Bioinformatics Analysis of Evolution and Human Disease Related Transposable Element-Derived microRNAs


1
National Primate Research Center, Korea Research Institute of Bioscience and Biotechnology, Cheongju 28116, Korea
2
Department of Functional Genomics, KRIBB School of Bioscience, Korea University of Science and Technology (UST), Daejeon 34113, Korea
3
Department of Biological Sciences, College of Natural Sciences, Pusan National University, Busan 46241, Korea
4
Institute of Systems Biology, Pusan National University, Busan 46241, Korea
*
Author to whom correspondence should be addressed.
Life 2020, 10(6), 95; https://doi.org/10.3390/life10060095
Submission received: 8 June 2020
/
Revised: 22 June 2020
/
Accepted: 23 June 2020
/
Published: 25 June 2020
(This article belongs to the Special Issue Genomic Impact of Transposable Elements)
Abstract
:Transposable element (TE) has the ability to insert into certain parts of the genome, and due to this event, it is possible for TEs to generate new factors and one of these factors are microRNAs (miRNA). miRNAs are non-coding RNAs made up of 19 to 24 nucleotides and numerous miRNAs are derived from TE. In this study, to support general knowledge on TE and miRNAs derived from TE, several bioinformatics tools and databases were used to analyze miRNAs derived from TE in two aspects: evolution and human disease. The distribution of TEs in diverse species presents that almost half of the genome is covered with TE in mammalians and less than a half in other vertebrates and invertebrates. Based on selected evolution-related miRNAs studies, a total of 51 miRNAs derived from TE were found and analyzed. For the human disease-related miRNAs, total of 34 miRNAs derived from TE were organized from the previous studies. In summary, abundant miRNAs derived from TE are found, however, the function of miRNAs derived from TE is not informed either. Therefore, this study provides theoretical understanding of miRNAs derived from TE by using various bioinformatics tools.
1. Introduction
In 1986, transposable elements (TEs) were found from Drosophila melanogaster that have the ability to adjust their position and replicate in host genome that categorizes into two large classes with different mechanisms [1]. TEs are abundant in the genome of eukaryotes, which is about approximately from 3% up to 85% in the genome of plants and 50% in mammalian’s genome. The discovery on specific function of TE is still ongoing assignments for researchers, however, numerous studies revealed that TE is involved in epigenetics [2,3,4], evolution [5,6,7,8,9], and disease [10,11,12]. An interesting fact about class I TE called retrotransposons and evolution is that superfamily of retrotransposon long interspersed nuclear element 1 (LINE1 or L1), short interspersed nuclear element (SINE)-variable number of tandem repeat (VNTR)-Alu (SVA), and human endogenous retrovirus K (HERV-K) are active TEs in recent human evolution [13,14,15,16,17]. As mentioned earlier, not only in evolution, TE has very close relation with human cancer and disease [18,19,20,21,22,23,24,25,26,27,28,29]. For instance, L1 encodes RNA pol II promoter and this L1 promoter is hypomethylated in tumors of lung, colon, and several types of cancers [19]. TEs merge into the host genome, and this event has the possibility to provide and create new sequences to gene expression regulators such as microRNAs (miRNA) [4,30,31,32,33,34,35,36,37,38] and transcription factors (TF) [39,40,41].
In 1993, miRNA was first found in Caenorhabditis elegans (C. elegans) and it is comprised with 19 to 24 nucleotides of non-coding RNAs which is associated with gene regulations by targeting mRNAs for cleavage or translational inhibition [42,43,44,45]. The essential function of miRNA is correlated with oncogenesis, immunity, developments, and cell differentiations. Generally, 3′ untranslated region (UTR) is the miRNA binding sites of target mRNA. miRNA recognizes the complementary binding sites of target gene for its seed regions which is approximately 6 nucleotides long in miRNA. Previously, numerous studies have provided the evidence of identification on miRNA derived from TE [30,31,33,34,36] and some of the miRNAs derived from TE had a strong correlation with human disease [46,47,48,49,50,51,52] as well as evolution [53,54,55,56,57]. Considering the many substantial aspects of TE, miRNAs derived from TE mimic the functions of TE.
Bioinformatics tools are useful for the initial steps before starting the experiments, which is what understanding the primary information on what the study will be about. In the case of TE, analyzing the sequences and predicting structure of TE is important considering the function of TE. A TE-based database called ‘Repeatmasker’ provides the proportion of each type of TEs among diverse species [15]. Additionally, there are more of TE related bioinformatics tools and databases that determines which TE has merged into the target sequences, however, TE-based databases and programs are still limited [58,59,60,61]. In contrast with TE, numerous miRNA-based databases provide basic information about miRNA, miRNA related cancer, target genes, TFs, and so on [60,62,63,64,65,66,67].
In this study, evolution as well as human disease-related miRNAs derived from TE were examined through published research papers. Those determined miRNAs derived from TE were analyzed by using several bioinformatics tools to provide fundamental information of miRNAs derived from TE.
2. Bioinformatics Analysis of Transposable Elements
The distribution of TEs in various species (human, chimpanzee, gorilla, orangutan, gibbon, macaque, rhesus, marmoset, mouse, horse, cow, cat, dog, chicken, zebrafish, and C. elegans) were verified by RepeatMasker Genomic Datasets [15]. The species that are examined for whole genome sequencing are listed in both UCSC genome browser and RepeatMasker [15,60]. Table 1 shows the percentage of which TE is or not include in the genome. After the name of the superfamily element such as SINE, LINE, and long terminal repeats (LTR), the specific name of the element is given. The ‘other’ after the superfamily elements are representing the unspecified elements. The primates, including humans and the other seven of the species, are reasonably chosen to present the percentage of each TEs for evolutionary aspects.
Chicken had the lowest percentage of TEs in the genome (9.3%), and orangutan had the highest percentage of TEs in the genome (48.5%). From primates to mammalians (human, chimpanzee, gorilla, orangutan, gibbon, macaque, rhesus, marmoset, mouse, horse, cow, cat, and dog), variation of TE is well spread out in each species genome, excluding SVA element. SVA element was exclusive in humans. C. elegans is another species with the lowest percentage of TE in the genome (9%) and few of the TEs were included (LINE-CR1, LTE-other, DNA-TcMar, DNA-hAT, and DNA-other). Zebrafish is representing species of fish in this table and zebrafish contains the highest percentage of DNA transposons-other (19.6%).
3. Selection of Microrna Related Papers and Bioinformatic Analyses of Transposable Element-Derived microRNAs
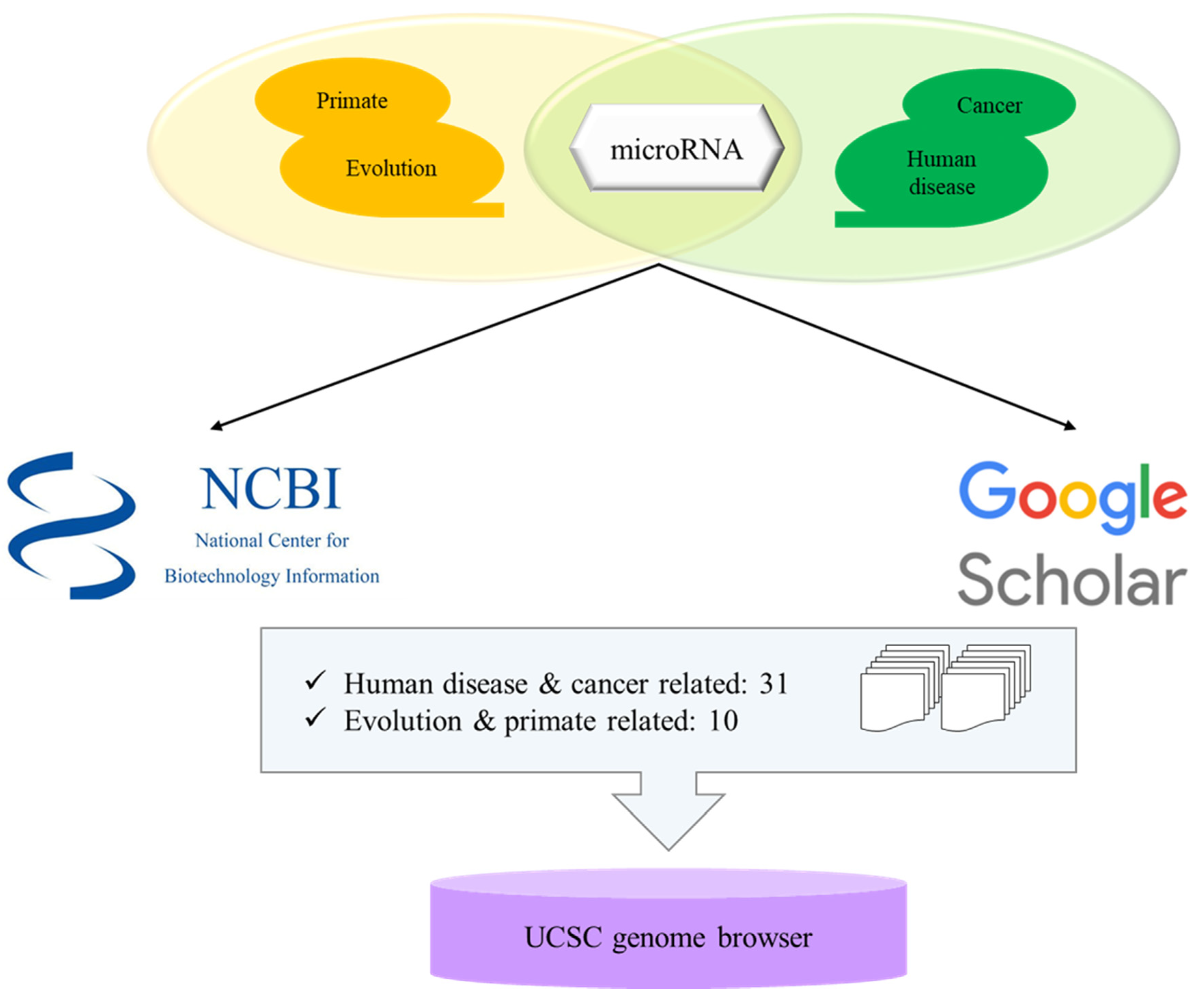
miRNAs related with keywords of ‘evolution and primates’ and ‘human disease and cancer’ were searched from National Center for Biotechnology Information (NCBI)-PubMed database [68] and google scholar [69] (Figure 1). Each paper contained numerous miRNAs and the information of miRNAs were examined from miRbase v22.1 (http://www.mirbase.org) [66]. Then each miRNA was localized in human genome (GRCh38) by UCSC Genome Browser (http://genome.ucsc.edu) [60].
To determine miRNAs derived from TE from human disease and cancer and evolution and primate-related miRNAs, a total of 41 papers were selected from NCBI-PubMed and google scholar with 31 studies on human disease and cancer and 10 studies on evolution and primate-related miRNAs. MiRNAs derived from TE are fully and partially derived from TE, and some of the miRNAs derived from TE share more than one TEs in the sequence.
4. Bioinformatic Analyses of Evolution Related Transposable Element-Derived microRNAs
The evolution and primate related miRNAs derived from TE from 10 studies were then localized in UCSC genome browser to check the location in the human genome and which type of TE that miRNAs are derived from (Table 2). From a total of 51 miRNAs derived from TE related with evolution and primates, 16 miRNAs were derived from LINE family, 19 from SINE, 3 from LTR, and 15 miRNAs were derived from DNA transposon. Ten of the miRNAs derived from TE were derived from more than one TEs, and interestingly, hsa-miR-548a-2 and hsa-miR-619 are derived from different types of TEs. For instance, hsa-miR-548a-2 is derived from two LTR16A2 at the terminal of miRNA and one DNA transposon MADE1 is in the middle and hsa-miR-619 has one of each L1MC4 and AluSz6.
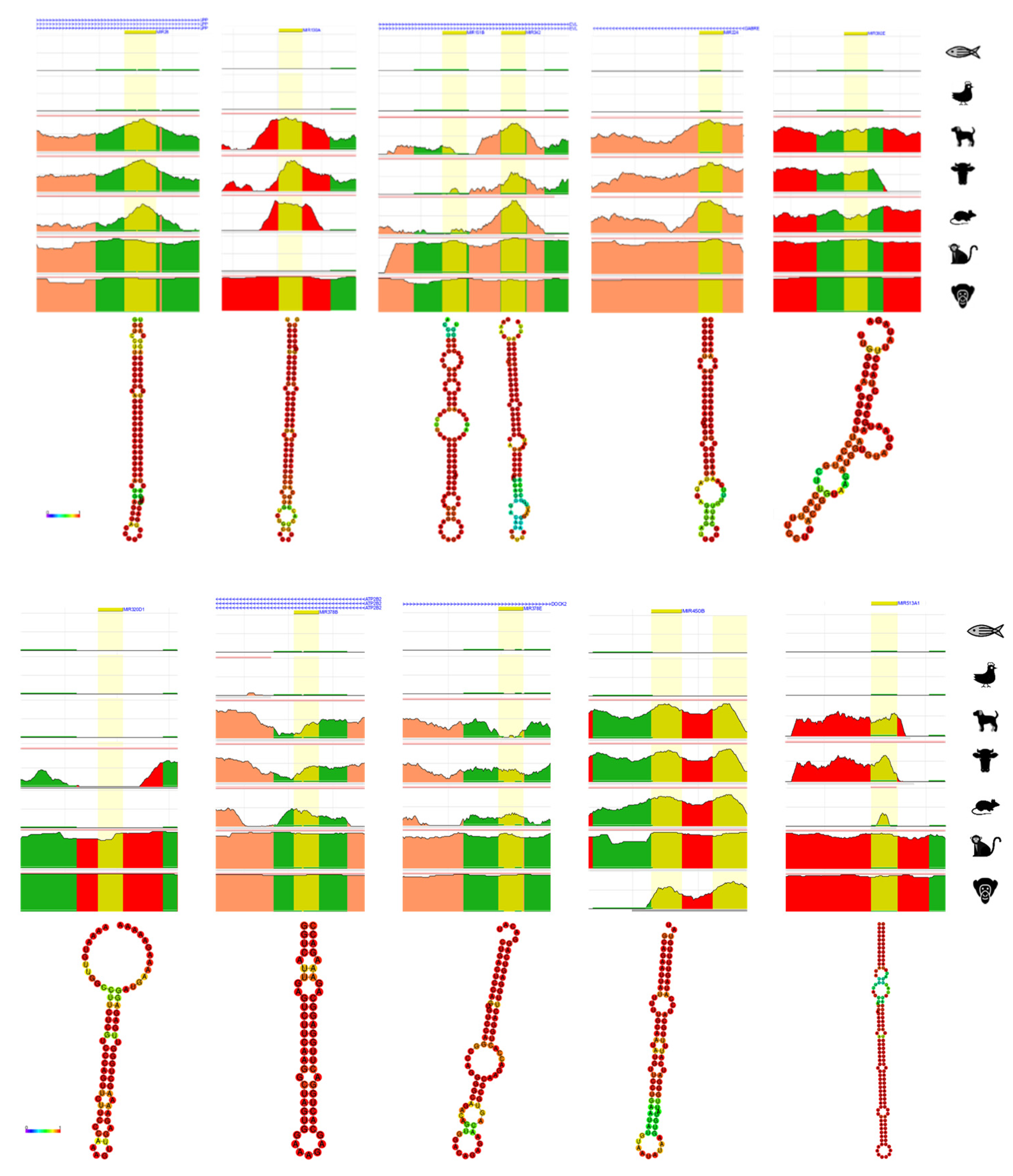
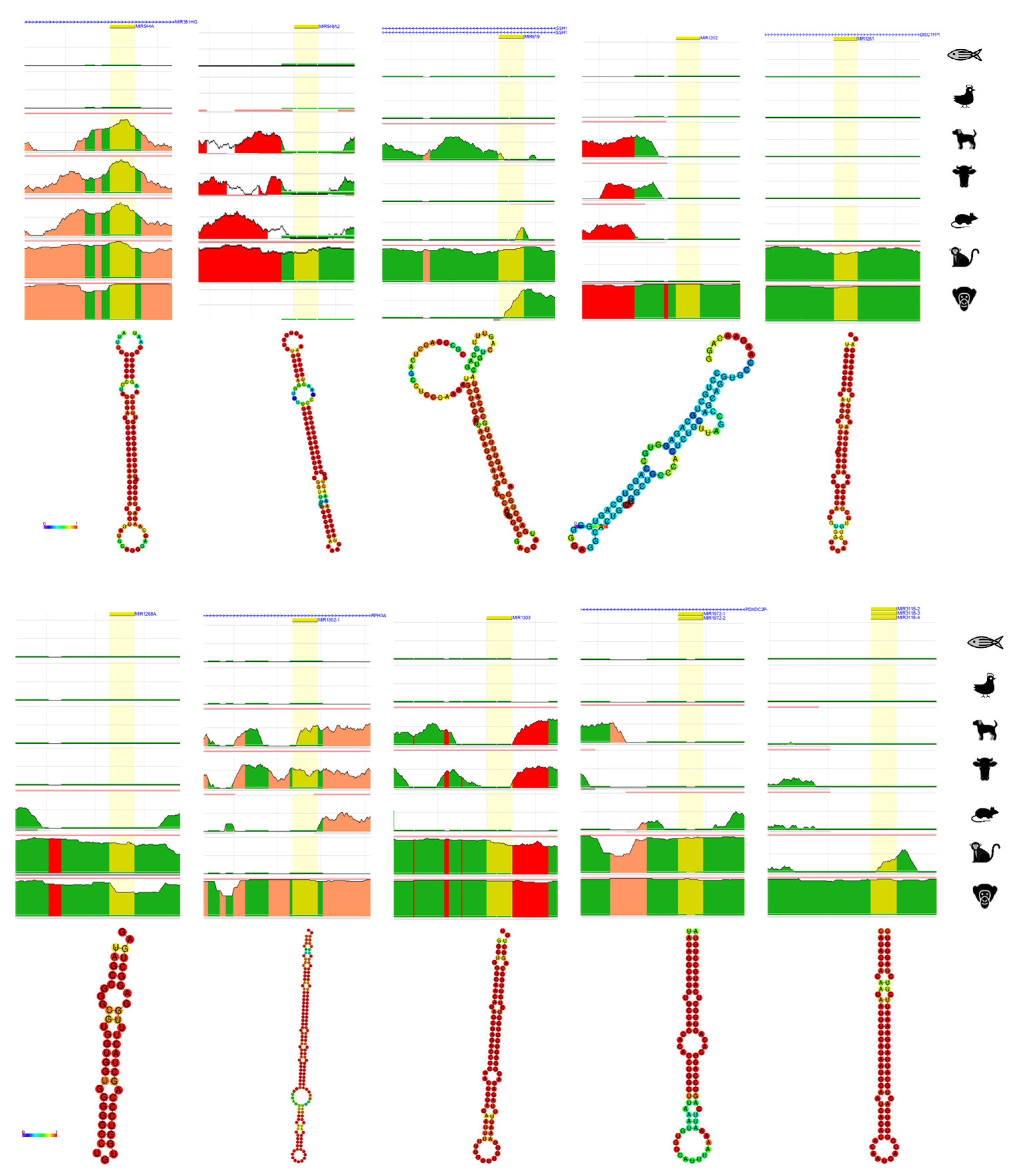
From a total of 51 evolution and primate-related miRNAs derived from TE, 21 of miRNAs derived from TE with different type of TEs were chosen for further bioinformatics analysis. The evolution and primate related miRNAs derived from TE were analyzed by ECR browser to briefly check the conservation in chimpanzee, rhesus, mouse, cow, dog, chicken, and zebrafish [72]. Additionally, the structure of each 21 miRNAs derived from TE were predicted by RNAfold webserver which generates the structure of minimum free energy (MFE) contributed by secondary structure of RNA sequences [73]. The strong base-pairing probability shows in color red with value close to 1 and weak base-pairing probability shows in color blue with value close to 0.
From the numerically ordered list of evolution and primate related miRNAs derived from TE, one of each miRNA was selected from miRNAs derived from the same TE family (hsa-miRNA-28, -130a, -151b, -224, -302e, -320d-1, -342, -378b, -378e, -450b, -513a-1, -544a, -548a-2, -619, -1202, -1261, -1268a, -1302-1, -1303, -1972-1, -3118-3) to examine the conservation throughout human, chimpanzee, rhesus, mouse, cow, dog, chicken and zebrafish (Figure 2). Amongst the 21 of analyzed evolution and primate related miRNAs derived from TE, hsa-miRNA-151b (chr14: 100,109,419-100,109,514) and -342 (chr14: 100,109,655-100,109,753) are located near each other, however, they do not share the same TE family. All 21 of miRNAs derived from TE were not conserved from chicken and zebrafish and hsa-miRNA-28, -224, and -544a was conserved from chimpanzee, rhesus, mouse, cow and dog, hsa-miRNA-342, -302e, -378b, and -378e was conserved in primates but partially conserved from mouse, cow and dog. Hsa-miRNA-1202 and -3118-3 showed conservation only in chimpanzee and hsa-miRNA-548a-2 and -619 showed conservation only in rhesus monkey. Hsa-miRNA-320d-1, -1261, 1268a, -1302-1, -1303 and -1972-1 shows conservation in primates. Strangely, most of miRNAs derived from TE were well conserved in chimpanzee, however, hsa-miRNA-548a-2 was not conserved in chimpanzee, hsa-miRNA-450b and -619 was partially conserved in chimpanzee, and hsa-miRNA-130a, -1202, -1302-1, and -3118-3 was not conserved in rhesus monkey. Hsa-miRNA-513a-1 was not found in mouse genome.
The secondary structure of 21 of evolution and primate related miRNAs derived from TE were predicted by RNAfold webserver [73]. Almost all the MFE structure of miRNAs derived from TE had strong base-pairing MFE values, with the exception of hsa-miRNA-1202 which shows weakest MFE structure.
5. Bioinformatic Analyses of Human Diseases Related Transposable Element-Derived microRNAs
The human disease and cancer related miRNAs derived from TE from 31 studies were then localized in the UCSC genome browser to check the location in the human genome and which type of TE that miRNAs are derived from (Table 3). As mentioned previously, miRNAs derived from TE are not only derived from one TE, however, it could be derived from more than one TE with different families. From a total of 34 human diseases and cancer-related miRNAs derived from TE, 16 miRNAs were derived from LINE, 6 from SINE, 2 from LTR, and 10 miRNAs were derived from DNA transposons. Unlike evolution and primate related miRNAs derived from TE which share two types of TEs in one miRNA, no more than one TE type share miRNA from human disease and cancer related miRNAs but 7 out of 34 miRNAs derived from TE share more than one same TE family shares miRNAs derived from TE.
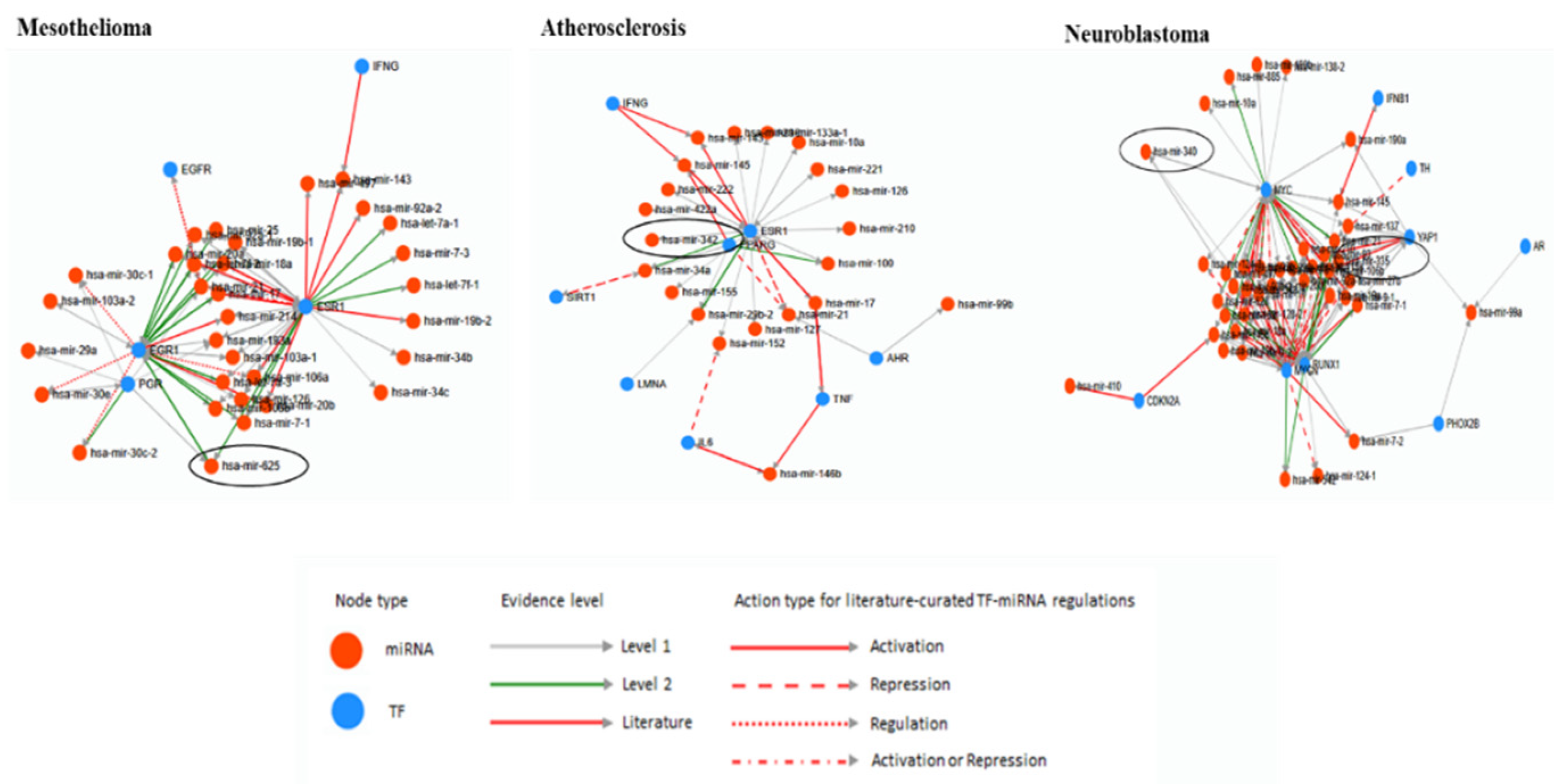
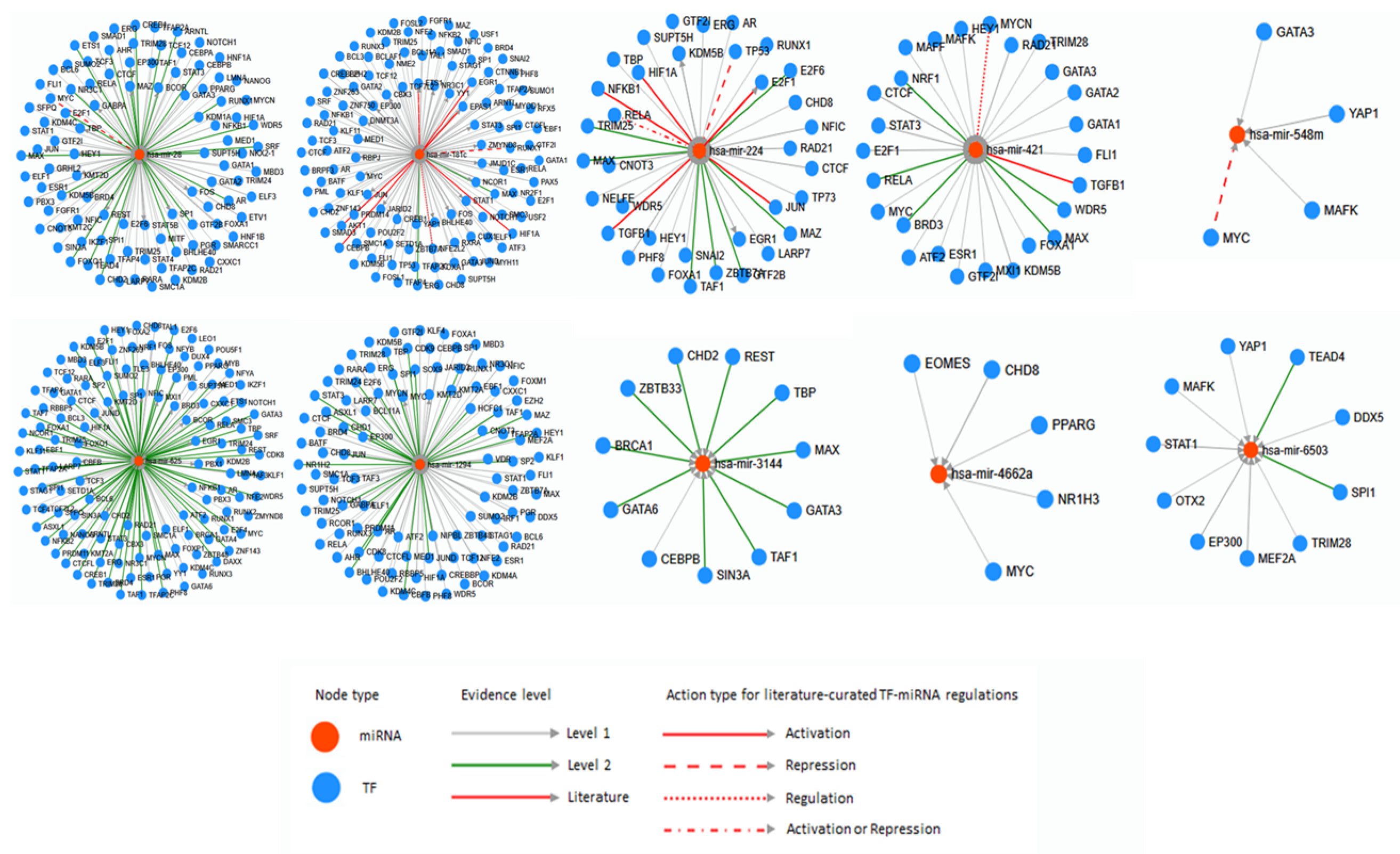
The database TransmiR v2.0 was used to predict the correlation of miRNAs and TFs [67]. From all the list of the human disease uploaded in TransmiR database, three diseases Mesothelioma, Atherosclerosis and Neuroblastoma related miRNAs and TF were found (Figure 3). Hsa-miRNA-625 is one of the Mesothelioma-related miRNA and it is activated by EGR1, PGR, and ESR1 TFs. The evidence level of EGR1 and ESR1 is level 2 and it is stricter than level 1 TF PGR. Atherosclerosis related miRNAs derived from TE is hsa-miRNA-342 and it is activated by ESR1 in level of 1. Two miRNAs derived from TE, hsa-miRNA-335 and -340 were related Neuroblastoma. The literature level evidence provided TF MYCN represses hsa-miRNA-335, and YAP1, RUNX1, and MYC activate hsa-miRNA-335 by level of 1. MYCN, RUNX1, and MYC activate hsa-miRNA-340 by level of 1.
From the list of human disease and cancer-related miRNAs derived from TE, numerically one of each miRNA was selected (hsa-miRNA-28, -181c, -224, -421, -548m, -625, -1294, -3144, -4662a, and -6503) to analyze by TransmiR v2.0. After all, it also provides correlation between numerous miRNA and TFs (Figure 4). Hsa-miRNA-28 is activated by various TFs, however, MYC represses hsa-miRNA-28 exclusively. Mostly, hsa-miRNA-28 was activated or repressed by TF and similarly, hsa-miRNA-181c was activated by many TFs. On the other hand, hsa-miRNA-181c was also activating abundant TFs. Additionally, RUNX1 and ETS1 represses and NFE2L2 regulates hsa-miRNA-181c. Hsa-miRNA-224 was activated by many TFs and it activates E2F1, EGR1, and KDM5B. TP53 represses and RELA activates or represses hsa-miRNA-224. Hsa-miRNA-421 was activated by most of related TFs, however MYCN regulates hsa-miRNA-421. Hsa-miRNA-548m is activated by GATA3, YAP1, and MAFK however, MYC represses hsa-miRNA-548m. Hsa-miRNA-625, -1294 is activated by abundant TFs and hsa-miRNA-625 activates several TFs. Hsa-miRNA-1294 also activates KMT2D, EP300, and MYC. Hsa-miRNA-3144 is activated by CHD2, SIN3A, GATA6, REST, TAF1, BRCA1, GATA3, MAX, ZBTB33, TBP, and CEBPB and hsa-miRNA-4662a is activated by CHD8, EOMES, MYC, NR1H3, and PPARG. Lastly, MEF2A, TEAD4, OTX2, MAFK, EP300, SPI1, STAT1, TRIM28, YAP1, and DDX5 activates hsa-miRNA-6503.
6. Discussion
Bioinformatics tools are useful and important when not much information is provided or studied for the target subjects. Numerous bioinformatics tools are provided online, and are ready to be used right away or downloaded. There are several bioinformatics tools of miRNAs, however, TE related bioinformatics tools are still insufficient. By using bioinformatics database related with TE, the distribution of TE has been modified (Table 1) [15]. The distribution of TE is highly scattered in the genome of most of the species. In the evolutionary aspects on distribution of TE, SVA element is exclusive in humans only. Alu element from SINE is a primate and mouse specific element excluding few mammalians (horse, cow, cat, and dog), chicken, zebrafish, and C. elegans. From LINE, the proportion of CR1 element is very low amongst all of mammalians, fish, and C. elegans, however, one study provided the evidence that CR1 element is moderately scattered in avian, crocodilian, turtle, and lepidosaurian, also known as diapsid reptiles [84]. The distribution of ERVK from LTR element presents in primates until rhesus monkey, mouse and cow amongst the mammalians. Most of the ERVK studies are performed in primates, thus mouse shows highest of percentages of ERVK element among all the species, and one study mentioned that human and mouse contain numerous LTR-derived TFBS which contributes in other TFs to bind, and they did not mention the reason why mouse has a high percentage of ERVK element, yet it might be due to embryonic stem cells of mice [85,86,87].
Approximately half of the genome is covered in TEs for mammalians and zebrafish, and over 10 percent for chicken and C. elegans and these highly distributed TEs are capable of generating miRNAs and TFBSs [30,31,34,87]. Based on the miRNA studies, miRNAs derived from TE were filtered into two types, primate and evolution and human disease and cancer. Table 2 shows 51 of primate and evolution related miRNAs derived from TE and Table 3 shows 34 of human disease and cancer-related miRNAs derived from TE. First, to analyze miRNAs derived from TE related in primates and evolution, ECR browser was used to check the conservation on few of selected miRNAs derived from TE. The conservation is influential to miRNAs derived from TE related in primates and evolution due to selecting the target species or samples before going into the actual experiments. Previous studies checked the conservation of each target miRNAs they found to applicate them on primate and evolution related miRNAs [54,70,88]. ECR browsers are used to predict the conservation of the target gene, miRNA, or the specific region of the genome. The conservation on few of the selected primate and evolution-related miRNAs derived from TE show conservation well until mammalians, however, few of miRNAs derived from TE are not randomly conserved (Figure 2). To examine the conservation precisely, the sequence of each miRNA is needed to be downloaded from each species. TargetScan database provides the sequences of conserved miRNAs in the target genes and this method is more accurate than the prediction from ECR browser [89,90]. The RNAfold result predicts the strongness of base pairing as well as the MFE value of the miRNAs by the colors. The miRNA with the weakest structure is predicted as miRNA-1202.
The human disease and cancer related miRNAs derived from TE were analyzed with TFs. TransmiR database provides the information on TFBS that regulates or correlates with miRNAs. First, the examination of all 34 human disease and cancer related miRNAs derived from TE from Table 3 were analyzed to check the correlation between miRNAs derived from TE and TFs by human disease and cancer provided from TransmiR database (Figure 3). Four miRNAs derived from TE were found from three human disease and cancers provided from TransmiR. Other miRNA and TF studies used TransmiR to predict which TFs that their target miRNA is targeting or correlate together and applicate them on further bioinformatics analysis or experiments [91,92]. In addition, few of TFs were determined on human disease and cancer related miRNAs derived from TE. As shown in Figure 4, some miRNAs derived from TE interact with numerous TFs and on the other hand, some miRNAs derived from TE interact with few TFs. The study of miRNA-548m and MYC supported the data of TransmiR based on the result of stroma-inducing miRNA-549m inhibition leads to the c-Myc overexpression [93]. By using TransmiR databases, the hypothesis was suggested that enhancer activity of miRNAs derived from TE is increased by TFs, and the report actually mentioned that the enhancer activity of miRNAs derived from TE OF-miRNA-307 might induced by the TFs near OF-miRNA-307 binds in 3′UTR of target gene [94].
The aim of this study was to introduce the basic bioinformatics tools used for TE and miRNAs derived from TE studies. The evolution and human disease-related miRNAs derived from TE were identified by published papers and they were analyzed with bioinformatics tools. Abundant miRNAs were derived from TEs and they have a close relation with primate and evolution and human disease and cancer. Here, fundamental information of miRNAs derived from TE by using several of the bioinformatics tools are analyzed.
Author Contributions
H.-E.L. designed and wrote the manuscript, J.-W.H. commented on the manuscript and H.-S.K. acquires funding. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Research Foundation of Korea (NRF), grant number (NRF-2018R1D1A1B07049460).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Finnegan, D.J.; Fawcett, D.H. Transposable elements in Drosophila melanogaster. Oxf. Surv. Eukaryot. Genes 1986, 3, 1–62. [Google Scholar] [PubMed]
- Hollister, J.D.; Gaut, B.S. Epigenetic silencing of transposable elements: A trade-off between reduced transposition and deleterious effects on neighboring gene expression. Genome Res. 2009, 19, 1419–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendizabal, I.; Keller, T.E.; Zeng, J.; Yi, S.V. Epigenetics and evolution. Integr. Comp. Biol. 2014, 54, 31–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lisch, D.; Bennetzen, J.L. Transposable element origins of epigenetic gene regulation. Curr. Opin. Plant. Biol. 2011, 14, 156–161. [Google Scholar] [CrossRef]
- Lee, H.-E.; Eo, J.; Kim, H.-S. Composition and evolutionary importance of transposable elements in humans and primates. Genes Genom. 2014, 37, 135–140. [Google Scholar] [CrossRef]
- Feschotte, C.; Pritham, E.J. DNA transposons and the evolution of eukaryotic genomes. In Annual Review of Genetics; Annual Reviews: Palo Alto, CA, USA, 2007; Volume 41, pp. 331–368. [Google Scholar]
- Finnegan, D.J. Eukaryotic transposable elements and genome evolution. Trends Genet. 1989, 5, 103–107. [Google Scholar] [CrossRef]
- Biemont, C.; Vieira, C. Genetics: Junk DNA as an evolutionary force. Nature 2006, 443, 521–524. [Google Scholar] [CrossRef]
- Deininger, P.L.; Moran, J.V.; Batzer, M.A.; Kazazian, H.H., Jr. Mobile elements and mammalian genome evolution. Curr. Opin. Genet. Dev. 2003, 13, 651–658. [Google Scholar] [CrossRef]
- Callinan, P.A.; Batzer, M.A. Retrotransposable elements and human disease. Genome Dyn. 2006, 1, 104–115. [Google Scholar] [CrossRef] [Green Version]
- Bourque, G.; Burns, K.H.; Gehring, M.; Gorbunova, V.; Seluanov, A.; Hammell, M.; Imbeault, M.; Izsvak, Z.; Levin, H.L.; Macfarlan, T.S.; et al. Ten things you should know about transposable elements. Genome Biol. 2018, 19. [Google Scholar] [CrossRef]
- Ayarpadikannan, S.; Lee, H.E.; Han, K.; Kim, H.S. Transposable element-driven transcript diversification and its relevance to genetic disorders. Gene 2015, 558, 187–194. [Google Scholar] [CrossRef]
- Kazazian, H.H., Jr.; Moran, J.V. The impact of L1 retrotransposons on the human genome. Nat. Genet. 1998, 19, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Cordaux, R.; Batzer, M.A. The impact of retrotransposons on human genome evolution. Nat. Rev. Genet. 2009, 10, 691–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, A.; Hubley, R.; Green, P. RepeatMasker. Available online: http://repeatmasker.org (accessed on 1 October 2019).
- Wang, H.; Xing, J.; Grover, D.; Hedges, D.J.; Han, K.; Walker, J.A.; Batzer, M.A. SVA elements: A hominid-specific retroposon family. J. Mol. Biol. 2005, 354, 994–1007. [Google Scholar] [CrossRef] [PubMed]
- Mills, R.E.; Bennett, E.A.; Iskow, R.C.; Devine, S.E. Which transposable elements are active in the human genome? Trends Genet. 2007, 23, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.S.; Shah, N.M.; Du, A.Y.; Dailey, Z.Z.; Pehrsson, E.C.; Godoy, P.M.; Zhang, D.; Li, D.; Xing, X.; Kim, S.; et al. Transposable elements drive widespread expression of oncogenes in human cancers. Nat. Genet. 2019, 51, 611–617. [Google Scholar] [CrossRef]
- Burns, K.H. Transposable elements in cancer. Nat. Rev. Cancer 2017, 17, 415–424. [Google Scholar] [CrossRef]
- Anwar, S.L.; Wulaningsih, W.; Lehmann, U. Transposable Elements in Human Cancer: Causes and Consequences of Deregulation. Int. J. Mol. Sci 2017, 18, 974. [Google Scholar] [CrossRef] [Green Version]
- Transposable Elements Regulate Oncogene Expression in Human Cancers. Cancer Discov. 2019, 9, 689. [CrossRef] [Green Version]
- Babaian, A.; Mager, D.L. Endogenous retroviral promoter exaptation in human cancer. Mob. DNA 2016, 7, 24. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.J.; Jung, Y.D.; Kim, T.O.; Kim, H.S. Alu-related transcript of TJP2 gene as a marker for colorectal cancer. Gene 2013, 524, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Lee, J.; Km, H.S.; Yang, K.; Yi, J.M. DNA methylation of mobile genetic elements in human cancers. Genes Genom. 2013, 35, 265–271. [Google Scholar] [CrossRef]
- Lee, S.H.; Kang, Y.J.; Jo, J.O.; Ock, M.S.; Baek, K.W.; Eo, J.; Lee, W.J.; Choi, Y.H.; Kim, W.J.; Leem, S.H.; et al. Elevation of human ERV3-1 env protein expression in colorectal cancer. J. Clin. Pathol. 2014, 67, 840–844. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.R.; Jung, Y.D.; Kim, Y.J.; Lee, H.E.; Jeong, H.; Kim, H.S. Identification of L1ASP-derived chimeric transcripts in lung cancer. Genes Genom. 2014, 36, 853–859. [Google Scholar] [CrossRef]
- Yi, J.M.; Kim, H.M.; Kim, H.S. Human endogenous retrovirus HERV-H family in human tissues and cancer cells: Expression, identification, and phylogeny. Cancer Lett. 2006, 231, 228–239. [Google Scholar] [CrossRef]
- Sin, H.S.; Huh, J.W.; Kim, D.S.; Kang, D.W.; Min, D.S.; Kim, T.H.; Ha, H.S.; Kim, H.H.; Lee, S.Y.; Kim, H.S. Transcriptional control of the HERV-H LTR element of the GSDML gene in human tissues and cancer cells. Arch. Virol. 2006, 151, 1985–1994. [Google Scholar] [CrossRef]
- Yi, J.M.; Schuebel, K.; Kim, H.S. Molecular genetic analyses of human endogenous retroviral elements belonging to the HERV-P family in primates, human tissues, and cancer cells. Genomics 2007, 89, 1–9. [Google Scholar] [CrossRef]
- Roberts, J.T.; Cardin, S.E.; Borchert, G.M. Burgeoning evidence indicates that microRNAs were initially formed from transposable element sequences. Mob. Genet. Elem. 2014, 4, e29255. [Google Scholar] [CrossRef]
- Piriyapongsa, J.; Jordan, I.K. A family of human microRNA genes from miniature inverted-repeat transposable elements. PLoS ONE 2007, 2, e203. [Google Scholar] [CrossRef]
- Ugolini, I.; Halic, M. Fidelity in RNA-based recognition of transposable elements. Philos. Trans. R Soc. Lond B Biol. Sci. 2018, 373. [Google Scholar] [CrossRef] [Green Version]
- Smalheiser, N.R.; Torvik, V.I. Mammalian microRNAs derived from genomic repeats. Trends Genet. 2005, 21, 322–326. [Google Scholar] [CrossRef]
- Piriyapongsa, J.; Marino-Ramirez, L.; Jordan, I.K. Origin and evolution of human microRNAs from transposable elements. Genetics 2007, 176, 1323–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lippman, Z.; Gendrel, A.V.; Black, M.; Vaughn, M.W.; Dedhia, N.; McCombie, W.R.; Lavine, K.; Mittal, V.; May, B.; Kasschau, K.D.; et al. Role of transposable elements in heterochromatin and epigenetic control. Nature 2004, 430, 471–476. [Google Scholar] [CrossRef]
- Qin, S.; Jin, P.; Zhou, X.; Chen, L.; Ma, F. The Role of Transposable Elements in the Origin and Evolution of MicroRNAs in Human. PLoS ONE 2015, 10, e0131365. [Google Scholar] [CrossRef]
- McCue, A.D.; Slotkin, R.K. Transposable element small RNAs as regulators of gene expression. Trends Genet. 2012, 28, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Feschotte, C. Transposable elements and the evolution of regulatory networks. Nat. Rev. Genet. 2008, 9, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Lambert, S.A.; Jolma, A.; Campitelli, L.F.; Das, P.K.; Yin, Y.; Albu, M.; Chen, X.; Taipale, J.; Hughes, T.R.; Weirauch, M.T. The Human Transcription Factors. Cell 2018, 172, 650–665. [Google Scholar] [CrossRef] [PubMed]
- Zemojtel, T.; Vingron, M. P53 binding sites in transposons. Front. Genet. 2012, 3, 40. [Google Scholar] [CrossRef] [Green Version]
- Kunarso, G.; Chia, N.Y.; Jeyakani, J.; Hwang, C.; Lu, X.; Chan, Y.S.; Ng, H.H.; Bourque, G. Transposable elements have rewired the core regulatory network of human embryonic stem cells. Nat. Genet. 2010, 42, 631–634. [Google Scholar] [CrossRef]
- Ambros, V.; Bartel, B.; Bartel, D.P.; Burge, C.B.; Carrington, J.C.; Chen, X.; Dreyfuss, G.; Eddy, S.R.; Griffiths-Jones, S.; Marshall, M.; et al. A uniform system for microRNA annotation. RNA 2003, 9, 277–279. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Kim, V.N. MicroRNA biogenesis: Coordinated cropping and dicing. Nat. Rev. Mol. Cell Biol. 2005, 6, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Yates, L.A.; Norbury, C.J.; Gilbert, R.J.C. The Long and Short of MicroRNA. Cell 2013, 153, 516–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, D.; Lee, H. Prioritizing cancer-related microRNAs by integrating microRNA and mRNA datasets. Sci. Rep. 2016, 6, 35350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulte, J.H.; Horn, S.; Otto, T.; Samans, B.; Heukamp, L.C.; Eilers, U.C.; Krause, M.; Astrahantseff, K.; Klein-Hitpass, L.; Buettner, R.; et al. MYCN regulates oncogenic MicroRNAs in neuroblastoma. Int. J. Cancer 2008, 122, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Dutta, A. MicroRNAs in cancer. Annu. Rev. Pathol. 2009, 4, 199–227. [Google Scholar] [CrossRef]
- Pinto, Y.; Buchumenski, I.; Levanon, E.Y.; Eisenberg, E. Human cancer tissues exhibit reduced A-to-I editing of miRNAs coupled with elevated editing of their targets. Nucleic Acids Res. 2018, 46, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Zhu, F.; Ding, Y. Differential microRNA expression profile in the plasma of preeclampsia and normal pregnancies. Exp. Ther. Med. 2019, 18, 826–832. [Google Scholar] [CrossRef]
- Murakami, Y.; Yasuda, T.; Saigo, K.; Urashima, T.; Toyoda, H.; Okanoue, T.; Shimotohno, K. Comprehensive analysis of microRNA expression patterns in hepatocellular carcinoma and non-tumorous tissues. Oncogene 2006, 25, 2537–2545. [Google Scholar] [CrossRef]
- Wang, H.; Peng, R.; Wang, J.; Qin, Z.; Xue, L. Circulating microRNAs as potential cancer biomarkers: The advantage and disadvantage. Clin. Epigenet. 2018, 10, 59. [Google Scholar] [CrossRef] [Green Version]
- McCreight, J.C.; Schneider, S.E.; Wilburn, D.B.; Swanson, W.J. Evolution of microRNA in primates. PLoS ONE 2017, 12, e0176596. [Google Scholar] [CrossRef] [PubMed]
- Guerra-Assuncao, J.A.; Enright, A.J. Large-scale analysis of microRNA evolution. BMC Genom. 2012, 13, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teferedegne, B.; Murata, H.; Quinones, M.; Peden, K.; Lewis, A.M. Patterns of microRNA expression in non-human primate cells correlate with neoplastic development in vitro. PLoS ONE 2010, 5, e14416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awan, H.M.; Shah, A.; Rashid, F.; Shan, G. Primate-specific Long Non-coding RNAs and MicroRNAs. Genom. Proteom. Bioinform. 2017, 15, 187–195. [Google Scholar] [CrossRef]
- Robles, A.I.; Harris, C.C. A primate-specific microRNA enters the lung cancer landscape. Proc. Natl. Acad. Sci. USA 2013, 110, 18748–18749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrusan, G.; Grundmann, N.; DeMester, L.; Makalowski, W. TEclass-a tool for automated classification of unknown eukaryotic transposable elements. Bioinformatics 2009, 25, 1329–1330. [Google Scholar] [CrossRef] [Green Version]
- Wicker, T.; Matthews, D.E.; Keller, B. TREP: A database for Triticeae repetitive elements. Trends Plant Sci. 2002, 7, 561–562. [Google Scholar] [CrossRef]
- Casper, J.; Zweig, A.S.; Villarreal, C.; Tyner, C.; Speir, M.L.; Rosenbloom, K.R.; Raney, B.J.; Lee, C.M.; Lee, B.T.; Karolchik, D.; et al. The UCSC Genome Browser database: 2018 update. Nucleic Acids Res. 2018, 46, D762–D769. [Google Scholar] [CrossRef] [Green Version]
- Jurka, J.; Kapitonov, V.V.; Pavlicek, A.; Klonowski, P.; Kohany, O.; Walichiewicz, J. Repbase Update, a database of eukaryotic repetitive elements. Cytogenet. Genome Res. 2005, 110, 462–467. [Google Scholar] [CrossRef]
- An, J.; Lai, J.; Lehman, M.L.; Nelson, C.C. miRDeep*: An integrated application tool for miRNA identification from RNA sequencing data. Nucleic Acids Res. 2013, 41, 727–737. [Google Scholar] [CrossRef]
- Liu, B.; Li, J.; Cairns, M.J. Identifying miRNAs, targets and functions. Brief. Bioinform. 2014, 15, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Heikkinen, L.; Wang, C.; Yang, Y.; Sun, H.; Wong, G. Trends in the development of miRNA bioinformatics tools. Brief. Bioinform. 2018, 20, 1836–1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Tong, Z.; Cui, Q.; Wang, J.; Zhou, Y. TransmiR v2.0: An updated transcription factor-microRNA regulation database. Nucleic Acids Res. 2019, 47, D253–D258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Center for Biotechnology Information-PubMed. Available online: https://www.ncbi.nlm.nih.gov/pubmed/ (accessed on 20 May 2020).
- Google Scholar. Available online: https://scholar.google.co.kr/ (accessed on 20 May 2020).
- Zhang, R.; Wang, Y.Q.; Su, B. Molecular evolution of a primate-specific microRNA family. Mol. Biol. Evol. 2008, 25, 1493–1502. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, P.N.; Huang, C.J.; Sugii, S.; Cheong, S.K.; Choo, K.B. Selective activation of miRNAs of the primate-specific chromosome 19 miRNA cluster (C19MC) in cancer and stem cells and possible contribution to regulation of apoptosis. J. Biomed. Sci. 2017, 24, 20. [Google Scholar] [CrossRef] [Green Version]
- Ovcharenko, I.; Nobrega, M.A.; Loots, G.G.; Stubbs, L. ECR Browser: A tool for visualizing and accessing data from comparisons of multiple vertebrate genomes. Nucleic Acids Res. 2004, 32, W280–W286. [Google Scholar] [CrossRef] [Green Version]
- Hofacker, I.L. Vienna RNA secondary structure server. Nucleic Acids Res. 2003, 31, 3429–3431. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Huang, J.; Yang, N.; Greshock, J.; Megraw, M.S.; Giannakakis, A.; Liang, S.; Naylor, T.L.; Barchetti, A.; Ward, M.R.; et al. microRNAs exhibit high frequency genomic alterations in human cancer. Proc. Natl. Acad. Sci. USA 2006, 103, 9136–9141. [Google Scholar] [CrossRef] [Green Version]
- Kutay, H.; Bai, S.; Datta, J.; Motiwala, T.; Pogribny, I.; Frankel, W.; Jacob, S.T.; Ghoshal, K. Downregulation of miR-122 in the rodent and human hepatocellular carcinomas. J. Cell Biochem. 2006, 99, 671–678. [Google Scholar] [CrossRef] [Green Version]
- Hayashita, Y.; Osada, H.; Tatematsu, Y.; Yamada, H.; Yanagisawa, K.; Tomida, S.; Yatabe, Y.; Kawahara, K.; Sekido, Y.; Takahashi, T. A polycistronic microRNA cluster, miR-17-92, is overexpressed in human lung cancers and enhances cell proliferation. Cancer Res. 2005, 65, 9628–9632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lal, A.; Kim, H.H.; Abdelmohsen, K.; Kuwano, Y.; Pullmann, R., Jr.; Srikantan, S.; Subrahmanyam, R.; Martindale, J.L.; Yang, X.; Ahmed, F.; et al. p16(INK4a) translation suppressed by miR-24. PLoS ONE 2008, 3, e1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roldo, C.; Missiaglia, E.; Hagan, J.P.; Falconi, M.; Capelli, P.; Bersani, S.; Calin, G.A.; Volinia, S.; Liu, C.G.; Scarpa, A.; et al. MicroRNA expression abnormalities in pancreatic endocrine and acinar tumors are associated with distinctive pathologic features and clinical behavior. J. Clin. Oncol. 2006, 24, 4677–4684. [Google Scholar] [CrossRef]
- Du, B.; Wang, Z.; Zhang, X.; Feng, S.; Wang, G.; He, J.; Zhang, B. MicroRNA-545 suppresses cell proliferation by targeting cyclin D1 and CDK4 in lung cancer cells. PLoS ONE 2014, 9, e88022. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, M.H.; Jensen, N.F.; Tarpgaard, L.S.; Qvortrup, C.; Romer, M.U.; Stenvang, J.; Hansen, T.P.; Christensen, L.L.; Lindebjerg, J.; Hansen, F.; et al. High expression of microRNA-625-3p is associated with poor response to first-line oxaliplatin based treatment of metastatic colorectal cancer. Mol. Oncol. 2013, 7, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Fan, Y.; Fa, Z.; Xu, J.; Yu, H.; Li, P.; Gu, J. microRNA-625 inhibits tumorigenicity by suppressing proliferation, migration and invasion in malignant melanoma. Oncotarget 2017, 8, 13253–13263. [Google Scholar] [CrossRef]
- Lin, L.; Cai, Q.; Zhang, X.; Zhang, H.; Zhong, Y.; Xu, C.; Li, Y. Two less common human microRNAs miR-875 and miR-3144 target a conserved site of E6 oncogene in most high-risk human papillomavirus subtypes. Prot. Cell 2015, 6, 575–588. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, X.; Yu, S.; Jeong, K.J.; Zhou, Z.; Han, L.; Tsang, Y.H.; Li, J.; Chen, H.; Mangala, L.S.; et al. Systematic characterization of A-to-I RNA editing hotspots in microRNAs across human cancers. Genome Res. 2017, 27, 1112–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, A.; Churakov, G.; Ramakodi, M.P.; Platt, R.N., 2nd; Jurka, J.; Kojima, K.K.; Caballero, J.; Smit, A.F.; Vliet, K.A.; Hoffmann, F.G.; et al. Multiple lineages of ancient CR1 retroposons shaped the early genome evolution of amniotes. Genome Biol. Evol. 2014, 7, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Sundaram, V.; Choudhary, M.N.; Pehrsson, E.; Xing, X.; Fiore, C.; Pandey, M.; Maricque, B.; Udawatta, M.; Ngo, D.; Chen, Y.; et al. Functional cis-regulatory modules encoded by mouse-specific endogenous retrovirus. Nat. Commun. 2017, 8, 14550. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, R.P.; Wildschutte, J.H.; Russo, C.; Coffin, J.M. Identification, characterization, and comparative genomic distribution of the HERV-K (HML-2) group of human endogenous retroviruses. Retrovirology 2011, 8, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, P.J.; Macfarlan, T.S.; Lorincz, M.C. Long Terminal Repeats: From Parasitic Elements to Building Blocks of the Transcriptional Regulatory Repertoire. Mol. Cell 2016, 62, 766–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perdomo, C.; Campbell, J.D.; Gerrein, J.; Tellez, C.S.; Garrison, C.B.; Walser, T.C.; Drizik, E.; Si, H.Q.; Gower, A.C.; Vick, J.; et al. MicroRNA 4423 is a primate-specific regulator of airway epithelial cell differentiation and lung carcinogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, 18946–18951. [Google Scholar] [CrossRef] [Green Version]
- Jo, A.; Im, J.; Lee, H.E.; Jang, D.; Nam, G.H.; Mishra, A.; Kim, W.J.; Kim, W.; Cha, H.J.; Kim, H.S. Evolutionary conservation and expression of miR-10a-3p in olive flounder and rock bream. Gene 2017, 628, 16–23. [Google Scholar] [CrossRef]
- Jo, A.; Lee, H.E.; Kim, H.S. Genomic Analysis of miR-21-3p and Expression Pattern with Target Gene in Olive Flounder. Genom. Inform. 2017, 15, 98–107. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Zhang, F.; Shugart, Y.Y.; Yang, L.; Li, X.; Liu, Z.; Sun, N.; Yang, C.; Guo, X.; Shi, J.; et al. The early growth response protein 1-miR-30a-5p-neurogenic differentiation factor 1 axis as a novel biomarker for schizophrenia diagnosis and treatment monitoring. Transl. Psychiatry 2017, 7, e998. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.C.; Jiang, W.; Mitra, R.; Cheng, F.; Yu, H.; Zhao, Z. Regulation rewiring analysis reveals mutual regulation between STAT1 and miR-155-5p in tumor immunosurveillance in seven major cancers. Sci. Rep. 2015, 5, 12063. [Google Scholar] [CrossRef] [Green Version]
- Lwin, T.; Zhao, X.; Cheng, F.; Zhang, X.; Huang, A.; Shah, B.; Zhang, Y.; Moscinski, L.C.; Choi, Y.S.; Kozikowski, A.P.; et al. A microenvironment-mediated c-Myc/miR-548m/HDAC6 amplification loop in non-Hodgkin B cell lymphomas. J. Clin. Investig. 2013, 123, 4612–4626. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.E.; Jo, A.; Im, J.; Cha, H.J.; Kim, W.J.; Kim, H.H.; Kim, D.S.; Kim, W.; Yang, T.J.; Kim, H.S. Characterization of the Long Terminal Repeat of the Endogenous Retrovirus-derived microRNAs in the Olive Flounder. Sci. Rep. 2019, 9, 14007. [Google Scholar] [CrossRef]
Figure 1.
Schematic step of analyzing microRNAs derived from transposable elements.

Figure 2.
Result of ECR browser and RNAfold on evolution related miRNAs derived from transposable element. On top each miRNA derived from TE, the nearest gene is presented. Yellow represents the region of searched miRNA, green represents transposons and simple repeats, salmon represents intronic region, and red represents intergenic regions. The x-axis represents the position in the human genome and the y-axis represents the conservation scale compared with human. Underneath the ECR browser conservation figure, each RNAhold structure of miRNAs derived from TE is shown. The scale bar on the left shows the minimum free energy (MFE) value of strong and weakest base-pairing values from 0 to 1.
Figure 2.
Result of ECR browser and RNAfold on evolution related miRNAs derived from transposable element. On top each miRNA derived from TE, the nearest gene is presented. Yellow represents the region of searched miRNA, green represents transposons and simple repeats, salmon represents intronic region, and red represents intergenic regions. The x-axis represents the position in the human genome and the y-axis represents the conservation scale compared with human. Underneath the ECR browser conservation figure, each RNAhold structure of miRNAs derived from TE is shown. The scale bar on the left shows the minimum free energy (MFE) value of strong and weakest base-pairing values from 0 to 1.


Figure 3.
The analyses of human diseases related miRNAs derived from TE and transcription factors by TransmiR. For evidence level, level 1 and 2 is based on ChIP-seq data and level 2 is more stringent than level 1. The Literature level is based on the research papers. Other information is shown in the figure.
Figure 3.
The analyses of human diseases related miRNAs derived from TE and transcription factors by TransmiR. For evidence level, level 1 and 2 is based on ChIP-seq data and level 2 is more stringent than level 1. The Literature level is based on the research papers. Other information is shown in the figure.

Figure 4.
The correlation of miRNAs derived from TE and transcription factors analyzed by TransmiR. For evidence level, level 1 and 2 are based on ChIP-seq data and level 2 is more stringent than level 1. The Literature level is based on the research papers. Other information is shown in the figure.
Figure 4.
The correlation of miRNAs derived from TE and transcription factors analyzed by TransmiR. For evidence level, level 1 and 2 are based on ChIP-seq data and level 2 is more stringent than level 1. The Literature level is based on the research papers. Other information is shown in the figure.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The distribution of transposable elements in various species.
| Human | Chimpanzee | Gorilla | Orangutan | Gibbon | Macaque | Rhesus | Marmoset | Mouse | Horse | Cow | Cat | Dog | Chicken | Zebrafish | C. elegans | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Non-TEs | 47.5% | 49.2% | 50.6% | 47.9% | 48.1% | 48.5% | 50.7% | 51.0% | 55.0% | 54.8% | 50.6% | 56.3% | 56.8% | 88.8% | 47.4% | 87.4% |
| SINE/MIR | 2.9% | 2.9% | 3.0% | 2.9% | 2.9% | 2.9% | 3.1% | 2.7% | 0.6% | 3.8% | 2.3% | 3.1% | 2.9% | 0.0% | 0.0% | 0.0% |
| SINE/Alu | 10.5% | 10.3% | 8.8% | 10.0% | 10.4% | 11.0% | 10.5% | 11.0% | 4.8% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| SINE/other | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 2.4% | 3.7% | 9.4% | 8.3% | 7.6% | 0.0% | 2.4% | 0.0% |
| SVA | 0.1% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| LINE/L2 | 3.7% | 3.8% | 3.8% | 3.7% | 3.8% | 3.8% | 4.0% | 3.3% | 0.4% | 5.2% | 2.7% | 4.0% | 3.7% | 0.0% | 1.7% | 0.0% |
| LINE/CR1 | 0.4% | 0.4% | 0.4% | 0.4% | 0.4% | 0.4% | 0.4% | 0.4% | 0.0% | 0.6% | 0.3% | 0.4% | 0.4% | 6.8% | 0.0% | 0.3% |
| LINE/RTE | 0.2% | 0.2% | 0.2% | 0.2% | 0.2% | 0.2% | 0.2% | 0.1% | 0.0% | 0.3% | 12.9% | 0.2% | 0.2% | 0.0% | 0.2% | 0.0% |
| LINE/L1 | 17.5% | 17.7% | 16.8% | 18.3% | 17.4% | 17.3% | 15.5% | 18.8% | 19.9% | 18.0% | 13.3% | 17.0% | 16.8% | 0.0% | 0.4% | 0.0% |
| LINE/other | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.5% | 0.0% |
| LTR/ERVK | 0.3% | 0.3% | 0.3% | 0.3% | 0.2% | 0.4% | 0.3% | 0.0% | 4.9% | 0.0% | 0.5% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| LTR/ERV1 | 2.9% | 2.8% | 2.7% | 2.9% | 2.6% | 2.7% | 2.8% | 2.2% | 1.2% | 1.9% | 1.4% | 1.0% | 1.0% | 0.2% | 0.4% | 0.0% |
| LTR/ERVL | 5.8% | 5.9% | 5.8% | 5.9% | 5.7% | 5.8% | 6.1% | 5.2% | 5.9% | 5.0% | 2.7% | 4.0% | 3.8% | 1.3% | 0.0% | 0.0% |
| LTR/Gypsy | 0.2% | 0.2% | 0.2% | 0.2% | 0.2% | 0.2% | 0.2% | 0.1% | 0.0% | 0.3% | 0.0% | 0.2% | 0.2% | 0.0% | 1.7% | 0.0% |
| LTR/other | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.2% | 0.0% | 0.0% | 0.0% | 0.0% | 3.1% | 0.3% |
| DNA/TcMar | 1.5% | 1.5% | 1.5% | 1.5% | 1.5% | 1.5% | 1.6% | 1.3% | 0.2% | 0.8% | 0.6% | 0.8% | 0.7% | 0.3% | 6.1% | 1.8% |
| DNA/hAT | 2.2% | 2.2% | 2.2% | 2.2% | 2.2% | 2.2% | 2.3% | 2.0% | 0.9% | 2.7% | 1.6% | 2.2% | 2.1% | 0.5% | 9.7% | 0.5% |
| DNA/other | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.2% | 19.6% | 6.1% |
| Other/Unknown | 4.3% | 2.6% | 3.7% | 3.6% | 4.4% | 3.1% | 2.3% | 1.9% | 3.8% | 2.7% | 1.7% | 2.5% | 3.8% | 1.9% | 6.8% | 3.6% |
| TOTAL: 100% | ||||||||||||||||
Table 2.
The list of evolution related miRNAs derived from transposable element. The coordinates of miRNAs derived from transposable element (TE) in the human genome, the type and name of TE that miRNAs are derived from, and the references are shown in each column.
Table 2.
The list of evolution related miRNAs derived from transposable element. The coordinates of miRNAs derived from transposable element (TE) in the human genome, the type and name of TE that miRNAs are derived from, and the references are shown in each column.
| microRNA | Coordinates | Type of TE | References | |
|---|---|---|---|---|
| 1 | miR-28 | chr3:188,688,781-188,688,866 | LINE_L2c, L2c | [53,55] |
| 2 | miR-130a | chr11:57,641,198-57,641,286 | LINE_MamRTE1 | [55] |
| 3 | miR-151a | chr8:140,732,564-140,732,653 | LINE_L2c, L2c | [55] |
| 4 | miR-151b | chr14:100,109,419-100,109,514 | LINE_L2b | [53,55] |
| 5 | miR-224 | chrX:151,958,578-151,958,658 | DNA_MER135 | [53] |
| 6 | miR-302e | chr11:7,234,766-7,234,837 | SINE_MIR | [70] |
| 7 | miR-320d-1 | chr13:40,727,816-40,727,887 | LINE_L1MEd | [53,55] |
| 8 | miR-342 | chr14:100,109,655-100,109,753 | SINE_MamSINE1 | [55] |
| 9 | miR-374a | chrX:74,287,286-74,287,357 | LINE_L2c | [53] |
| 10 | miR-378a | chr5:149,732,825-149,732,890 | SINE_MIRc, MIRc | [55] |
| 11 | miR-378b | chr3:10,330,229-10,330,285 | SINE_MIR3, MIRc | [54,55] |
| 12 | miR-378d-1 | chr4:5,923,275-5,923,328 | SINE_MIRb | [54,55] |
| 13 | miR-378d-2 | chr8:93,916,022-93,916,119 | SINE_MIRc | [54,55] |
| 14 | miR-378e | chr5:170,028,488-170,028,566 | SINE_MIRb, MIRc | [55] |
| 15 | miR-378f | chr1:23,929,070-23,929,147 | SINE_MIRc | [54,55] |
| 16 | miR-378h | chr5:154,829,458-154,829,540 | SINE_MIRc | [55] |
| 17 | miR-378i | chr22:41,923,222-41,923,297 | SINE_MIRc | [55] |
| 18 | miR-450b | chrX:134,540,185-134,540,262 | LINE_L1ME4a | [53] |
| 19 | miR-466 | chr3:31,161,704-31,161,787 | LINE_L1ME3 | [54] |
| 20 | miR-513a-1 | chrX:147,213,463-147,213,591 | DNA_MER91C | [54] |
| 21 | miR-513a-2 | chrX:147,225,826-147,225,952 | DNA_MER91C | [54] |
| 22 | miR-513b | chrX:147,199,044-147,199,127 | DNA_MER91C | [54] |
| 23 | miR-513c | chrX:147,189,704-147,189,787 | DNA_MER91C | [54] |
| 24 | miR-518d | chr19:53,734,877-53,734,963 | LINE_MamRTE1 | [54,71] |
| 25 | miR-544a | chr14:101,048,658-101,048,748 | DNA_MER5A1 | [54] |
| 26 | miR-544b | chr3:124,732,439-124,732,516 | DNA_MER5A1 | [54] |
| 27 | miR-548a-1 | chr6:18,571,784-18,571,880 | DNA_MADE1 | [54] |
| 28 | miR-548a-2 | chr6:135,239,160-135,239,256 | LTR_LTR16A2, DNA_MADE1, LTR_LTR16A2 | [54] |
| 29 | miR-570 | chr3:195,699,401-195,699,497 | DNA_MADE1 | [54] |
| 30 | miR-603 | chr10:24,275,685-24,275,781 | DNA_MADE1 | [56] |
| 31 | miR-619 | chr12:108,836,908-108,837,006 | LINE_L1MC4, SINE_AluSz6 | [54] |
| 32 | miR-637 | chr19:3,961,414-3,961,512 | LINE_L1MC4a | [57] |
| 33 | miR-652 | chrX:110,055,329-110,055,426 | DNA_MER91C | [54] |
| 34 | miR-1202 | chr6:155,946,797-155,946,879 | LTR_MER52A | [56] |
| 35 | miR-1261 | chr11:90,869,121-90,869,202 | DNA_Tigger1 | [54] |
| 36 | miR-1268a | chr15:22,225,278-22,225,329 | SINE_AluJo | [54] |
| 37 | miR-1268b | chr17:80,098,828-80,098,877 | SINE_AluSx1 | [54] |
| 38 | miR-1273c | chr6:154,853,360-154,853,436 | SINE_AluJo | [54] |
| 39 | miR-1273h | chr16:24,203,116-24,203,231 | SINE_AluJb | [54] |
| 40 | miR-1302-1 | chr12:112,695,034-112,695,176 | DNA_MER54 | [54] |
| 41 | miR-1303 | chr5:154,685,776-154,685,861 | SINE_AluJr, FLAM_A | [54] |
| 42 | miR-1304 | chr11:93,733,674-93,733,764 | SINE_AluJo | [54] |
| 43 | miR-1587 | chrX:39,837,561-39,837,613 | LTR_MLT1H2 | [54] |
| 44 | miR-1972-1 | chr16:15,010,321-15,010,397 | SINE_AluSx, FLAM_A | [54] |
| 45 | miR-2355 | chr2:207,109,987-207,110,073 | LINE_MamRTE1, MamRTE1 | [53] |
| 46 | miR-3118-1 | chr21:13,644,775-13,644,850 | LINE_L1PA12 | [54] |
| 47 | miR-3118-2 | chr15:20,832,795-20,832,869 | LINE_L1PA14 | [54] |
| 48 | miR-3118-3 | chr15:21,406,385-21,406,459 | LINE_L1PA13 | [54] |
| 49 | miR-3118-4 | chr15:21,843,750-21,843,824 | LINE_L1PA13 | [54] |
| 50 | miR-4452 | chr4:86,542,482-86,542,552 | SINE_AluJo | [54] |
| 51 | miR-6303 | chr10:24,275,685-24,275,781 | DNA_MADE1 | [54] |
Table 3.
The list of human disease related miRNAs derived from transposable element. The coordinates of miRNAs derived from TE in the human genome, the type and name of TE that miRNAs are derived from related disorders and the references are shown in each column.
Table 3.
The list of human disease related miRNAs derived from transposable element. The coordinates of miRNAs derived from TE in the human genome, the type and name of TE that miRNAs are derived from related disorders and the references are shown in each column.
| microRNA | Coordinates | Type of TE | Related Disorders | References | |
|---|---|---|---|---|---|
| 1 | miRNA-28 | chr3:188,688,781-188,688,866 | LINE_L2c, L2c | OC, M | [74] |
| 2 | miRNA-95 | chr4:8,005,301-8,005,381 | LINE_L2b, L2c | OC, BrC | [74] |
| 3 | miRNA-130a | chr11:57,641,198-57,641,286 | LINE_MamRTE1 | LuC, OC, LiC | [46,48,75,76] |
| 4 | miRNA-151a | chr8:140,732,564-140,732,653 | LINE_L2c | G, OC, BrC, M | [49,74] |
| 5 | miRNA-151b | chr14:100,109,419-100,109,514 | LINE_L2b | OC, BrC, M | [74] |
| 6 | miRNA-181c | chr19:13,874,699-13,874,808 | LINE_MamRTE1 | LiC, A, OC, BrC | [74,77] |
| 7 | miRNA-224 | chrX: 151,958,578-151,958,658 | DNA_MER135 | LiC | [48,51] |
| 8 | miRNA-320d-1 | chr13:40,727,816-40,727,887 | LINE_L1MEd | OC, BrC, M, N | [47,48,74] |
| 9 | miRNA-335 | chr7:130,496,111-130,496,204 | SINE_MIRb | OC, M | [74] |
| 10 | miRNA-340 | chr5:180,015,303-180,015,397 | DNA_MARNA | OC | [74] |
| 11 | miRNA-342 | chr14:100,109,655-100,109,753 | SINE_MamSINE1 | EC, ACC, PaC | [48,78] |
| 12 | miRNA-361 | chrX: 85,903,636-85,903,707 | DNA_MER5A | LuC | [52] |
| 13 | miRNA-378a | chr5:149,732,825-149,732,890 | SINE_MIRc, MIRc | BlC | [49] |
| 14 | miRNA-378b | chr3:10,330,229-10,330,285 | SINE_MIR3, MIRc | L | [49] |
| 15 | miRNA-421 | chrX:74,218,377-74,218,461 | LINE_L2c, L2c | Ph, Pa | [49] |
| 16 | miRNA-513a-1 | chrX:147,213,463-147,213,591 | DNA_MER91C | LuC | [79] |
| 17 | miRNA-513a-2 | chrX:147,225,826-147,225,952 | DNA_MER91C | LuC | [79] |
| 18 | miRNA-518d | chr19:53,734,877-53,734,963 | LINE_MamRTE1 | LuC | [79] |
| 19 | miRNA-545 | chrX:74,287,104-74,287,209 | LINE_L2c | LuC | [79] |
| 20 | miRNA-546b | chr6:119,069,022-119,069,167 | DNA_MADE1 | LuC | [79] |
| 21 | miRNA-548l | chr11:94,466,495-94,466,580 | DNA_MADE1 | LuC | [79] |
| 22 | miRNA-548m | chrX:95,063,141-95,063,226 | DNA_MADE1 | LuC | [79] |
| 23 | miRNA-625 | chr14:65,471,102-65,471,186 | LINE_L1MCa | M, GC, CoC, LuC, KC | [49,80,81] |
| 24 | miRNA-626 | chr15:41,691,585-41,691,678 | LINE_L1MB8, L1MB8 | HPV | [82] |
| 25 | miRNA-646 | chr20:60,308,474-60,308,567 | LTR_LTR67B | LuC | [79] |
| 26 | miRNA-659 | chr22:37,847,678-37,847,774 | DNA_Arthur1B | D | [77] |
| 27 | miRNA-1290 | chr1:18,897,071-18,897,148 | DNA_Tigger4a | LuC | [79] |
| 28 | miRNA-1294 | chr5:154,347,071-154,347,283 | SINE_MIRb | LuC | [79] |
| 29 | miRNA-2355 | chr2:207,109,987-207,110,073 | LINE_MamRTE1, MamRTE1 | HPV | [82] |
| 30 | miRNA-1304 | chr11:93,733,674-93,733,764 | SINE_AluJo | LuC, Pr, UCEC | [50,83] |
| 31 | miRNA-3144 | chr6:120,015,179-120,015,257 | LINE_L1MA8 | LiC PrC, CeC, HPV | [82,83] |
| 32 | miRNA-3681 | chr2:12,199,130-12,199,201 | LTR_LTR16D1 | CeC, LiC | [49] |
| 33 | miRNA-4662a | chr8:124,821,985-124,822,051 | LINE_L1ME4b | LuC | [49] |
| 34 | miRNA-6503 | chr11:60,209,071-60,209,156 | LINE_MLT1D | LiC, L | [49,83] |
Abbreviations: Ovarian cancer (OC), Melanoma (M), Breast cancer (BrC), Lung cancer (LuC), Glioblastoma (G), Alzheimer (A), Neuroblastoma (N), Endocrine cancer (EC), Acrinar cell carcinoma (ACC), Pancreatic cancer (PaC), Bladder cancer (BlC), Leukemia (L), Pheochromocytoma (Ph), Paraganglioma (Pa), Gastric cancer (GC), Colorectal cancer (CoC), Kidney cancer (KC), Human papillomavirus (HPV), Dementia (D), Preeclampsia (Pr), Uterine Corpus Endometrial Carcinoma (UCEC), Prostate cancer (PrC), Cervical cancer (CeC), Liver cancer (LiC).
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lee, H.-E.; Huh, J.-W.; Kim, H.-S. Bioinformatics Analysis of Evolution and Human Disease Related Transposable Element-Derived microRNAs. Life 2020, 10, 95. https://doi.org/10.3390/life10060095
AMA Style
Lee H-E, Huh J-W, Kim H-S. Bioinformatics Analysis of Evolution and Human Disease Related Transposable Element-Derived microRNAs. Life. 2020; 10(6):95. https://doi.org/10.3390/life10060095
Chicago/Turabian StyleLee, Hee-Eun, Jae-Won Huh, and Heui-Soo Kim. 2020. "Bioinformatics Analysis of Evolution and Human Disease Related Transposable Element-Derived microRNAs" Life 10, no. 6: 95. https://doi.org/10.3390/life10060095
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.