactate media containing S. pasteurii and B. thuringiensis were prepared at an inoculation ratio of 10:0, 8:2, or 5:5, to which mortar specimens with predefined cracks were exposed. The obtained results indicate enhanced CaCO3 precipitation by coculturing ureolytic and non-ureolytic bacteria and elucidate the principles involved in such phenomena. The findings of this study can be summarized as below:
- (1)
- The urea–calcium lactate medium in which B. thuringiensis and S. pasteurii were co-cultured showed enhanced consumption of Ca2+, thereby leading to a higher extent of CaCO3 precipitation in comparison cultures solely containing S. pasteurii.
- (2)
- The faster growth rate of the non-ureolytic organism B. thuringiensis compared to the ureolytic bacteria led to more rapid respiration producing CO2 which decrease pH and provided nucleation site that effectively enhanced CaCO3 precipitation.
- (3)
- The rapid growth of B. thuringiensis was also effective for NB decomposition, as evidenced by the higher concentration of NH4+ in the coculturing medium, whereby production of CO32− and CaCO3 were significantly increased.
- (4)
- The performance of the bacteria for crack filling in mortar specimens was largely dictated by the width of crack. Even though a higher amount of CaCO3 precipitation was achieved in the co-cultured medium, the rate of crack filling was found to be more dependent on the crack width than the co-culture ratios.
Author Contributions
H.M.S. and H.Y.K. came up with main conceptual idea, performed the planned experiments and wrote the manuscript with support from S.M.P. and H.K.L. All of them contributed to the implementation of the paper, to discussion of the results and to the preparing of the figures, tables and the manuscript.
Acknowledgments
This research was supported by the National Research Foundation (NRF) of the Korean government funded by the Korean government (Ministry of Science, ICT & Future Planning) (2015R1A2A1A10055694).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Benboudjema, F.; Meftah, F.; Torrenti, J.M. Interaction between drying, shrinkage, creep and cracking phenomena in concrete. Eng. Struct. 2005, 27, 239–250. [Google Scholar] [CrossRef]
- Cusson, D.; Hoogeveen, T. Internal curing of high-performance concrete with pre-soaked fine lightweight aggregate for prevention of autogenous shrinkage cracking. Cem. Concr. Res. 2008, 38, 757–765. [Google Scholar] [CrossRef]
- Jonkers, H.M. Self healing concrete: A biological approach. In Self Healing Materials; Springer: Dordrecht, The Netherlands, 2007; pp. 195–204. [Google Scholar]
- Van Tittelboom, K.; De Belie, N.; De Muynck, W.; Verstraete, W. Use of bacteria to repair cracks in concrete. Cem. Concr. Res. 2010, 40, 157–166. [Google Scholar] [CrossRef]
- Şahmaran, M.; Li, V.C. Influence of microcracking on water absorption and sorptivity of ECC. Mater. Struct. 2009, 42, 593–603. [Google Scholar] [CrossRef]
- Jang, J.G.; Park, S.M.; Lee, H.K. Physical barrier effect of geopolymeric waste form on diffusivity of cesium and strontium. J. Hazard. Mater. 2016, 318, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Yao, W. Influence of damage degree on self-healing of concrete. Constr. Build. Mater. 2008, 22, 1137–1142. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, J.; Lv, Y.; Wang, D.; Ye, J. Study on the expansion of concrete under attack of sulfate and sulfate–chloride ions. Constr. Build. Mater. 2013, 39, 26–32. [Google Scholar] [CrossRef]
- Basista, M.; Weglewski, W. Micromechanical modeling of sulphate corrosion in concrete: Influence of ettringite forming reaction. Theor. Appl. Mech. 2008, 35, 29–52. [Google Scholar] [CrossRef]
- De Anna, P.L. The effects of water and chloride ions on the electrochemical behaviour of iron and 304L stainless steel in alcohols. Corros. Sci. 1985, 25, 43–53. [Google Scholar] [CrossRef]
- Ma, Y.; Li, Y.; Wang, F. Corrosion of low carbon steel in atmospheric environments of different chloride content. Corros. Sci. 2009, 51, 997–1006. [Google Scholar] [CrossRef]
- Lee, N.K.; Lee, H.K. Influence of the slag content on the chloride and sulfuric acid resistances of alkali-activated fly ash/slag paste. Cem. Concr. Compos. 2016, 72, 168–179. [Google Scholar] [CrossRef]
- Wong, T.S.; Kang, S.H.; Tang, S.K.; Smythe, E.J.; Hatton, B.D.; Grinthal, A.; Aizenberg, J. Bioinspired self-repairing slippery surfaces with pressure-stable omniphobicity. Nature 2011, 477, 443. [Google Scholar] [CrossRef] [PubMed]
- Morgan, D.R. Compatibility of concrete repair materials and systems. Constr. Build. Mater. 1996, 10, 57–67. [Google Scholar] [CrossRef]
- Basheer, L.; Cleland, D.J. Freeze–thaw resistance of concretes treated with pore liners. Constr. Build. Mater. 2006, 20, 990–998. [Google Scholar] [CrossRef]
- Ibrahim, M.; Al-Gahtani, A.S.; Maslehuddin, M.; Almusallam, A.A. Effectiveness of concrete surface treatmentmaterials in reducing chloride-induced reinforcement corrosion. Constr. Build. Mater. 1997, 11, 443–451. [Google Scholar] [CrossRef]
- Wiktor, V.; Jonkers, H.M. Quantification of crack-healing in novel bacteria-based self-healing concrete. Cem. Concr. Compos. 2011, 33, 763–770. [Google Scholar] [CrossRef]
- Wang, J.; Van Tittelboom, K.; De Belie, N.; Verstraete, W. Use of silica gel or polyurethane immobilized bacteria for self-healing concrete. Constr. Build. Mater. 2012, 26, 532–540. [Google Scholar] [CrossRef]
- Jonkers, H.M.; Thijssen, A.; Muyzer, G.; Copuroglu, O.; Schlangen, E. Application of bacteria as self-healing agent for the development of sustainable concrete. Ecol. Eng. 2010, 36, 230–235. [Google Scholar] [CrossRef]
- Wang, J.Y.; De Belie, N.; Verstraete, W. Diatomaceous earth as a protective vehicle for bacteria applied for self-healing concrete. J. Ind. Microbiol. Biotechnol. 2012, 39, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, H.W.; Jooss, M. Permeability and self-healing of cracked concrete as a function of temperature and crack width. Cem. Concr. Res. 2003, 33, 981–985. [Google Scholar] [CrossRef]
- Wang, J.Y.; Soens, H.; Verstraete, W.; De Belie, N. Self-healing concrete by use of microencapsulated bacterial spores. Cem. Concr. Res. 2014, 56, 139–152. [Google Scholar] [CrossRef]
- Kim, H.K.; Park, S.J.; Han, J.I.; Lee, H.K. Microbially mediated calcium carbonate precipitation on normal and lightweight concrete. Constr. Build. Mater. 2013, 38, 1073–1082. [Google Scholar] [CrossRef]
- Wang, J. Self-Healing Concrete by Means of Immobilized Carbonate Precipitating Bacteria. Ph.D. Thesis, Ghent University, Ghent, Belgium, 2013. [Google Scholar]
- Luo, M.; Qian, C.X.; Li, R.Y. Factors affecting crack repairing capacity of bacteria-based self-healing concrete. Constr. Build. Mater. 2015, 87, 1–7. [Google Scholar] [CrossRef]
- Achal, V.; Mukherjee, A.; Basu, P.C.; Reddy, M.S. Strain improvement of Sporosarcina pasteurii for enhanced urease and calcite production. J. Ind. Microbiol. Biotechnol. 2009, 36, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Achal, V.; Mukerjee, A.; Reddy, M.S. Biogenic treatment improves the durability and remediates the cracks of concrete structures. Constr. Build. Mater. 2013, 48, 1–5. [Google Scholar] [CrossRef]
- Mobley, H.L.; Hausinger, R.P. Microbial ureases: Significance, regulation, and molecular characterization. Microbiol. Rev. 1989, 53, 85–108. [Google Scholar] [PubMed]
- Bang, S.S.; Galinat, J.K.; Ramakrishnan, V. Calcite precipitation induced by polyurethane-immobilized Bacillus pasteurii. Enzym. Microb. Technol. 2001, 28, 404–409. [Google Scholar] [CrossRef]
- Ghosh, S.; Biswas, M.; Chattopadhyay, B.D.; Mandal, S. Microbial activity on the microstructure of bacteria modified mortar. Cem. Concr. Compos. 2009, 31, 93–98. [Google Scholar] [CrossRef]
- De Muynck, W.; Debrouwer, D.; De Belie, N.; Verstraete, W. Bacterial carbonate precipitation improves the durability of cementitious materials. Cem. Concr. Res. 2008, 38, 1005–1014. [Google Scholar] [CrossRef]
- Chahal, N.; Siddique, R.; Rajor, A. Influence of bacteria on the compressive strength, water absorption and rapid chloride permeability of fly ash concrete. Constr. Build. Mater. 2012, 28, 351–356. [Google Scholar] [CrossRef]
- Tiwari, P.K.; Joshi, K.; Rehman, R.; Bhardwaj, V.; Shamsudheen, K.V.; Sivasubbu, S.; Scaria, V. Draft genome sequence of urease-producing Sporosarcina pasteurii with potential application in biocement production. Genome Announc. 2014, 2, e01256-13. [Google Scholar] [CrossRef] [PubMed]
- Schnepf, E.; Crickmore, N.V.; Van Rie, J.; Lereclus, D.; Baum, J.; Feitelson, J.; Zeigler, D.; Dean, D.H. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 775–806. [Google Scholar] [PubMed]
- Živković, S.; Stojanović, S.; Ivanović, Ž.; Gavrilović, V.; Popović, T.; Balaž, J. Screening of antagonistic activity of microorganisms against Colletotrichum acutatum and Colletotrichum gloeosporioides. Arch. Biol. Sci. 2010, 62, 611–623. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Cheng, X.; Guo, H. Heavy metal removal by biomineralization of urease producing bacteria isolated from soil. Int. Biodeterior. Biodegrad. 2013, 76, 81–85. [Google Scholar] [CrossRef]
- Achal, V.; Mukherjee, A.; Reddy, M.S. Biocalcification by Sporosarcina pasteurii using corn steep liquor as the nutrient source. Ind. Biotechnol. 2010, 6, 170–174. [Google Scholar] [CrossRef]
- Kim, H.J.; Eom, H.J.; Park, C.; Jung, J.; Shin, B.; Kim, W.; Kim, W.; Chung, N.; Choi, I.L.; Park, W. Calcium carbonate precipitation by Bacillus and Sporosarcina strains isolated from concrete and analysis of the bacterial community of concrete. J. Microbiol. Biotechnol. 2016, 26, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Obst, M.; Dittrich, M.; Kuehn, H. Calcium adsorption and changes of the surface microtopography of cyanobacteria studied by AFM, CFM, and TEM with respect to biogenic calcite nucleation. Geochem. Geophys. Geosyst. 2006, 7, Q06011. [Google Scholar] [CrossRef]
- De Muynck, W.; Verbeken, K.; De Belie, N.; Verstraete, W. Influence of urea and calcium dosage on the effectiveness of bacterially induced carbonate precipitation on limestone. Ecol. Eng. 2010, 36, 99–111. [Google Scholar] [CrossRef]
Figure 1.
Optical density measurement for cell growth in urea–TSB medium over time.

Figure 2.
Bacteria growth images in: (a) urea–TSB solid medium; and (b) urea agar base medium.

Figure 3.
The changes of dissolved NH4+ concentration in urea–calcium lactate medium over time.

Figure 4.
Ca2+ consumption by S. pasteurii and B. thuringiensis in urea–calcium lactate medium over time.
Figure 4.
Ca2+ consumption by S. pasteurii and B. thuringiensis in urea–calcium lactate medium over time.

Figure 5.
Dry weight of precipitated CaCO3 at urea–calcium lactate medium at 33 h.

Figure 6.
XRD pattern of CaCO3 precipitated in urea–calcium lactate medium at 33 h.

Figure 7.
The changes of pH in urea–calcium lactate medium over time.

Figure 8.
The changes of zeta potential in urea–calcium lactate medium over time.

Figure 9.
SEM images of calcite precipitated in the co-culture medium at magnification of: (a) 30,000; and (b) 7000.
Figure 9.
SEM images of calcite precipitated in the co-culture medium at magnification of: (a) 30,000; and (b) 7000.

Figure 10.
Microscopic observation of crack healing after one and three days of incubation.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Microbial inoculation ratio by volume ratio.
| Media Code | S. pasteurii | B. thuringiensis |
|---|---|---|
| A | 10 | 0 |
| B | 8 | 2 |
| C | 5 | 5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Son, H.M.; Kim, H.Y.; Park, S.M.; Lee, H.K. Ureolytic/Non-Ureolytic Bacteria Co-Cultured Self-Healing Agent for Cementitious Materials Crack Repair. Materials 2018, 11, 782. https://doi.org/10.3390/ma11050782
AMA Style
Son HM, Kim HY, Park SM, Lee HK. Ureolytic/Non-Ureolytic Bacteria Co-Cultured Self-Healing Agent for Cementitious Materials Crack Repair. Materials. 2018; 11(5):782. https://doi.org/10.3390/ma11050782
Chicago/Turabian StyleSon, Hyeong Min, Ha Yeon Kim, Sol Moi Park, and Haeng Ki Lee. 2018. "Ureolytic/Non-Ureolytic Bacteria Co-Cultured Self-Healing Agent for Cementitious Materials Crack Repair" Materials 11, no. 5: 782. https://doi.org/10.3390/ma11050782
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.