
Characterization of a Type II L-Asparaginase from the Halotolerant Bacillus subtilis CH11
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria Strains, Medium, and Chemicals
2.2. Bioinformatic Analysis
2.3. Cloning of the ansZP21 Gene Encoding L-ASNasaZP21
2.4. Expression and Purification of L-ASNasaZP21
2.5. Molecular Weight Determination
2.6. SDS-PAGE and Zymography
2.7. L-asparaginase Activity and Protein Assay
2.8. Biochemical Characterization
2.9. Data Collection and Analysis
3. Results and Discussion
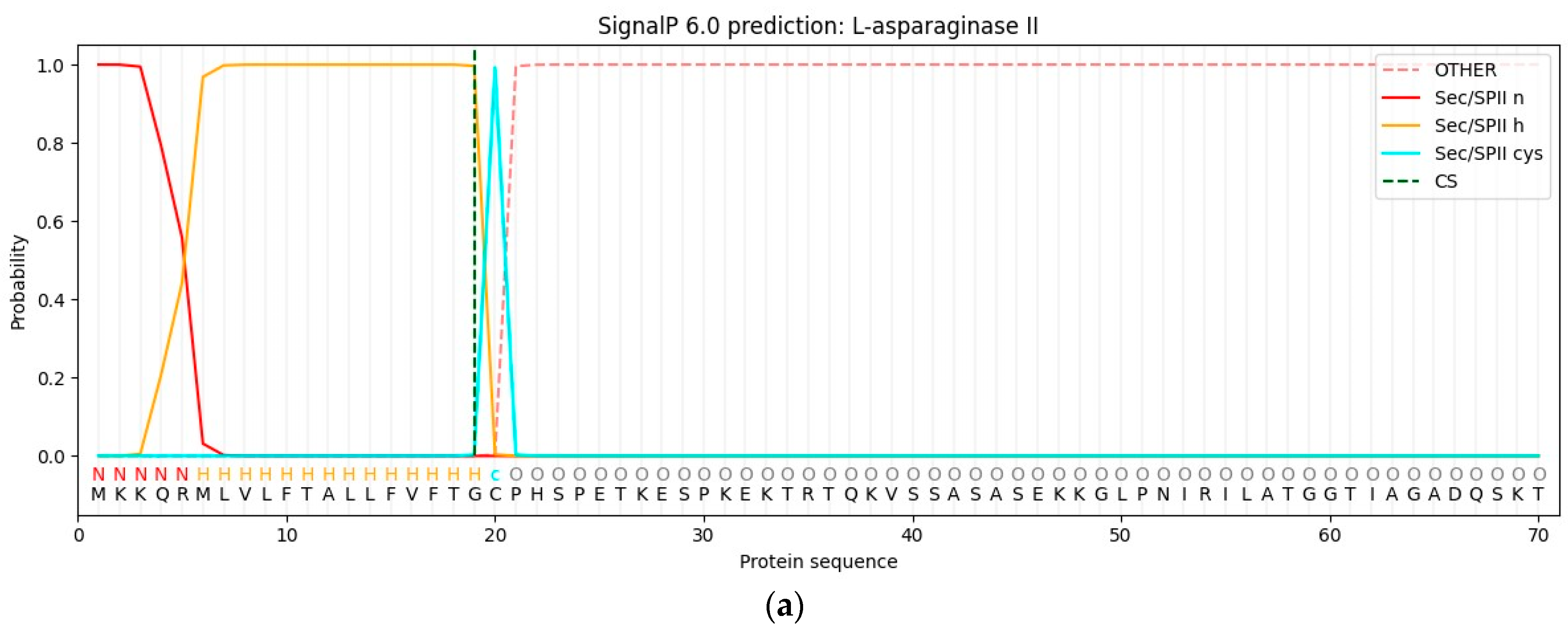
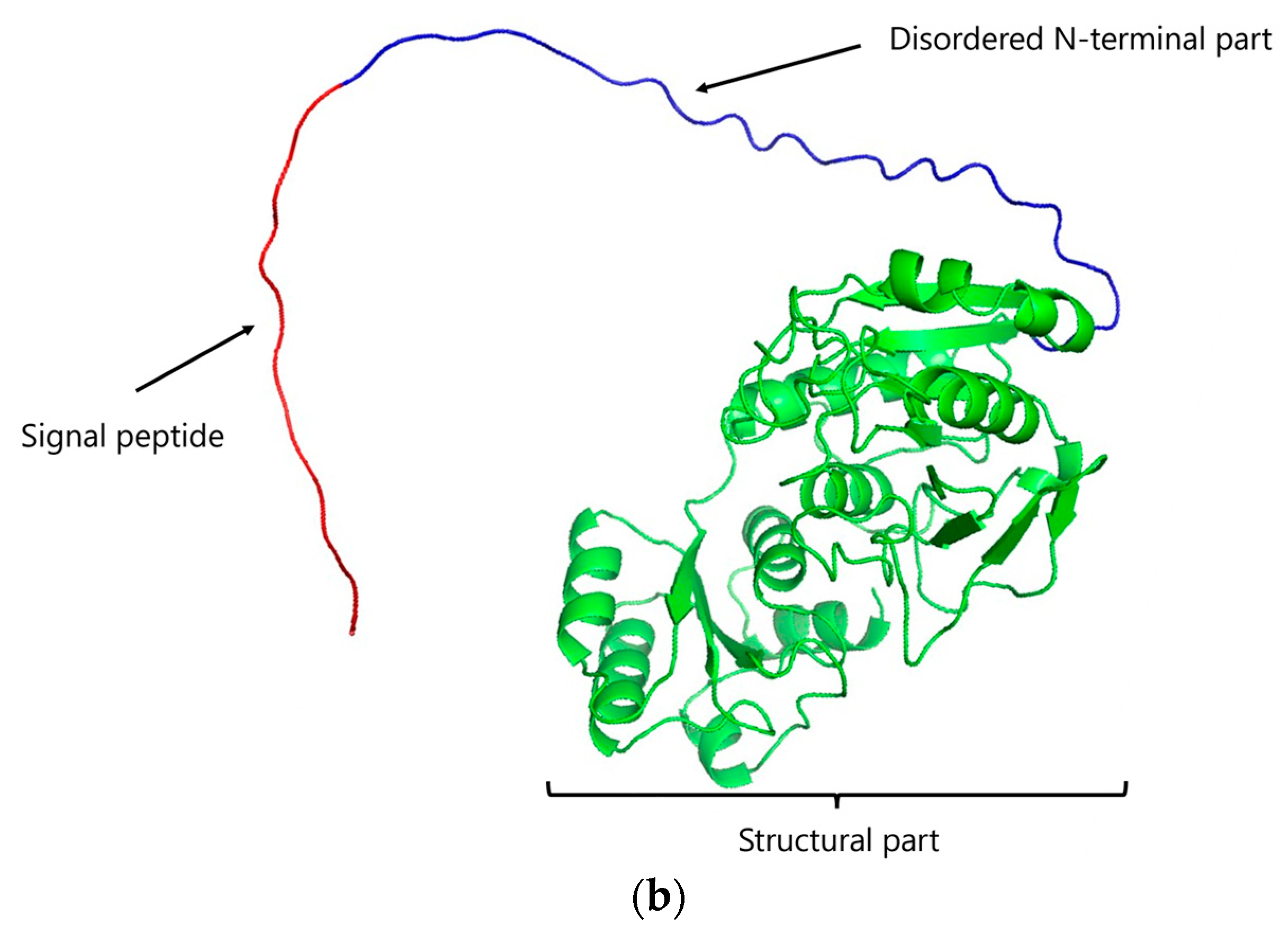
3.1. Cloning of the ansZP21 Gene and Sequence Analysis
3.2. Expression and Purification of L-ASNasaZP21
3.3. Molecular Weight Determination, SDS-PAGE, and Zymography
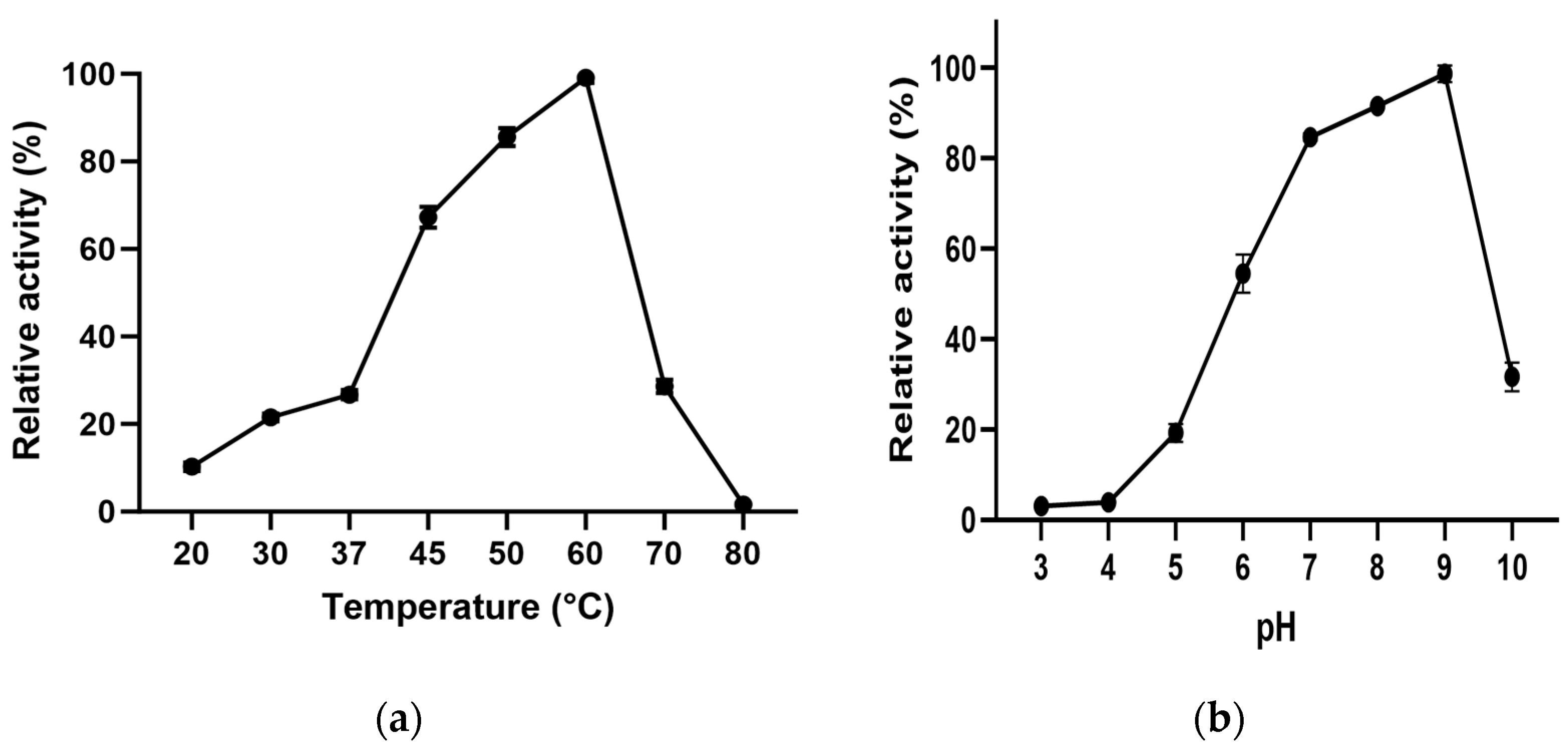
3.4. Effect of Temperature and pH
3.5. Molecular Weight Determination, SDS-PAGE, and Zymography
3.6. Effect of Temperature and pH
3.7. Effect of Metal Ions and Inhibitors
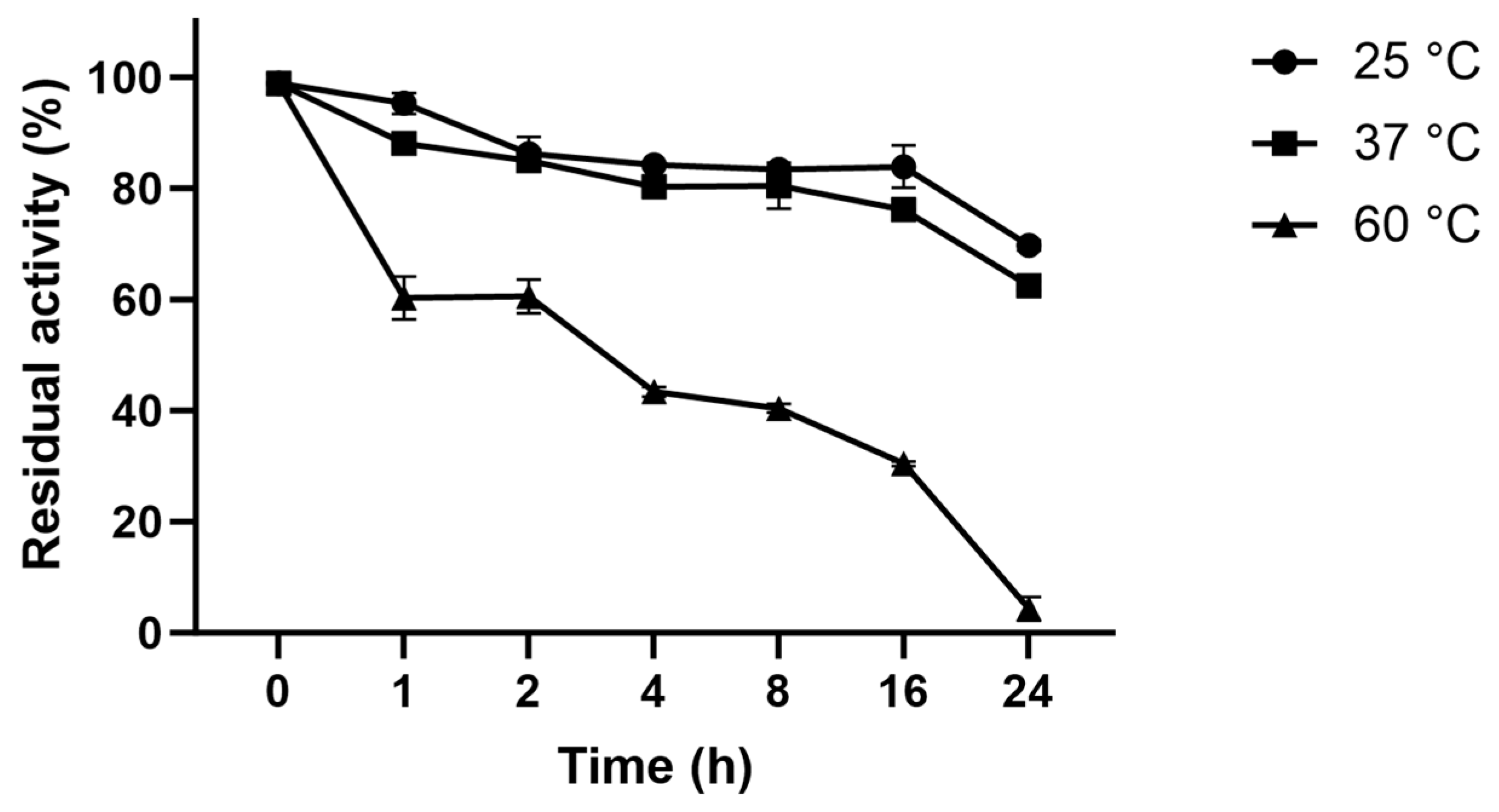
3.8. Thermostability of L-ASNasaZP21
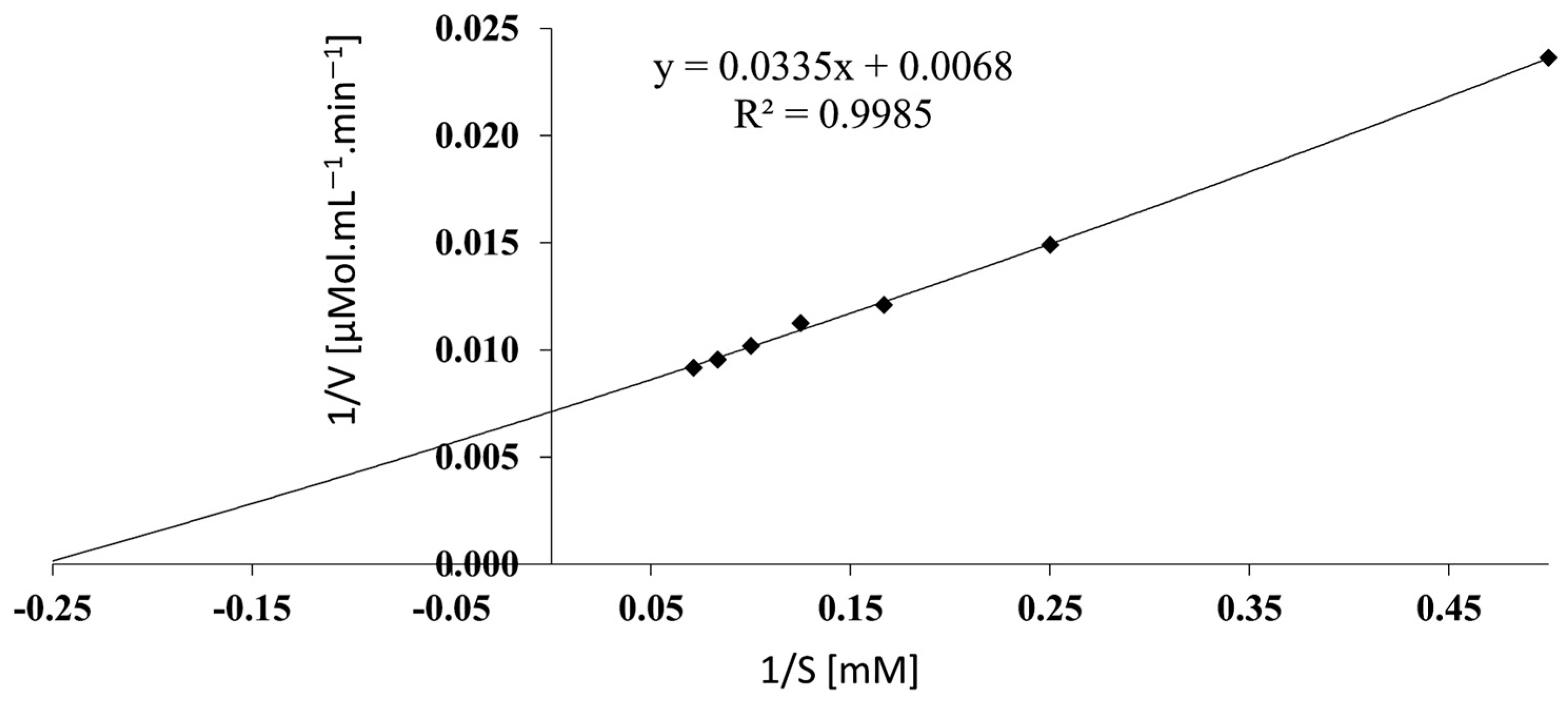
3.9. Determination of Kinetic Parameters
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shakambari, G.; Ashokkumar, B.; Varalakshmi, P. L-asparaginase—A promising biocatalyst for industrial and clinical applications. Biocatal. Agric. Biotechnol. 2019, 17, 213–224. [Google Scholar] [CrossRef]
- Mostafa, Y.; Alrumman, S.; Alamri, S.; Hashem, M.; Al-izran, K.; Alfaifi, M.; Elbehairi, S.E.; Taha, T. Enhanced production of glutaminase-free L-asparaginase by marine Bacillus velezensis and cytotoxic activity against breast cancer cell lines. Electron. J. Biotechnol. 2019, 42, 6–15. [Google Scholar] [CrossRef]
- Nunes, C.F.; Almeida, M.R.; De Paiva, G.B.; Pedrolli, D.B.; Neves, M.C.; Freire, M.G.; Tavares, A.P.M. A flow-through strategy using supported ionic liquids for L-asparaginase purification. Sep. Purif. Technol. 2023, 315, 123718. [Google Scholar] [CrossRef]
- Asselin, B.; Rizzari, C. Asparaginase pharmacokinetics and implications of therapeutic drug monitoring. Leuk. Lymphoma 2015, 56, 2273–2280. [Google Scholar] [CrossRef]
- Dhankhar, R.; Gupta, V.; Kumar, S.; Kapoor, R.K.; Gulati, P. Microbial enzymes for deprivation of amino acid metabolism in malignant cells: Biological strategy for cancer treatment. Appl. Microbiol. Biotechnol. 2020, 104, 2857–2869. [Google Scholar] [CrossRef]
- Cachumba, J.J.M.; Antunes, F.A.F.; Peres, G.F.D.; Brumano, L.P.; Dos Santos, J.C.; Da Silva, S.S. Current applications and different approaches for microbial L-asparaginase production. Braz. J. Microbiol. 2016, 47, 77–85. [Google Scholar] [CrossRef] [PubMed]
- El-Ghonemy, D.H. Medical microbiology & Diagnosis microbial amidases and their industrial applications: A review. Med. Microbiol. Diagn. 2014, 4, 4–9. [Google Scholar] [CrossRef]
- Yim, S.; Kim, M. Purification and characterization of thermostable L-asparaginase from Bacillus amyloliquefaciens MKSE in Korean soybean paste. LWT 2019, 109, 415–421. [Google Scholar] [CrossRef]
- Ran, T.; Jiao, L.; Wang, W.; Chen, J.; Chi, H.; Lu, Z.; Zhang, C.; Xu, D.; Lu, F. Structures of L-asparaginase from Bacillus licheniformis reveal an essential residue for its substrate stereoselectivity. J. Agric. Food Chem. 2021, 69, 223–231. [Google Scholar] [CrossRef]
- Chakravarty, N.; Priyanka; Singh, J.; Singh, R.P. A potential type-II L-asparaginase from marine isolate Bacillus australimaris NJB19: Statistical optimization, in silico analysis and structural modeling. Int. J. Biol. Macromol. 2021, 174, 527–539. [Google Scholar] [CrossRef]
- Costa-Silva, T.A.; Costa, I.M.; Biasoto, H.P.; Lima, G.M.; Silva, C.; Pessoa, A.; Monteiro, G. Critical overview of the main features and techniques used for the evaluation of the clinical applicability of L-asparaginase as a biopharmaceutical to treat blood cancer. Blood Rev. 2020, 43, 100651. [Google Scholar] [CrossRef] [PubMed]
- Zolfaghar, M.; Amoozegar, M.A.; Khajeh, K.; Babavalian, H.; Tebyanian, H. Isolation and screening of extracellular anticancer enzymes from halophilic and halotolerant bacteria from different saline environments in Iran. Mol. Biol. Rep. 2019, 46, 3275–3286. [Google Scholar] [CrossRef] [PubMed]
- Calderón-toledo, S.; Tapia-Bañez, Y.; Jiménez-Aliaga, K.; Esquerre-Hullpa, C.; Zavaleta, A.I. Caracterización bioinformática y producción de L-asparaginasa de Bacillus sp. M62 aislado de las salinas de Maras, Cusco, Perú. Rev. Peru. Biol. 2023, 30, e22411. [Google Scholar] [CrossRef]
- Gholamian, S.; Gholamian, S.; Nazemi, A.; Nargesi, M. Isolation and characterization of a novel Bacillus sp. strain that produces L-asparaginase, an antileukemic drug. Asian J. Biol. 2013, 6, 106–115. [Google Scholar] [CrossRef]
- Onishi, Y.; Yano, S.; Thongsanit, J.; Takagi, K.; Yoshimune, K.; Wakayama, M. Expression in Escherichia coli of a gene encoding type II L-asparaginase from Bacillus subtilis, and characterization of its unique properties. Ann. Microbiol. 2011, 61, 517–524. [Google Scholar] [CrossRef]
- Fisher, S.H.; Wray, L.V. Bacillus subtilis 168 contains two differentially regulated genes encoding L-asparaginase. J. Bacteriol. 2002, 184, 2148–2154. [Google Scholar] [CrossRef]
- Thenmozhi, C.; Sankar, R.; Karuppiah, V.; Sampathkumar, P. L-Asparaginase production by mangrove derived Bacillus cereus MAB5: Optimization by response surface methodology. Asian Pac. J. Trop. Med. 2011, 4, 486–491. [Google Scholar] [CrossRef]
- Feng, Y.; Liu, S.; Jiao, Y.; Gao, H.; Wang, M.; Du, G.; Chen, J. Enhanced extracellular production of L-asparaginase from Bacillus subtilis 168 by B. subtilis WB600 through a combined strategy. Appl. Microbiol. Biotechnol. 2017, 101, 1509–1520. [Google Scholar] [CrossRef]
- Ismail, N.F.; Hamdan, S.; Mahadi, N.M.; Murad, A.M.A.; Rabu, A.; Bakar, F.D.A.; Klappa, P.; Illias, R.M. A mutant L-asparaginase II signal peptide improves the secretion of recombinant cyclodextrin glucanotransferase and the viability of Escherichia coli. Biotechnol. Lett. 2011, 33, 999–1005. [Google Scholar] [CrossRef]
- De Moura, W.A.F.; Schultz, L.; Breyer, C.A.; de Oliveira, A.L.P.; Tairum, C.A.; Fernandes, G.C.; Toyama, M.H.; Pessoa-Jr, A.; Monteiro, G.; de Oliveira, M.A. Functional and structural evaluation of the antileukaemic enzyme L-asparaginase II expressed at low temperature by different Escherichia coli strains. Biotechnol. Lett. 2020, 42, 2333–2344. [Google Scholar] [CrossRef]
- Singh, Y.; Gundampati, R.K. Extracellular L-asparaginase from a protease-deficient Bacillus aryabhattai ITBHU02: Purification, biochemical characterization, and evaluation of antineoplastic activity in vitro. Appl. Biochem. Biotechnol. 2013, 171, 1759–1774. [Google Scholar] [CrossRef]
- Aly, N.; El-Ahwany, A.; Ataya, F.S.; Saeed, H. Bacillus sonorensis L. asparaginase: Cloning, expression in E. coli and characterization. Protein J. 2020, 39, 717–729. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xie, Y.; Zhang, C.; Bie, X.; Zhao, H.; Lu, F.; Lu, Z. Biochemical characterization of a novel L-asparaginase from Bacillus megaterium H-1 and its application in french fries. FRIN 2015, 77, 527–533. [Google Scholar] [CrossRef]
- Pillaca-Pullo, O.S. Biodiversidad microbiana en la producción del biofármaco L-asparaginasa: Una revisión sobre su potencial terapéutico. Rev. Investig. Univ. Norbert Wien. 2022, 11, r0005. [Google Scholar] [CrossRef]
- Montes Cjuno, J.Z. Caracterización Molecular de Bacterias Con Actividad L-Asparaginasa Aisladas de Las Salinas de Pilluana, Maras y Chilca. Bachelor Thesis, Pharmaceutical Chemist, Universidad Nacional Mayor de San Marcos, Lima, Peru, 2018. [Google Scholar]
- Mahajan, R.V.; Kumar, V.; Rajendran, V.; Saran, S.; Ghosh, P.C.; Saxena, R.K. Purification and characterization of a novel and robust L-asparaginase having low-glutaminase activity from Bacillus licheniformis: In vitro evaluation of anti-cancerous properties. PLoS ONE 2014, 9, e99037. [Google Scholar] [CrossRef]
- Hurtado, A.; Flores-Santos, J.C.; Flores-Fernández, C.N.; Saavedra, S.; Santos, J.H.P.M.; Pessoa-Júnior, A.; Lienqueo, M.E.; Bayro, M.J.; Zavaleta, A.I. A novel L-asparaginase from Enterobacter sp. strain M55 from Maras salterns in Peru. Chem. Biochem. Eng. Q. 2022, 36, 167–173. [Google Scholar] [CrossRef]
- Shifrin, S.; Parrott, C.L.; Luborsky, S.W. Substrate binding and intersubunit interactions in L-asparaginase. J. Biol. Chem. 1974, 249, 1335–1340. [Google Scholar] [CrossRef]
- Rahimzadeh, M.; Poodat, M.; Javadpour, S.; Qeshmi, F.I.; Shamsipour, F. Purification, characterization and comparison between two new L-asparaginases from PG03 and PG04. Open Biochem. J. 2016, 10, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Sant’Anna, V.; Cladera-Olivera, F.; Brandelli, A. Kinetic and thermodynamic study of thermal inactivation of the antimicrobial peptide P34 in milk. Food Chem. 2012, 130, 84–89. [Google Scholar] [CrossRef]
- Tjalsma, H.; Bolhuis, A.; Jongbloed, J.D.H.; Bron, S.; van Dijl, J.M. Signal peptide-dependent protein transport in Bacillus subtilis: A genome-based survey of the secretome. Microbiol. Mol. Biol. Rev. 2000, 64, 515–547. [Google Scholar] [CrossRef]
- Bengtsson, J.; Tjalsma, H.; Rivolta, C.; Hederstedt, L. Subunit II of Bacillus subtilis Cytochrome C oxidase is a lipoprotein. J. Bacteriol. 1999, 181, 685–688. [Google Scholar] [CrossRef]
- Nesmeyanova, M.A.; Karamyshev, A.L.; Karamysheva, Z.N.; Kalinin, A.E.; Ksenzenko, V.N.; Kajava, A.V. Positively charged lysine at the N-terminus of the signal peptide of the Escherichia coli alkaline phosphatase provides the secretion efficiency and is involved in the interaction with anionic phospholipids. FEBS Lett. 1997, 403, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold protein structure database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Liu, S.; Jiao, Y.; Wang, Y.; Wang, M.; Du, G. Gene cloning and expression of the L-asparaginase from Bacillus cereus BDRD-ST26 in Bacillus subtilis WB600. J. Biosci. Bioeng. 2019, 127, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Xu, M.; He, B.; Rao, Z. Cloning, expression, and characterization of L-asparaginase from a newly isolated Bacillus subtilis B11-06. J. Agric. Food Chem. 2013, 61, 9428–9434. [Google Scholar] [CrossRef]
- Falak, S.; Sajed, M.; Rashid, N. Strategies to enhance soluble production of heterologous proteins in Escherichia coli. Biologia 2022, 77, 893–905. [Google Scholar] [CrossRef]
- Rodriguez, E.L.; Poddar, S.; Iftekhar, S.; Suh, K.; Woolfork, A.G.; Ovbude, S.; Pekarek, A.; Walters, M.; Lott, S.; Hage, D.S. Affinity chromatography: A review of trends and developments over the past 50 years. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2020, 1157, 122332. [Google Scholar] [CrossRef]
- Lu, X.; Chen, J.; Jiao, L.; Zhong, L.; Lu, Z.; Zhang, C.; Lu, F. Improvement of the activity of L-asparaginase I improvement of the catalytic activity of L-asparaginase I from Bacillus megaterium H-1 by in vitro directed evolution. J. Biosci. Bioeng. 2019, 128, 683–689. [Google Scholar] [CrossRef]
- Sanghvi, G.; Bhimani, K.; Vaishnav, D.; Oza, T.; Dave, G.; Kunjadia, P. Mitigation of acrylamide by L-asparaginase from Bacillus subtilis KDPS1 and analysis of degradation products by HPLC and HPTLC. Springerplus 2016, 5, 1–11. [Google Scholar] [CrossRef]
- Chand, S.; Mahajan, R.; Prasad, J.P.; Sahoo, D.K.; Mihooliya, K.N.; Dhar, M.S.; Sharma, G. A comprehensive review on microbial L-asparaginase: Bioprocessing, characterization, and industrial applications. Biotechnol. Appl. Biochem. 2020, 67, 619–647. [Google Scholar] [CrossRef] [PubMed]
- Lubkowski, J.; Wlodawer, A. Structural and biochemical properties of L-asparaginase. FEBS J. 2021, 288, 4183–4209. [Google Scholar] [CrossRef]
- Lakshmi, A.V.; Mari, D.S. Screening and identification of asparaginase and glutaminase producing halophilic bacteria from natural saline habitats. Int. J. Recent Adv. Biotechnol. Nanotechnol. 2020, 3, 34–53. [Google Scholar]
- Krishnapura, P.R.; Belur, P.D.; Subramanya, S. A critical review on properties and applications of microbial L-asparaginases. Crit. Rev. Microbiol. 2016, 42, 720–737. [Google Scholar] [CrossRef]
- Zuo, S.; Zhang, T.; Jiang, B.; Mu, W. Recent research progress on microbial L-asparaginases. Appl. Microbiol. Biotechnol. 2015, 99, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Venkata Dasu, V.; Pakshirajan, K. Purification and characterization of glutaminase-free L-asparaginase from Pectobacterium carotovorum MTCC 1428. Bioresour Technol 2011, 102, 2077–2082. [Google Scholar] [CrossRef] [PubMed]
- Warangkar, S.C.; Khobragade, C.N. Purification, characterization, and effect of thiol compounds on activity of the Erwinia carotovora L-asparaginase. Enzym. Res. 2010, 2010, 165878. [Google Scholar] [CrossRef] [PubMed]
- Moorthy, V.; Ramalingam, A.; Sumantha, A.; Shankaranaya, R. Production, purification and characterization of extracellular L-asparaginase from a soil isolate of Bacillus sp. Afr. J. Microbiol. Res. 2010, 4, 1862–1867. [Google Scholar]
- Beckett, A.; Gervais, D. What makes a good new therapeutic L-asparaginase? World J. Microbiol. Biotechnol. 2019, 35, 152. [Google Scholar] [CrossRef]
- Costa, I.M.; Schultz, L.; De Araujo Bianchi Pedra, B.; Leite, M.S.M.; Farsky, S.H.P.; De Oliveira, M.A.; Pessoa, A.; Monteiro, G. Recombinant L-asparaginase 1 from Saccharomyces cerevisiae: An allosteric enzyme with antineoplastic activity. Sci. Rep. 2016, 6, 36239. [Google Scholar] [CrossRef]
- Wlodarczyk, S.R.; Costa-Silva, T.A.; Pessoa, A., Jr.; Madeira, P.; Monteiro, G. Effect of osmolytes on the activity of anti-cancer enzyme L-asparaginase II from Erwinia chrysanthemi. Process Biochem. 2019, 81, 123–131. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Activity (U) | Total Protein (mg) | Activity (U mg−1) | Purification-Fold | Yield (%) | |
|---|---|---|---|---|---|
| Crude extract | 2.38 × 102 | 86.39 | 2.75 | 1.0 | 100.0 |
| Ni-affinity | 1.47 × 102 | 0.63 | 234.38 | 85.2 | 61.9 |
| Source | pH | Temperature (°C) | Activity (U mg−1) | Km (mM) | Chromatography 1 | References |
|---|---|---|---|---|---|---|
| Bacillus subtilis CH11 | 9.0 | 60 | 234.38 | 4.75 | AC | This study |
| B. subtilis BDRD-ST26 | - | 65 | 162.90 | 5.29 | HIC, IEX, GC | Feng et al. [36] |
| B. subtilis 168 | 8.0 7.5 | 65 50 | 45.40 31.90 | 2.06 7.06 | IEX, HIC | Onishi et al. [15] |
| B. subtilis B11-06 | 7.5 | 40 | 92.45 | 0.43 | HIC | Jia et al. [37] |
| B. subtilis KDPS1 | 5.0 | 37 | - | - | IEX | Sanghvi et al. [41] |
| B. aryabhattai ITBHU02 | 8.5 | 40 | 680.50 | 0.25 | HIC, GF | Singh et al. [21] |
| B. amyloliquefaciens MKSE | 8.5 | 65 | 136.30 | 1.15 | AC | Yim et al. [8] |
| B. cereus | 9.0 | 50 | 550.80 | 9.38 | HIC, IEX, GF | Feng et al. [36] |
| B. megaterium H-1 | 8.0 | 40 | 1146.29 | 21.63 | AC | Lu et al. [40] |
| B. sonorensis | 7.0 | 45 | 4438.62 | 2.00 | AC | Aly et al. [22] |
| B. velenzensis | 7.5 | 37 | 31.77 | 0.04 | GF | Mostafa et al. [2] |
| Ions/Inhibitors | Final Concentration | Relative Activity (%) 1 |
|---|---|---|
| Control | - | 100.00 |
| KCl | 100 mM | 124.500 ± 1.85 * |
| NaCl | 106.300 ± 0.09 | |
| MgCl2 | 149.800 ± 4.04 * | |
| CaCl2 | 310.700 ± 3.28 * | |
| BaCl2 | 95.070 ± 2.73 | |
| MnCl2 | 0.0 * | |
| CuCl2 | 0.0 * | |
| CoCl2 | 0.0 * | |
| PMFS | 10 mM | 118.700 ± 5.77 |
| Urea | 96.260 ± 7.36 | |
| Mercaptoethanol | 139.800 ± 3.52 * | |
| DL-dithiothreitol | 271.100 ± 37.00 * | |
| SDS | 0.0 * | |
| EDTA | 58.850 ± 1.46 * | |
| Glutathione | 5 mM | 97.860 ± 4.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arredondo-Nuñez, A.; Monteiro, G.; Flores-Fernández, C.N.; Antenucci, L.; Permi, P.; Zavaleta, A.I. Characterization of a Type II L-Asparaginase from the Halotolerant Bacillus subtilis CH11. Life 2023, 13, 2145. https://doi.org/10.3390/life13112145
Arredondo-Nuñez A, Monteiro G, Flores-Fernández CN, Antenucci L, Permi P, Zavaleta AI. Characterization of a Type II L-Asparaginase from the Halotolerant Bacillus subtilis CH11. Life. 2023; 13(11):2145. https://doi.org/10.3390/life13112145
Chicago/Turabian StyleArredondo-Nuñez, Annsy, Gisele Monteiro, Carol N. Flores-Fernández, Lina Antenucci, Perttu Permi, and Amparo Iris Zavaleta. 2023. "Characterization of a Type II L-Asparaginase from the Halotolerant Bacillus subtilis CH11" Life 13, no. 11: 2145. https://doi.org/10.3390/life13112145