
The Alteration of Intestinal Microbiota Profile and Immune Response in Epinephelus coioides during Pathogen Infection


1
Marine Biotechnology, National Taiwan Ocean University, Keelung 20224, Taiwan
2
Marine Biotechnology, Academia Sinica, Taipei 11529, Taiwan
3
Marine Research Station, Institute of Cellular and Organismic Biology, Academia Sinica, Taipei 11529, Taiwan
4
Institute of Cellular and Organismic Biology, Academia Sinica, Taipei 11529, Taiwan
5
Department of Aquaculture, National Taiwan Ocean University, Keelung 20224, Taiwan
6
Center of Excellence for the Oceans, National Taiwan Ocean University, Keelung 20224, Taiwan
*
Author to whom correspondence should be addressed.
Life 2021, 11(2), 99; https://doi.org/10.3390/life11020099
Submission received: 7 January 2021
/
Revised: 24 January 2021
/
Accepted: 26 January 2021
/
Published: 28 January 2021
(This article belongs to the Special Issue Microbiome–Animal Interaction)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Epinephelus coioides, or grouper, is a high economic value fish species that plays an important role in the aquaculture industry in Asia. However, both viral and bacterial diseases have threatened grouper for many years, especially nervous necrosis virus, grouper iridovirus and Vibrio harveyi, which have caused a bottleneck in the grouper industry. Currently, intestinal microbiota can provide novel insights into the pathogenesis-related factors involved in pathogen infection. Hence, we investigated the comparison of intestinal microbiota communities in control group and pathogen-infected grouper through high-throughput sequencing of the 16S rRNA gene. Our results showed that microbial diversity was decreased, whereas microbial richness was increased during pathogen infection. The individuals in each group were distributed distinctly on the PLSDA diagram, especially the GIV group. Proteobacteria and Firmicutes were the most abundant bacterial phyla in all groups. Interestingly, beneficial genera, Faecalibacterium and Bifidobacterium, predominated in the intestines of the control group. In contrast, the intestines of pathogen-infected grouper had higher levels of harmful genera such as Sphingomonas, Atopostipes, Staphylococcus and Acinetobacter. Additionally, we investigated the expression levels of innate and adaptive immune-related genes after viral and bacterial infection. The results revealed that immunoglobulin T and proinflammatory cytokine levels in the intestine increased after pathogen infection. Through these unique bacterial compositions in diseased and uninfected fish, we could establish a novel therapeutic approach and bacterial marker for preventing and controlling these diseases.
1. Introduction
Orange-spotted grouper (E. coioides) is an important species with high economic value in Asia. Owing to the fast growing and spawning of grouper year-round at temperatures from 22 °C to 28 °C, the orange-spotted grouper is an easier species to culture compared to other grouper species. Unfortunately, the grouper has suffered from the spread of infectious diseases, which have caused mass mortalities, in recent years. Nervous necrosis virus (NNV), grouper iridovirus (GIV) and V. harveyi were reported as the leading causes of death in the early life cycle of grouper [1,2,3].
The main cause of disease in cultured grouper is viral pathogens including NNV and GIV. NNV is one of the most threatening viral diseases affecting over 120 species of cultured marine fishes worldwide, especially post-hatch larvae of Epinephelus spp., and the mortality ranges from 90 to 100% [1,4,5]. NNV mainly attacks the central nervous system and is associated with viral encephalopathy and retinopathy (VER), vacuolation in the brain and eyes, abnormal swimming, dark body color and even anorexia [4,6]. Other major pathogens, such as GIV of the genus Ranavirus, also cause severe mortalities of up to 60% and lead to huge economic loss in the grouper industry [7,8]. The symptoms of infectious grouper include darker skin color, and the gills may bleed and become congested; additionally, the target organs (spleen and head kidney) become swollen [3,9]. Furthermore, the family Vibrionaceae is most often associated with bacterial infection in grouper [10]. V. harveyi has also been referred to as opportunistic bacteria in Lates calcarifer [11], Lateolabrax japonicus [12] and Litopenaeus vannamei [13]. According to previous studies, V. harveyi destroys the tissues on the epidermis and cause skin ulceration called skin ulcer disease [10].
Recently, many studies have revealed that diseases influence the intestinal microbiota composition [14], not only in mammals but also in fish species. The intestine is a complex organ composed of beneficial and harmful bacteria; thus, a beneficial intestinal microbiota has been highlighted as having a key role in maintaining proper digestive functioning [15,16]. The disruption of intestinal microbiota has been reported to correspond with fish after pathogen infections, including infections in Salmo salar, Oncorhynchus mykiss, Larimichthys crocea and Hippocampus kuda [17,18,19,20]. As mentioned above, a series of studies have stated that the composition of intestinal flora is altered by pathogen infection; however, the actual mechanism is still unknown. Therefore, the interaction of pathogens and intestinal microbiota is becoming an attractive topic for microbiological and medical research [21].
We previously developed an intestinal microbiota database of healthy grouper throughout metamorphosis stages. This database revealed that distinct bacterial composition dominated in each stage [22]. To enrich the knowledge about the alteration in intestinal microbiota upon viral and bacterial pathogen infection, we focused on comparing microbial communities of control group with NNV, GIV-and V. harveyi-infected grouper in this study. In addition, we evaluated the mRNA gene expression levels of cytokines-mediated inflammatory factors and immunoglobulins in grouper intestine after viral and bacterial infection. We expected that the intestinal microbiota profile could help us understand the interaction between the host and intestinal microbiomes. Through these data, we established a novel therapeutic approach and bacterial marker for preventing and controlling these diseases.
2. Materials and Methods
2.1. Sampling and Challenge of Juvenile Grouper
In this study, juvenile of orange-spotted groupers were collected from hatcheries at Pingtung, Taiwan. The animal experiment was conducted at National Taiwan Ocean University (Keelung, Taiwan) by following the institutional IACUC guideline (approval number: 106009). The juveniles (1–2 cm) were daily fed a mixture of commercial feeds and Artemia nauplii (0.2 g). NNV and GIV titers were determined using TCID50 (50% tissue culture infective dose) in grouper fin cells (GF-1) and grouper kidney cells (GK) [23]; the LD50 test of V. harveyi was performed according to Li J et al., 2019 [24]. The healthy groupers were separated into four groups in duplicate: the control (n = 20), NNV (n = 20), GIV (n = 20) and Vibrio (n = 20) groups. Fish were acclimatized in 5L tanks equipped with a storage tank system at 30 ℃ with a salinity of 32‰ and constant aeration. The juveniles were injected intraperitoneally with NNV (106.6 TCID50/mL), GIV (108 TCID50/mL) and V. harveyi (6.4 × 107 CFU/mL). The sample collection procedures were performed according to Wayne Knibb et al., 2017 [25]. We collected the intestine samples from five individuals in each tank. All the pathogen infected grouper displayed obvious clinical signs with abnormal swimming behavior (NNV), lethargy (GIV), lesions on skin and tail (Vibrio) with the mortality occurring after seven and 14 days post infection (dpi) with virus and bacteria, respectively. No clinical signs were observed in the control groups. Furthermore, the affected tissues (NNV: brain; GIV: head kidney; V. harveyi: liver) were homogenized and the viruses and bacteria detected by PCR using specific primers for NNV, GIV or V. harveyi. In addition, the intestine and head kidney samples of all groups were collected for RNA and DNA extraction. After dissection, intestine and head kidney samples were washed twice with PBS and immediately stored at −80 ℃ until subsequent use.
2.2. RNA Extraction and cDNA Synthesis
Total RNA was extracted from intestine and head kidney samples by using TRIzol® Reagent (Invitrogen, Carlsbad, CA, USA) following the manufacturer’s instructions, and DNase I treatment (Thermo Scientific™, Waltham, MA, USA) according to the manufacturer’s instructions. The quality of extracted RNA was determined on a 1% agarose gel by electrophoresis. After that, total RNA (1μg) was used for cDNA synthesis by using HiScript I Reverse Transcriptase (BIONOVAS, Halifax, NS, Canada). Reverse transcription was conducted according to the manufacturer’s protocol with random primer. The synthesis condition of cDNA was set at: 65 °C for 5 min, 30 °C for 10 min, 42 °C for 60 min and 70 °C for 15 min.
2.3. Quantitative Real-Time PCR Analysis
Gene expression profiles were performed using the Applied BiosystemTM 7500 Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) on a TOptical Thermocycler® (Analytik Jena AG, Jena, Germany). The qPCR reaction volume was 20 μL in total, containing 1 μL of the cDNA template, 10 μL of the 2X qPCRBIO syGreen Master Mix, 0.8 μL each of the forward and reverse primer (10 pmol/uL) and 7.4 μL distilled water. The amplification condition was initial denaturation at 95 °C for 5 min, followed by 40 cycles of 95 °C for 5 s, 65 °C for 30 s. The melting curve and cooling were performed at the last step of qPCR. The primers used in this study are listed in Table S1. The relative expression levels of the target gene were normalized to beta-actin, a housekeeping gene and calculated by the standard 2−ΔΔCt method. The changes were analyzed by unpaired sample t-test. Statistical significance was accepted at p < 0.05, and high significance was accepted at p < 0.01. All data were expressed as mean ± standard deviation (mean ± SD).
2.4. Genomic DNA Extraction and Gene Sequencing
Total genomic DNA from the intestine was extracted using the Genomic DNA Mini Kit (Geneaid) according to the manufacturer’s instructions. The genomic DNA was monitored on 1% agarose gels to analyze the DNA concentration and purity. Amplicon sequencing (16S V4: 515F-806R) was performed by using 300 bp paired-end raw reads and all the paired-end reads were assembled using FLASH v.1.2.7 [26] at Biotools, Co., Ltd. Demultiplexing was carried out based on barcode identification. As a quality control, low-quality reads (Q < 20) were discarded by the QIIME 1.9.1 pipeline [27]. If three consecutive bases were < Q 20, the read was truncated and the resulting read was retained in the data set only if it was at least 75% of the original length using split_libraries_fastq.py script in QIIME [28]. Sequences were chimera-checked using UCHIME to obtain the effective tags [29,30] and filtered from the data set; then, operational taxonomic unit (OTU) clustering at 97% sequence identity using the UPARSE [31] function in the USEARCH v.7 pipeline [32] was performed. For each representative sequence, the RDP classifier (v.2.2) algorithm [33] was employed to annotate taxonomy classification based on the information retrieved from the Silva Database v.132 [34,35]. This analysis was performed with a minimum confidence threshold of 80% for each assignment. Sequences with one occurrence (singletons) or present in only one sample were filtered out. To analyze the sequence similarities among different OTUs, multiple sequence alignment was conducted by using PyNAST software (v.1.2) [36] against the core-set dataset in the Silva database.
2.5. Bioinformatics and Statistical Analysis
Subsequent analysis (NCBI-Accession number: SUB7303081) of alpha and beta diversities was performed using the normalized data. Alpha diversity was indicative of the species complexity within individual samples based on the observed OTU output from the QIIME pipeline. The number of observed OTUs is the number of different species represented in the microbial community. Beta diversity parameters, the unweighted UniFrac [37,38], were calculated by using the QIIME pipeline. Normally, UniFrac is used for analyzing the comparison between two samples to achieve a UniFrac distance matrix. To further increase the group distinction, a supervised partial-least-squares discriminant analysis (PLS-DA) was used to evaluate and visualize variance based on the OTU level of the intestinal microbiota composition among the groups. PLS-DA was performed using the R package mixOmics. For statistical analysis, the significance of all species among groups at various taxonomic levels was detected using differential abundance analysis with a zero-inflated Gaussian (ZIG) log-normal model as implemented in the “fitFeatureModel” function of the Bioconductor metagenomeSeq package [39]. Moreover, Welch’s t-test was performed using STAMP software (v2.1.3) to identify significant differences among the groups (p value < 0.05) [40].
3. Results
3.1. The Intestinal Microbiota Richness and Diversity Associated with Grouper Disease
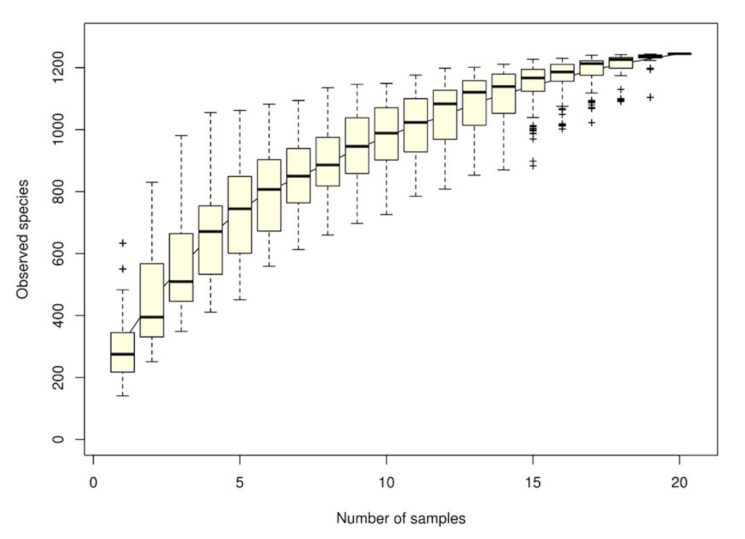
The species accumulation curve (SAC) indicated that the number of microbial OTU had a positive effect on species richness, indicating that the adequacy of sample size could generate a sample survey. When the curve became flat, the number of intestinal microbial species did not increase with the sample size. Figure 1 presents a sharp increase in the curve until a sample size of 20, indicating that the survey sample size of intestinal microbial species was adequate.
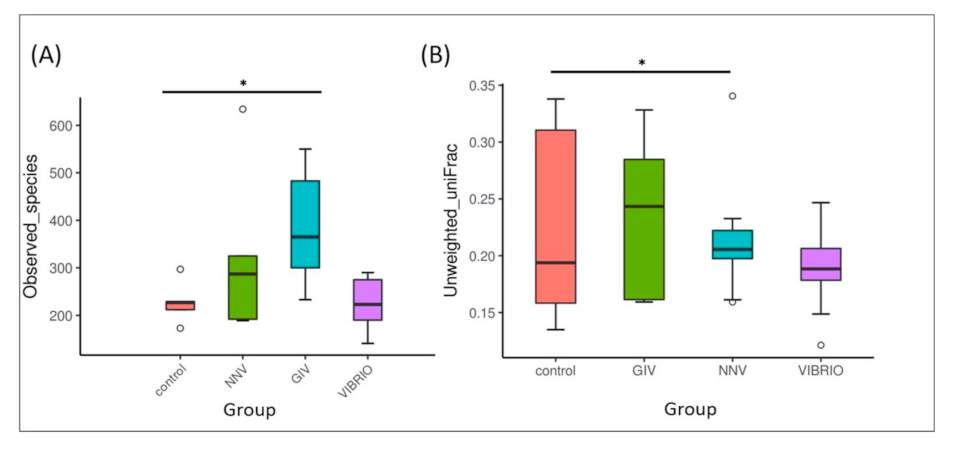
In the alpha diversity analysis, the observed species indices in Figure 2A represent differences in the species richness of control group and pathogen-infected group. The variation in the GIV group was higher than that in the other groups, the NNV and Vibrio groups, while the control group became the least abundant. Similarly, unweighted UniFrac is a qualitative beta diversity measure that compares the significant microbial differences in each sample. As shown in Figure 2B, the microbial beta-diversity was increased in the order of Vibrio, NNV, and GIV, followed by the control group. These results indicated that an immense diversity of intestinal microbiota in healthy grouper could maintain immune homeostasis in the gastrointestinal tract.
3.2. The Intestinal Flora Community Composition of Uninfected and Diseased Grouper
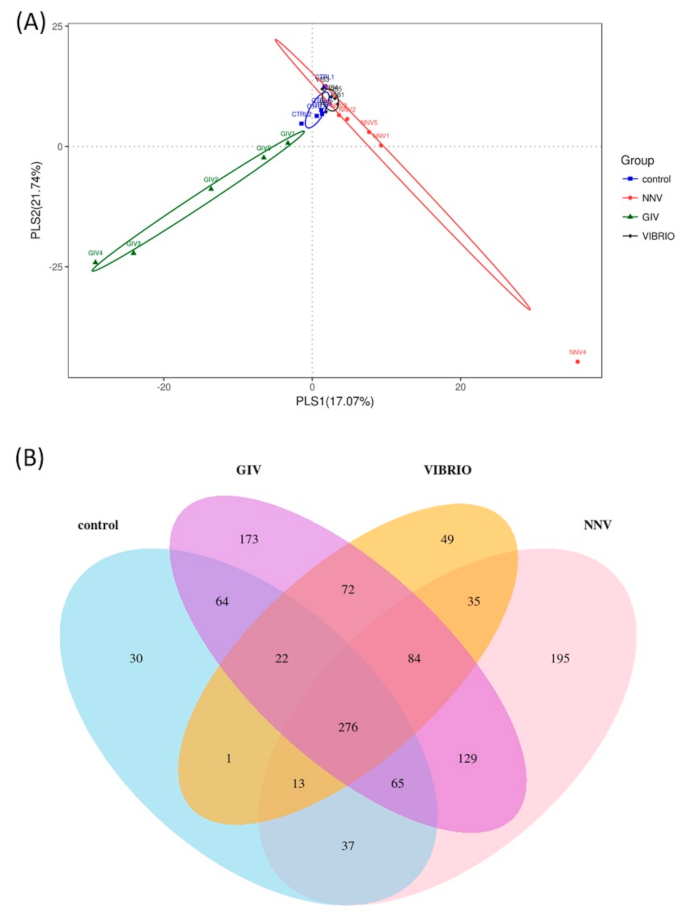
Based on PLS discriminant analysis (PLSDA), each dot represents a sample. The PLSDA diagram of the distribution of microbes significantly separated the GIV group and the other groups, is shown in Figure 3A. Similar distributions were found in the control and Vibrio groups. The NNV group was correlated with these two groups. These results provided a good dataset for clearly understanding the variability and correlation in control group and pathogen-infected groups. A Venn diagram shows the different relationships among the control, NNV, GIV and Vibrio groups (Figure 3B). In total, 1245 OTUs were found in these four groups. There were 276 OTUs in the overlapping area; these OTUs were common to all groups. Furthermore, the nonoverlapping region revealed the unique OTUs in each group, including 30 OTUs, 173 OTUs, 49 OTUs and 195 OTUs that belonged to the control, GIV, Vibrio and NNV groups, respectively. Sixty-four OTUs were shared between the control and GIV groups; one OTU was shared between the control and Vibrio groups; and 37 OTUs were shared between the control and NNV groups. A small number of OTUs were shared between the NNV, GIV, Vibrio groups and the control group, and the overall variation of intestinal microbial communities in pathogen-infected grouper seemed distinct from that of uninfected grouper.
3.3. Interaction between Disease and Intestinal Microbiomes
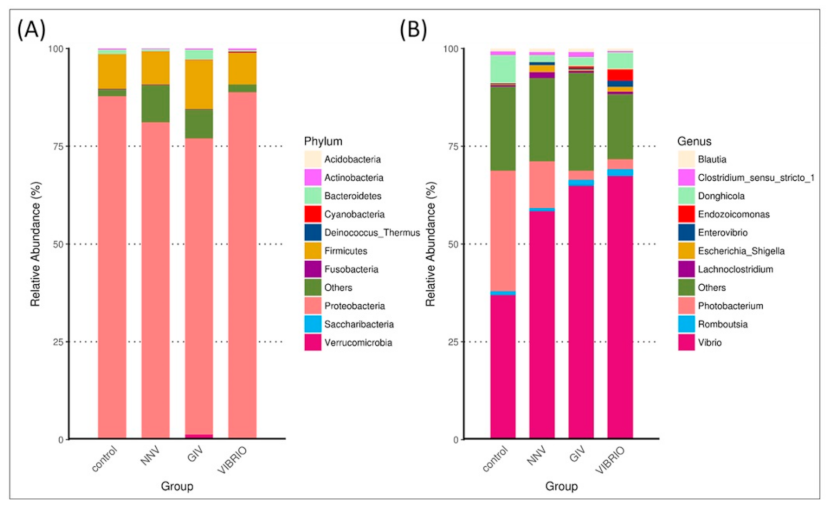
We determined the top ten taxonomic classifications to examine the relative abundance and proportions of microbial phyla (Figure 4A) and genera (Figure 4B) in uninfected and diseased grouper.
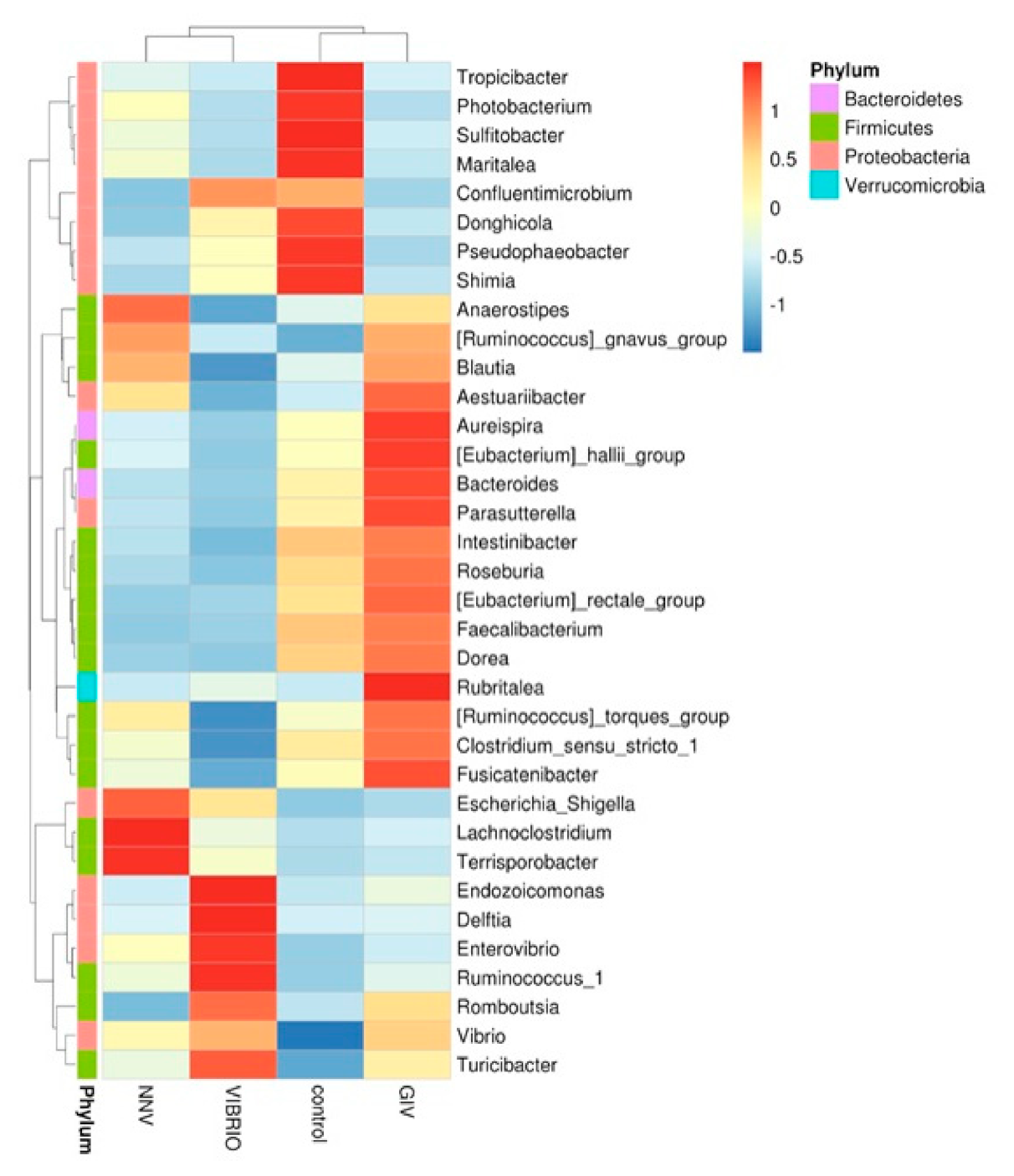
The intestinal microbiome (at the phylum and genus level) showed different microbial communities in each treatment group. Intestinal microbial communities in each group were dominated by two bacterial phyla: Proteobacteria and Firmicutes, with control (88% and 10%), NNV group (81% and 9%), GIV group (76% and 13%) and Vibrio group (89% and 8%), respectively. In the control group, the most abundant genus level was Vibrio (36%), followed by Photobacterium and Donghicola (30% and 7%, respectively). On the other hand, the genus Vibrio had higher connectivity with the NNV, GIV and Vibrio-infected groups, indicating that both viral and bacterial diseases affect the communities of harmful microorganisms. Endozoicomonas accounted for 2% and was overrepresented in the Vibrio group, while the proportion of Escherichia Shigella in the NNV and Vibrio groups (1%) was greater than that in the control and GIV groups, accounting for 0.2% and 0.3%, respectively. Compared with the NNV (1%) and GIV groups (2%), Donghicola was abundant in the control (7%) and Vibrio groups (4%). The results of taxonomic classification (Figure 5) revealed that Proteobacteria might play a key role in both heathy and pathogen-infected grouper. Thus, we further analyzed the top 35 genera and generated a taxa heatmap to analyze the microbiome annotation and abundance information. The microbial communities of the control group consisted of Tropicibacter, Photobacterium, Sulfidobacter, Maritalea, Donghicola, Pseudophaebacter and Shimia. Lachnoclostridium and Terrisporobacter were most abundant genera in NNV-infected grouper. Four genera were enriched in the Vibrio group: Endozoicomonas, Delftia, Enterovibrio and Ruminococcus. In addition, Aureispira, Eubacterium hallii, Bacteroides, Parasutterella, Rubritalea and Fusicatenibacter were highly abundant in the GIV group.
3.4. Differential Abundance Analysis and Statistical Analysis
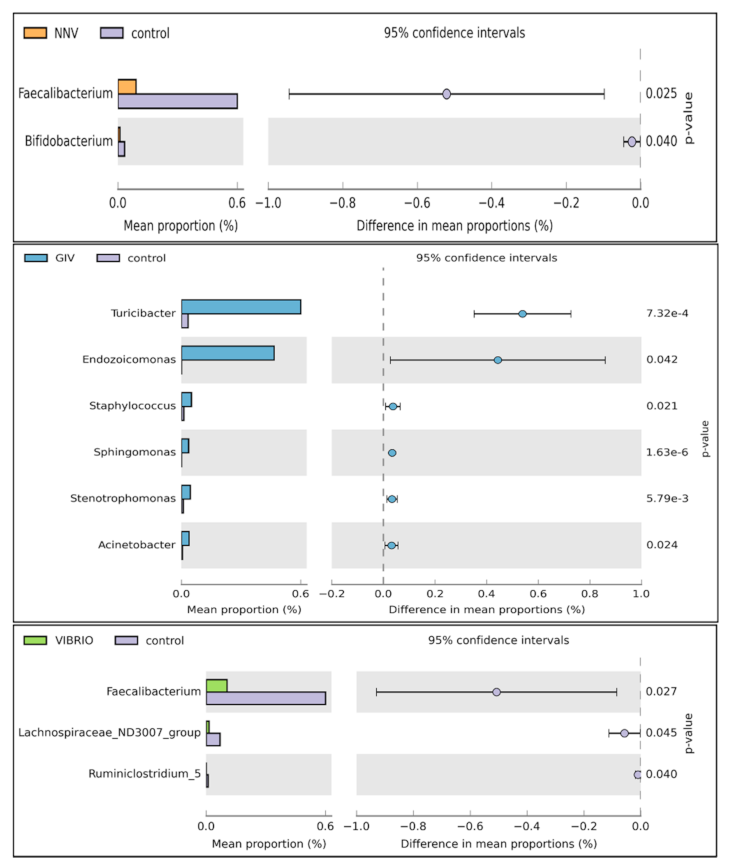
Welch’s t-test was performed on different pathogen-infected groups to further study the microbial community structure. Through statistical analysis, the microbiomes with significant difference in abundance between groups could be found depending on the OTU abundance features. As shown in Figure 6, the induction of Faecalibacterium and Bifidobacterium was significantly affected in the control group compared to the NNV group. Increases in Endozoicomonas, Staphylococcus, Sphingomonas, Stenotrophomonas and Acinetobacter were observed in the GIV group. In the Vibrio and control groups, Faecalibacterium was more abundant and was clearly observed in healthy grouper. From these results, we found that the pathogen may drive changes in the abundance of microbiomes. To support this view, we further investigated the relationship between the intestinal microbiomes of uninfected grouper and NNV, GIV and Vibrio-infected grouper. Therefore, we tried to clearly understand the potential roles of healthy and diseased grouper, and we subsequently assayed the differentially abundant species by metagenomeSeq in the control, NNV, GIV and Vibrio groups (Figure 7).
Compared with control group, NNV, GIV and Vibrio groups showed an increase in the abundances of genus Sphingomonas, Atopostipes, Staphylococcus and Acinetobacter. Among these, Sphingomonas spp. and Staphylococcus spp., which are known to be harmful bacteria in both human and aquatic animals. In addition, Mongoliitalea spp. was clearly superior to the NNV group, whereas the vibrio group was enriched with Ralstonia spp. In conclusion, there was a significant difference among uninfected, NNV-infected, GIV-infected and Vibrio-infected microbiota, which implied that the normal and healthy composition of grouper might be altered through viral and bacterial infection.
3.5. Pathogenic Infection Up-Regulates the Pro-Inflammatory Cytokine Gene Expression in E. coioides Intestine and Head Kidney
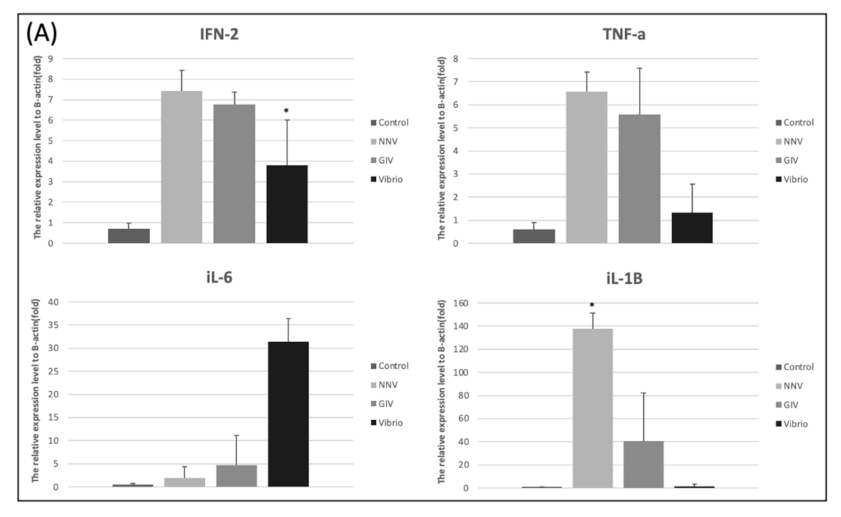
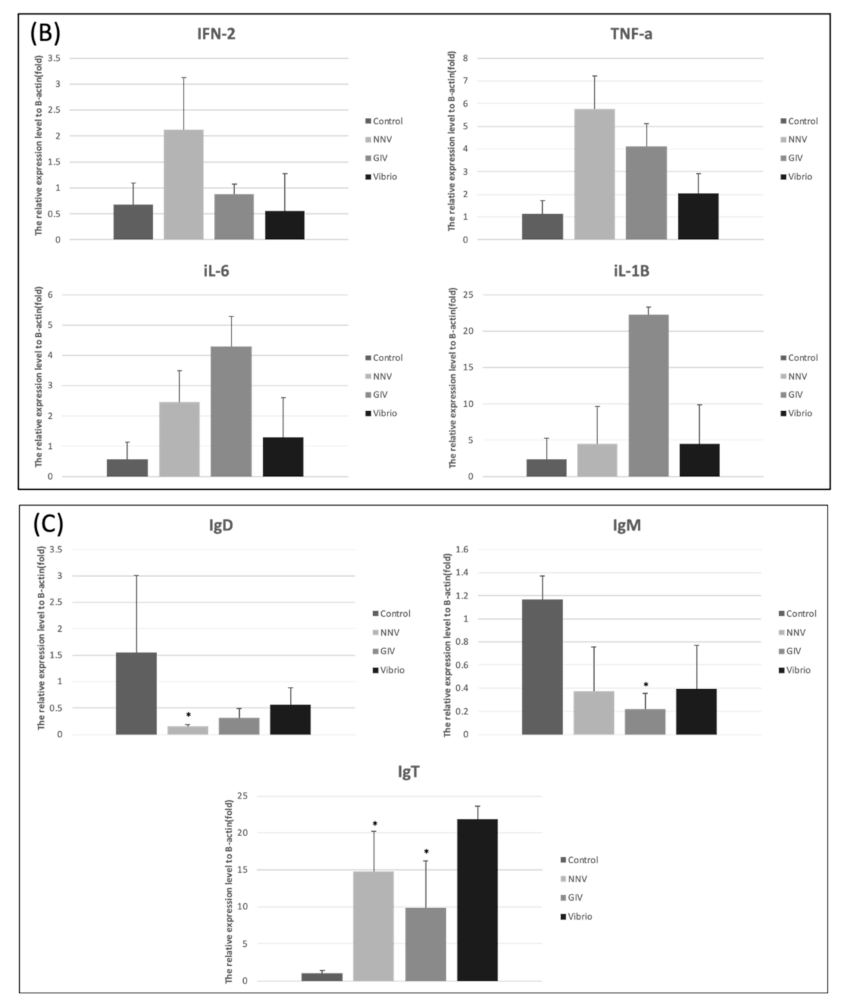
As shown in Figure 8A, treatment with NNV, GIV and Vibrio caused an increase of proinflammatory response in intestines. The expression level of IFN-2 and TNF-α were significantly increased in the Vibrio group. Similarly, iL-6 and iL-1β expression levels in Vibrio group were higher than control group. On the other hand, the higher induction TNF-α, iL-6 and iL-1β were observed in NNV and GIV group compared with the control group. In head kidney, we found the IFN-2 and TNF-α highly expressed in NNV group. iL-6 and iL-1β expressions in the GIV and Vibrio groups were higher than control group. Our results showed the innate immune genes were upregulated in both intestine and head kidney after pathogen infection. To evaluate whether the immune-related genes were expressed in E. coioides after pathogen infection, we analyzed the immunoglobulins (antibodies) in the intestines (Figure 8C). The expression of IgD and IgM was the most upregulated in the control group compared to pathogen infected group, whereas no significant changes were shown in IgT expression. In contrast, we found that IgT expression of the pathogen infected group increased when compared to the control group.
4. Discussion
Recent studies have suggested that microbiome profiling has the capacity to become a feasible application for screening candidate microbiomes against animal or plant diseases, including colorectal cancer and irritable bowel disease in humans, leaf spot disease in plants, and bleaching disease in aquatic animals [41,42,43,44]. Based on our case study, we used metagenomics analysis to analyze the function of the microbiome and the interaction between the microbiome and host. Indeed, this tool can help identify potential therapeutics [45,46].
We aimed to understand the differences in the intestinal microbiota of Epinephelus coioides under normal conditions and pathogen pressure. In this study, we investigated the composition of intestinal microbiomes from uninfected individuals and NNV, GIV and V. harveyi-infected individuals. NNV usually affects the nervous system, causing behavioral abnormalities and extremely high mortalities [25,47], whereas iridovirus infection leads to high mortality in many economic marine and freshwater fish, especially GIV-infected grouper, the mortality of which ranges from 30% to 100% [48,49,50]. In addition, V. harveyi is an opportunistic pathogen causing skin injury and muscle necrosis disease in grouper [51]. As stated above, grouper is susceptible to these infectious diseases and causes significant economic losses in aquaculture [52]. We further examined whether these diseases could alter the composition of the intestinal flora. In Figure 2A, we assumed that the increase in species richness in the intestinal microbiota, during grouper infection with both virus and bacterial pathogens, might help to enhance resistance against the pathogens. Figure 2B indicates that an immense diversity of intestinal microbiota in healthy grouper could maintain immune homeostasis in the gastrointestinal tract. Our findings, which indicated no significant differences in intestinal microbiome diversity (data not shown), are consistent with a previous report [53]. Apart from that, PLSDA (Figure 3A) presented the intestinal microbiome communities from uninfected and infected grouper. The GIV group clustered distinctly among all groups. The NNV and Vibrio groups remained close to the control group, indicating that GIV might affect the enrichment and diversity of intestine communities. Previous studies revealed that the environment, dietary habits or disease also influence the structure of the intestinal bacteria in aquatic animals [18,22,54,55].
The intestinal communities of a majority of marine fish harbor a higher proportion of Proteobacteria and Firmicutes [56,57,58]. In our study, these two phyla were enriched in all groups, revealing that they are the most common bacteria in the intestinal flora of marine fish [59]. At the genus level, the intestine communities of the control group were dominated by Tropicibacter, Photobacterium, Sulfidobacter, Maritalea, Donghicola, Pseudophaeobacter and Shimia. Among these, many Sulfitobacter strains, which are considered sulfite oxidizers, have the ability to produce sulfite oxidases, including the oxidation of organic and inorganic sulfur compounds and carbon monoxide, which can be used in biosensor systems for detecting sulfite [60,61,62,63,64]. Researchers from Iran discovered that Maritalea is a radiation-resistant microorganism that acts as a remoistening agent to endure extreme dryness [65]. Phaeobacter spp., such as P. inhibens and P. gallaeciensis, have been used as probiotics against Vibrio pathogens in scallop and algae cultures [66,67]. Tropicibacter, Photobacterium, Donghicola and Shimia are genera that are abundant in marine organisms or seawater, and Photobacterium mainly live in symbiotic relationships with aquatic organisms [68,69,70,71]. Additionally, we detected an abundance of Lachnoclostridium and Terrisporobacter in the NNV group. Previous studies identified that Lachnoclostridium is related to several viral diseases and has high potential to become a bacterial marker for colorectal adenoma detection [72,73]. In addition, Terrisporobacter was formerly known as a pathogen in humans [74]. The genera Enterovibrio, Delftia, Endozoicomonas and Ruminococcus were abundant in the Vibrio group; these genera are opportunistic bacterial pathogens in both humans and fish [75,76,77,78,79,80]. After infection with GIV, the intestinal microbiota was associated with a wide range of Aureispira, Eubacterium hallii, Bacteroides, Parasutterella, Rubritalea and Fusicatenibacter. Bacteroides and Parasutterella often act as virulence factors in the intestine to form intestinal chronic inflammation [81,82]. Nonetheless, Eubacterium hallii has the ability to maintain intestine metabolic balance due to the formation of propionate [83]. In short, the relationships between these commensal bacteria and their hosts remain unknown, and they may play an important role in the intestinal immune system of grouper, suggesting that the key bacteria present in each group could be further validated as screening microbiome markers for NNV, GIV and V. harveyi infection.
In order to better understand the differential abundance of significantly different microbiomes, we explored the expression profile of microbiomes via Welch’s t-test. The upregulation of Faecalibacterium and Bifidobacterium in control group is consistent with the results of previous microbiota studies [84,85,86,87,88]. Similar studies have shown that Faecalibacterium is predominantly expressed in healthy individuals [89]. These results demonstrated that these two microbiomes could be novel complex supplements considered for use in aquaculture. In contrast, the infection group had an increased abundance of pathogenic bacteria. For example, Staphylococcus warneri was generally recognized as a fish pathogen [90] and is the major cause of infectious diseases in spotted rose snapper [91] and rainbow trout [92,93]. In MetagenomeSeq analysis, we found Mongoliitalea spp. was clearly superior to the NNV group, whereas the vibrio group was enriched with Ralstonia spp., some species reported as infectious bacteria on human, which cause meningitis, respiratory infection and catheter related infection [94]. Acinetobacter spp., which are abundant in NNV, GIV and Vibrio-infected grouper, were identified as serious pathogens that could lead to major disease outbreak in Labeo catla [95]. In the Vibrio group, Ralstonia spp. was a common bacterium isolated from diseased fish [96]. This analysis showed that grouper have a weak immune system that may allow some harmful bacteria to colonize the intestinal tract.
Fish immunity consists of two components, innate and adaptive immune system [97]. Recent studies revealed that cytokines play appreciable innate immune factors in fish which displayed as antibacterial, antifungal, antiviral and antiprotozoal agents [98]. The gut-associated lymphoid tissues (GALT) contain a variety of immune cells and play a critical role in maintaining the balance between pathogenic and commensal microbiota [99]. Moreover, the microbiota has a positive effect on immune regulatory functions of the intestine by stimulating the immune response during pathogen infection [100]. Valero Y et al., [101] noticed that antibacterial and antiviral activity were modulated by several innate and adaptive immune genes. We analyzed different genes, including proinflammatory cytokines and immunoglobulins, to determine the innate and adaptive immune responses triggered by viral and bacterial infections. Previous studies reported that different proinflammatory cytokines production was induced by the pathogens and host commensal communities [102,103,104]. Similarly, all the expression of proinflammatory cytokines (IFN-2, TNF-α, iL-6 and iL-1β) showed upregulation in the intestines and head kidneys of pathogen-infected groupers compared with the control group. As expected, the immunoglobulin-based adaptive immune system (Figure 8B) showed high expression of IgT in intestines of pathogen-infected grouper, whereas IgM and IgD in intestines showed no substantial difference between uninfected and pathogen-infected grouper. This result revealed that immunoglobulin T presented a specialized function in the intestinal mucosa immunity after infection with viral and bacterial pathogens. Strong evidence from a USA team indicated IgT has a specialized function in mucosal immunity, acting as mammalian IgA, which plays a critical role during viral and bacterial pathogen infection in teleost [105].
In summary, we examined the intestinal microbiota profile of uninfected E. coioides and E. coioides infected with different types of pathogens. Our results clearly indicated that disease dramatically affected the diversity and composition of the fish intestinal microbiota. Interestingly, we discovered that the intestinal flora of diseased grouper contained a high abundance of harmful bacteria. In contrast, beneficial bacteria were overrepresented in the control group. However, the symbiotic relationships between either bacteria and bacteria or bacteria and host are currently unknown, and they might be key to helping the host against both viral and bacterial disease. Thus, we speculated that infectious diseases are associated with the intestine immune system, which eventually gives rise to the proinflammatory response [106]. Perhaps a better understanding of the roles these bacteria play may allow us to develop a novel dietary strategy, such as multistrain probiotics, and identify novel bacterial markers for the diagnosis of NNV, GIV and V. harveyi infections. Additionally, identification of the composition and function of disease-associated microorganisms in inflammatory and immune response can be further investigated and considered for the prevention and control of infectious diseases in the aquaculture industry.
Supplementary Materials
The following are available online at https://www.mdpi.com/2075-1729/11/2/99/s1, Table S1: Sequence of primers used in this study.
Author Contributions
Author Contributions: J.-L.W., M.-W.L. and Y.-C.T. conceived and designed the experiments. J.T.X.J. performed the fish experiments. J.-L.W. and Y.-C.T. contributed materials. J.T.X.J. analyzed the metagenomic data. J.T.X.J. and M.-W.L. wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financially supported by the Ministry of Science and Technology, Taiwan (grant numbers: MOST-108-2313-B-019-006, MOST-108-2313-B-019-003 and MOST-109-2823-8-019-001) for supporting this work.
Acknowledgments
We would like to thank Jyh-Yih Chen at the Marine Research Station, Institute of Cellular and Organismic Biology, Academia Sinica, Taiwan for the instrument instructions in the fish trial and sampling.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jeong, K.H.; Kim, H.J. Current status and future directions of fish vaccines employing virus-like particles. Fish Shellfish Immunol. 2020, 100, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Jian, J.; Lu, Y.; Cai, S.; Wang, B.; Tang, J.; Pang, H.; Ding, Y.; Wu, Z.-H. Draft genome sequence of the fish pathogen Vibrio harveyi strain ZJ0603. J. Bacteriol. 2012, 194, 6644–6645. [Google Scholar]
- Peng, C.; Ma, H.; Su, Y.; Wen, W.; Feng, J.; Guo, Z.-X.; Qiu, L. Susceptibility of farmed juvenile giant grouper Epinephelus lanceolatus to a newly isolated grouper iridovirus (genus Ranavirus). Vet. Microbiol. 2015, 177, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Munday, B.; Kwang, J.; Moody, N. Betanodavirus infections of teleost fish: A review. J. Fish Dis. 2002, 25, 127–142. [Google Scholar] [CrossRef]
- Costa, J.; Thompson, K.D. Understanding the interaction between Betanodavirus and its host for the development of prophylactic measures for viral encephalopathy and retinopathy. Fish Shellfish Immunol. 2016, 53, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Souto, S.; Olveira, J.G.; Vázquez-Salgado, L.; Dopazo, C.P.; Bandín, I. Betanodavirus infection in primary neuron cultures from sole. Vet. Res. 2018, 49, 86. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.Y.; Cheng, C.F.; Chiou, P.P.; Liou, C.J.; Yiu, J.C.; Lai, Y.S. Identification and characterization of a late gene encoded by grouper iridovirus 2L (GIV-2L). J. Fish Dis. 2015, 38, 881–890. [Google Scholar] [CrossRef]
- Chen, C.W.; Wu, M.S.; Huang, Y.J.; Lin, P.W.; Shih, C.J.; Lin, F.P.; Chang, C.Y. Iridovirus CARD protein inhibits apoptosis through intrinsic and extrinsic pathways. PLoS ONE 2015, 10, e0129071. [Google Scholar] [CrossRef]
- Chao, C.B.; Chen, C.Y.; Lai, Y.Y.; Lin, C.S.; Huang, H.T. Histological, ultrastructural, and in situ hybridization study on enlarged cells in grouper Epinephelus hybrids infected by grouper iridovirus in Taiwan (TGIV). Dis. Aquat. Org. 2004, 58, 127–142. [Google Scholar] [CrossRef] [Green Version]
- Ningqiu, L.; Junjie, B.; Shuqin, W.; Xiaozhe, F.; Haihua, L.; Xing, Y.; Cunbin, S. An outer membrane protein, OmpK, is an effective vaccine candidate for Vibrio harveyi in Orange-spotted grouper (Epinephelus coioides). Fish Shellfish Immunol. 2008, 25, 829–833. [Google Scholar] [CrossRef]
- Chin, Y.K.; Ina-Salwany, M.Y.; Zamri-Saad, M.; Amal, M.N.A.; Mohamad, A.; Lee, J.Y.; Annas, S.; Al-saari, N. Effects of skin abrasion in immersion challenge with Vibrio harveyi in Asian seabass Lates calcarifer fingerlings. Dis. Aquat. Org. 2020, 137, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Dang, Y.; Nie, L.; Chen, J. Molecular and functional characterisation of a mannose-binding lectin-like gene from Japanese sea bass (Lateolabrax japonicus). Dev. Comp. Immunol. 2020, 104, 103558. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Liu, Y.; Wu, Y.; Jia, L.; Liu, X.; Wang, Q.; Zhang, Y.; Yuanxing, Z. An attenuated Vibrio harveyi surface display of envelope protein VP28 to be protective against WSSV and vibriosis as an immunoactivator for Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 95, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Becattini, S.; Taur, Y.; Pamer, E.G. Antibiotic-induced changes in the intestinal microbiota and disease. Trends Mol. Med. 2016, 22, 458–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.S.; Kao, C.Y. Current understanding of the gut microbiota shaping mechanisms. J. Biomed. Sci. 2019, 26, 59. [Google Scholar] [CrossRef]
- Guinane, C.M.; Cotter, P.D. Role of the gut microbiota in health and chronic gastrointestinal disease: Understanding a hidden metabolic organ. Ther. Adv. Gastroenterol. 2013, 6, 295–308. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Pan, X.; Xu, Y. Altered intestinal microbiota composition associated with enteritis in yellow seahorses Hippocampus kuda (Bleeker, 1852). Curr. Microbiol. 2020, 77, 730–737. [Google Scholar] [CrossRef]
- Pérez-Sánchez, T.; Mora-Sánchez, B.; Vargas, A.; Balcázar, J.L. Changes in intestinal microbiota and disease resistance following dietary postbiotic supplementation in rainbow trout (Oncorhynchus mykiss). Microb. Pathog. 2020, 142, 104060. [Google Scholar] [CrossRef]
- Karlsen, C.; Ottem, K.F.; Brevik, Ø.J.; Davey, M.; Sørum, H.; Winther-Larsen, H.C. The environmental and host-associated bacterial microbiota of Arctic seawater-farmed Atlantic salmon with ulcerative disorders. J. Fish Dis. 2017, 40, 1645–1663. [Google Scholar] [CrossRef]
- Li, C.H.; Xiong, J.B.; Ding, F.F.; Chen, J. Immune and gut bacterial successions of large yellow croaker (Larimichthys crocea) during Pseudomonas plecoglossicida infection. Fish Shellfish Immunol. 2020, 99, 176–183. [Google Scholar] [CrossRef]
- Manzo, V.E.; Bhatt, A.S. The human microbiome in hematopoiesis and hematologic disorders. Blood 2015, 126, 311–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joe, J.T.X.; Chiou, P.P.; Kuo, C.Y.; Lin, J.H.J.; Wu, J.L.; Lu, M.W. The microbiota profile and transcriptome analysis of immune response during metamorphosis stages in orange spotted grouper (Epinephelus coioides). Fish Shellfish Immunol. 2019, 90, 141–149. [Google Scholar]
- Periyasamy, T.; Joe, J.T.X.; Lu, M.W. Cloning and expression of Malabar grouper (Epinephelus malabaricus) ADAR1 gene in response to immune stimulants and nervous necrosis virus. Fish Shellfish Immunol. 2017, 71, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, Z.B.; Zhang, Z.; Zha, J.W.; Qu, S.Y.; Qi, X.Z.; Wang, G.X.; Ling, F. Effects of potential probiotic Bacillus velezensisK2 on growth, immunity and resistance to Vibrio harveyi infection of hybrid grouper (Epinephelus lanceolatus♂ × E. fuscoguttatus♀). Fish Shellfish Immunol. 2019, 93, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Knibb, W.; Luu, G.; Premachandra, H.K.A.; Lu, M.W.; Nguyen, N.H. Regional genetic diversity for NNV grouper viruses across the Indo-Asian region—Implications for selecting virus resistance in farmed groupers. Sci. Rep. 2017, 7, 10658. [Google Scholar] [CrossRef] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.T.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glöckner, F.O. The SILVA and “All-species Living Tree Project (LTP)” taxonomic frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; DeSantis, T.Z.; Andersen, G.L.; Knight, R. PyNAST: A flexible tool for aligning sequences to a template alignment. Bioinformatics 2010, 26, 266–267. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An effective distance metric for microbial community comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Paulson, J.N.; Stine, O.C.; Bravo, H.C.; Pop, M. Differential abundance analysis for microbial marker-gene surveys. Nat. Methods 2013, 10, 1200–1202. [Google Scholar] [CrossRef] [Green Version]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [Green Version]
- CLozupone, A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative beta diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagpal, R.; Kumar, M.; Yadav, A.K.; Hemalatha, R.; Yadav, H.; Marotta, F.; Yamashiro, Y. Gut microbiota in health and disease: An overview focused on metabolic inflammation. Benef. Microbes 2016, 7, 181–194. [Google Scholar] [CrossRef]
- JZackular, P.; Rogers, M.A.; Ruffin, M.T.t.; Schloss, P.D. The human gut microbiome as a screening tool for colorectal cancer. Cancer Prev. Res. 2014, 7, 1112–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bass, D.; Stentiford, G.D.; Wang, H.C.; Koskella, B.; Tyler, C.R. The pathobiome in animal and plant diseases. Trends Ecol. Evol. 2019, 34, 996–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, M.R.; Sample, H.A.; Zorn, K.C.; Arevalo, S.; Yu, G.; Neuhaus, J.; Federman, S.; Stryke, D.; Briggs, B.; Langelier, C.; et al. Clinical metagenomic sequencing for aiagnosis of meningitis and encephalitis. N. Engl. J. Med. 2019, 380, 2327–2340. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Gilbert, J.; Meyer, F. Metagenomics—A guide from sampling to data analysis. Microb. Inform. Exp. 2012, 2, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, M.W.; Ngou, F.H.; Chao, Y.-M.; Lai, Y.S.; Chen, N.-Y.; Lee, F.Y.; Chiou, P.P. Transcriptome characterization and gene expression of Epinephelus spp in endoplasmic reticulum stress-related pathway during betanodavirus infection in vitro. BMC Genom. 2012, 13, 651. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.Y.; Chiou, P.P.; Liou, C.J.; Lai, Y.S. Monoclonal antibody against a putative myristoylated membrane protein encoded by grouper iridovirus 59L gene. Dis. Aquat. Org. 2015, 113, 215–226. [Google Scholar] [CrossRef]
- Kuo, H.C.; Wang, T.Y.; Hsu, H.H.; Chen, P.P.; Lee, S.H.; Chen, Y.M.; Tsai, T.J.; Wang, C.-K.; Ku, H.-T.; Lee, G.-B.; et al. Nervous necrosis virus replicates following the embryo development and dual infection with iridovirus at juvenile stage in grouper. PLoS ONE 2012, 7, e36183. [Google Scholar] [CrossRef]
- Wu, M.S.; Chen, C.W.; Liu, Y.C.; Huang, H.H.; Lin, C.H.; Tzeng, C.S.; Chang, C.Y. Transcriptional analysis of orange-spotted grouper reacting to experimental grouper iridovirus infection. Dev. Comp. Immunol. 2012, 37, 233–242. [Google Scholar] [CrossRef]
- Zhu, Z.M.; Dong, C.F.; Weng, S.P.; He, J.G. The high prevalence of pathogenic Vibrio harveyi with multiple antibiotic resistance in scale drop and muscle necrosis disease of the hybrid grouper, Epinephelus fuscoguttatus (♀) × E. lanceolatus (♂), in China. J. Fish Dis. 2018, 41, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Qin, Y.; Sheng, X.; Xing, J.; Zhan, W. Generation, characterization and application of monoclonal antibodies against matrix protein of hirame novirhabdovirus (HIRRV) in flounder. Dis. Aquat. Org. 2018, 128, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Chen, C.; Jia, L.; He, X.; Zhang, B. Comparison of the intestinal microbiota composition and function in healthy and diseased Yunlong Grouper. AMB Express 2019, 9, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turgay, E.; Steinum, T.M.; Eryalçın, K.M.; Yardımcı, R.E.; Karataş, S. The influence of diet on the microbiota of live-feed rotifers (Brachionus plicatilis) used in commercial fish larviculture. FEMS Microbiol. Lett. 2020, 367, fnaa020. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, S.; Prasad, A. Microflora in fish digestive tract plays significant role in digestion and metabolism. Rev. Fish Biol. Fish. 2011, 22, 11–16. [Google Scholar] [CrossRef]
- Tran, N.T.; Zhang, J.; Xiong, F.; Wang, G.T.; Li, W.X.; Wu, S.G. Altered gut microbiota associated with intestinal disease in grass carp (Ctenopharyngodon idellus). World J. Microbiol. Biotechnol. 2018, 34, 71. [Google Scholar] [CrossRef]
- Qi, X.Z.; Tu, X.; Zha, J.W.; Huang, A.G.; Wang, G.X.; Ling, F. Immunosuppression-induced alterations in fish gut microbiota may increase the susceptibility to pathogens. Fish Shellfish Immunol. 2019, 88, 540–545. [Google Scholar] [CrossRef]
- Parshukov, A.; Kashinskaya, E.; Simonov, E.; Hlunov, O.; Izvekova, G.; Andree, K.; Solovyev, M. Variations of the intestinal gut microbiota of farmed rainbow trout, Oncorhynchus mykiss (Walbaum), depending on the infection status of the fish. J. Appl. Microbiol. 2019, 127, 379–395. [Google Scholar] [CrossRef]
- Liu, H.; Guo, X.; Gooneratne, S.R.; Lai, R.; Zeng, C.; Zhan, F.; Wang, W. The gut microbiome and degradation enzyme activity of wild freshwater fishes influenced by their trophic levels. Sci. Rep. 2016, 6, 24340. [Google Scholar] [CrossRef] [Green Version]
- Long, C.; Lu, X.L.; Gao, Y.; Jiao, B.H.; Liu, X.Y. Description of a sulfitobacter strain and its extracellular cyclodipeptides. Evid.-Based Complement. Altern. Med. 2011, 2011, 393752. [Google Scholar] [CrossRef]
- Buchan, A.; González, J.M.; Moran, M.A. Overview of the marine roseobacter lineage. Appl. Environ. Microbiol. 2005, 71, 5665–5677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, E.P.; Gorshkova, N.M.; Sawabe, T.; Zhukova, N.V.; Hayashi, K.; Kurilenko, V.V.; Alexeeva, Y.; Buljan, V.; Nicolau, D.V.; Mikhailov, V.V.; et al. Sulfitobacter delicatus sp. nov. and Sulfitobacter dubius sp. nov., respectively from a starfish (Stellaster equestris) and sea grass (Zostera marina). Int. J. Syst. Evol. Microbiol. 2004, 54, 475–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Jang, H.J.; Joung, Y.; Kang, I.; Cho, J.C. Sulfitobacter profundi sp. nov., isolated from deep seawater. J. Microbiol. 2019, 57, 661–667. [Google Scholar] [CrossRef] [PubMed]
- González, J.M.; Kiene, R.P.; Moran, M.A. Transformation of sulfur compounds by an abundant lineage of marine bacteria in the alpha-subclass of the class Proteobacteria. Appl. Environ. Microbiol. 1999, 65, 3810–3819. [Google Scholar] [CrossRef] [Green Version]
- Shirsalimian, M.S.; Sepahy, A.A.; Amoozegar, M.A.; Kalantar, S.M.; Dabbagh, R. Isolation of Two Radiation Resistant and Desiccation Tolerant Bacteria, Modestobacter sp. A2 and Maritalea sp. B9, from Gandom Beryan Hill in the Lut Desert of Iran. Microbiology 2018, 87, 363–371. [Google Scholar] [CrossRef]
- Genard, B.; Larouche, O.; Nicolas, J.-L.; Miner, P.; Beaudin, M.-L.; Tremblay, R. Effect of the probiotic strain Phaeobacter gallaeciensis after bacterial challenge on the complete larval development of Pecten maximus. Aquat. Living Resour. 2014, 27, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Yuan, T.; Piva, C.; Spinard, E.J.; Schuttert, C.W.; Rowley, D.C.; Nelson, D.R. The Probiotic Bacterium Phaeobacter inhibens Downregulates Virulence Factor Transcription in the Shellfish Pathogen Vibrio coralliilyticus by N-Acyl Homoserine Lactone Production. Appl. Environ. Microbiol. 2019, 85, e01545-18. [Google Scholar]
- Sung, H.R.; Lee, J.M.; Kim, M.; Yun, B.R.; Shin, K.S. Donghicola tyrosinivorans sp. nov., a tyrosine-degrading bacterium isolated from seawater. Int. J. Syst. Evol. Microbiol. 2015, 65, 4140–4145. [Google Scholar] [CrossRef]
- Choi, D.H.; Cho, B.C. Shimia marina gen. nov., sp. nov., a novel bacterium of the Roseobacter clade isolated from biofilm in a coastal fish farm. Int. J. Syst. Evol. Microbiol. 2006, 56, 1869–1873. [Google Scholar] [CrossRef]
- Ruby, E.G.; Nealson, K.H. Symbiotic association of Photobacterium fischeri with the marine luminous fish Monocentris japonica; a model of symbiosis based on bacterial studies. Biol. Bull. 1976, 151, 574–586. [Google Scholar] [CrossRef]
- Lucena, T.; Pujalte, M.J.; Ruvira, M.A.; Garay, E.; Macián, M.C.; Arahal, D.R. Tropicibacter multivorans sp. nov., an aerobic alphaproteobacterium isolated from surface seawater. Int. J. Syst. Evol. Microbiol. 2012, 62, 844–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangifesta, M.; Mancabelli, L.; Milani, C.; Gaiani, F.; De’Angelis, N.; De’Angelis, G.L.; Van Sinderen, D.; Ventura, M.; Turroni, F. Mucosal microbiota of intestinal polyps reveals putative biomarkers of colorectal cancer. Sci. Rep. 2018, 8, 13974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.Q.; Li, T.; Nakatsu, G.; Chen, Y.-X.; Yau, T.O.; Chu, E.; Wong, S.H.; Szeto, C.H.; Ng, S.C.; Chan, F.K.; et al. A novel faecal Lachnoclostridium marker for the non-invasive diagnosis of colorectal adenoma and cancer. Gut 2019, 69, 1248–1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, M.P.; Domingo, M.C.; Lévesque, S.; Yansouni, C.P. A case report of a deep surgical site infection with Terrisporobacter glycolicus/T. Mayombei and review of the literature. BMC Infect. Dis. 2016, 16, 529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, W.; Cascarano, M.C.; Schlapbach, R.; Katharios, P.; Vaughan, L.; Seth-Smith, H.M.B. Ca. Endozoicomonas cretensis: A novel fish pathogen characterized by genome plasticity. Genome Biol. Evol. 2018, 10, 1363–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andree, K.B.; Rodgers, C.J.; Furones, D.; Gisbert, E. Co-infection with Pseudomonas anguilliseptica and Delftia acidovorans in the European eel, Anguilla anguilla (L.): A case history of an illegally trafficked protected species. J. Fish Dis. 2013, 36, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Davin-Regli, A.; Pagès, J.M. Enterobacter aerogenes and Enterobacter cloacae; versatile bacterial pathogens confronting antibiotic treatment. Front. Microbiol. 2015, 6, 392. [Google Scholar] [CrossRef] [Green Version]
- Novotny, L.; Dvorska, L.; Lorencova, A.; Beran, V.; Pavlik, I. Fish: A potential source of bacterial pathogens for human beings. Vet. Med. 2004, 49, 343–358. [Google Scholar] [CrossRef]
- Sekar, V.T.; Santiago, T.; Vijayan, K.; Alavandi, S.; Raj, V.S.; Rajan, J.; Sanjuktha, M.; Kalaimani, N. Involvement of Enterobacter cloacae in the mortality of the fish, Mugil cephalus. Lett. Appl. Microbiol. 2008, 46, 667–672. [Google Scholar] [CrossRef]
- Henke, M.T.; Kenny, D.J.; Cassilly, C.D.; Vlamakis, H.; Xavier, R.J.; Clardy, J. Ruminococcus gnavus, a member of the human gut microbiome associated with Crohn’s disease, produces an inflammatory polysaccharide. Proc. Natl. Acad. Sci. USA 2019, 116, 12672–12677. [Google Scholar] [CrossRef] [Green Version]
- Wexler, H.M. Bacteroides: The good, the bad, and the nitty-gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.J.; Wu, H.; Wu, S.D.; Lu, N.; Wang, Y.T.; Liu, H.N.; Dong, L.; Liu, T.T.; Shen, X.Z. Parasutterella, in association with irrit2 able bowel syndrome and intestinal chronic inflammation. J. Gastroenterol. Hepatol. 2018, 33, 1844–1852. [Google Scholar] [CrossRef] [PubMed]
- Engels, C.; Ruscheweyh, H.J.; Beerenwinkel, N.; Lacroix, C.; Schwab, C. The Common Gut Microbe Eubacterium hallii also Contributes to Intestinal Propionate Formation. Front. Microbiol. 2016, 7, 713. [Google Scholar] [CrossRef] [PubMed]
- Shigwedha, N.; Jia, L. Bifidobacterium in human GI tract: Screening, isolation, survival and growth kinetics in simulated gastrointestinal conditions. In Lactic Acid Bacteria—R & D for Food, Health and Livestock Purposes; InTech: Rijeka, Croatia, 2013; pp. 281–308. [Google Scholar]
- Gao, Y.M.; Zou, K.S.; Zhou, L.; Huang, X.D.; Li, Y.Y.; Gao, X.Y.; Chen, X.; Zhang, X. Deep insights into gut microbiota in four carnivorous coral reef fishes from the South China Sea. Microorganisms 2020, 8, 426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín, R.; Miquel, S.; Benevides, L.; Bridonneau, C.; Robert, V.; Hudault, S.; Chain, F.; Berteau, O.; Azevedo, V.; Chatel, J.M.; et al. Functional characterization of novel Faecalibacterium prausnitzii strains isolated from healthy volunteers: A step forward in the use of F. prausnitzii as a next-generation probiotic. Front. Microbiol. 2017, 8, 1226. [Google Scholar]
- Arboleya, S.; Watkins, C.; Stanton, C.; Ross, R.P. Gut Bifidobacteria populations in human health and aging. Front. Microbiol. 2016, 7, 1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picard, C.; Fioramonti, J.; Francois, A.; Robinson, T.; Neant, F.; Matuchansky, C. Review article: Bifidobacteria as probiotic agents—Physiological effects and clinical benefits. Aliment. Pharm. Ther. 2005, 22, 495–512. [Google Scholar] [CrossRef]
- Laursen, M.F.; Laursen, R.P.; Larnkjær, A.; Mølgaard, C.; Michaelsen, K.F.; Frøkiær, H.; Bahl, M.I.; Licht, T.R. Faecalibacterium gut colonization accelerated by presence older siblings. mSphere 2017, 2, e00448-17. [Google Scholar] [CrossRef] [Green Version]
- Bunnoy, A.; Na-Nakorn, U.; Kayansamruaj, P.; Srisapoome, P. Acinetobacter strain KUO11TH, a unique organism related to Acinetobacter pittii and isolated from the skin mucus of healthy bighead catfish and its efficacy against several fish pathogens. Microorganisms 2019, 7, 549. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Gil, B.; Fajer-Avila, E.; García-Vargas, F. Vibrios of the spotted rose snapper Lutjanus guttatus Steindachner, 1869 from northwestern Mexico. J. Appl. Microbiol. 2007, 102, 1518–1526. [Google Scholar] [CrossRef]
- Austin, B.; Austin, D.; Sutherland, R.; Thompson, F.; Swings, J. Pathogenicity of vibrios to rainbow trout (Oncorhynchus mykiss, Walbaum) and Artemia nauplii. Environ. Microbiol. 2005, 7, 1488–1495. [Google Scholar] [CrossRef]
- Musharrafieh, R.; Tacchi, L.; Trujeque, J.; LaPatra, S.; Salinas, I. Staphylococcus warneri, a resident skin commensal of rainbow trout (Oncorhynchus mykiss) with pathobiont characteristics. Vet. Microbiol. 2014, 169, 80–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, M.P.; Adley, C.C. Ralstonia spp emerging global opportunistic pathogens. Eur. J. Clin Microbiol. Infect. 2014, 33, 291–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malick, R.C.; Bera, A.K.; Chowdhury, H.; Bhattacharya, M.; Abdulla, T.; Swain, H.S.; Baitha, R.; Kumar, V.; Das, B.K. Identification and pathogenicity study of emerging fish pathogens Acinetobacter junii and Acinetobacter pittii recovered from a disease outbreak in Labeo catla (Hamilton, 1822) and Hypophthalmichthys molitrix (Valenciennes, 1844) of freshwater wetland in West Bengal, India. Aquac. Res. 2020, 51, 2410–2420. [Google Scholar]
- Wamala, S.P.; Mugimba, K.K.; Mutoloki, S.; Evensen, Ø.; Mdegela, R.; Byarugaba, D.K.; Sørum, H. Occurrence and antibiotic susceptibility of fish bacteria isolated from Oreochromis niloticus (Nile tilapia) and Clarias gariepinus (African catfish) in Uganda. Fish Aquat. Sci. 2018, 21, 6. [Google Scholar] [CrossRef]
- Salinas, I. The Mucosal Immune System of Teleost Fish. Biology 2015, 4, 525–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neshani, A.; Zare, H.; Akbari Eidgahi, M.R.; Khaledi, A.; Ghazvini, K. Epinecidin-1, a highly potent marine antimicrobial peptide with anticancer and immunomodulatory activities. BMC Pharm. Toxicol. 2019, 20, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, T.; Balcázar, J.L.; Ruiz-Zarzuela, I.; Halaihel, N.; Vendrell, D.; Blas, I.D.; Múzquiz, J.L. Host-microbiota interactions within the fish intestinal ecosystem. Mucosal Immunol. 2010, 3, 355–560. [Google Scholar] [CrossRef]
- Arrazuria, R.; Pérez, V.; Molina, E.; Juste, R.A.; Khafipour, E.; Elguezabal, N. Diet induced changes in the microbiota and cell composition of rabbit gut associated lymphoid tissue (GALT). Sci. Rep. 2018, 8, 14103. [Google Scholar] [CrossRef]
- Valero, Y.; García-Alcázar, A.; Esteban, M.Á.; Cuesta, A.; Chaves-Pozo, E. Antimicrobial response is increased in the testis of European sea bass, but not in gilthead seabream, upon nodavirus infection. Fish Shellfish Immunol. 2015, 44, 203–213. [Google Scholar] [CrossRef]
- Maehr, T.; Costa, M.M.; Vecino, J.L.G.; Wadsworth, S.; Martin, S.; Wang, T.; Secombes, C.J. Transforming growth factor-β1b: A second TGF-β1 paralogue in the rainbow trout (Oncorhynchus mykiss) that has a lower constitutive expression but is more responsive to immune stimulation. Fish Shellfish Immunol. 2013, 34, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chang, X.; Wu, H.; Xiao, J.; Gao, Y.; Zhang, Y. Role of intestinal inflammation in predisposition of Edwardsiella tarda infection in zebrafish (Danio rerio). Fish Shellfish Immunol. 2014, 41, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Cook, J.A.; Gilkeson, G.S.; Luttrell, L.M.; Wang, L.; Borg, K.; Halushka, P.V.; Fan, H. Increased expression of beta-arrestin 1 and 2 in murine models of rheumatoid arthritis: Isoform specific regulation of inflammation. Mol. Immunol. 2011, 49, 64–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piazzon, M.C.; Galindo-Villegas, J.; Pereiro, P.; Estensoro, I.; Calduch-Giner, J.A.; Gómez-Casado, E.; Novoa, B.; Mulero, V.; Sitjà-Bobadilla, A.; Pérez-Sánchez, J. Differential modulation of IgT and IgM upon parasitic, bacterial, viral, and dietary challenges in a perciform fish. Front. Immunol. 2016, 7, 637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parolini, C. Effects of fish n-3 PUFAs on intestinal microbiota and immune system. Mar. Drugs 2019, 17, 374. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Species accumulation curve derived by grouper intestine samples and estimations of the number of intestinal microbiomes in the gastrointestinal tract of grouper. Each dot represents the total richness for all samples pooled. The x-axis is the number of samples and the y-axis is the number of observed species.
Figure 1.
Species accumulation curve derived by grouper intestine samples and estimations of the number of intestinal microbiomes in the gastrointestinal tract of grouper. Each dot represents the total richness for all samples pooled. The x-axis is the number of samples and the y-axis is the number of observed species.

Figure 2.
Species diversity. (A) Alpha diversity analysis with observed species indices was used to evaluate the number of different species in the intestinal flora. (B) Beta diversity analysis with unweighted UniFrac was used to evaluate the differences in microbial communities among samples in terms of species complexity [40]. The asterisk (*) represents significant difference at * p < 0.05; ** p < 0.01; *** p < 0.001 from controls. Note: The upper and lower range of the standard deviation are indicated in each box with a horizontal line with dots as outliers. The interquartile range is Q3-Q1 (IQ), the maximum range is Q3 + 1.5 * IQ, and the minimum range is Q1 − 1.5 * IQ, which generate the area between the minimum and maximum; a line inside the boxplot shows the median. The group names are plotted on the X-axis.
Figure 2.
Species diversity. (A) Alpha diversity analysis with observed species indices was used to evaluate the number of different species in the intestinal flora. (B) Beta diversity analysis with unweighted UniFrac was used to evaluate the differences in microbial communities among samples in terms of species complexity [40]. The asterisk (*) represents significant difference at * p < 0.05; ** p < 0.01; *** p < 0.001 from controls. Note: The upper and lower range of the standard deviation are indicated in each box with a horizontal line with dots as outliers. The interquartile range is Q3-Q1 (IQ), the maximum range is Q3 + 1.5 * IQ, and the minimum range is Q1 − 1.5 * IQ, which generate the area between the minimum and maximum; a line inside the boxplot shows the median. The group names are plotted on the X-axis.

Figure 3.
Microbial community composition. (A) Partial least-square-discriminant analysis (PLS-DA) of sample distribution (blue squares represent the control, red circles represent NNV, green triangles represent GIV and black dots represent Vibrio). (B) Venn diagram representing OTU richness and overlap of microbial communities in the control, GIV, Vibrio and NNV groups (red), which are represented by blue, purple, yellow and red color, respectively. The total OTU richness was 6372 OTUs among the four groups.
Figure 3.
Microbial community composition. (A) Partial least-square-discriminant analysis (PLS-DA) of sample distribution (blue squares represent the control, red circles represent NNV, green triangles represent GIV and black dots represent Vibrio). (B) Venn diagram representing OTU richness and overlap of microbial communities in the control, GIV, Vibrio and NNV groups (red), which are represented by blue, purple, yellow and red color, respectively. The total OTU richness was 6372 OTUs among the four groups.

Figure 4.
Species abundance distribution displayed the top ten most abundant at the (A) phylum and (B) genus levels. The distribution was visualized as microbiomes with different relative abundances at classification levels and their relative abundance proportions. Typically, the data intuitively distinguished the differences in the abundance of bacteria between healthy and diseased grouper. The X-axis shows the group names; the Y-axis shows the percentage of relative abundance.
Figure 4.
Species abundance distribution displayed the top ten most abundant at the (A) phylum and (B) genus levels. The distribution was visualized as microbiomes with different relative abundances at classification levels and their relative abundance proportions. Typically, the data intuitively distinguished the differences in the abundance of bacteria between healthy and diseased grouper. The X-axis shows the group names; the Y-axis shows the percentage of relative abundance.

Figure 5.
Bacterial abundance heatmap at the genus level was established for the control, nervous necrosis virus (NNV), grouper iridovirus (GIV) and Vibrio groups. The blue color indicates that the abundance level was less than the mean level, while the red color indicates that the abundance level was higher than the mean level.
Figure 5.
Bacterial abundance heatmap at the genus level was established for the control, nervous necrosis virus (NNV), grouper iridovirus (GIV) and Vibrio groups. The blue color indicates that the abundance level was less than the mean level, while the red color indicates that the abundance level was higher than the mean level.

Figure 6.
Comparison of uninfected and different disease grouper intestinal microbiota to characterize differential abundances between the two groups. Note: The left chart shows the mean abundance level and ratio between two groups, while the right chart shows the 95% confidence intervals. The probability value (p value < 0.05) is plotted on the right-hand side.
Figure 6.
Comparison of uninfected and different disease grouper intestinal microbiota to characterize differential abundances between the two groups. Note: The left chart shows the mean abundance level and ratio between two groups, while the right chart shows the 95% confidence intervals. The probability value (p value < 0.05) is plotted on the right-hand side.

Figure 7.
MetagenomeSeq annotated the species with significant differences (* p value < 0.05; ** p value < 0.01) among the groups to conduct an intensive study of community structure differences. Top row, genus Sphingomonas, Atopostipes, Staphylococcus; Bottom row, genus Mongoliitalea, Acinetobacter and Ralstonia.
Figure 7.
MetagenomeSeq annotated the species with significant differences (* p value < 0.05; ** p value < 0.01) among the groups to conduct an intensive study of community structure differences. Top row, genus Sphingomonas, Atopostipes, Staphylococcus; Bottom row, genus Mongoliitalea, Acinetobacter and Ralstonia.

Figure 8.
Innate immune-related genes expression in the intestine and head kidney of E. coioides after infection with NNV, GIV and V. harveyi. (A) The expression levels of IFN-2, TNF-α, iL-6 and iL-1β in intestines. (B) The expression levels of IFN-2, TNF-α, iL-6 and iL-1β in head kidney. Adaptive immune-related genes expression in the intestine of E. coioides after infection with NNV, GIV and V. harveyi. (C)The different Ig isotopes (IgD, IgM and IgT) expression in the intestines of control group and pathogen infected (NNV, GIV and Vibrio group) group. The relative mRNA levels were normalized by β-actin. Data presented are presented as mean ± SD (n = 5) and the asterisk (*) represented significant difference at * p < 0.05; ** p < 0.01; *** p < 0.001 from controls.
Figure 8.
Innate immune-related genes expression in the intestine and head kidney of E. coioides after infection with NNV, GIV and V. harveyi. (A) The expression levels of IFN-2, TNF-α, iL-6 and iL-1β in intestines. (B) The expression levels of IFN-2, TNF-α, iL-6 and iL-1β in head kidney. Adaptive immune-related genes expression in the intestine of E. coioides after infection with NNV, GIV and V. harveyi. (C)The different Ig isotopes (IgD, IgM and IgT) expression in the intestines of control group and pathogen infected (NNV, GIV and Vibrio group) group. The relative mRNA levels were normalized by β-actin. Data presented are presented as mean ± SD (n = 5) and the asterisk (*) represented significant difference at * p < 0.05; ** p < 0.01; *** p < 0.001 from controls.


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xiao Joe, J.T.; Tseng, Y.-C.; Wu, J.-L.; Lu, M.-W. The Alteration of Intestinal Microbiota Profile and Immune Response in Epinephelus coioides during Pathogen Infection. Life 2021, 11, 99. https://doi.org/10.3390/life11020099
AMA Style
Xiao Joe JT, Tseng Y-C, Wu J-L, Lu M-W. The Alteration of Intestinal Microbiota Profile and Immune Response in Epinephelus coioides during Pathogen Infection. Life. 2021; 11(2):99. https://doi.org/10.3390/life11020099
Chicago/Turabian StyleXiao Joe, Joan Tang, Yung-Che Tseng, Jen-Leih Wu, and Ming-Wei Lu. 2021. "The Alteration of Intestinal Microbiota Profile and Immune Response in Epinephelus coioides during Pathogen Infection" Life 11, no. 2: 99. https://doi.org/10.3390/life11020099
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.