
Molecular and Biochemical Mechanisms of Elicitors in Pest Resistance
by
 , and
, and
Saif ul Malook
1,2,*,† ,
,
Saiqa Maqbool
1,3,†,
Muhammad Hafeez
4,
Samantha Chandranath Karunarathna
5 and
and
Nakarin Suwannarach
1,*
1
Research Center of Microbial Diversity and Sustainable Utilization, Chiang Mai University, Chiang Mai 50200, Thailand
2
Shenzhen Branch, Guangdong Laboratory for Lingnan Modern Agriculture, Genome Analysis Laboratory of the Ministry of Agriculture, Agricultural Genomics Institute Shenzhen, Chinese Academy of Agricultural Sciences, Shenzhen 518100, China
3
Department of Chemistry, University of Agriculture Faisalabad, Sub-Campus Burewala, Faisalabad 38000, Pakistan
4
State Key Laboratory of Rice Biology, Ministry of Agriculture, Key Laboratory of Molecular Biology of Crop Pathogens and Insects, Institute of Insect Sciences, Zhejiang University, Hangzhou 310058, China
5
Center for Yunnan Plateau Biological Resources Protection and Utilization, College of Biological Resource and Food Engineering, Qujing Normal University, Qujing 655011, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Life 2022, 12(6), 844; https://doi.org/10.3390/life12060844
Submission received: 3 May 2022
/
Revised: 30 May 2022
/
Accepted: 3 June 2022
/
Published: 6 June 2022
(This article belongs to the Special Issue Advances in Plant Defensive Biochemical Activity)
Abstract
:Insect herbivores have a variety of life cycles and feeding habits, making them extremely diverse. With their host plants, they form close relationships and suppress their defense mechanisms. Molecular elicitors are the key bio-elements in the detection and recognition of attacking enemies in tissue consumption. Insect oral secretion, frass, and fluid of egg deposition contain biologically active molecules called herbivore-associated elicitors (HAEs) that are recognized by pattern-recognition receptors (PRRs). Many plants distinguish insect feeding from wounding by HAEs present in their oral secretions (OS) and induce local and/or systemic responses against arthropod feeding. PRRs perceive HAEs in the oral secretion of caterpillars in a species-specific manner to elicit exclusive defense responses. HAEs-PRRs interactions induce plant resistance by reprogramming plant metabolism and transcriptional machinery. Quantitative, timely, and coordinated plant response initiate early signaling events, including Ca2+, reactive oxygen species (ROS), and mitogen-activated protein kinases (MAPKs). However, in insect herbivory, little is known about the molecular basis of signal transduction and regulation of plant resistance. We discuss here how early signaling cascades converge into the accumulation of phytohormones that regulate downstream special metabolites against herbivores. In this review, we propose a hypothetical model of PPRs-HAEs-mediated-induced responses in plants and discuss how PRRs-HAEs interactions elicit short- and long-term induced defenses in plants. The understanding of PRRs-HAEs interactions will help to explore the fundamental molecular mechanisms of host manipulation and may generate prospects to develop novel pest-resistance strategies.
1. Introduction
As sessile organisms, plants cannot escape from herbivore arthropods and are substantially challenged by insect herbivores. Over millions of years of coevolution with insects, plants have evolved exquisite defense mechanisms to fend off insect herbivory on plants [1]. The recognition of herbivore attacks requires the ability of plants to detect chemical cues (Herbivore-associated elicitors; HAEs) generated by insects during infestation, and these receptors are also called receptor kinases (RKs). Plants distinguish insect feeding from wounding by recognizing specific conserved molecules present in their oral secretions (OS; shown in Figure 1) [2,3,4,5]. In literature, based on plant-insect interactions, few reports have revealed that OS constituents depend on the insect feeding of host plants and their associated microbes [6].
During insect herbivory, deposition of OS on the wounds causes manipulation of plant responses against insect herbivores by changing plant metabolism and gene expression [4,7]. HAEs in nature are diverse in structure and exist in the form of enzymes [e.g., glucose oxidase (GOX) and β-glucosidase], lipids [fatty acid-amino acid conjugates (FACs) such as volicitin and caeliferins], cell wall fragments (e.g., pectin and oligogalacturonides), and plant peptides (e.g., inceptin: proteolytic fragments of the chloroplastic ATP synthase subunit), but none of them were found to affect the induced defenses of tomato [4,8,9,10]. The HAEs are not general elicitors in all plants, and plant responses to insect herbivores are restricted to plant-insect associations that depend upon the specific mode of feeding style of insects [7,11,12]. This specificity reflects the evolutionary history of both plants and insects living and surviving together in nature, and it is important to understand the mechanism of plant-elicitors interactions in an evolutionary context [4]. Herbivore-induced defenses are mediated by signaling molecules and are employed to maintain crop resilience during insect herbivory [12,13,14]. Thus, despite the need for a clear understanding of induced responses, plant receptor interactions with their HAEs remain an emerging research topic in plant-insect interaction.
Upon the recognition of insect elicitors, plants activate defense responses by triggering calcium ion influx (Ca2+), plasma membrane depolarization (Vm), mitogen-activated protein kinases (MAPKs), NADPH oxidase, production of reactive oxygen species (ROS), and activation of nitrogen species (NO) [15]. The molecular signaling cascades elicit the production of defense hormones, mainly jasmonic acid (JA), ethylene, and salicylic acid (SA) as well as transcription factors (TFs). Defense hormone, especially JA, is the central component to regulate downstream defense metabolites including but not limited to glucosinolates, benzoxazinoids (Bxs), cyanogenic glucosides, alkaloids, phenolics, and proteinase inhibitors in damaged and systemic leaves, as shown in Figure 1 [13]. Several excellent reviews have been published on the discussion of herbivore-induced plant responses [13,16,17,18]. Molecular breeding for pest resistance traits is of great importance in developing crops with enhanced insect resistance. To develop pesticide-free food for an increasing world population, a clear understanding of the underlying mechanism in the perception of crop attack by insect herbivores and how plant receptors are employed to induce plant defense responses against herbivory is needed. The prime objective of this review is to provide cutting-edge research updates about plant receptor interaction with HAEs and the perception of insect herbivory to induce signal transduction-mediated defense responses accordingly.
2. Plant Receptors Perceive Insect Herbivory
Despite the continuous battle for the survival between plants and insects, the evolution of plant defenses is the main weapon that has a decisive power to determine victory in favor of plants or insects. Molecular and chemical ecologists mainly focused on investigating HAEs in the last two decades, but information on HAEs interactions with PRRs is still at an emerging stage. The first step in the mechanism of HAEs recognition is the detection of HAEs in the oral secretion (OS) of herbivorous insects, which is mediated by membrane-bound receptors [19,20]. However, many receptors of HAEs were discovered, and we have listed the known receptors and their respective HAEs in Table 1. The weapons of plant perception are divided into receptor-like kinases (RLKs) or proteins with ecto- and cytosolic domains in the plasma membrane that are actively involved in ligand binding. Plasma membrane-localized, leucine-rich repeat receptor-like kinases (LRR-RLK) genes belong to the subfamily of receptor-like kinases (RLKs). The role of RLKs is largely known for the recognition of insect herbivory when OS elicitors bind with the plasma membrane in plants. For example, biological active tritiated volicitin [(3H)-L-volicitin) binds to the plasma membrane in maize, and this binding was increased by foliar application of methyl jasmonate (MeJA). Similarly, actual feeding by Spodoptera exigua enhanced four-folds binding capacity of volicitin with the plasma membrane, suggesting that volicitin binding with plasma membrane involves FAC-specific receptors. The receptors perceive MeJA and insect herbivory and activate gene transcription-encoding protein-binding with the plasma membrane, which depends on unknown receptors and JA signaling [21]. Our understanding of the perception of MeJA and volicitin binding with plasma membrane is not yet clear and needs further investigations. The Bph3, a cluster of three genes identified in rice, is known as G-type lectin receptor kinases (OsLecRK1-OsLecRK3) involved in resistance to brown planthopper and white-back planthopper. Molecular cloning and introgression of Bph3 into susceptible rice cultivars lines demonstrated increased resistance to brown planthopper [22]. Leucine-rich repeat receptor kinases (LRR-RK) has been demonstrated to be involved in perception and resistance against Chilo suppressalis (striped stem borer; SSB). Feeding by SSB and treatment with OS of Spodoptera frugiperda increased expression level of OsLRR-RLK1, JA, SA, ethylene biosynthesis genes, and activity of trypsin protease inhibitor (TrypPI). The RNAi-mediated silencing of plants for OsLRR-RLK1 expression showed attenuated activity of TrypPI and resistance to stem borer. We hypothesize that OsLRR-RLK1 may bind with HAEs in rice to induce plant responses [23]. Nicotiana attenuata, a native tobacco plant to western North America, is well-equipped with defense strategies to overcome insect herbivory. Lectin receptor kinases (LecRK1) in tobacco functions in perceiving the elicitors in larval OS. FAC 18:3Glu was found in OS when Manduca sexta feeds on tobacco leaves. Silencing of LecRK1 gene by virus-induced gene silencing (VIGS) and inverted repeated RNA interference revealed the susceptibility of ir-LecRK1 plants to M. sexta. Larvae were grown better and gained higher weight on ir-LecRK1 than wild-type plants, suggesting that LecRK1 is essential to perceive and regulate plant responses to M. sexta [24].
Oviposition produces a necrotic zone at the egg-laying site, and the activity of egg deposition is perceived by LRKs to make the alert signal for plants to elicit defense responses. Mutant plants with a T-DNA insertion in the coding sequence of the Lecrk-I.8 gene were detected to induce plant responses by egg extract. Treatment with egg extracts partially induce the relative PR-1 expression and the level of PR-1 expression was significantly increased in Col-0 plants, suggesting that Lecrk-I.8 is the main receptor in perceiving the egg-derived elicitors in Arabidopsis [40]. It would be interesting to test whether Lecrk-I.8 perceives other elicitors.
The plant receptor-like kinase somatic embryogenesis receptor kinase 1 (SERK1) a distinct member of SERK family, is required for a nucleotide-binding, leucine-rich repeat protein (NB-LRR) called Mi-1 to function in conferring resistance against caterpillar larvae, including Macrosiphum euphorbiae (The potato aphid). Mutant tomato plants silenced with SlSERK1 expression exhibited a higher rate of survival of potato aphids than wild-type plants, where the aphids died during feeding, suggesting the important role of SlSERK1 in the perception of potato aphid attack [41]. Endogenous peptides are released into the apoplast upon wounding or insect feeding and are considered secondary danger signals. Systemin is an 18-amino-acid-long peptide molecule cleaved by prosystemin and spreads throughout a plant body to activate proteinase inhibitor that negatively affects the growth of caterpillars [29]. Many signaling events of plant resistance were initially described in tomatoes, including systemin induce responses, which are conserved in the plant kingdom. In Arabidopsis, AtPep peptide elicitors are recognized by two closely related leucine-rich repeat receptors, AtPEPR1 and AtPEPR2 [27,42,43]. In tomatoes, the high-affinity receptor SYR1 of LRR-RLK family binds to systemin. Introgression lines that lack a single transgene SYR1 were tested for Spodoptera litoralis resistance on tomatoes. Larvae of S. littoralis grow better and gain highly significant weight on introgression line IL3-3, which is deficient in SYR1, than wild-type tomato plants. However, the expression level of the proteinase inhibitor gene (PIN1) was not affected in local and systemic tissues of both transgenic lines, suggesting that SYR1 triggers other defense molecules against S. litoralis larvae [29]. Exogenous treatment with ATP increased defense responses, including cytosolic Ca+ in Arabidopsis. ATP-insensitive mutant DOES NOT RESPOND TO NUCLEOTIDE 1 (DORN1), which is defective in lectin receptor kinase I.9 (Lecrk-I.9), which binds with ATP and is required for ATP-induced responses. Overexpression of DORN1 increased plant responses to wounding, indicating that DORN1 is involved in the perception of extracellular ATP [28].
Recently, researchers in the Schmelz group used a forward genetic mapping approach to show that leucine-rich receptors in Vigna unguiculata (cowpea) and Phaseolus vulgaris (common bean) recognized proteolytic fragments of chloroplastic ATP synthase (inceptin) in OS and elicit the defense responses. Heterologous expression of leucine-rich repeat receptor in tobacco showed that inceptin receptor (INR) regulates plant defenses and confers resistance to Spodoptera exigua [44,45]. In contrast to the pathogen, receptors employed by plants to perceive insect herbivory are not well studied and need further investigations.
3. Detection of Herbivory and Encounter Mechanism
Plants have the ability to distinguish between insect herbivory and mechanical damage by the pattern of tissue feeding and are mediated by the perception of elicitor constituents present in the OS. For example, representative elicitors were chosen and applied to the wounds of different plant species to monitor the induction of phytohormones and volatiles. Results indicated that plants differently respond to insect-derived elicitors. suggesting that plant defense response to various insect herbivores is a species-specific phenomenon [46]. Metabolic study of fatty acid-amino acid conjugates (FACs), the main constituent in the OS of M. sexta, revealed that N-linolenoyle-glutamic acid (18:3-Gln) was metabolized within 30 s after applying to puncture wounds of N. attenuata leaves. Similarly, application of M. sexta OS into wounded leaves showed a 50% decline and changed into a modified form of 18:3 Gln that corresponded to 13-hydroxy-18:3-Glu, 13-hydroperoxy-18:3-Glu, and 13-oxo-13:2-Glu. Nicotiana attenuata silenced plants in the expression of lipoxygenase 2 (LOX2) and lipoxygenase 3 (LOX3), showing a strong reduction in the modified form of 18:3-Gln and suggesting that modified forms of FACs are responsible to elicit plant responses in N. attenuata [47].
Egg deposition of herbivores poses a severe threat to the survival of plants as they change into feeding caterpillars, and plants have developed necrotic arsenals at oviposition regions that are associated with high mortality and reduced hatching rate [48]. Oviposition by Pieris brassicae elicited an increased expression of hundreds of genes after egg deposition. Importantly, transcriptome signature by P. brassicae oviposition was strikingly different in Arabidopsis than observations drawn by chewing herbivores feeding, but oviposition shared similarities in defense-related gene expression changes induced by the biotrophic pathogen during infection [49]. Similarly, oviposition by Pieris brassicae elicited increased accumulation of salicylic acid (SA), and a similar response was observed after perception of pathogen-associated molecular patterns (PAMP). In addition, treatment with egg extract elicited a rapid and strong induction of PAMP-responsive genes, unraveling the shared plant receptors in insect oviposition and PAMP [40].
Frass deposition in a plant’s whorls can suppress plant defenses. FAW larvae deposit copious amounts of frass in whorls during foliar herbivory on maize plants. Applying fall armyworm frass extract to wounded leaves increased the performance of FAW larvae more than caterpillars grown on wounded plants, and frass treatment attenuated the transcript accumulation of defense-related genes, including JA, which indicates insects have encountered proteins that cheat/modulate the plant defenses [50]. During larval feeding on maize, frass can accumulate in maize whorls and deposit for a long period, damaging the plant tissue. Infestation by FAW induces maize chitinases Pr4 and endochitinase A, and these chitinases deposit in the frass and mediate suppression of FAW-induced maize proteinase inhibitor, thereby increasing caterpillar growth in plants [51,52]. Natural enemies of pest herbivores become attracted upon emission of HIPVs, and maize specialist fall armyworm can suppress indirect defenses similar to direct defenses [11,53]. HIPVs emitted by fall armyworm infestation were much weaker than S. littoralis, S. exigua, and Helicoverpa armigera in maize, and FAW feeding could not suppress emission of HIPVs in Gossypium herbaceum (cotton), suggesting that HIPVs suppression is specific for maize [54].
4. Plant and Insect Origin Elicitors
Plants are exposed to biotic stresses by microbes, insects, and animal feeding. To fend off insect herbivory, plants have adapted responses and recognition systems that depend on specific HAEs. HAEs take part in signaling pathways and can activate the defense reaction system in plants. Apart from components of OS, HAEs originate in bacteria, caterpillar frass, the oviposition fluid, and some insect pheromone compounds that can either disrupt or induce plant defenses [55]. In other words, many molecules in OS can cause the plant to manipulate its defense response, involving enzymes such as glucose oxidase and β-glycosidase, peptides such as inceptin, and fatty acid conjugates such as volicitin (Table 1) [4,6,56]. However, as time passes, some plants can overcome this inhibition when they have adapted themselves to recognize the molecules from the insect [57]. Therefore, fatty acid-amino acid conjugates (FACs), or fatty acid amides, were one of the first types identified as an elicitor in the saliva of insects [10]. A two-pronged methodology to study FACs in M. sexta exhibited increased indirect defense response in a host plant by the inducing the volatiles organic compounds (VOCs) and attracting the predators [58]. Since the initial discovery, other types of elicitors have been identified, with their specific molecular activity varying greatly between plant species [46]. Furthermore, inceptins and caeliferins in oral secretions activate insect defensive pathways [59]. Moreover, in previous studies, the induction of defense signaling has been reported in response to the presence of glucose oxidase (GOX) in insect saliva, for example, the Proteinase Inhibitor 2 (PIN2) produced by the salivary component of Ostrinia nubilalis induces in maize and tomato [60,61]. However, some OS inhibit the defense pathway in plants. According to the literature, it has been observed in the larval stages of S. littoralis and P. brassicae, where salivary secretions inhibited defense to allow larvae to grow [62]. As a result, depending on which organism oversees the evolutionary process at that time, the plant or the insect, these molecules can either activate or repress plant defense responses, respectively.
5. Elicitors of Plants’ Intracellular Products
The simplest form of plant elicitors is the intracellular products released upon leaf damage by insect feeding [63]. The intracellular liquid moves to the apoplast and is recognized by DORN1/P2K1 in neighboring, undamaged cells and activates ATP-induced Ca2+ defenses (Figure 2 and Table 1) [28]. In tomatoes, degradation activity of adenosine-5-triphosphate (ATP) was detected in Helicoverpa zea OS assay with tomato leaf fluid. On the other hand, salivary glands of H. zea secrete apyrase and ATP-hydrolyzing enzymes that interfere with ATP signaling and suppress defense-related genes in tomatoes [64]. During insect feeding, regurgitation on leaves provides signaling cues recognized by plants. For example, lignocellulose deposited in the herbivore gut during feeding and digested products can be recognized by receptors in Arabidopsis upon e digestion, functioning as elicitors. However, it is not clear whether these compounds are gut-derived or are plant cell wall degradation products [2,65]. In maize and lima bean, insect feeding produces extracellular self-DNA (esDNA), and plants exposed to esDNA and extracellular heterologous DNA increased plasma membrane potential (Vm) and calcium flux (Ca2+), confirming that esDNA trigger plant responses [25]. Whether the perception of esDNA requires specific receptors other than insect herbivory is another interesting research question to answer.
6. Peptide Elicitors
Endogenous peptide molecules are secreted in plant cells and exclusively found in Solanaceae family and function to elicit plant defenses in response to herbivore feeding (Figure 2). In tomatoes, prosystemin is accumulated and processed in the cytosol by proteolytic cleavage and transported to the apoplast to trigger plant defenses by interacting with membrane-localized leucine-rich receptors (LRR) called PEPRs [66]. In Arabidopsis, endogenous peptide signals of AtPep1 amplify the defense responses by PEP receptor (PEPR1 and PEPR2) [67]. Systemin, a plant peptide hormone of 18 amino acids derived from prosystemin, is a larger precursor protein of about 200 amino acids. Systemin spreads throughout the plant upon wounding and negatively affects the growth of chewing herbivores by activating JA-dependent defenses. Genetic analysis revealed that systemin produces or enlarges the production of systemic signals. Spodoptera lituralis induced a greater level of transcript accumulation of PEPR1, PEPR2, and PROSPEP3 in Arabidopsis. Genetic evidence by using pepr1 and pepr2 mutant plants showed that larvae grew much bigger on mutant plants than wild-types [68], suggesting that PEPR1 and PEPR2 play an important role in the perception of insect herbivory. In addition, systemic trigger the emission of green leaf volatiles (GLVs) to attract the natural enemies of caterpillars as a part of the role in tri-trophic interactions as well as direct defenses by increasing the accumulation of reactive oxygen species (ROS) and phytohormones [69,70]. Peptide elicitors (Pep), similar to systemin, are recognized in Arabidopsis (AtPep1–8) and maize (ZmPep1,3). In maize, ZmPep3 induced an increased level of JA, ethylene, proteinase inhibitors, production of volatiles, benzoxazinoids, and genes encoding defense proteins. The induced plant responses by zmPep3 were similar to those elicited by S. exigu [26]. In rice, brown planthopper infestation significantly induced transcript levels of both OsPep receptors and Pep precursors. Knockout mutant plants impaired in function of OsPEPRs demonstrated susceptibility to brown planthopper feeding, whereas exogenous application of OsPep3 improved the resistance level in rice seedlings against brown planthopper infestation as well as fungal pathogen Magnaporthe oryzae and bacterial pathogen Xanthamonas oryzae pv. oryzae [71]. It is clearly demonstrated that the OsPEPRs signaling is essential for plants to defend against insect herbivores.
7. Elicitors in OS of Insects
Among well-known HAEs, fatty acid–amino acid conjugates (FACs) are a best-studied group of elicitors, which trigger defense responses upon herbivore feeding in many plant species, including maize, soybean, eggplant, and tobacco [4,10,46]. A maize elicitor, volicitin, a hydroxyl FAC [N-(17-hydroxylinolenoyl)-L-glutamine], was isolated from the OS of S. exigua larvae by Alborn and his colleagues in 1997. Applying the volicitin onto the wounds of maize leaves elicits the emission of an increased level of volatiles and attracts parasitic wasps, natural enemies of S. exigua. Wounding without the application of volicitin did not emit the blend of volatiles that attract the natural enemies of herbivores [10]. Since the discovery of volicitin, several FACs have been found in OS of lepidopteran species, and their biological functions are well studied in N. attenuate. Manduca sexta larvae induced increased accumulation of MAPKs, JA, ethylene biosynthesis genes, metabolome, and transcriptome reprogramming in infested and systemic leaves [72,73,74,75,76,77]. Wounding elicits the increased transcript level of transcription factor (TF) WRKY3, and applying the FACs into the wounds of nicotiana leaves caused increased WRKY6 transcript accumulation. Importantly, WRKY3 is required for the elicitation of WRKY6, and silencing of either gene made plants susceptible to herbivores [75]. Manduca sexta herbivory induced high levels of salicylic acid-induced protein kinase (SIPK) and wound-induced protein kinase (WIPK). Silencing of SIPK and WIPK showed decreased defense levels, but M. sexta larvae grown on transgenic plants did not show a difference compared to wild-type plants.
However, green leaf volatiles (GLVs) were attenuated in both transgenic plants and wild-type plants, and the addition of synthetic GLVs restore the increased M. sexta performance in transgenic plants [74]. Glucose oxidase (GOX) is the main component of OS in H. zea, which acts as a salivary protein to suppress the herbivore-induced plant responses in Nicotiana tabacum (tobacco) [36]. In response to larval herbivory, Medicago trancatula (alfalfa) responds by saponins and terpenoid production. Spodoptera exigua (Beat armyworm) suppressed the transcript accumulation of saponins biosynthetic genes for terpenoid production. Researchers hypothesized that GOX may involve in the suppression of gene expression following insect feeding. Further experiments by comparing wounding, the addition of GOX into wounds, and insect feeding confirmed the function of GOX in suppressing defense responses in herbivore attacks [78].
Chemical analysis of the OS in S. exigua indicated that FACs are composed of fatty acid (linoleic acid (LA)/linolenic acid), which is plant originated and insect-derived amino acid (Glu/Gln). Interestingly, 17 hydroxylation and conjugation occur in the midgut of insects, which is important for the biological activity that emits plant volatiles to attract natural enemies of herbivores [79]. Feeding experiments with radio-labeled glutamine, glutamic acid, and linolenic acid to S. litura caterpillars revealed that FACs are involved in nitrogen assimilation and function as glutamine storage in the insect. Glutamine is the main component in insect nitrogen metabolism, and hence, it is not possible for caterpillars to stop production of FACs when feeding on plants, even though plants perceive caterpillar feeding in the presence of FACs [80].
In addition to FAC, there are several other elicitors reported in insect OS. Inceptin, a proteolytic fragment of the chloroplastic ATP synthase γ-subunit, an elicitor isolated in OS of S. frugiperda, perceives the insect herbivory and enhances production of ethylene as well as increases accumulation of phenylpropanoid, VOCs, and protease inhibitor in Vigna unguiculata (cowpea) [9]. Comparing treatment with FAW, OS of Anticarsia gemmatalis, a legume specialist herbivore (Velvetbean caterpillar; VBC), did not induce large production of ethylene and direct herbivory to induce a smaller level of predominant volatile (E)-4,8-dimethyl-1,3,7-nonatriene (DMNT). The examination of OS in VBC for the discovery of truncated form of inceptin in VBC suggested that truncated form of inceptin may be recognized by PPRs and thus suppress the plant defense [81].
8. Elicitors in OS of Non-Lepidopteran Insects
A new class of non-lepidopteran elicitors called “caeliferins” was isolated in OS of Schistocerca americana (grasshopper). FACs found in non-lepidopteran species, for instance, Yoshinaga and her colleagues identified FACs in the gut of two closely related cricket species, Teleogryllus taiwanemma and T. Emma, and larvae of fruit fly, with a similar composition of N-linolenoyl-L-glutamic acid and N-linoleoyl-L-glutamic [82]. Maize seedlings treated with OS of grasshopper emit a blend of volatiles similar to herbivores. Treatment with 5 μL OS of grasshopper emitted a blend of volatiles equal to emission by seedling treated with 100 pmol of volicitin [31].
In the event of coevolution, plants have developed a sophisticated system for the perception of herbivorous feeding through cues derived from insect feeding, saliva, OS, eggs, volatiles, and microbes [13,48,83]. In rice, Nilaparvata lugens-secreted mucin-like protein (NlMLP) was identified by transcriptome and proteome analyses. NlMLP was highly expressed in salivary glands and secreted into rice tissues during brown planthopper feeding. As an elicitor, NlMLP induced the expression of pathogen-responsive genes and increased level of callose deposition, suggesting the important role as an elicitor in rice-insect interactions [38]. In Pisum sativum (pea), oviposition by Bruchus pisorum (pea weevil) increased cell division and created tumor-like growth (neoplasm) at the egg-laying site. Neoplasm delays the entry of pea weevil larvae into pods, and this resistance is mediated by bruchins and can cause neoplastic growth by application of 0.5 pg into the pods [35].
9. Plant-Induced Responses
In response to insect feeding, plants trigger a cascade of short signaling molecules and production of defense-related phytohormones as shown in Figure 2 [13]. Within minutes of tissue damage in Phaseolus lunatus (lima bean), Spodoptera littoralis elicited membrane depolarization and Ca2+ influx. The membrane depolarization was high at the site of S. littoralis feeding and was decreased as the distance increased from the wound site [84]. Interestingly, a research report revealed that herbivore feeding and mechanical wounding induces membrane depolarization at long distances from damaged tissues, and membrane depolarization was dependent on glutamate receptor-like (GRL) channels. In coordination with electric signals, ROS signaling plays an important role in plant response to chewing insects. For instance, this long-distance wound-induced signaling system requires increased production of ROS by membrane-bound RBOHD protein [85]. The superoxide (O2−) generates NADPH oxidases, which are considered important pathogen defense, and the transcript level of Narboh D homologe in N. attenuate was increased by wounding and further amplified by S. littoralis. Consistently, rbohD-silenced plants were susceptible to S. littoralis [86]. Aphids induced a strong accumulation of ROS 3 h after feeding on a resistant near-isogenic line but not in a susceptible line. ROS accumulation was increased with the increase of NADPH oxidase activity only in resistant cultivars, suggesting the involvement of H2O2 in oxidase activity. In addition, insect feeding can rapidly activate the MAPK signaling cascade, which is a highly conserved signaling mechanism among all eukaryotes [76]. In rice, OsMPK3 has been found to positively regulate the defense response against rice striped stem borer (SSB, C. suppressalis) by modulating JA biosynthesis [87].
Early signaling of membrane depolarization, ROS, and MAPK cascade converge into the accumulation of phytohormones JA and conjugate with amino acid isoleucine (Ile). JA-Ile binds with its receptor complex consisting of CORONATINE-INSENSITIVE1 (COI1) and JASMONATE-ZIM DOMAIN (JAZ). The interaction of JA-Ile and its receptor complex leads to the degradation of JAZ and releases transcription factors (TFs) constitutively suppressed by JAZ, activating the expression of downstream defense genes against insect herbivores [13]. Therefore, an early signaling mechanism could be essential for all aspects of plant defense and lead to the induction of plant defense downstream in the form of secondary metabolites and toxic proteins to herbivores. The early signaling events ensure the quantitative, coordinated, spatial, and temporal defense responses. Further research is required to identify and characterize receptors that perceive chemical compounds to elicit defense pathways in plants.
10. Plant Defense against Gall-Inducing Insects
Galls are induced on plants by viruses, mycoplasma, bacteria, fungi, nematodes, insects, mites, and other plants. They are defined by an abnormal plant organ development with ectopic cell proliferation and expansion, generating a wide range of gall morphologies [88,89]. Among them, insect-induced galls have attracted the attention of many researchers because of their unique shapes and wide range of variation. The estimated number of gall-inducing insects ranges from 21,000 to 211,000 [90]. Furthermore, host plant species span numerous phylogenetic lineages, suggesting that gall-inducing systems have evolved independently during the insect evolution [91,92]. Insect galls can be induced on plant leaves, stems, floral buds, flowers, fruits, or roots and exhibit unique shapes [93,94]. Gall-inducing effector candidates have been identified from the transcriptome analysis of ovaries and venom glands of two cynipid gall wasps, Biorhiza pallida and Diplolepis rosae, inducing galls on oak and rose, respectively [95], or the analysis of salivary gland proteome of root-galling grape phylloxera, Daktulosphaira vitifoliae [96]. However, there is no direct evidence showing that these effector candidates have gall-inducing activity in their host plants.
The host plants produce tannins for the protection and gall-inducing insects from herbivores. Aphid galls on R. chinensis accumulate gallotannin, and genes involved in gallotannin biosynthesis [97], gallic acid synthesis [98], and lignin biosynthesis [99] have been identified. In the developing gall of the chestnut gall wasp, Dryocosmus kuriphilus, on the Chinese chestnut, Castanea mollissima, the expression of genes related to metabolic processes, such as phenylpropanoid biosynthesis, secondary metabolism, and plant–pathogen interactions, was altered compared to that of non-infested leaves [100]. Galls induced on elm leaves by a gall-inducing aphid, Tetraneura akinire, were shown to express the genes encoding lignocellulose synthase, suggesting the reinforcement of cell walls to improve resistance to damage by aphids [101].
11. Regulation of Plant Responses at Primed Stage
Priming response is well-documented in plant-pathogen interaction [102]. In the last decade, studies have documented the phenomenon of priming that is triggered by HIPVs, egg deposition, insect herbivory BABA, systemin, and cytokinin to explain enhanced defense plant responses [103,104]. For example, perception of indole signal primes neighboring plants for enhanced release of herbivore-induced plant volatiles (HIPVs) as well as early defense signaling genes within a plant [105,106]. Similarly, HIPVs released upon Mythimna separata infestation (Zea mays L. cv. Royal Dent) primes the maize resistance to insect herbivory and leads to increasing the relative transcript levels of Bowman-Birk type trypsin inhibitor (TI) for five days, and in a promoter region of TI, a set of methylation sites were found demethylated [107]. Recently, M. separata herbivory direct feeding was shown to prime the maize defenses and showed elevated accumulation of benzoxazinoids, JA/JA-Ile, as well as an increased level of defense-related transcripts to M. separata feeding in maize systemic leaves [108]. Diabrotica virgifera infestation on roots of maize seedlings increased DIMBOA content in leaves, and the leaves were primed for the accumulation of chlorogenic acid after subsequent infestation by S. littoralis [109]. However, priming is not only limited to VOCs and direct insect feeding, but the exposure of plants with oviposition equally primes plant defenses and exhibits elevated resistance to insect herbivores. For example, oviposition by the lepidopteran generalist insect S. exigua causes higher mortality, retarded development, and inflicted less feeding damage on oviposition-experienced than on oviposition-unexperienced Nicotiana attenuata plants. In addition, oviposited plants showed a stronger induction of caffeoylputrescine (CP) and trypsin protease inhibitors (TPIs) [110]. A cross-resistance experiment showed that S. exigua larvae suffered reduced performance on M. sexta-oviposited. attenuata plants as they did on S. exigua-oviposited plants [102]. Insect feeding by Pieris rapae and S. exigua in the previous generation on Arabidopsis plants primed enhanced resistance to P. rapae caterpillars in the next generation. Arabidopsis mutants deficient in JA perception or biogenesis of small interfering RNAs were not able to show inherited resistance, suggesting that JA signaling and epigenetics are likely to be involved in transgenerational priming [111].
Although growing lines of evidence have been implicated in the mechanism of priming with epigenetic-based histone modifications and DNA methylation, these epigenetic modifications are responsible for the changes in gene expression of defense-related genes that enable priming response stronger and faster in plants [112]. The hypothetical model of priming phenomena in maize induced by lepidopteran insects has been demonstrated (Figure 3). The mechanism of priming and involvement of epigenetic regulation is at a premature stage, and it would be worthy to investigate how epigenetic modifications regulate priming responses in plants. However, the mechanism of priming induced by insect herbivory not only in local but also in systemic, undamaged leaves has not been investigated so far and deserves much attention.
12. Conclusions
12.1. Concluding Remarks
- Although the mechanism of induced responses is important to understand for better protection of plants, recently, the debate on how plants perceive HAEs to activate downstream-induced defenses has received great attention. However, investigations are required to explore the receptors that perceive insect herbivory.
- Plant receptors perceive elicitors from endo- and exogenous danger signals that are both plant and insect-derived to activate the long- and short-term downstream defenses.
- Upon the perception of herbivory, plants can respond by using exquisite defense strategies. As the perception is strong, plant responses are more robust against caterpillars. The potential of plants to recognize and distinguish between mechanical damage and the kind of insect herbivory indicate the capability of perception of the chemical cues present in the OS of attacker herbivores and feeding on specific host plants.
- Plant responses to insect herbivory are very specific according to the HAEs, and the specificity of plants largely depend on the perception of the nature of elicitors.
- As plants respond stronger and faster to repeated herbivore attacks, it would be interesting to know whether, how, and to what extent plant receptors are involved in the induction of long-term responses in plants.
12.2. Outstanding Questions
- To date, molecular signaling and biosynthesis mechanism of HAEs, such as FACs, caeliferins, egg deposition, and frass, to elicit the defense responses have not been extensively studied. It would be noteworthy to investigate the molecular signal transduction mechanism and biosynthesis of HAEs in plants and insects. Genome editing (e.g., knockout lines) and comparative transcriptomic approaches could be used for functional characterization.
- Indirect defenses are major shareholders in the repellence of herbivores. Genetic and functional characterization are required for the demonstration of genetic control of indirect responses and whether receptors perceiving insect herbivory could also function to emit volatiles to attract natural enemies to fend off insect herbivory.
- Insect infestation triggers short- and long-term plant responses. Defense is costly. There is a need to investigate how plant-defined long-term defenses are sustainable. Studies designed on the hypothesis of trade-off mechanisms could explain long-term and short-term sustainable defense responses.
- The molecular mechanism of induction of defense priming is still at immature stage. According to the specificity of plant responses to insect herbivory, it would be interesting to identify the HAEs that elicit plant responses and prime for enhanced resistance.
- Plants are constantly facing threats to their survival. To what extent plants manage resources for growth and defense by employing the receptors of pathogens and insects requires further research.
Author Contributions
Conceptualization, S.u.M., S.M. and S.C.K.; methodology, S.u.M. and S.M.; software, S.u.M.; validation, S.u.M., S.M., M.H. and S.C.K.; formal analysis, S.u.M., S.M. and M.H.; investigation, S.u.M., S.M., M.H. and S.C.K.; resources, S.u.M.; data curation, S.u.M. and S.M.; writing—original draft preparation, S.u.M., S.M., M.H., S.C.K. and N.S.; writing—review and editing, S.u.M., S.M., M.H., S.C.K. and N.S.; visualization, M.H. and S.C.K.; supervision, S.C.K.; project administration, S.C.K.; funding acquisition, N.S. All authors have read and agreed to the published version of the manuscript.
Funding
S.C.K. thanks the Center for Yunnan Plateau Biological Resources Protection and Utilization, College of Biological Resource and Food Engineering, Qujing Normal University, Qujing, Yunnan, China, for providing research facilities. N.S. thank Chiang Mai University, Thailand for partial financial support.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We are thankful to all the reviewers for their critical suggestions to improve the quality of the manuscript. We also thank Adil Zafar, University of California, Riverside, for editing and improving the quality of our manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wu, J.Q.; Baldwin, I.T. New insights into plant responses to the attack from insect herbivores. Annu. Rev. Genet. 2010, 44, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Arimura, G.I. Making Sense of the Way Plants Sense Herbivores. Trends Plant Sci. 2021, 26, 288–298. [Google Scholar] [CrossRef]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonaventure, G.; VanDoorn, A.; Baldwin, I.T. Herbivore-associated elicitors: FAC signaling and metabolism. Trends Plant Sci. 2011, 16, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Reymond, P. Receptor kinases in plant responses to herbivory. Curr. Opin. Biotechnol. 2021, 70, 143–150. [Google Scholar] [CrossRef]
- Kallure, G.S.; Kumari, A.; Shinde, B.A.; Giri, A.P. Characterized constituents of insect herbivore oral secretions and their influence on the regulation of plant defenses. Phytochemistry 2022, 193, 113008. [Google Scholar] [CrossRef] [PubMed]
- Felton, G.W.; Tumlinson, J.H. Plant-insect dialogs: Complex interactions at the plant-insect interface. Curr. Opin. Plant Biol. 2008, 11, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Mattiacci, L.; Dicke, M.; Maartin, A.P. β-Glucosidase- an elicitor of herbivore-induced plant odor that attracts host-searching parasitic wasps. Proc. Natl. Acad. Sci. USA 1995, 92, 2036–2040. [Google Scholar] [CrossRef] [Green Version]
- Schmelz, E.A.; Carroll, M.J.; LeClere, S.; Phipps, S.M.; Meredith, J.; Chourey, P.S.; Alborn, H.T.; Teal, P.E. Fragments of ATP synthase mediate plant perception of insect attack. Proc. Natl. Acad. Sci. USA 2006, 103, 8894–8899. [Google Scholar] [CrossRef] [Green Version]
- Alborn, H.T.; Turlings, T.C.J.; Jones, T.H.; Stenhagen, G.; Loughrin, J.H.; Tumlinson, J.H. An elicitor of plant volatiles from beet armyworm oral secretion. Science 1997, 276, 945–949. [Google Scholar] [CrossRef]
- Acevedo, F.E.; Rivera-Vega, L.J.; Chung, S.H.; Ray, S.; Felton, G.W. Cues from chewing insects—The intersection of DAMPs, HAMPs, MAMPs and effectors. Curr. Opin. Plant Biol. 2015, 26, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Schmelz, E.A. Impacts of insect oral secretions on defoliation-induced plant defense. Curr. Opin. Insect Sci. 2015, 9, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.; Reymond, P. Molecular interactions between plants and insect herbivores. Annu. Rev. Plant Biol. 2019, 70, 527–557. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-Y.; Mao, Y.-B. Research advances in plant-insect molecular interaction. F1000Research 2020, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zebelo, S.A.; Maffei, M.E. Role of early signalling events in plant-insect interactions. J. Exp. Bot. 2015, 66, 435–448. [Google Scholar] [CrossRef] [Green Version]
- Erb, M.; Kliebenstein, D.J. Plant Secondary Metabolites as Defenses, Regulators, and Primary Metabolites: The Blurred Functional Trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef]
- Qi, J.; Malook, S.u.; Shen, G.; Gao, L.; Zhang, C.; Li, J.; Zhang, J.; Wang, L.; Wu, J. Current understanding of maize and rice defense against insect herbivores. Plant Divers. 2018, 40, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Divekar, P.A.; Narayana, S.; Divekar, B.A.; Kumar, R.; Gadratagi, B.G.; Ray, A.; Singh, A.K.; Rani, V.; Singh, V.; Singh, A.K.; et al. Plant Secondary Metabolites as Defense Tools against Herbivores for Sustainable Crop Protection. Int. J. Mol. Sci. 2022, 23, 2690. [Google Scholar] [CrossRef] [PubMed]
- Howe, G.A.; Jander, G. Plant immunity to insect herbivores. Annu. Rev. Plant Biol. 2008, 59, 41–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.Q.; Zhou, W.W.; Pottinger, S.; Baldwin, I.T. Herbivore associated elicitor-induced defences are highly specific among closely related Nicotiana species. BMC Plant Biol. 2015, 15, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truitt, C.L.; Wei, H.X.; Pare, P.W. A plasma membrane protein from Zea mays binds with the herbivore elicitor volicitin. Plant Cell 2004, 16, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.Q.; Wu, H.; Chen, H.; Liu, Y.L.; He, J.; Kang, H.Y.; Sun, Z.G.; Pan, G.; Wang, Q.; Hu, J.L.; et al. A gene cluster encoding lectin receptor kinases confers broad-spectrum and durable insect resistance in rice. Nat. Biotechnol. 2015, 33, 301–305. [Google Scholar] [CrossRef]
- Hu, L.F.; Ye, M.; Kuai, P.; Ye, M.F.; Erb, M.; Lou, Y.G. OsLRR-RLK1, an early responsive leucine-rich repeat receptor-like kinase, initiates rice defense responses against a chewing herbivore. New Phytol. 2018, 219, 1097–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilardoni, P.A.; Hettenhausen, C.; Baldwin, I.T.; Bonaventure, G. Nicotiana attenuata LECTIN RECEPTOR KINASE1 Suppresses the Insect-Mediated Inhibition of Induced Defense Responses during Manduca sexta Herbivory. Plant Cell 2011, 23, 3512–3532. [Google Scholar] [CrossRef] [Green Version]
- Barbero, F.; Guglielmotto, M.; Capuzzo, A.; Maffei, M.E. Extracellular Self-DNA (esDNA), but Not Heterologous Plant or Insect DNA (etDNA), Induces Plasma Membrane Depolarization and Calcium Signaling in Lima Bean (Phaseolus lunatus) and Maize (Zea mays). Int. J. Mol. Sci. 2016, 17, 1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huffaker, A.; Pearce, G.; Veyrat, N.; Erb, M.; Turlings, T.C.J.; Sartor, R.; Shen, Z.X.; Briggs, S.P.; Vaughan, M.M.; Alborn, H.T.; et al. Plant elicitor peptides are conserved signals regulating direct and indirect antiherbivore defense. Proc. Natl. Acad. Sci. USA 2013, 110, 5707–5712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, Y.; Huffaker, A.; Bryan, A.C.; Tax, F.E.; Ryan, C.A. PEPR2 Is a Second Receptor for the Pep1 and Pep2 Peptides and Contributes to Defense Responses in Arabidopsis. Plant Cell 2010, 22, 508–522. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Tanaka, K.; Yangrong, C.; Yue, Q.; Qiu, J.; Liang, Y.; Yeol Lee, S.; Stacey, G. Identification of a Plant Receptor for Extracellular ATP. Science 2014, 343, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Einig, E.; Almeida-Trapp, M.; Albert, M.; Fliegmann, J.; Mithofer, A.; Kalbacher, H.; Felix, G. The systemin receptor SYR1 enhances resistance of tomato against herbivorous insects. Nat. Plants 2018, 4, 152–156. [Google Scholar] [CrossRef]
- Hopke, J.; Donatha, J.; Blechertb, S.; Boland, W. Herbivore-induced volatiles—The emission of acyclic homoterpenes from leaves of Phaseolus lunatus and Zea mays can be triggered by a b-glucosidase and jasmonic acid. FEBS Lett. 1994, 352, 146–150. [Google Scholar] [CrossRef] [Green Version]
- Alborn, H.T.; Hansen, T.V.; Jones, T.H.; Bennett, D.C.; Tumlinson, J.H.; Schmelz, E.A.; Teal, P.E.A. Disulfooxy fatty acids from the American bird grasshopper Schistocerca americana, elicitors of plant volatiles. Proc. Natl. Acad. Sci. USA 2007, 104, 12976–12981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schafer, M.; Fischer, C.; Meldau, S.; Seebald, E.; Oelmuller, R.; Baldwin, I.T. Lipase Activity in Insect Oral Secretions Mediates Defense Responses in Arabidopsis. Plant Physiol. 2011, 156, 1520–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.J.; Wielsch, N.; Hafke, J.B.; Svatos, A.; Mithofer, A.; Boland, W. A porin-like protein from oral secretions of Spodoptera littoralis larvae induces defense-related early events in plant leaves. Insect Biochem. Mol. Biol. 2013, 43, 849–858. [Google Scholar] [CrossRef]
- Bricchi, I.; Occhipinti, A.; Bertea, C.M.; Zebelo, S.A.; Brillada, C.; Verrillo, F.; De Castro, C.; Molinaro, A.; Faulkner, C.; Maule, A.J.; et al. Separation of early and late responses to herbivory in Arabidopsis by changing plasmodesmal function. Plant J. 2013, 73, 14–25. [Google Scholar] [CrossRef] [Green Version]
- Doss, R.P.; Oliver, J.E.; Proebsting, W.M.; Potter, S.W.; Kuy, S.R.; Clement, S.L.; Williamson, R.T.; Carney, J.R.; DeVilbiss, E.D. Bruchins: Insect-derived plant regulators that stimulate neoplasm formation. Proc. Natl. Acad. Sci. USA 2000, 97, 6218–6223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musser, R.O.; Hum-Musser, S.M.; Eichenseer, H.; Peiffer, M.; Ervin, G.; Murphy, J.B.; Felton, G.W. Caterpillar saliva beats plant defences. Nature 2002, 416, 599–600. [Google Scholar] [CrossRef]
- Diezel, C.; von Dahl, C.C.; Gaquerel, E.; Baldwin, I.T. Different Lepidopteran Elicitors Account for Cross-Talk in Herbivory-Induced Phytohormone Signaling. Plant Physiol. 2009, 150, 1576–1586. [Google Scholar] [CrossRef] [Green Version]
- Shangguan, X.X.; Zhang, J.; Liu, B.F.; Zhao, Y.; Wang, H.Y.; Wang, Z.Z.; Guo, J.P.; Rao, W.W.; Jing, S.L.; Guan, W.; et al. A Mucin-Like Protein of Planthopper Is Required for Feeding and Induces Immunity Response in Plants. Plant Physiol. 2018, 176, 552–565. [Google Scholar] [CrossRef] [Green Version]
- Doares, S.H.; Syrovets, T.; Weiler, E.W.; Ryan, C.A. Oligogalacturonides and Chitosan Activate Plant Defensive Genes through the Octadecanoid Pathway. Proc. Natl. Acad. Sci. USA 1995, 92, 4095–4098. [Google Scholar] [CrossRef] [Green Version]
- Gouhier-Darimont, C.; Schmiesing, A.; Bonnet, C.; Lassueur, S.; Reymond, P. Signalling of Arabidopsis thaliana response to Pieris brassicae eggs shares similarities with PAMP-triggered immunity. J. Exp. Bot. 2013, 64, 665–674. [Google Scholar] [CrossRef] [Green Version]
- Mantelin, S.; Peng, H.C.; Li, B.B.; Atamian, H.S.; Takken, F.L.W.; Kaloshian, I. The receptor-like kinase SlSERK1 is required for Mi-1-mediated resistance to potato aphids in tomato. Plant J. 2011, 67, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Krol, E.; Mentzel, T.; Chinchilla, D.; Boller, T.; Felix, G.; Kemmerling, B.; Postel, S.; Arents, M.; Jeworutzki, E.; Al-Rasheid, K.A.; et al. Perception of the Arabidopsis danger signal peptide 1 involves the pattern recognition receptor AtPEPR1 and its close homologue AtPEPR2. J. Biol. Chem. 2010, 285, 13471–13479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gust, A.A.; Pruitt, R.; Nurnberger, T. Sensing Danger: Key to Activating Plant Immunity. Trends Plant Sci. 2017, 22, 779–791. [Google Scholar] [CrossRef]
- Steinbrenner, A.D.; Munoz-Amatriain, M.; Chaparro, A.F.; Aguilar-Venegas, J.M.; Lo, S.; Okuda, S.; Glauser, G.; Dongiovanni, J.; Shi, D.; Hall, M.; et al. A receptor-like protein mediates plant immune responses to herbivore-associated molecular patterns. Proc. Natl. Acad. Sci. USA 2020, 117, 31510–31518. [Google Scholar] [CrossRef]
- Snoeck, S.; Guayazán-Palacios, N.; Steinbrenner, A.D. Molecular tug-of-war: Plant immune recognition of herbivory. Plant Cell 2022, 34, 1497–1513. [Google Scholar] [CrossRef]
- Schmelz, E.A.; Engelberth, J.; Alborn, H.T.; Tumlinson, J.H.; Teal, P.E.A. Phytohormone-based activity mapping of insect herbivore-produced elicitors. Proc. Natl. Acad. Sci. USA 2009, 106, 653–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanDoorn, A.; Kallenbach, M.; Borquez, A.A.; Baldwin, I.T.; Bonaventure, G. Rapid modification of the insect elicitor N-linolenoyl-glutamate via a lipoxygenase-mediated mechanism on Nicotiana attenuata leaves. BMC Plant Biol. 2010, 10, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilker, M.; Fatouros, N.E. Plant responses to insect egg deposition. Annu. Rev. Entomol. 2015, 60, 493–515. [Google Scholar] [CrossRef]
- Little, D.; Gouhier-Darimont, C.; Bruessow, F.; Reymond, P. Oviposition by pierid butterflies triggers defense responses in Arabidopsis. Plant Physiol. 2007, 143, 784–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, S.; Gaffor, I.; Acevedo, F.E.; Helms, A.; Chuang, W.P.; Tooker, J.; Felton, G.W.; Luthe, D.S. Maize Plants Recognize Herbivore-Associated Cues from Caterpillar Frass. J. Chem. Ecol. 2015, 41, 781–792. [Google Scholar] [CrossRef]
- Ray, S.; Alves, P.C.; Ahmad, I.; Gaffoor, I.; Acevedo, F.E.; Peiffer, M.; Jin, S.; Han, Y.; Shakeel, S.; Felton, G.W.; et al. Turnabout Is Fair Play: Herbivory-Induced Plant Chitinases Excreted in Fall Armyworm Frass Suppress Herbivore Defenses in Maize. Plant Physiol. 2016, 171, 694–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, S.; Basu, S.; Rivera-Vega, L.J.; Acevedo, F.E.; Louis, J.; Felton, G.W.; Luthe, D.S. Lessons from the Far End: Caterpillar FRASS-Induced Defenses in Maize, Rice, Cabbage, and Tomato. J. Chem. Ecol. 2016, 42, 1130–1141. [Google Scholar] [CrossRef]
- Basu, S.; Varsani, S.; Louis, J. Altering Plant Defenses: Herbivore-Associated Molecular Patterns and Effector Arsenal of Chewing Herbivores. Mol. Plant Microbe Interact. 2018, 31, 13–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lange, E.S.; Laplanche, D.; Guo, H.J.; Xu, W.; Vlimant, M.; Erb, M.; Ton, J.; Turlings, T.C.J. Spodoptera frugiperda Caterpillars Suppress Herbivore-Induced Volatile Emissions in Maize. J. Chem. Ecol. 2020, 46, 344–360. [Google Scholar] [CrossRef]
- Abdul Malik, N.A.; Kumar, I.S.; Nadarajah, K. Elicitor and Receptor Molecules: Orchestrators of Plant Defense and Immunity. Int. J. Mol. Sci. 2020, 21, 963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Baldwin, I.T. Herbivory-induced signalling in plants: Perception and action. Plant Cell Environ. 2009, 32, 1161–1174. [Google Scholar] [CrossRef]
- Hogenhout, S.A.; Bos, J.I.B. Effector proteins that modulate plant–insect interactions. Curr. Opin. Plant Biol. 2011, 14, 422–428. [Google Scholar] [CrossRef]
- Gaquerel, E.; Weinhold, A.; Baldwin, I.T. Molecular Interactions between the Specialist Herbivore Manduca sexta (Lepidoptera, Sphigidae) and Its Natural Host Nicotiana attenuata. VIII. An Unbiased GCxGC-ToFMS Analysis of the Plant’s Elicited Volatile Emissions. Plant Physiol. 2009, 149, 1408–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, P.A.; Bos, J.I. Toward understanding the role of aphid effectors in plant infestation. Mol. Plant Microbe Interact. 2013, 26, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Tian, D.; Peiffer, M.; Shoemaker, E.; Tooker, J.; Haubruge, E.; Francis, F.; Luthe, D.S.; Felton, G.W. Salivary glucose oxidase from caterpillars mediates the induction of rapid and delayed-induced defenses in the tomato plant. PLoS ONE 2012, 7, e36168. [Google Scholar] [CrossRef] [Green Version]
- Louis, J.; Shah, J. Arabidopsis thaliana–Myzus persicae interaction: Shaping the understanding of plant defense against phloem-feeding aphids. Front. Plant Sci. 2013, 4, 213. [Google Scholar] [CrossRef] [Green Version]
- Consales, F.; Schweizer, F.; Erb, M.; Gouhier-Darimont, C.; Bodenhausen, N.; Bruessow, F.; Sobhy, I.; Reymond, P. Insect oral secretions suppress wound-induced responses in Arabidopsis. J. Exp. Bot. 2011, 63, 727–737. [Google Scholar] [CrossRef] [Green Version]
- Jeter, C.R.; Tang, W.Q.; Henaff, E.; Butterfield, T.; Roux, S.J. Evidence of a novel cell signaling role for extracellular adenosine triphosphates and diphosphates in Arabidopsis. Plant Cell 2004, 16, 2652–2664. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Peiffer, M.; Luthe, D.S.; Felton, G.W. ATP Hydrolyzing Salivary Enzymes of Caterpillars Suppress Plant Defenses. PLoS ONE 2012, 7, e41947. [Google Scholar] [CrossRef] [Green Version]
- Vadassery, J.; Reichelt, M.; Mithofer, A. Direct Proof of Ingested Food Regurgitation by Spodoptera littoralis Caterpillars during Feeding on Arabidopsis. J. Chem. Ecol. 2012, 38, 865–872. [Google Scholar] [CrossRef]
- Maffei, M.E.; Mithofer, A.; Boland, W. Before gene expression: Early events in plant-insect interaction. Trends Plant Sci. 2007, 12, 310–316. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Pearce, G.; Ryan, C.A. The cell surface leucine-rich repeat receptor for AtPep1, an endogenous peptide elicitor in Arabidopsis, is functional in transgenic tobacco cells. Proc. Natl. Acad. Sci. USA 2006, 103, 10104–10109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klauser, D.; Desurmont, G.A.; Glauser, G.; Vallat, A.; Flury, P.; Boller, T.; Turlings, T.C.J.; Bartels, S. The Arabidopsis Pep-PEPR system is induced by herbivore feeding and contributes to JA-mediated plant defence against herbivory. J. Exp. Bot. 2015, 66, 5327–5336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degenhardt, D.C.; Refi-Hind, S.; Stratmann, J.W.; Lincoln, D.E. Systemin and jasmonic acid regulate constitutive and herbivore-induced systemic volatile emissions in tomato, Solanum lycopersicum. Phytochemistry 2010, 71, 2024–2037. [Google Scholar] [CrossRef] [PubMed]
- Degenhardt, J. Indirect Defense Responses to Herbivory in Grasses. Plant Physiol. 2009, 149, 96–102. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.; Zhang, X.; Liu, J.; Tao, K.; Li, C.; Xiao, S.; Zhang, W.; Li, J.-F. Plant elicitor peptide signalling confers rice resistance to piercing-sucking insect herbivores and pathogens. Plant Biotechnol. J. 2022, 20, 991–1005. [Google Scholar] [CrossRef]
- Giri, A.P.; Wunsche, H.; Mitra, S.; Zavala, J.A.; Muck, A.; Svatos, A.; Baldwin, I.T. Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. VII. Changes in the plant’s proteome. Plant Physiol. 2006, 142, 1621–1641. [Google Scholar] [CrossRef] [Green Version]
- Halitschke, R. Molecular Interactions between the Specialist Herbivore Manduca sexta (Lepidoptera, Sphingidae) and Its Natural Host Nicotiana attenuata. III. Fatty Acid-Amino Acid Conjugates in Herbivore Oral Secretions Are Necessary and Sufficient for Herbivore-Specific Plant Responses. Plant Physiol. 2001, 125, 711–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meldau, S.; Wu, J.; Baldwin, I.T. Silencing two herbivory-activated MAP kinases, SIPK and WIPK, does not increase Nicotiana attenuata’s susceptibility to herbivores in the glasshouse and in nature. New Phytol. 2009, 181, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Skibbe, M.; Qu, N.; Galis, I.; Baldwin, I.T. Induced plant defenses in the natural environment: Nicotiana attenuata WRKY3 and WRKY6 coordinate responses to herbivory. Plant Cell 2008, 20, 1984–2000. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Hettenhausen, C.; Meldau, S.; Baldwin, I.T. Herbivory rapidly activates MAPK signaling in attacked and unattacked leaf regions but not between leaves of Nicotiana attenuata. Plant Cell 2007, 19, 1096–1122. [Google Scholar] [CrossRef] [Green Version]
- Pohnert, G.; Jung, V.; Haukioja, E.; Lempa, K.; Boland, W. New fatty acid amides from regurgitant of lepidopteran (Noctuidae, Geometridae) caterpillars. Tetrahedron 1999, 55, 11275–11280. [Google Scholar] [CrossRef]
- Bede, J.C.; Musser, R.O.; Felton, G.W.; Korth, K.L. Caterpillar herbivory and salivary enzymes decrease transcript levels of Medicago truncatula genes encoding early enzymes in terpenoid biosynthesis. Plant Mol. Biol. 2006, 60, 519–531. [Google Scholar] [CrossRef]
- Pare, P.W.; Alborn, H.T.; Tumlinson, J.H. Concerted biosynthesis of an insect elicitor of plant volatiles. Proc. Natl. Acad. Sci. USA 1998, 95, 13971–13975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshinaga, N.; Aboshi, T.; Abe, H.; Nishida, R.; Alborn, H.T.; Tumlinson, J.H.; Mori, N. Active role of fatty acid amino acid conjugates in nitrogen metabolism in Spodoptera litura larvae. Proc. Natl. Acad. Sci. USA 2008, 105, 18058–18063. [Google Scholar] [CrossRef] [Green Version]
- Schmelz, E.A.; Huffaker, A.; Carroll, M.J.; Alborn, H.T.; Ali, J.G.; Teal, P.E.A. An Amino Acid Substitution Inhibits Specialist Herbivore Production of an Antagonist Effector and Recovers Insect-Induced Plant Defenses. Plant Physiol. 2012, 160, 1468–1478. [Google Scholar] [CrossRef] [Green Version]
- Yoshinaga, N.; Aboshi, T.; Ishikawa, C.; Fukui, M.; Shimoda, M.; Nishida, R.; Lait, C.G.; Tumlinson, J.H.; Mori, N. Fatty acid amides, previously identified in caterpillars, found in the cricket Teleogryllus taiwanemma and fruit fly Drosophila melanogaster larvae. J. Chem. Ecol. 2007, 33, 1376–1381. [Google Scholar] [CrossRef] [PubMed]
- Hilker, M.; Meiners, T. How do plants “notice” attack by herbivorous arthropods? Biol. Rev. 2010, 85, 267–280. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.; Bossi, S.; Spiteller, D.; Mithofer, A.; Boland, W. Effects of feeding Spodoptera littoralis on lima bean leaves. I. Membrane potentials, intracellular calcium variations, oral secretions, and regurgitate components. Plant Physiol. 2004, 134, 1752–1762. [Google Scholar] [CrossRef] [Green Version]
- Toyota, M.; Spencer, D.; Sawai-Toyota, S.; Jiaqi, W.; Zhang, T.; Koo, A.J.; Howe, G.A.; Gilroy, S. Glutamate triggers long-distance calcium-based plant defense signaling. Sci. Adv. 2018, 361, 1112–1115. [Google Scholar] [CrossRef]
- Wu, J.; Wang, L.; Wunsche, H.; Baldwin, I.T. Narboh D, a respiratory burst oxidase homolog in Nicotiana attenuata, is required for late defense responses after herbivore attack. J. Integr. Plant Biol. 2013, 55, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, J. The essential role of jasmonic acid in plant-herbivore interactions—Using the wild tobacco Nicotiana attenuata as a model. J. Genet. Genom. 2013, 40, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Murakami, R.; Ushima, R.; Sugimoto, R.; Tamaoki, D.; Karahara, I.; Hanba, Y.; Wakasugi, T.; Tsuchida, T. A new galling insect model enhances photosynthetic activity in an obligate holoparasitic plant. Sci. Rep. 2021, 11, 13013. [Google Scholar] [CrossRef]
- Price, P.W.; Fernandes, G.W.; Waring, G.L. Adaptive Nature of Insect Galls. Environ. Entomol. 1987, 16, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Espírito-Santo, M. How Many Species of Gall-Inducing Insects Are There on Earth, and Where Are They? Ann. Entomol. Soc. Am. 2009, 100, 95–99. [Google Scholar] [CrossRef]
- Giron, D.; Huguet, E.; Stone, G.N.; Body, M. Insect-induced effects on plants and possible effectors used by galling and leaf-mining insects to manipulate their host-plant. J. Insect Physiol. 2016, 84, 70–89. [Google Scholar] [CrossRef]
- Stone, G.N.; Schönrogge, K. The adaptive significance of insect gall morphology. Trends Ecol. Evol. 2003, 18, 512–522. [Google Scholar] [CrossRef]
- Takeda, S.; Hirano, T.; Ohshima, I.; Sato, M.H. Recent Progress Regarding the Molecular Aspects of Insect Gall Formation. Int. J. Mol. Sci. 2021, 22, 9424. [Google Scholar] [CrossRef] [PubMed]
- Zagorchev, L.; Atanasova, A.; Albanova, I.; Traianova, A.; Mladenov, P.; Kouzmanova, M.; Goltsev, V.; Kalaji, H.M.; Teofanova, D. Functional Characterization of the Photosynthetic Machinery in Smicronix Galls on the Parasitic Plant Cuscuta campestris by JIP-Test. Cells 2021, 10, 1399. [Google Scholar] [CrossRef]
- Cambier, S.; Ginis, O.; Moreau, S.J.M.; Gayral, P.; Hearn, J.; Stone, G.N.; Giron, D.; Huguet, E.; Drezen, J.-M. Gall Wasp Transcriptomes Unravel Potential Effectors Involved in Molecular Dialogues with Oak and Rose. Front. Physiol. 2019, 10, 926. [Google Scholar] [CrossRef]
- Eitle, M.W.; Carolan, J.C.; Griesser, M.; Forneck, A. The salivary gland proteome of root-galling grape phylloxera (Daktulosphaira vitifoliae Fitch) feeding on Vitis spp. PLoS ONE 2019, 14, e0225881. [Google Scholar] [CrossRef]
- Chen, H.; Liu, J.; Cui, K.; Lu, Q.; Wang, C.; Wu, H.; Yang, Z.; Ding, W.; Shao, S.; Wang, H.; et al. Molecular mechanisms of tannin accumulation in Rhus galls and genes involved in plant-insect interactions. Sci. Rep. 2018, 8, 9841. [Google Scholar] [CrossRef]
- Wang, H.; Cui, K.; Shao, S.; Liu, J.; Chen, H.; Wang, C.; Wu, H.; Yang, Z.; Lu, Q.; King-Jones, K.; et al. Molecular response of gall induction by aphid Schlechtendalia chinensis (Bell) attack on Rhus chinensis Mill. J. Plant Interact. 2017, 12, 465–479. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T.; Kimura, S.; Sakamoto, T.; Okamoto, A.; Nakayama, T.; Matsuura, T.; Ikeda, Y.; Takeda, S.; Suzuki, Y.; Ohshima, I.; et al. Reprogramming of the Developmental Program of Rhus javanica During Initial Stage of Gall Induction by Schlechtendalia chinensis. Front. Plant Sci. 2020, 11, 471. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Shi, F.; Chen, Y.; Wang, M.; Zhao, Y.; Geng, G. Transcriptome Analysis of Chinese Chestnut (Castanea mollissima Blume) in Response to Dryocosmus kuriphilus Yasumatsu Infestation. Int. J. Mol. Sci. 2019, 20, 855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.B.; Jin, L.P.; Wei, D.; Huang, Z.H. Study on the differential gene expression of elm leaves fed on by Tetraneura akinire Sasaki. Genes Genom. 2019, 41, 1505–1516. [Google Scholar] [CrossRef] [PubMed]
- Galis, I.; Gaquerel, E.; Pandey, S.P.; Baldwin, I.T. Molecular mechanisms underlying plant memory in JA-mediated defence responses. Plant Cell Environ. 2009, 32, 617–627. [Google Scholar] [CrossRef]
- Engelberth, J.; Alborn, H.T.; Schmelz, E.A.; Tumlinson, J.H. Airborne signals prime plants against insect herbivore attack. Proc. Natl. Acad. Sci. USA 2004, 101, 1781–1785. [Google Scholar] [CrossRef] [Green Version]
- Frost, C.J.; Mescher, M.C.; Carlson, J.E.; De Moraes, C.M. Plant Defense Priming against Herbivores: Getting Ready for a Different Battle1. Plant Physiol. 2008, 146, 818–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erb, M.; Veyrat, N.; Robert, C.A.; Xu, H.; Frey, M.; Ton, J.; Turlings, T.C. Indole is an essential herbivore-induced volatile priming signal in maize. Nat. Commun. 2015, 6, 6273–6283. [Google Scholar] [CrossRef] [Green Version]
- Ye, M.; Glauser, G.; Lou, Y.; Erb, M.; Hu, L. Molecular Dissection of Early Defense Signaling Underlying Volatile-Mediated Defense Regulation and Herbivore Resistance in Rice. Plant Cell 2019, 31, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.; Sugimoto, K.; Ramadan, A.; Arimura, G. Memory of plant communications for priming anti-herbivore responses. Sci. Rep. 2013, 3, 1872–1876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malook, S.U.; Xu, Y.; Qi, J.; Li, J.; Wang, L.; Wu, J. Mythimna separata herbivory primes maize resistance in systemic leaves. J. Exp. Bot. 2021, 72, 3792–3805. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.; Gordon-Weeks, R.; Flors, V.; Camanes, G.; Turlings, T.C.; Ton, J. Belowground ABA boosts aboveground production of DIMBOA and primes induction of chlorogenic acid in maize. Plant Signal. Behav. 2009, 4, 636–638. [Google Scholar] [CrossRef]
- Bandoly, M.; Hilker, M.; Steppuhn, A. Oviposition by Spodoptera exigua on Nicotiana attenuata primes induced plant defence against larval herbivory. Plant J. 2015, 83, 661–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmann, S.; De Vos, M.; Casteel, C.L.; Tian, D.L.; Halitschke, R.; Sun, J.Y.; Agrawal, A.A.; Felton, G.W.; Jander, G. Herbivory in the previous generation primes plants for enhanced insect resistance. Plant Physiol. 2012, 158, 854–863. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.H.; Shan, W.X.; Ayliffe, M.A.; Wang, M.B. Epigenetic Mechanisms: An Emerging Player in Plant-Microbe Interactions. Mol. Plant Microbe Interact. 2016, 29, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
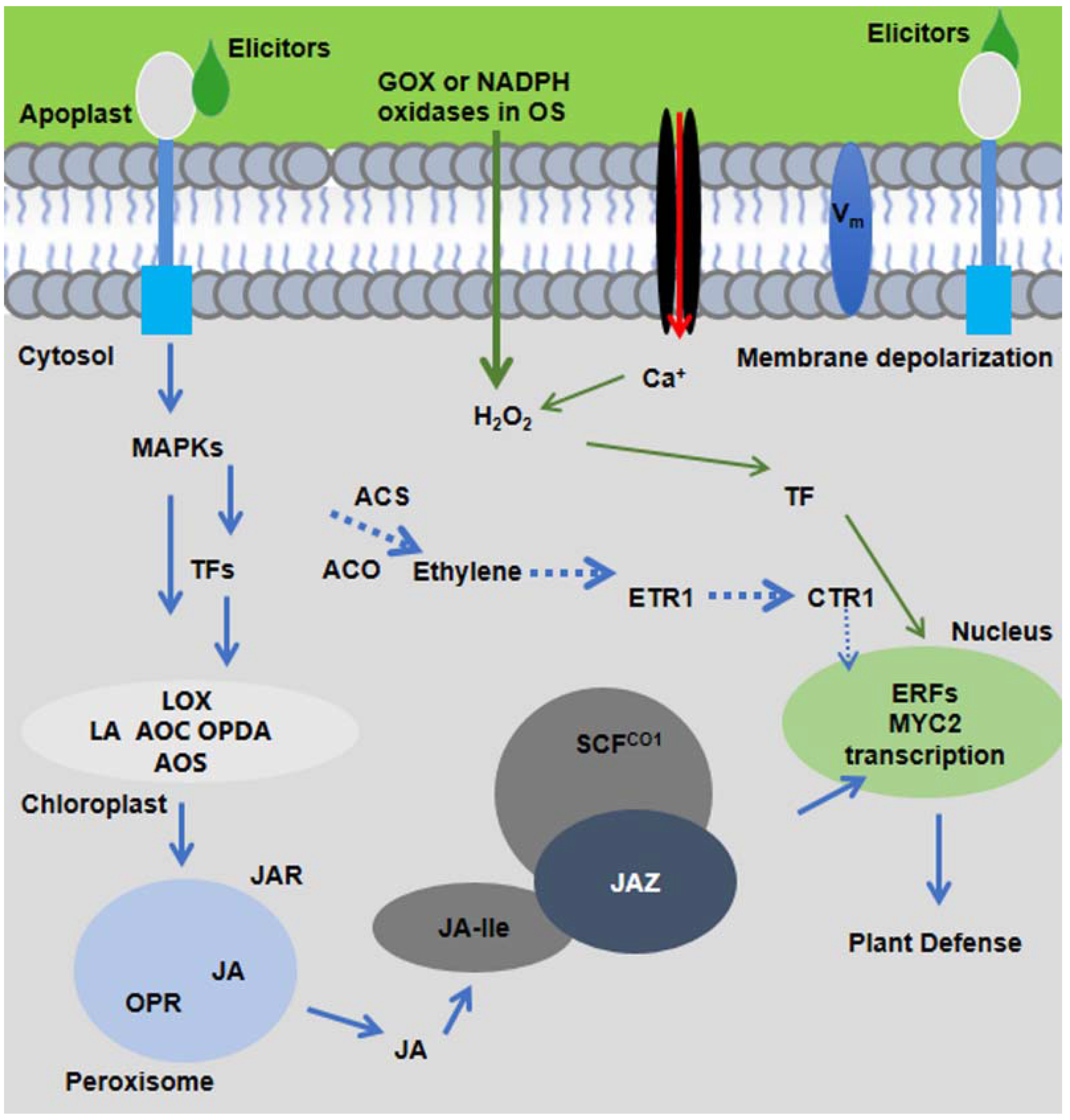
Figure 1.
The molecular signaling model of plant response to insect herbivory. HAEs from oral secretion (OS) of insect herbivores are perceived by plant receptors present in plasma membrane. Within minutes of herbivore feeding, short signaling molecules, such as ROS, Ca2+, MAPK signaling, and membrane depolarization (Vm), are activated and elicit the JA-Ile production. JA-Ile binds with SCFCOI1 and triggers the degradation of JAZ and activates downstream plant defenses.
Figure 1.
The molecular signaling model of plant response to insect herbivory. HAEs from oral secretion (OS) of insect herbivores are perceived by plant receptors present in plasma membrane. Within minutes of herbivore feeding, short signaling molecules, such as ROS, Ca2+, MAPK signaling, and membrane depolarization (Vm), are activated and elicit the JA-Ile production. JA-Ile binds with SCFCOI1 and triggers the degradation of JAZ and activates downstream plant defenses.

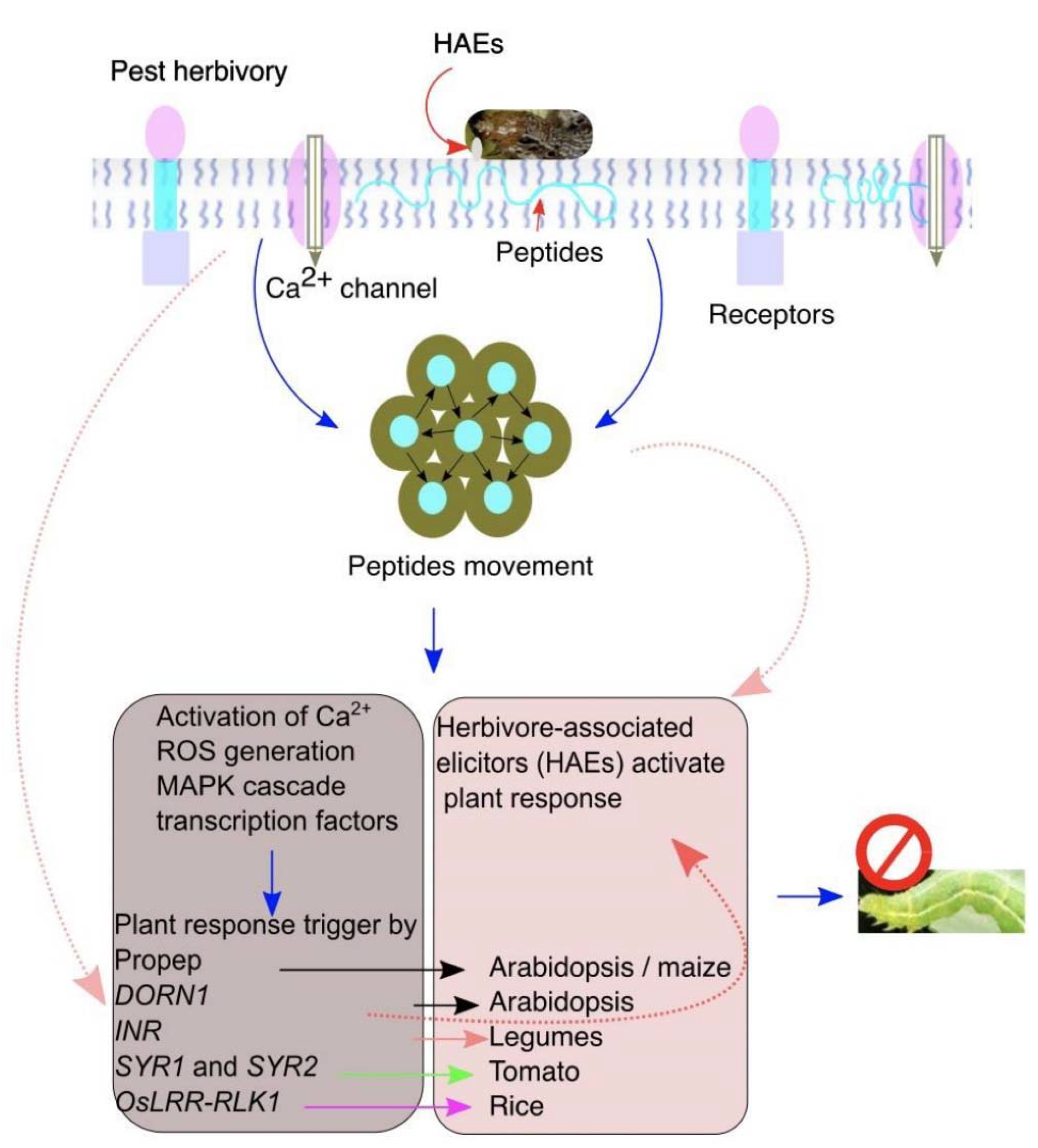
Figure 2.
The plant pattern-recognition receptors (PRRs) recognize herbivore-associated elicitors (HAEs) to elicit plant responses against insect herbivory. A plant perceives and detects herbivory by recognizing HAEs on insect feeding. Damaged cells release intracellular molecules that move into the apoplast and to the undamaged neighboring cells to induce the plant responses. Insect feeding induces endogenous peptides that bind with HAEs to elicit the downstream plant defenses.
Figure 2.
The plant pattern-recognition receptors (PRRs) recognize herbivore-associated elicitors (HAEs) to elicit plant responses against insect herbivory. A plant perceives and detects herbivory by recognizing HAEs on insect feeding. Damaged cells release intracellular molecules that move into the apoplast and to the undamaged neighboring cells to induce the plant responses. Insect feeding induces endogenous peptides that bind with HAEs to elicit the downstream plant defenses.

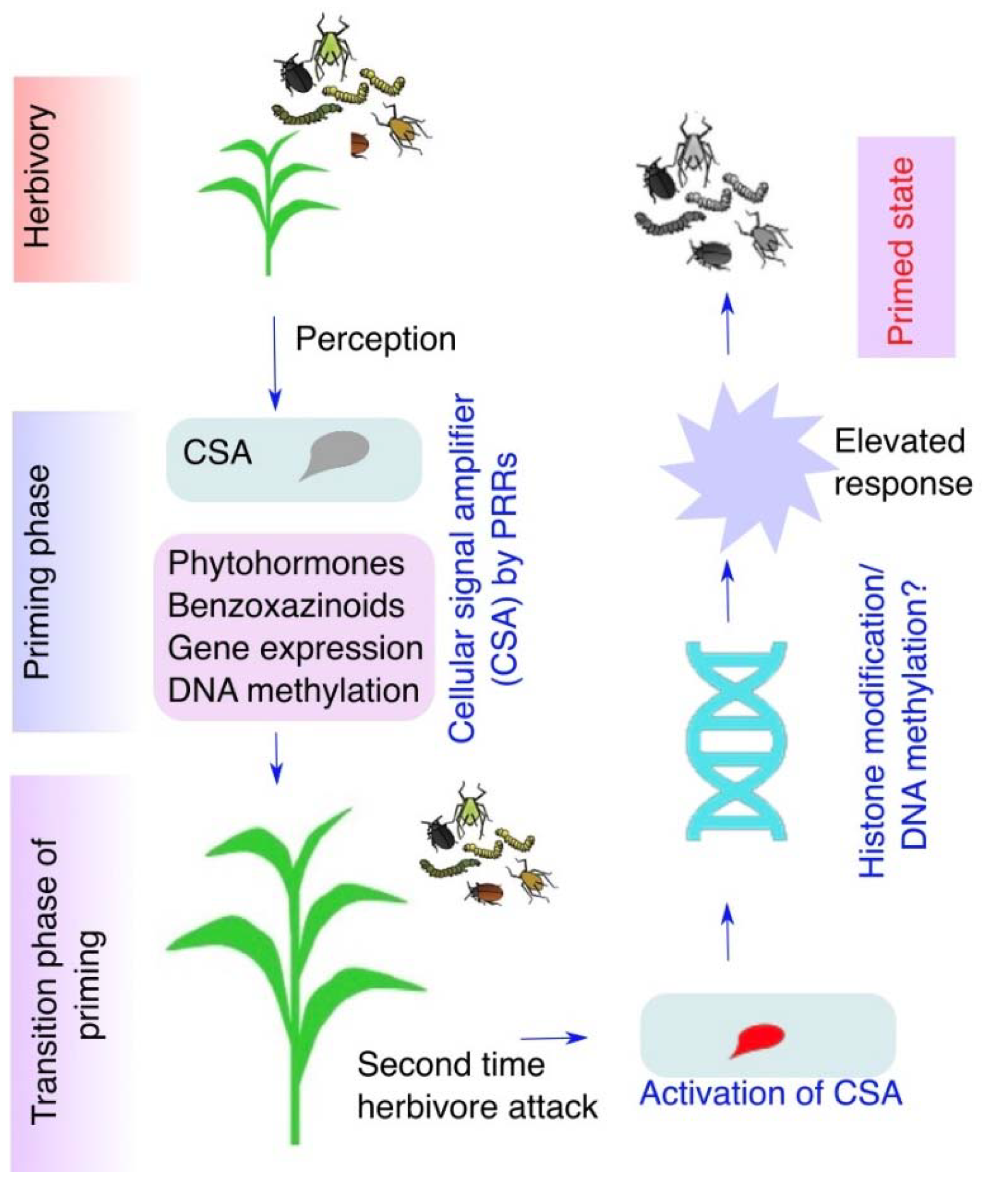
Figure 3.
Caterpillar feeding activates the cellular signal amplifier (CSA) and primes plant responses for enhanced resistance. Caterpillars secrete oral secretion during feeding on plant leaves. Plant pattern-recognition receptors on the surface of plasma membrane specifically perceive the elicitors in the OS and trigger the activation of the inactive cellular signal amplifier (CSA). In response to herbivory CSA enhances the defense response by jasmonic acid (JA), benzoxazinoids (Bxs), gene expression, and DNA methylation to herbivorous feeding in plants.
Figure 3.
Caterpillar feeding activates the cellular signal amplifier (CSA) and primes plant responses for enhanced resistance. Caterpillars secrete oral secretion during feeding on plant leaves. Plant pattern-recognition receptors on the surface of plasma membrane specifically perceive the elicitors in the OS and trigger the activation of the inactive cellular signal amplifier (CSA). In response to herbivory CSA enhances the defense response by jasmonic acid (JA), benzoxazinoids (Bxs), gene expression, and DNA methylation to herbivorous feeding in plants.


{kind=link}
{kind=link}
{kind=link}
Table 1.
The list of HAEs and their known receptors against insect herbivory.
| Elicitors | Receptors | Source of Elicitors | Host Plant | References |
|---|---|---|---|---|
| DNA | n.d. | These elicitors are of plant source | Bean, maize | [25] |
| Pep | Pep receptor (PEPR) | Maize | [26,27] | |
| ATP | ATP receptors (DORN1/P2K1) | Arabidopsis | [28] | |
| Systemin | Systemin receptor (SYR1) | Tomato | [29] | |
| FACs (volicitin) | Unknown membrane proteins | Spodoptera exigua | Maize | [10] |
| β-Glucosidase | n.d. | Pieris brassicae | Maize | [30] |
| Caeliferins | n.d. | Schistocerca americana | Maize | [31] |
| Inceptin | Inceptin receptor (INR) | Spodoptera frugiperda | Maize | [9] |
| Lipase | n.d. | Schistocerca gregaria | Arabidopsis | [32] |
| Porin-like proteins | n.d. | Spodoptera littoralis | Arabidopsis | [33] |
| β-Galactofuranose polysaccharide | HAK/PBL27 | Spodoptera spp. | Arabidopsis | [34] |
| Bruchins | n.d. | Bruchus pisorum, Nilaparvata lugens | Cowpea, pea | [35] |
| Glucose oxidase | n.d. | Helicoverpa zea, Spodoptera exigua, Helicoverpa armigera | Nicotiana | [36,37] |
| Mucin-like protein | n.d. | Callosobruchus maculatus | Rice | [38] |
| Oligouronides | n.d. | Produced by breakdown of plant cell walls by insect feeding | Tomato | [39] |
n.d. = not detected.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Malook, S.u.; Maqbool, S.; Hafeez, M.; Karunarathna, S.C.; Suwannarach, N. Molecular and Biochemical Mechanisms of Elicitors in Pest Resistance. Life 2022, 12, 844. https://doi.org/10.3390/life12060844
AMA Style
Malook Su, Maqbool S, Hafeez M, Karunarathna SC, Suwannarach N. Molecular and Biochemical Mechanisms of Elicitors in Pest Resistance. Life. 2022; 12(6):844. https://doi.org/10.3390/life12060844
Chicago/Turabian StyleMalook, Saif ul, Saiqa Maqbool, Muhammad Hafeez, Samantha Chandranath Karunarathna, and Nakarin Suwannarach. 2022. "Molecular and Biochemical Mechanisms of Elicitors in Pest Resistance" Life 12, no. 6: 844. https://doi.org/10.3390/life12060844
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.