
Population Structure and Genetic Diversity Analysis of “Yufen 1” H Line Chickens Using Whole-Genome Resequencing

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and WGS Analysis
2.2. Read Mapping and SNP Calling
2.3. Genomic Diversity Analysis
2.4. Population Genetic Structure and Gene Flow Analysis
3. Results
3.1. SNP Calling and Annotation
3.2. Genetic Diversity and LD Analysis
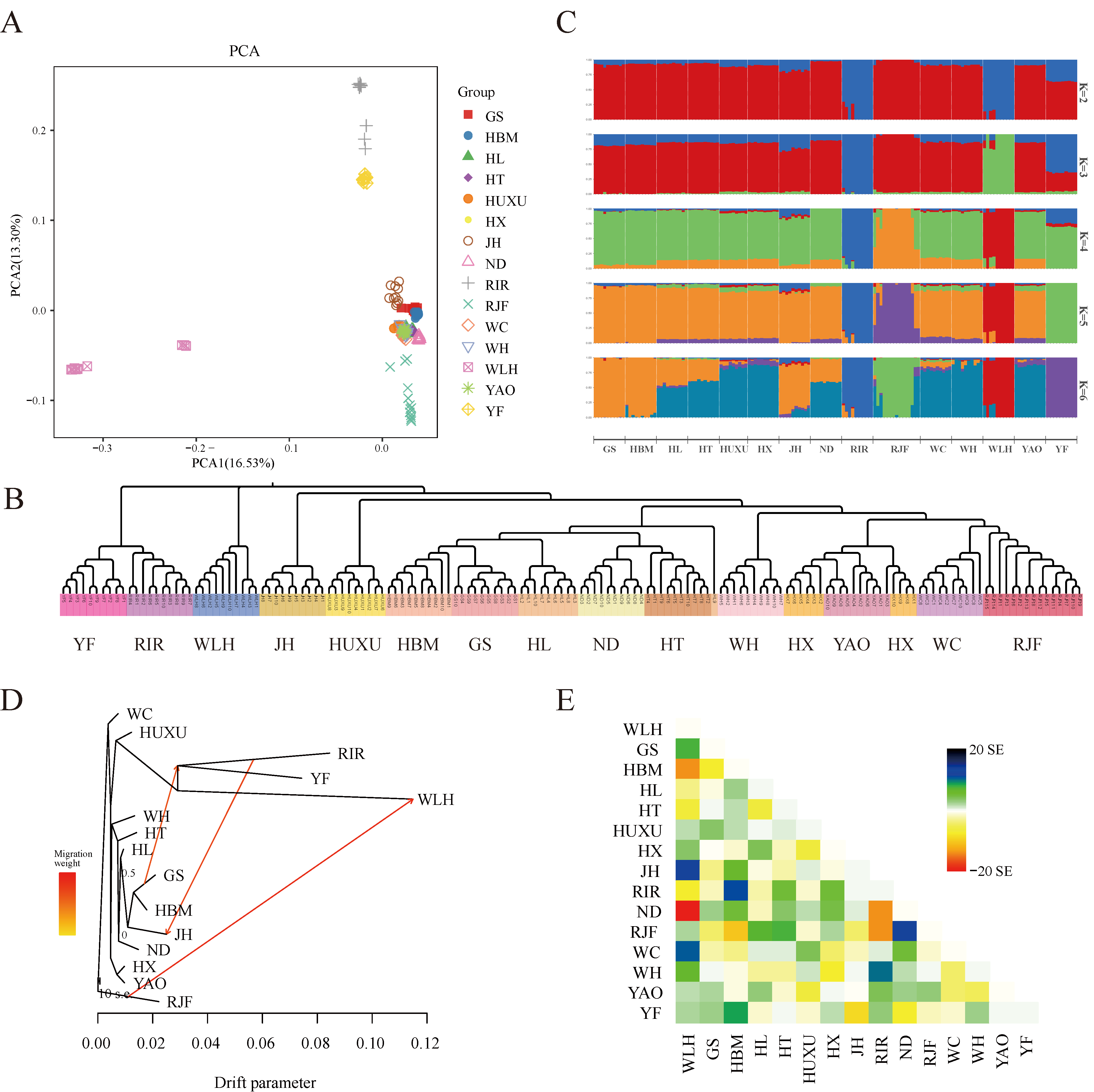
3.3. Population Genetic Structure and Gene Flow Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Luo, W.; Luo, C.; Wang, M.; Guo, L.; Nie, Q. Genome diversity of Chinese indigenous chicken and the selective signatures in Chinese gamecock chicken. Sci. Rep. 2020, 10, 14532. [Google Scholar] [CrossRef] [PubMed]
- Changsheng, N.; Pedro, A.; Yaxiong, J.; Haigang, B.; Zhonghua, N.; Lujiang, Q. Genome-Wide Single-Nucleotide Polymorphism Data Unveil Admixture of Chinese Indigenous Chicken Breeds with Commercial Breeds. Genome Biol. Evol. 2019, 11, 1847–1856. [Google Scholar]
- Zhang, C.; Lin, D.; Wang, Y.; Peng, D.; Li, H.; Fei, J.; Chen, K.; Yang, N.; Hu, X.; Zhao, Y.; et al. Widespread introgression in Chinese indigenous chicken breeds from commercial broiler. Evol. Appl. 2019, 12, 610–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sha, Y.; Gao, C.; Liu, M.; Zhao, S. Evaluation of the genetic diversity of six Chinese indigenous chickens. Asian-Australas. J. Anim. Sci. 2020, 33, 1566–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherf, B.D.; Fao, R.; Pilling, D. The Second Report on the State of the World’s Animal Genetic Resources for Food and Agriculture; Food and Agriculture Organization: Rome, Italy, 2015. [Google Scholar]
- Cheng, J.; Zhao, H.; Chen, N.; Cao, X.; Chen, H. Population structure, genetic diversity, and selective signature of Chaka sheep revealed by whole genome sequencing. BMC Genom. 2020, 21, 520. [Google Scholar] [CrossRef]
- Li, D.; Sun, G.; Zhang, M.; Cao, Y.; Kang, X. Breeding history and candidate genes responsible for black skin of Xichuan black-bone chicken. BMC Genom. 2020, 21, 511. [Google Scholar] [CrossRef] [PubMed]
- David, D.; Hubert, A.S.; Kossigan, D.G.; Chrysostome, C.C.A.A.M.; Olorounto, K.D. Genetic diversity and phylogenetic relationships within local pigs in southern Benin. Trop. Anim. Health Prod. 2021, 53, 434. [Google Scholar]
- Yang, Z.; Zou, L.; Sun, T.; Xu, W.; Zeng, L.; Jia, Y.; Jiang, J.; Deng, J.; Yang, X. Genome-Wide Association Study Using Whole-Genome Sequencing Identifies a Genomic Region on Chromosome 6 Associated With Comb Traits in Nandan-Yao Chicken. Front. Genet. 2021, 12, 682501. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Chen, T.; Zhu, M. Whole-genome sequencing revealed genetic diversity and selection of Guangxi indigenous chickens. PLoS ONE 2022, 17, e0250392. [Google Scholar] [CrossRef]
- Chen, L.; Wang, X.; Cheng, D.; Chen, K.; Fan, Y.; Wu, G.; You, J.; Liu, S.; Mao, H.; Ren, J. Population genetic analyses of seven Chinese indigenous chicken breeds in a context of global breeds. Anim. Genet. 2019, 50, 82–86. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, D.; Song, S.; Zhang, Y.; Kang, X. Combined transcriptomics and proteomics forecast analysis for potential genes regulating the Columbian plumage color in chickens. PLoS ONE 2019, 14, e0210850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Hu, H.; Tian, Y.; Li, J.; Kang, X. The chicken pan-genome reveals gene content variation and a promoter region deletion in IGF2BP1 affecting body size. Molecular Biology and Evolution. Mol. Biol. Evol. 2021, 38, 5066–5081. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Otecko, N.O.; Peng, M.; Weng, Z.; Du, B. Genome-wide genetic structure and selection signatures for color in 10 traditional Chinese yellow-feathered chicken breeds. BMC Genom. 2020, 21, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qanbari, S.; Rubin, C.J.; Maqbool, K.; Weigend, S.; Weigend, A.; Geibel, J.; Kerje, S.; Wurmser, C.; Peterson, A.T.; Brisbin, I.L., Jr.; et al. Genetics of adaptation in modern chicken. PLoS Genet. 2019, 15, e1007989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heng, L. Toward better understanding of artifacts in variant calling from high-coverage samples. Bioinformatics 2014, 30, 2843–2851. [Google Scholar]
- Heng, L.; Richard, D. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar]
- Lin, Y.L.; Chang, P.C.; Hsu, C.; Hung, M.Z.; Chien, Y.H.; Hwu, W.L.; Lai, F.; Lee, N.C. Comparison of GATK and DeepVariant by trio sequencing. Sci. Rep. 2022, 12, 1809. [Google Scholar] [CrossRef] [PubMed]
- Bayer, P.E.; Gill, M.; Danilevicz, M.F.; Edwards, D. Producing High-Quality Single Nucleotide Polymorphism Data for Genome-Wide Association Studies. Methods Mol. Biol. 2022, 2481, 153–159. [Google Scholar] [PubMed]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X. Ruden DM: A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Mcquillan, R.; Leutenegger, A.L.; Abdelrahman, R.; Franklin, C.S.; Pericic, M.; Baraclauc, L.; Smolejnarancic, N.; Janicijevic, B.; Polasek, O.; Tenesa, A. Runs of homozygosity in European populations. Am. J. Hum. Genet. 2008, 83, 359–372. [Google Scholar] [CrossRef] [Green Version]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; Depristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]
- Santiago, E.; Novo, I.; Pardiñas, A.F.; Saura, M.; Wang, J.; Caballero, A. Recent demographic history inferred by high-resolution analysis of linkage disequilibrium. Mol. Biol. Evol. 2020, 37, 3642–3653. [Google Scholar] [CrossRef] [PubMed]
- Groenen, M.A.; Cheng, H.H.; Bumstead, N.; Benkel, B.F.; Briles, W.E.; Burke, T.; Burt, D.W.; Crittenden, L.B.; Dodgson, J.; Hillel, J. A consensus linkage map of the chicken genome. Genome Res. 2000, 10, 137–147. [Google Scholar] [PubMed]
- Warren, W.C.; Hillier, L.W.; Tomlinson, C.; Minx, P.; Kremitzki, M.; Graves, T.; Markovic, C.; Bouk, N.; Pruitt, K.D.; Thibaud-Nissen, F. A new chicken genome assembly provides insight into avian genome structure. G3 Genes Genomes Genet. 2017, 7, 109–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Dong, S.S.; Xu, J.Y.; He, W.M.; Yang, T.L. PopLDdecay: A fast and effective tool for linkage disequilibrium decay analysis based on variant call format files. Bioinformatics 2019, 35, 1786–1788. [Google Scholar] [CrossRef]
- Felsenstein, J. PHYLIP-Phylogeny Inference Package (Version 3.2). Cladistics 1989, 5, 164–166. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Alexander, D.H.; Lange, K. Enhancements to the ADMIXTURE algorithm for individual ancestry estimation. BMC Bioinform. 2011, 12, 246. [Google Scholar] [CrossRef] [Green Version]
- Pickrell, J.K.; Pritchard, J.K.; Tang, H. Inference of population splits and mixtures from genome-wide allele frequency data. PLoS Genet. 2012, 8, e1002967. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Dong, R.; Li, X.; Cui, C.; Yu, G. Analysis of the Genetic Diversity and Family Structure of the Licha Black Pig Population on Jiaodong Peninsula, Shandong Province, China. Animals 2022, 12, 1045. [Google Scholar] [CrossRef]
- Strillacci, M.G.; Cozzi, M.C.; Gorla, E.; Mosca, F.; Schiavini, F.; Román-Ponce, S.; López, F.R.; Schiavone, A.; Marzoni, M.; Cerolini, S. Genomic and genetic variability of six chicken populations using single nucleotide polymorphism and copy number variants as markers. Animal 2017, 11, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Cendron, F.; Perini, F.; Mastrangelo, S.; Tolone, M.; Cassandro, M. Genome-Wide SNP Analysis Reveals the Population Structure and the Conservation Status of 23 Italian Chicken Breeds. Animals 2020, 10, 1441. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Nie, C.; Li, X.; Ning, Z.; Qu, L. Genome-Wide Population Genetic Analysis of Commercial, Indigenous, Game, and Wild Chickens Using 600K SNP Microarray Data. Front. Genet. 2020, 11, 543294. [Google Scholar] [CrossRef] [PubMed]
- Setianto, J.; Zain, B.; Sutriyono; Prakoso, H. Domestication of red jungle fowl: A case study of the red jungle fowl chicks procurement by the communities in Central Bengkulu, Indonesia. Biodiversitas 2017, 18, 183–189. [Google Scholar] [CrossRef]
- Akrim, F.; Awan, M.S.; Mahmood, T.; Anjum, M.Z.; Qasim, S.; Khalid, J.; Andleeb, S. Threats to Red Junglefowl (Gallus gallus murghi) in Deva Vatala National Park, District Bhimber, Azad Jammu and Kashmir, Pakistan. Sciencedomain 2015, 6, 59–65. [Google Scholar] [CrossRef]
- Meuwissen, T.; Woolliams, J.A. Effective sizes of livestock populations to prevent a decline in fitness. Theor. Appl. Genet. 1994, 89, 1019–1026. [Google Scholar] [CrossRef] [PubMed]
- Zanetti, E.; Marchi, M.D.; Dalvit, C.; Cassandro, M. Genetic characterization of local Italian breeds of chickens undergoing in situ conservation. Poult. Sci. 2010, 89, 420–427. [Google Scholar] [CrossRef]
- Mengmeng, Z.; Wei, H.; Hui, T.; Guohui, L.; Minjie, Z.; Ran, X.; Yijun, L.; Tao, Y.; Wenting, L.; Jianmin, Z. Genomic diversity dynamics in conserved chicken populations are revealed by genome-wide SNPs. BMC Genom. 2018, 19, 598. [Google Scholar]
- Ballou, J.D.; Lacy, R.C.; Traylor-Holzer, K.; Bauman, K.; Ivy, J.A.; Asa, C. Strategies for establishing and using genome resource banks to protect genetic diversity in conservation breeding programs. Zoo Biol. 2022. Online ahead of print. [Google Scholar] [CrossRef]


{kind=link}
| Breeds | N 1 | Breed Assignment | Ho 2 | He 3 | FROH 4 | Π 5 | Ne 6 | MAF 7 |
|---|---|---|---|---|---|---|---|---|
| WLH 8 | 10 | Commercial breeds (Italy) | 0.1332 | 0.1757 | 0.4427 | 0.0016 | 11 | 0.1279 |
| RIR 9 | 10 | Commercial breeds (America) | 0.1915 | 0.2128 | 0.3096 | 0.0019 | 59 | 0.1582 |
| GS 10 | 10 | Native breeds (Henan province, China) | 0.3129 | 0.3206 | 0.0793 | 0.0028 | 120 | 0.2372 |
| HBM 11 | 10 | Native breeds (Anhui province, China) | 0.2992 | 0.3056 | 0.0469 | 0.0027 | 167 | 0.2273 |
| HL 12 | 10 | Native breeds (Hunan province, China) | 0.3042 | 0.3174 | 0.0284 | 0.0028 | 497 | 0.2353 |
| HT 13 | 10 | Native breeds (Fujian province, China) | 0.3019 | 0.3095 | 0.0472 | 0.0028 | 306 | 0.2301 |
| HUXU 14 | 9 | Native breeds (Guangdong province, China) | 0.2794 | 0.3072 | 0.0424 | 0.0027 | 183 | 0.2283 |
| HX 15 | 10 | Native breeds (Guangdong province, China) | 0.2991 | 0.3133 | 0.0405 | 0.0028 | 262 | 0.2325 |
| JH 16 | 10 | Native breeds (Hubei province, China) | 0.2817 | 0.3064 | 0.0784 | 0.0027 | 52 | 0.2278 |
| ND 17 | 10 | Native breeds (Jiangxi province, China) | 0.2891 | 0.3055 | 0.0540 | 0.0027 | 262 | 0.2269 |
| WC 18 | 10 | Native breeds (Hainan province, China) | 0.2936 | 0.3134 | 0.0374 | 0.0028 | 216 | 0.2324 |
| WH 19 | 10 | Native breeds (Anhui province, China) | 0.3033 | 0.3078 | 0.0352 | 0.0027 | 52 | 0.2288 |
| YAO 20 | 10 | Native breeds (Guangxi province, China) | 0.3026 | 0.3136 | 0.0422 | 0.0028 | 264 | 0.2328 |
| RJF 21 | 15 | Wild breeds (Thailand) | 0.2724 | 0.2903 | 0.1696 | 0.0026 | 43 | 0.2155 |
| YF 22 | 10 | Native breeds (Henan province, China) | 0.2766 | 0.2630 | 0.3273 | 0.0024 | 70 | 0.1932 |
| Breeds | WLH 1 | RIR 2 | GS 3 | HBM 4 | HL 5 | HT 6 | HUXU 7 | HX 8 | JH 9 | ND 10 | WC 11 | WH 12 | YAO 13 | RJF 14 | YF 15 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WLH 1 | 0 | 0.3006 | 0.2232 | 0.2239 | 0.2039 | 0.2123 | 0.2043 | 0.2010 | 0.2079 | 0.2164 | 0.1989 | 0.2092 | 0.2020 | 0.2127 | 0.2636 |
| RIR 2 | 0 | 0.1648 | 0.1648 | 0.1580 | 0.1650 | 0.1619 | 0.1584 | 0.1480 | 0.1712 | 0.1593 | 0.1654 | 0.1589 | 0.1947 | 0.2066 | |
| GS 3 | 0 | 0.0278 | 0.0272 | 0.0429 | 0.0460 | 0.0437 | 0.0405 | 0.0424 | 0.0428 | 0.0492 | 0.0437 | 0.0947 | 0.1404 | ||
| HBM 4 | 0 | 0.0193 | 0.0365 | 0.0405 | 0.0386 | 0.0333 | 0.0350 | 0.0371 | 0.0438 | 0.0390 | 0.0921 | 0.1411 | |||
| HL 5 | 0 | 0.0166 | 0.0193 | 0.0149 | 0.0262 | 0.0151 | 0.0135 | 0.0240 | 0.0163 | 0.0683 | 0.1347 | ||||
| HT 6 | 0 | 0.0307 | 0.0278 | 0.0417 | 0.0254 | 0.0244 | 0.0355 | 0.0288 | 0.0761 | 0.1427 | |||||
| HUXU 7 | 0 | 0.0205 | 0.0398 | 0.0320 | 0.0170 | 0.0293 | 0.0221 | 0.0746 | 0.1372 | ||||||
| HX 8 | 0 | 0.0390 | 0.0274 | 0.0166 | 0.0272 | 0.0056 | 0.0696 | 0.1368 | |||||||
| JH 9 | 0 | 0.0410 | 0.0375 | 0.0452 | 0.0393 | 0.0903 | 0.1339 | ||||||||
| ND 10 | 0 | 0.0253 | 0.0349 | 0.0291 | 0.0757 | 0.1472 | |||||||||
| WC 11 | 0 | 0.0249 | 0.0179 | 0.0650 | 0.1378 | ||||||||||
| WH 12 | 0 | 0.0280 | 0.0781 | 0.1432 | |||||||||||
| YAO 13 | 0 | 0.0706 | 0.1372 | ||||||||||||
| RJF 14 | 0 | 0.1780 | |||||||||||||
| YF 15 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Wang, D.; He, Y.; Liang, W.; Li, W.; Wang, K.; Li, D.; Li, Z.; Tian, Y.; Kang, X.; et al. Population Structure and Genetic Diversity Analysis of “Yufen 1” H Line Chickens Using Whole-Genome Resequencing. Life 2023, 13, 793. https://doi.org/10.3390/life13030793
Liu C, Wang D, He Y, Liang W, Li W, Wang K, Li D, Li Z, Tian Y, Kang X, et al. Population Structure and Genetic Diversity Analysis of “Yufen 1” H Line Chickens Using Whole-Genome Resequencing. Life. 2023; 13(3):793. https://doi.org/10.3390/life13030793
Chicago/Turabian StyleLiu, Cong, Dongxue Wang, Yuehua He, Wenjie Liang, Wenting Li, Kejun Wang, Donghua Li, Zhuanjian Li, Yadong Tian, Xiangtao Kang, and et al. 2023. "Population Structure and Genetic Diversity Analysis of “Yufen 1” H Line Chickens Using Whole-Genome Resequencing" Life 13, no. 3: 793. https://doi.org/10.3390/life13030793