
Ecological and Syntaxonomic Analysis of Pinus halepensis Mill. in the Iberian Peninsula and Balearic Islands

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
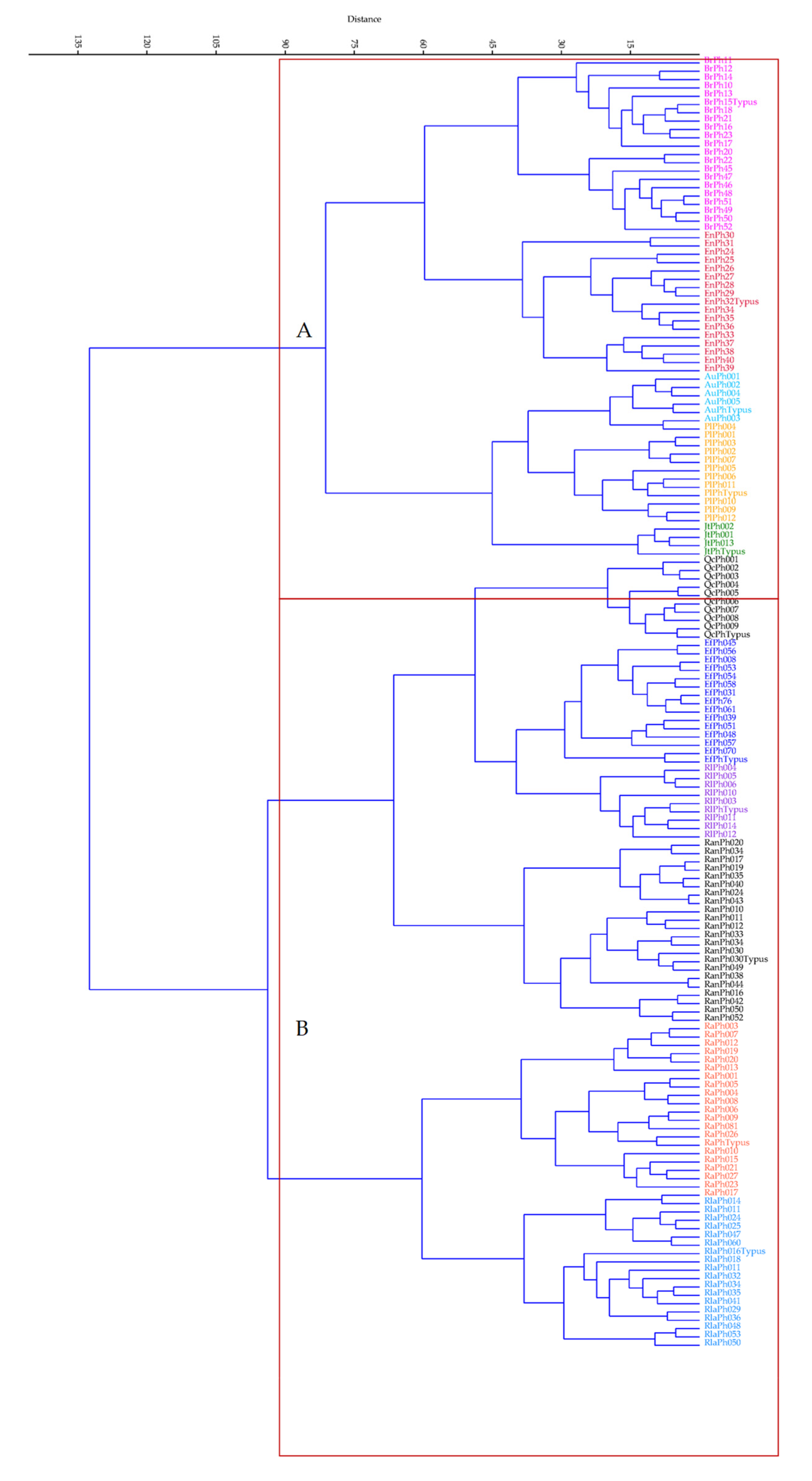
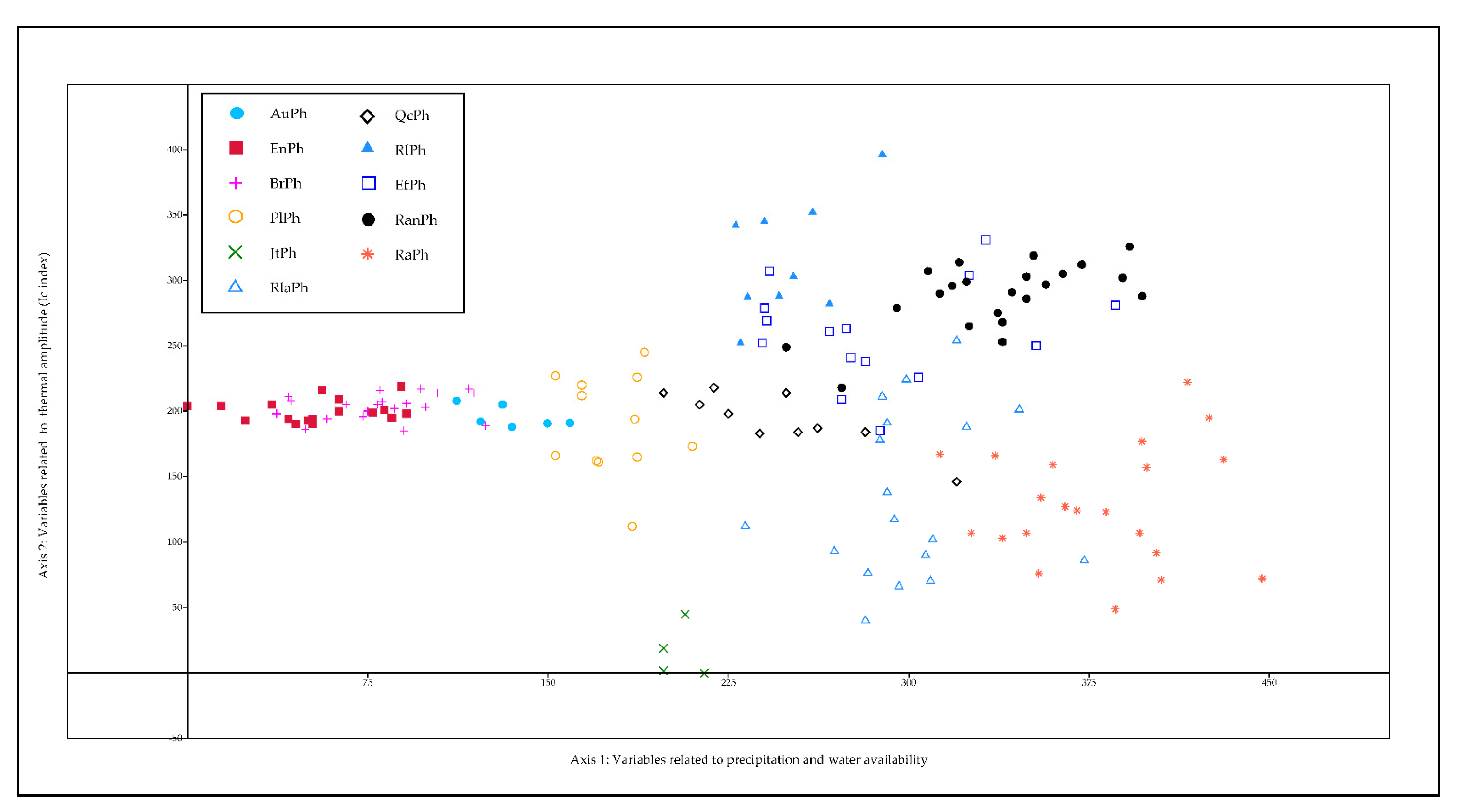
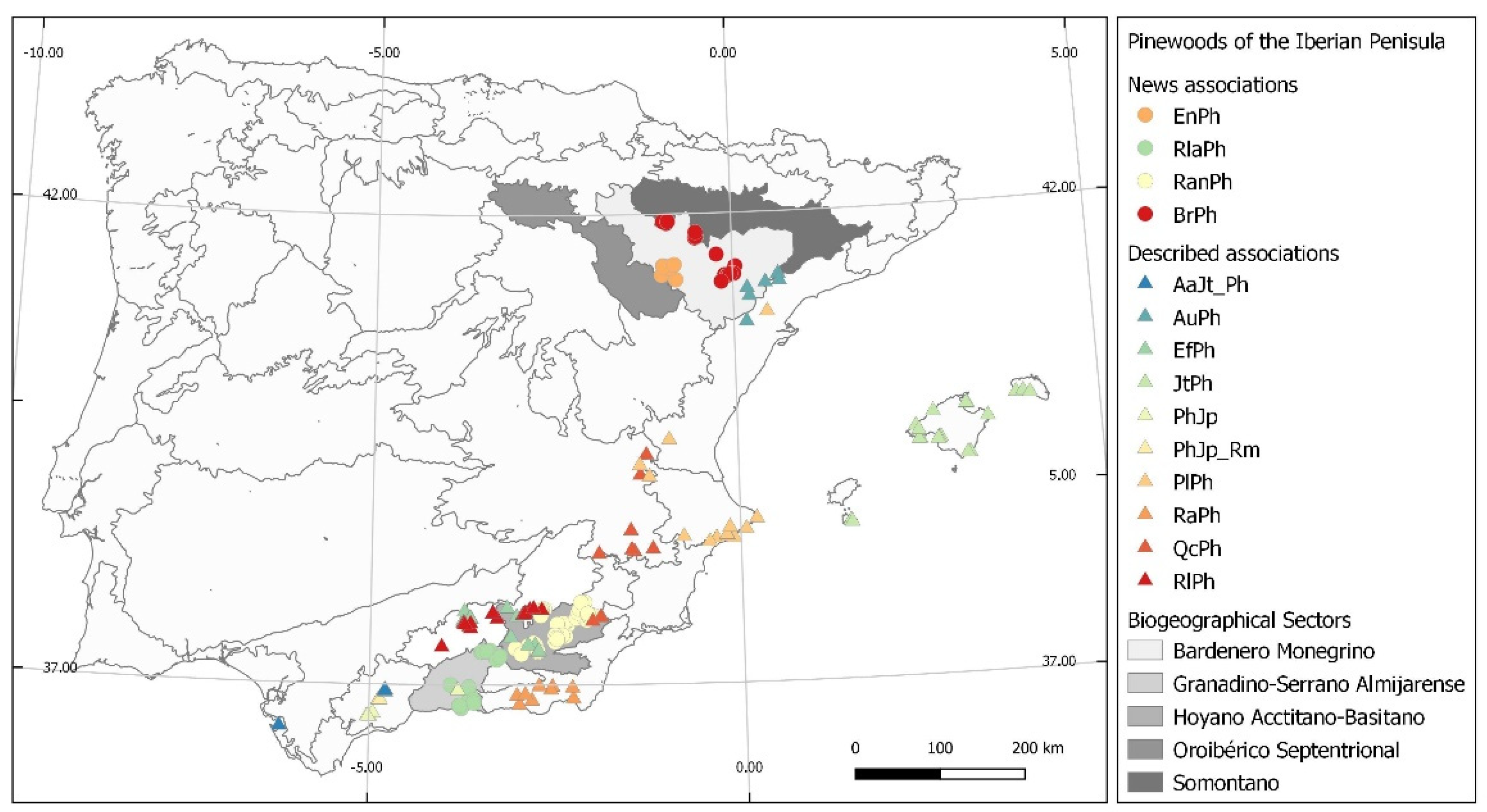
3. Results
3.1. Ass. Arbuto-Pinetum halepensis
3.2. Ass. Ephedro nebrodensis-Pinetum halepensis
3.3. Ass. Bupleuro rigidi-Pinetum halepensis
3.4. Ass. Ceratonio siliquae-Pinetum halepensis
3.5. Ass. Querco cocciferae-Pinetum halepensis
3.6. Ass. Junipero turbinatae-Pinetum halepensis
3.7. Ass. Rhamno angustifoliae-Pinetum halepensis
3.8. Ass. Rhamno almeriensis-Pinetum almeriensis
3.9. Ass. Rhamno laderoi-Pinetum halepensis
4. Discussion
5. Conclusions
Syntaxonomical Checklist
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Surface m2 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 |
| Atitude in m | 629 | 601 | 547 | 596 | 593 | 570 | 595 | 536 | 619 | 585 | 662 | 587 | 562 | 594 | 582 |
| Orientation | N | N | NE | SE | NE | NE | N | E | NO | E | N | N | E | N | NE |
| Slope in decimal degrees | 22.14 | 19.32 | 10.8 | 13.36 | 10.44 | 24.86 | 29.53 | 11.24 | 5.07 | 16.69 | 24.94 | 17.33 | 11.36 | 5.9 | 5.95 |
| Cover rate % | 90 | 80 | 90 | 90 | 80 | 80 | 90 | 90 | 70 | 90 | 90 | 60 | 80 | 60 | 80 |
| Ic | 17.77 | 17.8 | 17.92 | 17.8 | 17.8 | 17.86 | 17.79 | 18.04 | 17.89 | 17.83 | 17.72 | 17.81 | 17.87 | 17.98 | 17.85 |
| Io | 1.97 | 2.5 | 1.96 | 1.99 | 2 | 1.99 | 2.03 | 2 | 2.41 | 1.95 | 2 | 2.02 | 2 | 2.07 | 1.94 |
| Itc | 269.7 | 269.98 | 272.45 | 270.93 | 270.98 | 271.47 | 270.46 | 269.02 | 261.71 | 271.93 | 268.24 | 270.66 | 271.64 | 265.48 | 272.15 |
| Nª inventory | EnPh24 | EnPh25 | EnPh26 | EnPh27 | EnPh28 | EnPh29 | EnPh30 | EnPh32Typus | EnPh33 | EnPh35 | EnPh36 | EnPh37 | EnPh38 | EnPh39 | EnPh40 |
| Nº Orden | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
| Characteristic species | |||||||||||||||
| Pinus halepensis Mill. | 5 | 4 | 5 | 5 | 4 | 4 | 5 | 5 | 3 | 5 | 5 | 3 | 4 | 3 | 4 |
| Quercus coccifera L. subsp. coccifera | 2 | 4 | 4 | 2 | 4 | 4 | 3 | 2 | 3 | + | 2 | + | 2 | + | |
| Juniperus oxycedrus L. subsp. oxycedrus | 1 | + | 1 | + | 1 | + | 3 | + | + | + | + | + | + | ||
| Juniperus phoenicea L. subsp. phoenicea | 2 | 2 | 1 | + | 1 | + | + | + | + | + | + | ||||
| Bupleurum rigidum L. | 1 | 1 | + | 2 | 3 | 1 | 3 | 2 | + | ||||||
| Rubia peregrina L. subsp. peregrina | + | 2 | + | 1 | 2 | 2 | + | + | 2 | + | |||||
| Ephedra nebrodensis Guss. | + | + | + | + | + | + | 1 | + | 1 | 1 | + | + | |||
| Rhamnus lycioides L. subsp. lycioides | + | + | + | + | 3 | + | |||||||||
| Pistacia lentiscus L. | + | + | + | ||||||||||||
| Carex halleriana Asso. | + | + | |||||||||||||
| Asparagus acutifolius L. | + | + | |||||||||||||
| Quercus rotundifolia Lam. | 2 | ||||||||||||||
| Rhamnus alaternus L. subsp. alaternus | + | ||||||||||||||
| Arctostaphylos uva-ursi (L.) Spreng. | + | ||||||||||||||
| Companion species | |||||||||||||||
| Carex humilis Leyss. | + | 1 | 1 | 1 | 1 | 1 | + | 2 | 2 | 1 | 1 | 1 | 2 | 2 | 2 |
| Brachypodium retusum (Pers.) P. Beauv. | + | 3 | 2 | 2 | 3 | 2 | 3 | 1 | 2 | 5 | 2 | + | 3 | ||
| Stipa orientalis Trin. | + | + | + | + | 3 | + | 1 | 1 | 2 | + | + | 1 | 3 | ||
| Rosmarinus officinalis L. | 2 | + | + | + | + | + | 2 | + | + | + | 2 | 1 | |||
| Centaurea linifolia L. | 1 | + | 1 | + | + | + | + | + | + | + | |||||
| Linum suffruticosum L. | + | + | + | + | + | + | + | + | 1 | + | |||||
| Coronilla minima L. subsp. lotoides (W.D.J. Koch) Nyman | 3 | + | + | + | + | + | + | + | + | + | |||||
| Echinops ritro L. | + | + | + | 3 | + | + | + | + | + | + | |||||
| Avenula bromoides (Gouan) H. Scholz | + | + | + | + | + | + | + | 1 | + | ||||||
| Bupleurum fruticescens Loefl. ex L. | + | + | + | + | + | + | + | + | |||||||
| Euphorbia minuta Loscos & J. Pardo | + | + | + | + | + | 3 | 1 | + | |||||||
| Polygala rupestris Pourr. | + | + | + | + | 3 | + | + | 3 | |||||||
| Pleurochaete squarrosa Linderg | 1 | 2 | 1 | + | + | + | 3 | ||||||||
| Salvia officinalis L. | + | + | + | + | + | + | 3 | ||||||||
| Helianthemum myrtifolium (Lam.) Samp. | + | + | + | 1 | + | + | + | ||||||||
| Teucrium chamaedrys L. subsp. pinnatifidum (Sennen) Rech. fil. | + | + | 3 | + | 1 | 1 | + | ||||||||
| Rhaponticum coniferum (L.) Greuter | + | + | + | + | + | + | |||||||||
| Aristolochia pistolochia L. | 1 | + | + | + | + | 3 | |||||||||
| Genista scorpius (L.) DC. in Lam. & DC. | + | + | + | + | 1 | + | |||||||||
| Aphyllanthes monspeliensis L. | + | 3 | + | 3 | 2 | ||||||||||
| Fumana ericoides (Cav.) Gand. in Magnier | 3 | + | + | + | + | ||||||||||
| Lavandula latifolia Medik. | + | + | + | + | + | ||||||||||
| Thymus vulgaris subsp. vulgaris L. | + | + | 1 | + | + | ||||||||||
| Viola rupestris F.W. Schmidt | + | + | + | + | + | ||||||||||
| Cistus albidus L. | + | + | + | + | |||||||||||
| Cistus clusii subsp. clusii Dunal | 3 | 1 | + | + | |||||||||||
| Cistus libanotis L. | 3 | 1 | + | + | |||||||||||
| Inula montana L. | + | + | + | ||||||||||||
| Koeleria vallesiana subsp. vallesiana (Honck.) Gaudin | + | + | 1 | ||||||||||||
| Lithodora fruticosa (L.) Griseb. | + | + | + | ||||||||||||
| Orobanche latisquama (F.W. Schultz) Batt. in Batt. & Trab. | + | 3 | + | ||||||||||||
| Stereodon cupressiforme (Hedw.) Brid. ex Mitt. | 2 | 2 | + | ||||||||||||
| Dorycnium pentaphyllum Scop. | + | + | + | ||||||||||||
| Euphorbia verrucosa Lam. subsp. mariolensis (Rouy) Vives | + | + | + | ||||||||||||
| Anacamptis pyramidalis (L.) Rich. | 3 | 3 | |||||||||||||
| Globularia vulgaris L. | + | + | |||||||||||||
| Hedysarum boveanum Bunge ex Basiner subsp. europaeum Guitt. & Kerguélen | + | + | |||||||||||||
| Helianthemum syriacum (Jacq.) Dum. Cours. | + | + | |||||||||||||
| Ononis fruticosa L. | + | 1 | |||||||||||||
| Staehelina dubia L. | + | + | |||||||||||||
| Teucrium polium L. subsp. aragonense (Loscos & J.Pardo) Nyman | + | + | |||||||||||||
| Centaurium quadrifolium (L.) G. López & Jarvis subsp. barrelieri (L.M. Dufour) G. López | + | + |
| Surface m2 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 |
| Atitude in m | 619 | 532 | 669 | 652 | 700 | 700 | 675 | 671 | 686 | 600 | 570 | 622 | 265 | 327 | 149 |
| Orientation | NE | S | N | N | N | N | O | N | E | N | E | NE | NO | N | E |
| Slope in decimal degrees | 11.1 | 6.3 | 19.15 | 4.4 | 17.78 | 20.85 | 15.68 | 22.72 | 20.21 | 22.91 | 21.64 | 16.26 | 16.01 | 24.17 | 23.67 |
| Cover rate % | 85 | 85 | 90 | 90 | 90 | 90 | 90 | 90 | 80 | 65 | 80 | 90 | 80 | 80 | 85 |
| Ic | 17.6 | 18 | 16.97 | 18.88 | 18.73 | 18.73 | 18.82 | 18.83 | 18.87 | 17.77 | 18.79 | 17.58 | 19.38 | 18.76 | 19.37 |
| Io | 2.77 | 2.55 | 2.8 | 2.06 | 2.11 | 2.11 | 2.09 | 2.09 | 2.22 | 2.73 | 1.92 | 2.78 | 1.73 | 1.73 | 1.71 |
| Itc | 237.72 | 249.11 | 230.97 | 258.34 | 255.4 | 255.42 | 256.52 | 256.7 | 249.56 | 241.03 | 268.36 | 237.14 | 283.24 | 284.66 | 291.59 |
| Nª inventory | BrPh10 | BrPh11 | BrPh12 | BrPh14 | BrPh15Typus | BrPh16 | BrPh17 | BrPh18 | BrPh20 | BrPh21 | BrPh22 | BrPh23 | BrPh45 | BrPh46 | BrPh52 |
| Nº Orden | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
| Characteristic species | |||||||||||||||
| Pinus halepensis Mill. | 4 | 4 | 5 | 5 | 5 | 5 | 5 | 5 | 4 | 3 | 4 | 5 | 4 | 4 | 4 |
| Quercus coccifera L. | 2 | 3 | 4 | 3 | 3 | 3 | 4 | 3 | 3 | 4 | 2 | 3 | 4 | 3 | 1 |
| Juniperus oxycedrus L. | 2 | + | + | + | 2 | 1 | + | 3 | 2 | 2 | + | + | + | ||
| Juniperus phoenicea L. | + | 3 | 1 | + | + | 1 | + | + | 2 | 3 | 2 | ||||
| Carex halleriana Asso. | 3 | 1 | + | + | + | 2 | + | 1 | 1 | 1 | + | + | 1 | + | |
| Rubia peregrina L. | + | + | + | 1 | + | + | 2 | 1 | + | 1 | + | 1 | + | ||
| Rhamnus lycioides L. | + | + | 3 | 1 | + | 2 | |||||||||
| Arctostaphylos uva-ursi (L.) Spreng. | + | 1 | + | + | + | 4 | 2 | 4 | 3 | ||||||
| Pistacia lentiscus L. | 1 | 2 | + | ||||||||||||
| Quercus rotundifolia Lam. | 3 | + | + | 2 | + | + | |||||||||
| Ephedra nebrodensis Guss. | + | 1 | + | ||||||||||||
| Rhamnus alaternus L. | + | + | + | ||||||||||||
| Bupleurum rigidum L. | + | 1 | 1 | 1 | 3 | + | + | ||||||||
| Juniperus thurifera L. | + | + | + | + | 1 | ||||||||||
| Phillyrea angustifolia L. | + | ||||||||||||||
| Quercus faginea Lam. | + | ||||||||||||||
| Companion species | |||||||||||||||
| Brachypodium retusum (Pers.) P. Beauv. | + | 2 | 1 | + | 1 | + | 1 | 1 | 2 | 1 | + | 1 | 1 | 1 | + |
| Teucrium chamaedrys L. subsp. pinnatifidum (Sennen) Rech. fil. | 1 | 1 | 1 | 1 | 1 | 3 | + | 2 | 3 | + | 1 | ||||
| Thymelaea tinctoria (Pourr.) Endl. | + | + | + | + | + | + | + | + | + | + | + | ||||
| Stereodon cupressiformis (Hedw.) Brid. ex Mitt. | 1 | + | 3 | 1 | 1 | 2 | 2 | 1 | 2 | 2 | 3 | ||||
| Helianthemum myrtifolium (Lam.) Samp. | 1 | 2 | + | + | + | + | + | + | + | ||||||
| Rosmarinus officinalis L. | 3 | + | + | + | + | + | 1 | 2 | |||||||
| Rhaponticum coniferum (L.) Greuter | + | + | + | + | + | + | |||||||||
| Asphodelus cerasiferus J. Gay | 3 | + | + | + | 3 | + | |||||||||
| Genista scorpius (L.) DC. in Lam. & DC. | + | + | + | + | + | ||||||||||
| Pleurochaete squarrosa Linderg | + | 1 | + | 1 | + | ||||||||||
| Festuca rubra L. | 3 | + | + | + | + | ||||||||||
| Viola rupestris F.W. Schmidt | 1 | 3 | + | + | + | ||||||||||
| Dorycnium pentaphyllum Scop. | 1 | 3 | + | + | 1 | ||||||||||
| Helianthemum origanifolium subsp. origanifolium (Lam.) Pers. | + | + | + | 1 | |||||||||||
| Salvia officinalis L. | + | + | + | + | |||||||||||
| Cladonia pyxidata (L.) Hoffm. | 3 | + | + | 3 | |||||||||||
| Thymus vulgaris L. | 3 | + | + | ||||||||||||
| Viscum album L. subsp. austriacum (Wiesb.) Vollm. | + | + | + | ||||||||||||
| Staehelina dubia L. | + | 3 | 3 | ||||||||||||
| Bupleurum fruticescens Loefl. ex L. | + | 3 | + | ||||||||||||
| Aristolochia pistolochia L. | + | + | |||||||||||||
| Epipactis microphylla (Ehrh.) Sw. | + | + | |||||||||||||
| Galium pumilum Murray | + | + | |||||||||||||
| Helianthemum violaceum (Cav.) Pers. | + | + | |||||||||||||
| Homalothecium sericeum W. P. Schimper | + | 3 | |||||||||||||
| Linum narbonense L. | 3 | 3 | |||||||||||||
| Thalictrum tuberosum L. | + | + | |||||||||||||
| Thesium humifusum DC. in Lam. & DC. | + | + |
| Surface m2 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 |
| Atitude in m | 1212 | 1233 | 1179 | 1150 | 1326 | 1192 | 1100 | 1209 | 1154 | 1179 | 1179 | 1254 | 1086 | 1160 | 1111 | 1011 | 1100 | 971 | 894 | 1212 |
| Orientation | NE | NE | SO | SE | NO | E | E | SO | NE | N | E | E | NO | E | NO | O | E | NO | SE | E |
| Slope in decimal degrees | 7.82 | 19.76 | 19.43 | 8.19 | 12.51 | 15.82 | 12.89 | 7.52 | 13.86 | 26.67 | 19.43 | 6.76 | 26.32 | 15.29 | 3.87 | 14.78 | 12.89 | 6.6 | 17.93 | 22.2 |
| Cover rate % | 80 | 80 | 80 | 55 | 95 | 70 | 65 | 55 | 65 | 65 | 40 | 55 | 55 | 55 | 65 | 65 | 65 | 65 | 55 | 55 |
| Ic | 18.02 | 17.85 | 17.87 | 18.62 | 18.34 | 18.57 | 18.07 | 18.58 | 18.08 | 18.59 | 17.87 | 18.54 | 18.51 | 17.78 | 17.85 | 17.7 | 18.07 | 17.6 | 17.45 | 18.52 |
| Io | 2.23 | 2.47 | 2.39 | 2.3 | 2.64 | 2.43 | 2.12 | 2.25 | 2.21 | 2.4 | 2.39 | 2.33 | 1.99 | 2.33 | 2.23 | 2.08 | 2.12 | 2.04 | 2.02 | 2.47 |
| Itc | 230.18 | 228.71 | 233.32 | 245.39 | 230.21 | 232.87 | 247.87 | 233.38 | 238.31 | 234.47 | 233.32 | 229.61 | 260.01 | 238.42 | 248.41 | 270.19 | 247.87 | 275.7 | 282.81 | 231.04 |
| Nª inventory | RanPh063 | RanPh065 | RanPh066 | RanPh010 | RanPh011 | RanPh012 | RanPh030 | RanPh033 | RanPh034 | RanPh049 | RanPh066 | RanPh069 | RanPh071 | RanPh017 | RanPh019 | RanPh024 | RanPh030Typus | RanPh038 | RanPh043 | RanPh044 |
| Nº Orden | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 |
| Characteristic species | ||||||||||||||||||||
| Pinus halepensis Mill. | 4 | 4 | 4 | 3 | 5 | 4 | 3 | 3 | 3 | 3 | 2 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Rhamnus oleoides subsp. angustifolia (Lange & Willk.) Rivas Mart. & J.M. Pizarro | 2 | 2 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 2 | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 1 | 1 |
| Quercus rotundifolia Lam. | 2 | 2 | 1 | 1 | + | + | + | + | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | |
| Juniperus oxycedrus subsp. oxycedrus L. | 1 | 3 | + | + | + | + | + | + | + | 2 | 2 | |||||||||
| Juniperus phoenicea subsp. phoenicea L. | + | + | + | 1 | 1 | |||||||||||||||
| Carex halleriana Asso. | 1 | 1 | 2 | 1 | ||||||||||||||||
| Rhamnus lycioides subsp. lycioides L. | 1 | |||||||||||||||||||
| Asparagus acutifolius L. | + | |||||||||||||||||||
| Daphne gnidium L. | + | + | 1 | 2 | 1 | |||||||||||||||
| Quercus coccifera subsp. coccifera L. | 2 | 1 | 2 | |||||||||||||||||
| Crataegus monogyna subsp. brevispina (Kunze) Franco | 1 | + | ||||||||||||||||||
| Pistacia lentiscus L. | ||||||||||||||||||||
| Pistacia terebinthus L. | + | + | ||||||||||||||||||
| Companion species | ||||||||||||||||||||
| Rosmarinus officinalis L. | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | ||||||
| Macrochloa tenacissima (L.) Kunth | 1 | 1 | 1 | 3 | + | 2 | 2 | 2 | 2 | 1 | 2 | 2 | 2 | |||||||
| Brachypodium retusum subsp. retusum (Pers.) P. Beauv. | 2 | 1 | 2 | 1 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | |||||||||
| Thymus zygis subsp. gracilis (Boiss.) R. Morales | 1 | 1 | 3 | 1 | 2 | 2 | 2 | 2 | 1 | 1 | ||||||||||
| Genista scorpius (L.) DC. in Lam. & DC. | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | 2 | |||||||||||
| Thymus membranaceus subsp. membranaceus Boiss. | 2 | 1 | 2 | 2 | 2 | 1 | 2 | 2 | ||||||||||||
| Erinacea anthyllis Link | 2 | + | 2 | 2 | 2 | 2 | ||||||||||||||
| Festuca scariosa (Lag.) Asch. & Graebn. | + | 2 | 2 | 2 | ||||||||||||||||
| Cistus clusii subsp. clusii Dunal | 1 | 1 | 1 | 2 | ||||||||||||||||
| Lavandula stoechas L. | 3 | 3 | 3 | |||||||||||||||||
| Helichrysum stoechas (L.) Moench | 1 | 1 | 1 | |||||||||||||||||
| Helianthemum almeriense Pau | 1 | 1 | 1 | |||||||||||||||||
| Bupleurum fruticescens subsp. spinosum (Gouan) O. Bolòs & Vigo | 2 | 1 | 2 | |||||||||||||||||
| Fumana thymifolia (L.) Spach ex Webb | 2 | 1 | 1 | |||||||||||||||||
| Genista cinerea subsp. speciosa Losa & Rivas Goday (Vill.) DC. | + | 2 | 2 | |||||||||||||||||
| Phlomis lychnitis L. | 3 | 2 | + | |||||||||||||||||
| Dorycnium pentaphyllum Scop. | 1 | 1 | ||||||||||||||||||
| Euphorbia nicaeensis All. | 3 | + | ||||||||||||||||||
| Sideritis incana L. | 1 | 1 | ||||||||||||||||||
| Stipa lagascae Roem. & Schult. | 1 | 1 | ||||||||||||||||||
| Thymus mastichina subsp. mastichina (L.) L. | 1 | + | ||||||||||||||||||
| Trifolium spumosum L. | 2 | 1 | ||||||||||||||||||
| Ulex parviflorus subsp. parviflorus Pourr. | + | 1 |
| Surface m2 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 | 300 |
| Atitude in m | 1270 | 1076 | 1034 | 1102 | 1040 | 1223 | 1071 | 226 | 422 | 552 | 144 | 560 | 1012 | 894 | 226 |
| Orientation | O | S | S | O | SO | NE | NE | NO | S | NE | S | S | O | SE | NO |
| Slope in decimal degrees | 22.39 | 10.1 | 28.19 | 13.39 | 35.8 | 14.09 | 11.58 | 25.48 | 12.33 | 27.98 | 28.51 | 31.64 | 40.25 | 33.22 | 25.48 |
| Cover rate % | 60 | 60 | 80 | 65 | 80 | 65 | 65 | 60 | 60 | 65 | 90 | 60 | 60 | 65 | 50 |
| Ic | 18.44 | 18.24 | 18.37 | 18.38 | 16.97 | 18.66 | 17.5 | 13.33 | 13.55 | 15.52 | 14.05 | 14.87 | 18.74 | 16.05 | 13.33 |
| Io | 3.75 | 3.21 | 3.11 | 3.23 | 3.37 | 3.69 | 3.44 | 2.36 | 2.59 | 2.78 | 1.94 | 2.88 | 2.54 | 3.12 | 2.36 |
| Itc | 244.61 | 282.46 | 284.49 | 272.54 | 296.91 | 238.15 | 272.02 | 434.04 | 423.51 | 371.18 | 424.74 | 379.4 | 277.37 | 329.67 | 434.04 |
| Nª inventory | RlaPh029 | RlaPh034 | RlaPh035 | RlaPh036 | RlaPh050 | RlaPh032 | RlaPh041 | RlaPh011 | RlaPh014 | RlaPh016Typus | RlaPh024 | RlaPh025 | RlaPh060 | RlaPh018 | RlaPh011 |
| Nº Orden | 1 | 2 | 3 | 4 | 6 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 16 | 17 | 18 |
| Characteristic species | |||||||||||||||
| Pinus halepensis Mill. | 3 | 3 | 4 | 3 | 4 | 3 | 3 | 3 | 3 | 3 | 5 | 3 | 3 | 3 | 2 |
| Rhamnus ladedroi Rivas-Mart. & J.M. Pizarro | 1 | 2 | 1 | 1 | 1 | + | 1 | + | 1 | + | + | + | 1 | + | |
| Juniperus oxycedrus subsp. oxycedrus L. | 2 | 2 | 1 | 2 | 2 | 2 | 2 | ||||||||
| Daphne gnidium L. | 1 | 1 | 1 | 2 | 1 | 2 | |||||||||
| Quercus coccifera L. subsp. coccifera | 2 | 2 | 2 | 2 | |||||||||||
| Olea europaea var. sylvestris (Mill.) Lehr | 2 | 2 | 2 | 2 | |||||||||||
| Rhamnus myrtifolia Willk. | 1 | 1 | 1 | 1 | 1 | ||||||||||
| Quercus rotundifolia Lam. | 1 | 2 | 1 | 1 | |||||||||||
| Chamaerops humilis L. | 2 | 2 | 3 | ||||||||||||
| Asparagus acutifolius L. | 2 | 2 | |||||||||||||
| Rhamnus alaternus L. subsp. alaternus | 2 | ||||||||||||||
| Rhamnus alaternus subsp. parvifolia Arcang. | + | ||||||||||||||
| Rhamnus velutina Boiss. subsp. velutina | 2 | ||||||||||||||
| Juniperus phoenicea L. subsp. phoenicea | 1 | ||||||||||||||
| Pinus pinaster Aiton | 1 | ||||||||||||||
| Aristolochia baetica L. | 2 | ||||||||||||||
| Buxus balearica Lam. | 2 | ||||||||||||||
| Carex halleriana Asso. | 1 | 1 | |||||||||||||
| Pistacia lentiscus L. | 2 | 2 | |||||||||||||
| Pistacia terebinthus L. | + | ||||||||||||||
| Quercus faginea subsp. alpestris (Boiss.) Maire | 1 | ||||||||||||||
| Rubia peregrina L. subsp. peregrina | 1 | ||||||||||||||
| Teucrium fruticans L. | + | ||||||||||||||
| Companion species | |||||||||||||||
| Rosmarinus officinalis L. | 2 | 2 | 1 | 2 | 1 | 2 | 2 | 2 | 2 | 2 | 2 | ||||
| Ulex parviflorus subsp. parviflorus Pourr. | 2 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | |||||||
| Brachypodium retusum subsp. retusum (Pers.) P. Beauv. | 2 | 2 | 2 | 2 | 2 | 2 | 2 | ||||||||
| Macrochloa tenacissima (L.) Kunth | 2 | 2 | 3 | ||||||||||||
| Cistus albidus L. | 2 | 2 | 2 | ||||||||||||
| Phlomis purpurea L. subsp. purpurea | 2 | 2 | 2 | ||||||||||||
| Genista scorpius (L.) DC. in Lam. & DC. | 2 | 2 | |||||||||||||
| Echinospartum boissieri (Spach) Rothm. | 2 | 1 | |||||||||||||
| Lavandula latifolia Medik. | 1 | 1 | |||||||||||||
| Lavandula stoechas L. | 3 | ||||||||||||||
| Helichrysum stoechas (L.) Moench | 1 | ||||||||||||||
| Genista umbellata subsp. equisetiformis (Spach) Rivas Goday & Rivas Mart. | 2 | ||||||||||||||
| Cistus clusii subsp. clusii Dunal | 1 | ||||||||||||||
| Astragalus incanus L. subsp. incanus | + | ||||||||||||||
| Berberis hispanica Boiss. & Teut. | 1 | ||||||||||||||
| Brachypodium retusum subsp. boissieri (Nyman) Romero García | 2 | ||||||||||||||
| Bupleurum fruticescens subsp. spinosum (Gouan) O. Bolòs & Vigo | 1 | ||||||||||||||
| Cistus salviifolius L. | 2 | ||||||||||||||
| Dactylis glomerata subsp. hispanica (Roth) Nyman | 2 | ||||||||||||||
| Erica multiflora L. | 2 | ||||||||||||||
| Festuca indigesta subsp. hackeliana | 1 |
References
- Do Amaral Franco, J. Pinus L. in Flora Ibéria Vol I; Castroviejo, S., Lain, M., López González, G., Monserrat, P., Muñoz-Garmendia, F., Paiva, J., Villar, L., Eds.; Real Jardín Botánico, CSIC: Madrid, Spain, 1986. [Google Scholar]
- Bolòs, O.; de Vigo, J.; Masalles, R.M.; Ninot, J.M. Manual Dels Paisos Catalans; Portic: Barcelona, Italy, 1990. [Google Scholar]
- Fernández-Galiano, E. Pasado, Presente y Futuro de los Boques de la Península Ibérica. Acta Bot. Malacit. 1990, 15, 135–143. [Google Scholar] [CrossRef]
- Postigo-Mijarra, J.M.; Manzaneque, F.G.; Juaristi, C.M.; Zazo, Z. Palaeoecological significance of Late Pleistocene pine macrofossils in the lower Guadalquivir Basin (Doñana natural park, southwestern Spain). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 295, 332–343. [Google Scholar] [CrossRef]
- Fiori, A. Nuova Flora Analitica D´italia; Library New York: Firenze, Italy, 1923; Volume I. [Google Scholar]
- Conti, F.; Abbate, G.; Alessandrini, A.; Blasi, C. (Eds.) An Annotated Checklist of the Italian Vascular Flora; Ministero dell´Ambiente de della Tutela del Territorio-Direzione per la Protezione della Natura, Dipartimento de BiologiaVegetale-Università degli Studi di Roma “La Sapienza”: Roma, Italy, 2005. [Google Scholar]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.N.M.G.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Galasso, G.; Conti, F.; Peruzzi, L.; Ardenghi, N.M.G.; Banfi, E.; Celesti-Grapow, L.; Albano, A.; Alessandrini, A.; Bacchetta, G.; Ballelli, S.; et al. An updated checklist of the vascular flora alien to Italy. Plant Biosyst. 2018, 152, 556–592. [Google Scholar] [CrossRef]
- Torres, J.A.; García Fuentes, A.; Salazar, C.; Cano, E.; Valle, F. Caracterización de los Pinares de Pinus halepensis Mill. en el sur de la Península Ibérica. Ecol. Mediterránea 1999, 25, 135–146. [Google Scholar] [CrossRef]
- Maire, R. Flore de l´Afrique de Nort, 1st ed.; Paul Lechevalier: Paris, Frence, 1987; Volume 1. [Google Scholar]
- Ighbareyeh, M.M.H.; Cano, E.; Cano-Ortiz, A.; Ighbareyeh, J.; Suliemieh, A.A.A. Phytosociology with other Characteristic Biologically and Ecologically of Plant in Palestine. Am. J. Plant Sci. 2014, 5, 3104–3118. [Google Scholar] [CrossRef] [Green Version]
- Quézel, P.; Barbero, M.; Benabid, A.; Rivas-Martínez, S. Contribution a l´etude des groupements foretiers et pre-forestiers du Maroc Oriental. Studa Bot. 1992, 10, 57–90. [Google Scholar]
- Parra, I. Análisis polínico del sondaje C.A.L. 81-I. (Casablanca-Almenara, Prov. Castellón). In Actas del IV Simposio de Palinología Española; APLE: Barcelona, Spain, 1983. [Google Scholar]
- Rivera, B.; Obón, C. Macrorrestos vegetales de los yacimientos 433–445 de la comarca noroeste en los inicios de la Edad de los Metales. In El cambio Cultural del IV al II Milenios a.C. en la Comarca Noroeste de Murcia; López, P., Ed.; CSIC: Madrid, Spain, 1991; Volume I, pp. 239–246. [Google Scholar]
- Ariza, M.O.R. Human-plant relationships during the Copper & Bronze Ages in the Baza & Guadix basins (Granada, Spain). Bull. Société Bot. France. Actual. Bot. 1992, 139, 451–464. [Google Scholar]
- Ariza, M.O.R.; Vernet, J.L. Premiers resutats peléoécologiques de l´établissement chalcothique de Los Millares (Santa Fé de Mondujar, Almería, Espagñe) d´après l´analyse anthracologique. In Lind Deya Conference of Prehistory 1–13; BAR International Series; Weldren, W.H., Ensenyal, J.A., Kennard, R.C., Eds.; BAR Publishing: Oxford, UK, 1991; Volume I. [Google Scholar]
- Ariza, M.O.R.; Aguayo, P.; Moreno, F. The environment in the Ronda Basin (Málaga, Sapin) during recent prehistory based on an anthracological study of Old Ronda. Bull. Société Bot. France. Actual. Bot. 1992, 139, 715–725. [Google Scholar]
- Rivas-Martínez, S.; Asensi, A.; Valle, F.; Garretas, B.D. Mapa de series, geoseries y geopermaseries de vegetación de España. Parte II. Itinera Geobot. 2011, 18, 425–800. [Google Scholar]
- Pesaresi, S.; Bioindi, E.; Vagge, I.; Galdenzi, D.; Casavecchia, S. The Pinus halepensis Mill. Forests in the central-eastern European Mediterranean basin. Plant Biosyst. 2017, 151, 512–529. [Google Scholar] [CrossRef]
- Bucci, G.; Anzidei, M.; Madaghiele, A.; Vendramin, G.G. Detection of haplotypic variation and natural hybridization in halepensis-complex pine species using chloroplast simple sequence repeat (SSR) markers. Mol. Ecol. 1998, 7, 1633–1643. [Google Scholar] [CrossRef]
- Panetsos, C.P. Natural hybridization between Pinus halepensis and Pinus brutia. Silvae Genet. 1975, 24, 163–168. [Google Scholar]
- Farjon, A. Pinus brutia. The IUCN Red List of Threatened Species 2013: Et42347a2974345. Available online: https://www.iucnredlist.org/ (accessed on 14 January 2020).
- Daniels, R.R.; Taylor, R.S.; Serra-Varela, M.J.; Vendramin, G.G.; González-Martínez, S.C.; Grivet, D. Inferring selection in instances of long-range colonization: The Aleppo pine (Pinus halepensis) in the Mediterranean Basin. Mol. Ecol. 2018, 27, 3331–3345. [Google Scholar] [CrossRef] [PubMed]
- de la Campa Fayos, J.M.Á. Vegetació del Massís del Port.; Colúlecció Pius Font i Quer, 3; Institut d’Estudis Ilerdencs: Lleida, Spain, 2004; p. 459. [Google Scholar]
- Bolòs, O.; de et Molinier, R. Recherches phytosociologiques dans l’île de Majorque. Collect. Bot. 1958, V, 699–865. [Google Scholar]
- Braun-Blanquet, J. i Bolòs, O. de. Les groupements végétaux du bassin moyen de l’Ebre et leur dynamisme. An. Estac. Exp. Aula Dei. 1957, 5, 1–266. (In Saragossa) [Google Scholar]
- Cabezudo, B.; Pérez-Latorre, A.V.; Navas, P.; Gil, Y.; Navas, D. Parque natural de la Sierra de las Nieves. ‘Cartografía y evaluación de la flora y vegetación’. In Memoria de Investigación; Departamento de Biología Vegetal, Universidad de Málaga: Málaga, Spain, 1998; p. 367. [Google Scholar]
- Olmedo, J.A. Análisis Biogeográfico y Cartografía de la Vegetación de la Sierra de Baza (Provincia de Granada). Ph.D. Thesis, Universidad de Granada, Granada, Spain, 2011; p. 783. [Google Scholar]
- Latorre, A.V.P.; Navas, P.; Navas, D.; Gil, Y.; Cabezudo, B. Datos sobre la flora y la vegetación de la Serranía de Ronda (Málaga, España). Acta Bot. Malacit. 1998, 23, 149–191. [Google Scholar] [CrossRef] [Green Version]
- Latorre, A.V.P.; Fernández, D.N.; Gavira, O.; Caballero, G.; Cabezudo, B. Vegetación del Parque Natural de Las Sierras Tejeda, Almijara y Alhama (Málaga-Granada, España). Acta Bot. Malacit. 2004, 29, 117–190. [Google Scholar] [CrossRef]
- Pérez-Latorre, A.V.; Casimiro, F.; Cabezudo, B. Flora y vegetación de la sierra de Alcaparaín (Málaga, España). Acta Bot. Malacit. 2015, 40, 107–156. [Google Scholar] [CrossRef]
- Pérez-Latorre, A.V.; Casimiero, F.; Sánchez, J.G.; Cabezudo, B. Flora y vegetación del Paraje Natural Desfiladero de los Gaitanes y su entorno (Málaga). Acta Bot. Malacit. 2014, 39, 129–177. [Google Scholar] [CrossRef]
- Llueca, C.; Udias, S.L. Estudio de las Comunidades Vegetales del Valle del Mijares (Teruel); Servicio de Conservación de la Biodiversidad; Gobierno de Aragón: Zaragoza, Spain, 2004; pp. 1–122. [Google Scholar]
- Gómez-Mercado, F. Vegetación y flora de la Sierra de Cazorla. Guineana 2011, 17, 1–481. [Google Scholar]
- Cantos, R.M.; Franzi, A.V.; Ariza, F.J.A. Flora y Vegetación del Tramo Medio del Valle del Río Júcar (Albacete); Instituto de estudios albacetenses ‘Don Juan Manuel’ de la excma: Alicante, Spain, 2008; p. 663. [Google Scholar]
- Badia, R.P. Flora Vascular y Vegetación de la Comarca de la Marina Alta (Alicante); Colección Técnica 16; Instituto de Cultura Juan Gil Albert: Alicante, Spain, 1997. [Google Scholar]
- Martínez, S.R.; Costa, M.; Soriano, P.; Badia, R.P.; Llorens, L.; Roselló, J.A. Datos sobre el paisaje vegetal de Mallorca e Ibiza (Islas Baleares, España). Itinera Geobot. 1992, 6, 5–98. [Google Scholar]
- Rivas-Martínez, S.; Pizarro, J.M. Taxonomical system advance to Rhamnus, L. & Frangula Mill. of Iberian Peninsula and Balearic Islands. Int. J. Geobot. Res. 2011, 1, 55–78. [Google Scholar]
- Rojo, M.P.R.; Úbeda, J.R.; Badia, R.P. La diversidad vegetal de La Manchuela Conquense: Una comarca manchega con influencias setabenses y celtibérico-alcarreñas. Lazaroa 2009, 30, 35–49. [Google Scholar]
- López, A.M.R.i. Estudi Fitogeogràfic de les Comarques Catalanes Compreses Entre Els Ports de Beseit, el Riu Ebre i Els Límits Aragonesos. Ph.D. Thesis, Universitat de Barcelona, Barcelona, Spain, 1986. [Google Scholar]
- SIVIM. Sistema de Información de la Vegetación Ibérica y Macaronésica. Available online: http://www.sivim.info/sivi/PissarraUTM.jsp?ruta=f2.1&utm=30SXK60 (accessed on 20 February 2022).
- Rivas-Martínez, S.; Penas, A.; González, T.E.D.; Cantó, P.; del Río, S.; Costa, C.; Herrero, L.; Molero, J. Biogeographic Units of the Iberian Peninsula and Balearic Islands to District Level. A Concise Synopsis. In The Vegetation of the Iberian Peninsula, Plant and Vegetation 12; Loidi, J., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 131–188. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Rivas-Sáenz, S.; Penas, A.S. Worldwide bioclimatic classification system. Glob. Geobot. 2011, 1, 1–634. [Google Scholar]
- Theurillat, J.P.; Willner, W.; Fernández-González, F.; Bültmann, H.; Čarni, A.; Gigante, D.; Mucina, L.; Weber, H. International Code of Phytosociological Nomenclature. 4th edition. Appl. Veg. Sci. 2020, 24, e12491. [Google Scholar] [CrossRef] [Green Version]
- Rivas-Martínez, S. Mapa de series, geoseries y geopermaseries de vegetación de España. Parte I. Itinera Geobot. 2007, 17, 5–436. [Google Scholar]
- De Marco, G.; Veri, L.; Caneva, G. Analisi Fitosociologica, Cartografiad ella Vegetazione e Trasformacioni Ambientali nel Periodo 1955–1981 delle Isole Tremiti (Adriatico Centro-Meridionale). Ann. Di Bot. 1984, LX, 17–47. [Google Scholar]
- Loisel, R. Séries de vegetation propres, en Provence, aux massifs des Maures et de l´Estérel (ripisilves exclues). Bull. Société Bot. Fr. 1971, 118, 203–236. [Google Scholar] [CrossRef]
- Mesa, J.M.; Raya, F.P. Estudio fitosociológico de los sabinares de Juniperus phoenicea L. en el sector Malacitano-Almijarense (provincia corológica Bética). Lazaroa 1987, 7, 301–306. [Google Scholar]
- Ortiz, A.C.; Gomes, C.J.P.; Musarella, C.M.; Cano, E. Expansion of the Juniperus genus due to anthropic activity. In Old-Growth Forest and Coniferous Forests; Weber, R.P., Ed.; Nova Science Publishers: New York, NY, USA, 2015; pp. 55–65. [Google Scholar]
- Fuentes, J.C.P.; Cano-Ortiz, A.; Musarella, C.M.; Gomes, C.J.P.; Spampinato, G.; Cano, E. Rupicolous habitats of interest for conservation in the central-southern Iberian Peninsula. Plant Sociol. 2017, 54, 29–42. [Google Scholar]
- Vila-Viçosa, C.; Vázquez, F.M.; Mendes, P.; Del Rio, S.; Musarella, C.; Cano-Ortiz, A.; Meireles, C. Syntaxonomic update on the relict groves of Mirbeck’s oak (Quercus canariensis Willd. and Q. marianica C. Vicioso) in southern Iberia. Plant Biosyst. 2015, 149, 512–526. [Google Scholar] [CrossRef]
- Barrile, V.; Malerba, A.; Fotia, A.; Calabrò, F.; Bernardo, C.; Musarella, C. Quarries Renaturation by Planting Cork Oaks and Survey with UAV. In New Metropolitan Perspectives; NMP 2020. Smart Innovation, Systems and Technologies; Bevilacqua, C., Calabrò, F., Spina, L.D., Eds.; Springer: Cham, Switzerland, 2015; Volume 178. [Google Scholar] [CrossRef]
- Cano, E.; Musarella, C.M.; Ortiz, A.C.; Fuentes, J.C.P.; Gomes, C.J.P. Vegetation series as a basis for habitats and species conservation: Methodological aspects. Botanique 2016, 1, 21–26. [Google Scholar]
- Cano, E.; Musarella, C.M.; Ortiz, A.C.; Fuentes, J.C.P.; Torres, A.R.; Del Río González, S.; Gomes, C.J.P.; Quinto-Canas, R.; Spampinato, G. Geobotanical Study of the Microforests of Juniperus oxycedrus subsp. badia in the Central and Southern. Sustainability 2019, 11, 1111. [Google Scholar] [CrossRef] [Green Version]
- Spampinato, G.; Crisarà, R.; Cannavò, S.; Musarella, C.M. Phytotoponims of Southern Calabria: A tool for the analysis of the landscape and its transformations. Atti Della Soc. Toscana Di Sci. Nat. Mem. Ser. B 2017, 124, 61–72. [Google Scholar]
- Musarella, C.M.; Mendoza-Fernández, A.J.; Mota, J.F.; Alessandrini, A.; Bacchetta, G.; Brullo, S.; Caldarella, O.; Ciaschetti, G.; Conti, F.; Martino, L.; et al. Checklist of gypsophilous vascular flora in Italy. PhytoKeys 2018, 103, 61–82. [Google Scholar] [CrossRef]
- Pérez-García, F.; Akhani, H.; Parsons, R.; Silcock, J.; Kurt, L.; Özdeniz, E.; Spampinato, G.; Musarella, C.; Salmerón-Sánchez, E.; Sola, F.; et al. A first inventory of gypsum flora in the Palearctic and Australia. Mediterr. Bot. 2018, 39, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Biondi, E.; Blasi, C.; Allegrezza, M.; Anzellotti, I.; Azzella, M.M.; Carli, E.; Casavecchia, S.; Copiz, R.; Del Vico, E.; Facioni, L.; et al. Plant communities of Italy: The Vegetation Prodrome. Plant Biosyst. 2014, 148, 728–814. [Google Scholar] [CrossRef] [Green Version]
- Meddour, R.; Meddour-Sahar, O.; Zeraia, L.; Mucina, L. Syntaxonomic synopsis of the forest and tall scrub vegetation of Northern Algeria. Lazaroa 2017, 38, 127–163. [Google Scholar] [CrossRef] [Green Version]
- Mucina, L.; Dengler, J.; Bergmeier, E.; Carni, A.; Dimopoulos, P.; Jahn, R.; Matevski, V. New and validated high-rank syntaxa from Europe. Lazaroa 2009, 30, 267–276. [Google Scholar]
- Rivas-Martínez, S.; Díaz, T.E.; Fernández-González, F.; Izco, J.; Loidi, J.; Lousa, M.; Penas, A. Vascular Plant Communities of Spain and Portugal. Itinera Geobot. 2002, 15, 433–922. [Google Scholar]
- Ortiz, A.C.; Biondi, E.; Gomes, C.J.P.; del Río González, S.; Cano, E. Soil and phytosociological caracterisation of grasslands in the western mediterranean. Am. J. Plant Sci. 2014, 5, 3213–3240. [Google Scholar] [CrossRef] [Green Version]
- Bonari, G.; Fernández-González, F.; Çoban, S.; Monteiro-Henriques, T.; Bergmeier, E.; Didukh, Y.P.; Xystrakis, F.; Angiolini, C.; Chytrý, K.; Acosta, A.T.; et al. Classification of the Mediterranean lowland to submontane pine forest vegetation. Appl. Veg. Sci. 2021, 24, e12544. [Google Scholar] [CrossRef]



| Variable | Factor 1 | Factor 2 | Variable | Factor 1 | Factor 2 |
|---|---|---|---|---|---|
| MAX_January | 0.9427 | 0.0131 | MIN_July | 0.9465 | 0.0996 |
| MAX_February | 0.9547 | 0.2017 | MIN_August | 0.9307 | 0.0102 |
| MAX_March | 0.9028 | 0.3500 | MIN_September | 0.9324 | 0.1066 |
| MAX_October | 0.9538 | 0.1694 | MIN_October | 0.9141 | 0.1079 |
| MAX_November | 0.9630 | 0.0933 | MIN_December | 0.9132 | −0.0326 |
| MED_January | 0.9515 | 0.0240 | PE | 0.9831 | 0.1891 |
| MED_February | 0.9790 | 0.1088 | PE2 | 0.9095 | 0.0257 |
| MED_March | 0.9642 | 0.2419 | PRE_December | −0.0853 | −0.9250 |
| MED_April | 0.9404 | 0.3390 | PRE_January | −0.0155 | −0.9076 |
| MED_May | 0.9112 | 0.3235 | Psw | −0.1292 | −0.9072 |
| MED_September | 0.9629 | 0.1047 | Ptrw | −0.1078 | −0.9238 |
| MED_October | 0.9527 | 0.1349 | Pw | −0.1078 | −0.9238 |
| MED_November | 0.9617 | 0.1111 | T | 0.9928 | 0.1393 |
| MED_December | 0.9302 | −0.0865 | Tmax | 0.9026 | 0.0616 |
| MIN_January | 0.9162 | 0.0326 | Tpw | 0.9647 | 0.0083 |
| MIN_February | 0.9385 | 0.0143 | Tpw1 | 0.9531 | −0.0004 |
| MIN_March | 0.9575 | 0.1315 | Tpw2 | 0.9579 | −0.0209 |
| MIN_April | 0.9474 | 0.2147 | Ts | 0.9263 | 0.1350 |
| MIN_May | 0.9564 | 0.2073 | Ts2 | 0.9102 | 0.0049 |
| MIN_June | 0.9579 | 0.2275 | Itc | 0.9768 | 0.0459 |
| Variable | (Observed Value) K | K (Critical Value) | GDL | p-Value (Bilateral) | Alpha |
|---|---|---|---|---|---|
| MAX_JANUARY | 130.52 | 18.31 | 10 | <0.0001 | 0.05 |
| MAX_FEBRUARY | 109.49 | 18.31 | 10 | <0.0001 | 0.05 |
| MAX_MARCH | 101.72 | 18.31 | 10 | <0.0001 | 0.05 |
| MAX_OCTOBER | 106.67 | 18.31 | 10 | <0.0001 | 0.05 |
| MAX_NOVEMBER | 111.62 | 18.31 | 10 | <0.0001 | 0.05 |
| MIN_JANUARY | 116.01 | 18.31 | 10 | <0.0001 | 0.05 |
| MIN_FEBRUARY | 107.89 | 18.31 | 10 | <0.0001 | 0.05 |
| MIN_MARCH | 111.94 | 18.31 | 10 | <0.0001 | 0.05 |
| MIN_APRIL | 119.85 | 18.31 | 10 | <0.0001 | 0.05 |
| MIN_MAY | 104.48 | 18.31 | 10 | <0.0001 | 0.05 |
| MIN_JUNE | 104.54 | 18.31 | 10 | <0.0001 | 0.05 |
| MIN_JULY | 103.18 | 18.31 | 10 | <0.0001 | 0.05 |
| MIN_AGOST | 107.26 | 18.31 | 10 | <0.0001 | 0.05 |
| MIN_SEPTEMBER | 118.58 | 18.31 | 10 | <0.0001 | 0.05 |
| MIN_OCTOBER | 115.00 | 18.31 | 10 | <0.0001 | 0.05 |
| MIN_DICEMBER | 106.77 | 18.31 | 10 | <0.0001 | 0.05 |
| MED_JANUARY | 122.17 | 18.31 | 10 | <0.0001 | 0.05 |
| MED_FEBRERO | 113.21 | 18.31 | 10 | <0.0001 | 0.05 |
| MED_MARZO | 111.55 | 18.31 | 10 | <0.0001 | 0.05 |
| MED_ABRIL | 114.40 | 18.31 | 10 | <0.0001 | 0.05 |
| MED_MAYO | 97.07 | 18.31 | 10 | <0.0001 | 0.05 |
| PRE_ENERO | 135.89 | 18.31 | 10 | <0.0001 | 0.05 |
| PRE_DICIEMBRE | 122.17 | 18.31 | 10 | <0.0001 | 0.05 |
| PE | 106.73 | 18.31 | 10 | <0.0001 | 0.05 |
| PE2 | 118.65 | 18.31 | 10 | <0.0001 | 0.05 |
| Ic | 99.15 | 18.31 | 10 | <0.0001 | 0.05 |
| Io | 75.88 | 18.31 | 10 | <0.0001 | 0.05 |
| It | 118.89 | 18.31 | 10 | <0.0001 | 0.05 |
| Itc | 117.39 | 18.31 | 10 | <0.0001 | 0.05 |
| AbPh | PlPh | TvPh | RoPh | SsPh | RlPh | |
|---|---|---|---|---|---|---|
| Alkanna graeca subsp. baeotica | x | |||||
| Alyssum euboeum | x | |||||
| Centaurea montoudi | x | |||||
| Jurinea mollis subsp. anatolica | x | |||||
| Onosma euboica | x | |||||
| Quercus trojana subsp. euboica | x | |||||
| Scorzonera serpentinica | x | |||||
| Rosmarinus officinalis | x | x | x | |||
| Daphne gnidium | x | x | ||||
| Micromeria graeca | x | |||||
| Cistus monspeliensis | x | |||||
| Coronilla valentina | x | |||||
| Hyppocrepis emerus subsp. emoroides | x | |||||
| Prasium majus | x | |||||
| Fumana thymifolia | x | |||||
| Juniperus macrocarpa | x | |||||
| Phyllirea latifolia | x | x | ||||
| Ruscus aculeatus | x | |||||
| Teucrium polium | x | |||||
| Thymus striatus | x | |||||
| Convolvulus elegantissimus | x | |||||
| Helianthemum jonium | x | |||||
| Alkanna tinctoria | x | |||||
| Cistus albidus | x | |||||
| Juniperus oxycedrus | x | x | ||||
| Thymus vulgaris | x | |||||
| Coriaria myrtiffolia | x | |||||
| Aphyllanthes monspeliensis | x | |||||
| Coris monspeliensis | x | |||||
| Eria arborea | x | x | ||||
| Arbutus unedo | x | x | ||||
| Myrtus communis | x | x | ||||
| Clematis flammula | x | |||||
| Calicotome spinosa | x | x | ||||
| Ruta angustifolia | x | |||||
| Phagnalon sordidum | x | |||||
| Dorycnium pentphyllum subsp. pentaphyllum | x | x | ||||
| Ampelodesmos mauritanicus | x | |||||
| Clematis cirrhosa | x | |||||
| Calicotome villosa | x | x | ||||
| Phillyrea angustifolia | x | x | ||||
| Genista tyrrhena | x | |||||
| Genista ephedroides | x | |||||
| Genista corsica | x | |||||
| Quercus coccifera s.l. | x | x | ||||
| Asparagus albus | x | |||||
| Chamaerops humilis | x | |||||
| Sarcopoterium spinosum | x | |||||
| Carlina graeca | x | |||||
| Campanula spatulata | x | |||||
| Hypericum empetrifolium | x | |||||
| Anthyllis hermanniae subsp. hermanniae | x | |||||
| Rhamnus lycioides s.l. | x | |||||
| Cercis siliquastrum | x | |||||
| Cistus creticus | x | |||||
| Arbutus andrachne | x | |||||
| Rhamnus myrtifolia | x | |||||
| Rhamnus bourgaena | x | |||||
| Rhamnus lycioides | x | |||||
| Juniperus phoenicea | x | |||||
| Rubia longifolia | x | |||||
| Rhamnus alaternus | x | |||||
| Asparagus horridus | x | |||||
| Ephedra fragilis | x | |||||
| Rhamnus oleoides subsp. angustifolia | x | |||||
| Rhamnus lycioides subsp. laderoi | x | |||||
| Stipa tenacissima (com. terr.) | x | |||||
| Ulex parviflorus (com. terr.) | x |
| Characteristic Species | AuPh | CsPh | JtPh | BrPh | EnPh | RlaPh | RaPh | RlPh | EfPh | QcPh | RanPh |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Pinus halepensis Mill. | V | V | V | V | V | V | V | V | V | V | V |
| Juniperus oxycedrus L. subsp. oxycedrus | IV | IV | IV | IV | II | I | V | III | V | II | |
| Pistacia lentiscus L. | V | V | IV | II | I | I | I | I | II | ||
| Rhamnus lycioides L. subsp. lycioides | IV | III | II | II | II | I | IV | II | IV | I | |
| Juniperus phoenicea L. subsp. phoenicea | III | III | IV | IV | I | V | II | I | I | ||
| Quercus coccifera L. subsp. coccifera | IV | V | V | V | II | I | V | I | |||
| Asparagus acutifolius L. | IV | I | II | I | I | I | I | I | |||
| Quercus rotundifolia Lam. | I | I | I | I | I | I | I | I | |||
| Olea europaea var. sylvestris (Mill.) Lehr | I | III | V | I | I | I | I | ||||
| Phillyrea angustifolia L. | III | I | I | I | I | ||||||
| Lonicera implexa Aiton | IV | I | |||||||||
| Rhamnus alaternus subsp. parvifolia Arcang. | IV | I | I | ||||||||
| Arbutus unedo L. | IV | II | I | ||||||||
| Bupleurum rigidum L. | III | I | III | ||||||||
| Pistacia terebinthus L. | I | I | I | III | |||||||
| Rubia peregrina subsp. longifolia (Poir.) O. Bolòs | II | II | IV | ||||||||
| Smilax aspera L. | I | I | |||||||||
| Clematis flammula L. | I | ||||||||||
| Viburnum tinus L. | I | ||||||||||
| Carex halleriana Asso. | III | IV | I | I | I | I | I | I | |||
| Asparagus horridus L. in Murray | II | IV | I | I | II | ||||||
| Daphne gnidium L. | I | II | I | II | II | I | |||||
| Chamaerops humilis L. | I | I | I | ||||||||
| Coronilla juncea L. | I | I | I | ||||||||
| Ephedra fragilis Desf. | II | II | I | I | |||||||
| Rhamnus alaternus L. subsp. alaternus | I | I | I | I | |||||||
| Ceratonia siliqua L. | III | ||||||||||
| Osyris quadripartita Salzm. ex Decne. | II | ||||||||||
| Rhamnus lycioides L. subsp. oleoides (L.) Maire in Jahand. & Maire | III | ||||||||||
| Jasminum fruticans L. | I | ||||||||||
| Calicotome spinosa (L.) Link | I | ||||||||||
| Asparagus albus L. | I | I | I | ||||||||
| Juniperus phoenicea subsp. turbinata (Guss.) Nyman | V | ||||||||||
| Phillyrea latifolia subsp. rodriguezii (P. Monts.) Romo | II | ||||||||||
| Arisarum simorrhinum Durieu | II | ||||||||||
| Phillyrea latifolia L. subsp. latifolia | I | ||||||||||
| Rubia peregrina L. subsp. peregrina | IV | III | I | I | |||||||
| Arctostaphylos uva-ursi (L.) Spreng. | II | I | |||||||||
| Ephedra major Host subsp. nebrodensis | I | I | |||||||||
| Juniperus thurifera L. | I | ||||||||||
| Quercus faginea subsp. alpestris (Boiss.) Nyman | I | ||||||||||
| Rhamnus myrtifolia Willk. | I | II | I | ||||||||
| Aristolochia baetica L. | I | I | |||||||||
| Rhamnus lycioides L. subsp. laderoi Rivas Mart. & J.M. Pizarro | III | ||||||||||
| Pinus pinaster Aiton | I | ||||||||||
| Buxus balearica Lam. | I | ||||||||||
| Quercus faginea subsp. faginea Lam. | I | ||||||||||
| Rhamnus velutina subsp. almeriensis Rivas Mart. & J.M. Pizarro | II | ||||||||||
| Osyris alba L. | I | ||||||||||
| Teucrium fruticans L. | II | ||||||||||
| Bupleurum fruticosum L. | I | ||||||||||
| Buxus sempervirens L. | I | ||||||||||
| Rhamnus velutina subsp. velutina Boiss. | I | ||||||||||
| Rhamnus oleoides L. subsp. angustifolia (Lange ex Willk.) Rivas Mart. & J.M. Pizarro | I | IV | |||||||||
| Crataegus monogyna subsp. brevispina (Kunze) Franco | I | ||||||||||
| Compañeras | |||||||||||
| Rosmarinus officinalis L. | V | IV | III | IV | IV | II | IV | III | III | III | |
| Cistus clusii subsp. clusii Dunal | III | I | I | II | I | I | I | I | I | ||
| Brachypodium retusum (Pers.) P. Beauv. | V | V | II | V | IV | III | |||||
| Genista scorpius (L.) DC. in Lam. & DC. | IV | II | I | II | I | I | III | I | III | ||
| Dorycnium pentaphyllum Scop. | II | II | I | I | I | ||||||
| Retama sphaerocarpa (L.) Boiss. | I | I | I | I | I | ||||||
| Erica multiflora L. | V | V | I | I | |||||||
| Linum suffruticosum L. | V | I | I | IV | |||||||
| Thymus vulgaris subsp. vulgaris L. | III | III | II | I | |||||||
| Centaurea linifolia L. | I | IV | |||||||||
| Cytisus fontanesii subsp. fontanesii Spach | I | I | I | ||||||||
| Helianthemum marifolium (L.) Mill. | V | I | |||||||||
| Stipa offneri Breistr. | I | II | |||||||||
| Thymelaea tinctoria (Pourr.) End L. | III | I | III | ||||||||
| Euphorbia characias L. | I | ||||||||||
| Ulex parviflorus subsp. parviflorus Pourr. | IV | III | II | I | I | I | |||||
| Macrochloa tenacissima (L.) Kunth | III | I | III | II | V | III | II | ||||
| Anthyllis cytisoides L. | II | II | I | I | I | ||||||
| Helictotrichon filifolium subsp. filifolium (Lag.) Henrard | II | II | |||||||||
| Bupleurum fruticescens Loefl. ex L. | I | I | III | I | |||||||
| Urginea maritima (L.) Baker | I | II | |||||||||
| Cistus albidus L. | I | I | I | III | I | I | |||||
| Globularia alypum L. | II | I | |||||||||
| Piptatherum miliaceum subsp. miliaceum (L.) Coss. | I | II | |||||||||
| Teucrium carolipaui Vicioso ex Pau | I | ||||||||||
| Teucrium homotrichum (Font Quer) Rivas Mart. | I | ||||||||||
| Arisarum vulgare Targ.-Tozz. | I | ||||||||||
| Lavandula dentata L. | I | ||||||||||
| Hippocrepis valentina Boiss. | I | ||||||||||
| Melica minuta L. | I | ||||||||||
| Viola arborescens L. | I | ||||||||||
| Sedum sediforme (Jacq.) Pau | II | I | I | I | |||||||
| Cistus salviifolius L. | I | I | |||||||||
| Rosmarinus officinalis L. subsp. palaui (O. Bolòs & Molinier) Malagarriga | II | ||||||||||
| Ruta angustifolia Pers. | II | ||||||||||
| Teucrium capitatum subsp. majoricum (Rouy) T. Navarro & Rosúa | II | ||||||||||
| Ampelodesmos mauritanica (Poir.) T. Durand & Schinz | I | ||||||||||
| Arum sagittifolium L. | I | ||||||||||
| Piptatherum coerulescens (Desf.) P. Beauv. | I | ||||||||||
| Suaeda vera Forssk. ex J.F. Gmel. | I | ||||||||||
| Thymus zygis subsp. gracilis (Boiss.) R. Morales | II | I | I | II | I | II | |||||
| Helichrysum stoechas (L.) Moench | I | I | I | II | |||||||
| Helianthemum origanifolium subsp. origanifolium (Lam.) Pers. | I | I | I | ||||||||
| Staehelina dubia L. | I | I | I | ||||||||
| Koeleria vallesiana subsp. vallesiana (Honck.) Gaudin | I | I | I | ||||||||
| Lygeum spartum L. | I | I | I | ||||||||
| Thesium humifusum DC. | I | I | |||||||||
| Asphodelus cerasiferus J. Gay | II | I | I | ||||||||
| Fumana ericoides (Cav.) Gand. in Magnier | I | I | I | ||||||||
| Helianthemum myrtifolium (Lam.) Samp. | III | III | |||||||||
| Stipa orientalis Trin. | I | IV | |||||||||
| Aphyllanthes monspeliensis L. | I | II | |||||||||
| Aristolochia pistolochia L. | I | II | |||||||||
| Avenula bromoides (Gouan) H. Scholz | I | III | |||||||||
| Coronilla minima L. subsp. lotoides (W.D.J. Koch) Nyman | I | III | |||||||||
| Cistus libanotis L. | I | II | |||||||||
| Echinops ritro L. | I | III | |||||||||
| Euphorbia minuta Loscos & J. Pardo | I | III | |||||||||
| Coris monspeliensis L. | I | I | |||||||||
| Anacamptis pyramidalis (L.) Rich. | I | I | |||||||||
| Festuca ovina L. | I | I | |||||||||
| Rhaponticum coniferum (L.) Greuter | III | II | |||||||||
| Teucrium chamaedrys L. subsp. pinnatifidum (Sennen) Rech. fil. | III | II | |||||||||
| Salvia officinalis L. | I | II | |||||||||
| Linum narbonense L. | I | I | |||||||||
| Viola rupestris F.W. Schmidt | I | II | |||||||||
| Viscum album L. subsp. austriacum (Wiesb.) Vollm. | II | I | |||||||||
| Helianthemum violaceum (Cav.) Pers. | I | I | |||||||||
| Iberis ciliata All. | I | I | |||||||||
| Atractylis humilis L. | I | ||||||||||
| Dictamnus hispanicus Webb ex Willk. | I | ||||||||||
| Epipactis microphylla (Ehrh.) Sw. | I | ||||||||||
| Erucastrum nasturtiifolium (Poir.) O.E. Schulz | I | ||||||||||
| Festuca rubra L. | I | ||||||||||
| Galium lucidum All. subsp. corrudifolium (Vill.) Hayek | I | ||||||||||
| Galium pumilum Murray | I | ||||||||||
| Sanguisorba verrucosa (Link ex G. Don) Ces. | I | ||||||||||
| Thalictrum tuberosum L. | I | ||||||||||
| Veronica tenuifolia Asso | I | ||||||||||
| Lavandula latifolia Medik. | II | I | I | I | I | ||||||
| Eryngium campestre L. | I | I | I | ||||||||
| Digitalis obscura subsp. obscura L. | I | I | |||||||||
| Helianthemum syriacum (Jacq.) Dum. Cours. | I | I | I | ||||||||
| Polygala rupestris Pourr. | III | I | |||||||||
| Lithodora fruticosa (L.) Griseb. | I | I | |||||||||
| Carex humilis Leyss. | V | ||||||||||
| Cephalaria leucantha (L.) Roem. & Schult. | I | ||||||||||
| Euphorbia verrucosa Lam. subsp. mariolensis (Rouy) Vives | I | ||||||||||
| Hedysarum boveanum Bunge ex Basiner subsp. europaeum Guitt. & Kerguélen | I | ||||||||||
| Helianthemum hirtum (L.) Mill. | I | ||||||||||
| Ononis fruticosa L. | I | ||||||||||
| Orobanche latisquama (F.W. Schultz) Batt. in Batt. & Trab. | I | ||||||||||
| Silene italica (L.) Pers. | I | ||||||||||
| Teucrium polium L. subsp. aragonense (Loscos & J.Pardo) Nyman | I | ||||||||||
| Aster squamatus (Spreng.) Hieron. | I | ||||||||||
| Fumana procumbens (Dunal) Gren. & Godron | I | ||||||||||
| Globularia vulgaris L. | I | ||||||||||
| Inula montana L. | I | ||||||||||
| Brachypodium retusum subsp. retusum (Pers.) P. Beauv. | II | III | II | V | III | ||||||
| Dactylis glomerata subsp. hispanica (Roth) Nyman | I | I | I | I | |||||||
| Echinospartum boissieri (Spach) Rothm. | I | III | I | I | |||||||
| Bupleurum fruticescens subsp. spinosum (Gouan) O. Bolòs & Vigo | I | I | I | I | |||||||
| Thymus mastichina subsp. mastichina (L.) L. | I | I | I | I | |||||||
| Festuca scariosa (Lag.) Asch. & Graebn. | I | I | I | I | |||||||
| Thymus orospedanus Huguet del Villar | I | II | I | ||||||||
| Phlomis lychnitis L. | I | I | I | I | |||||||
| Genista mugronensis Vierh. | I | I | |||||||||
| Lavandula stoechas Lam. | I | I | |||||||||
| Thymus longiflorus Boiss. | I | I | |||||||||
| Phlomis purpurea subsp. purpurea L. | I | I | I | ||||||||
| Teucrium pseudochamaepitys L. | I | I | |||||||||
| Genista spartioides Spach | I | I | |||||||||
| Genista umbellata subsp. equisetiformis (Spach) Rivas Goday & Rivas Mart. | I | I | |||||||||
| Genista umbellata subsp. umbellata (L’Hér.) Dum. Cours. | I | II | |||||||||
| Phlomis purpurea subsp. almeriensis (Pau) Losa & Rivas Goday | I | I | |||||||||
| Spartium junceum L. | I | I | |||||||||
| Thymbra capitata (L.) Cav. | I | I | |||||||||
| Berberis hispanica Boiss. & Reut. | I | ||||||||||
| Brachypodium retusum subsp. boissieri (Nyman) Romero García | I | ||||||||||
| Brachypodium retusum subsp. boissieri (Nyman) Romero García | I | ||||||||||
| Festuca indigesta subsp. hackeliana | I | ||||||||||
| Ceterach officinarum Willd. | I | ||||||||||
| Fumana thymifolia (L.) Spach ex Webb | I | I | I | ||||||||
| Artemisia herba-alba Asso | I | I | I | ||||||||
| Helianthemum almeriense Pau | I | I | I | ||||||||
| Genista cinerea subsp. speciosa Losa & Rivas Goday (VilL.) DC. | I | I | I | ||||||||
| Salsola oppositifolia Desf. | I | I | I | ||||||||
| Lavandula lanata Boiss. | I | I | |||||||||
| Santolina villosa Mill. | I | I | |||||||||
| Satureja obovata subsp. canescens (Rouy) Rivas Martínez | I | I | |||||||||
| Artemisia barrelieri Besser | I | I | |||||||||
| Artemisia campestris subsp. glutinosa (J. Gay ex DC.) Batt. In Batt. & Trab. | I | I | |||||||||
| Teucrium capitatum subsp. gracillimum (Rouy) Valdés-Bermejo | I | I | |||||||||
| Ballota hirsuta Benth. | I | ||||||||||
| Capparis spinosa subsp. spinosa L. | I | ||||||||||
| Cistus monspeliensis L. | I | ||||||||||
| Fumana laevipes (L.) Spach | I | ||||||||||
| Launaea lanifera Pau | I | ||||||||||
| Lavatera oblongifolia Boiss. | I | ||||||||||
| Satureja cuneifolia Ten. | I | ||||||||||
| Scorzonera angustifolia L. | I | ||||||||||
| Sideritis lasiantha Juss. ex Pers. | I | ||||||||||
| Suaeda vera J. F. Gmelin in L. | I | ||||||||||
| Teucrium lusitanicum Schreb. | I | ||||||||||
| Thapsia villosa var. villosa L. | I | ||||||||||
| Thymelaea hirsuta (L.) Endl. | I | ||||||||||
| Thymus baeticus Boiss. ex Lacaita | I | ||||||||||
| Thymus hyemalis Lange | I | ||||||||||
| Ulex parviflorus subsp. willkommii (Webb) Borja & al. | I | ||||||||||
| Umbilicus rupestris (Salisb.) Dandy in Ridd. & al. | I | ||||||||||
| Santolina canescens Lag. | I | I | |||||||||
| Helianthemum croceum (Desf.) Pers. | II | ||||||||||
| Teucrium capitatum L. | II | ||||||||||
| Prunus spinosa L. | I | ||||||||||
| Ptilostemon hispanicus (Lam.) Greuter | I | ||||||||||
| Asperula hirsuta Desf. | I | ||||||||||
| Asphodelus albus Mill. | I | ||||||||||
| Centaurea boissieri DC. subsp. willkommii (Sch. Bip.) Dostál | I | ||||||||||
| Centaurea granatensis Boiss. ex DC. | I | ||||||||||
| Convolvulus boissieri Steudel | I | ||||||||||
| Fumana ericoides (Cav.) Gand. in Magnier subsp. paradoxa (Heywood) Charpin & Fern. Casas | I | ||||||||||
| Globularia spinosa L. | I | ||||||||||
| Helianthemum cinereum (Cav.) Pers. subsp. rotundifolium (Dunal) Greuter & Burdet | I | ||||||||||
| Helianthemum frigidulum Cuatrec. | I | ||||||||||
| Knautia spathulata Lag. | I | ||||||||||
| Lactuca tenerrima Pourr. | I | ||||||||||
| Leucanthemopsis spathulifolia (Gay) Fern. Casas | I | ||||||||||
| Paronychia suffruticosa (L.) DC. in Lam. | I | ||||||||||
| Phagnalon saxatile (L.) Cass. | I | ||||||||||
| Scorzonera albicans Coss. | I | ||||||||||
| Sedum dasyphyllum L. | I | ||||||||||
| Silene legionensis Lag. | I | ||||||||||
| Sideritis incana L. | I | I | |||||||||
| Astragalus clusianus Soldano | I | ||||||||||
| Capparis canescens Banks ex DC. | I | ||||||||||
| Anthyllis onobrychioides Cav. | I | ||||||||||
| Micromeria graeca subsp. longiflora (C. Presl) Nyman | I | ||||||||||
| Salsola genistoides Juss. ex Poir. in Lam. | I | ||||||||||
| Astragalus incanus subsp. incanus L. | I | ||||||||||
| Astragalus sesameus L. | I | ||||||||||
| Brachypodium phoenicoides (L.) Roem. & Schult. | I | ||||||||||
| Cytisus scoparius subsp. reverchonii (Degen & Hervier) Rivas Goday & Rivas Mart. | I | ||||||||||
| Erinacea anthyllis Link | I | ||||||||||
| Euphorbia nicaeensis All. | I | ||||||||||
| Fumana hispidula Losc. & J. Pardo in Willk. | I | ||||||||||
| Halimium umbellatum subsp. viscosum (Willk.) O. Bolòs & Vigo | I | ||||||||||
| Helianthemum squamatum (L.) Dum. Cours. | I | ||||||||||
| Helichrysum italicum subsp. serotinum (Boiss.) P. Fourn. | I | ||||||||||
| Linum suffruticosum subsp. suffruticosum L. | I | ||||||||||
| Matthiola fruticulosa (Loefl. ex L.) Maire in Jahand. & Maire | I | ||||||||||
| Ononis tridentata L. | I | ||||||||||
| Stipa lagascae Roem. & Schult. | I | ||||||||||
| Teucrium webbianum Boiss. | I | ||||||||||
| Thymus serpylloides subsp. gadorensis (Pau) Jalas | I | ||||||||||
| Thymus membranaceus subsp. membranaceus Boiss. | I | ||||||||||
| Trifolium spumosum L. | I | ||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cano, E.; Cano-Ortiz, A.; Fuentes, J.C.P.; Quinto-Canas, R.; Igbareyeh, J.; del Río, S.; Gomes, C.J.P. Ecological and Syntaxonomic Analysis of Pinus halepensis Mill. in the Iberian Peninsula and Balearic Islands. Land 2022, 11, 369. https://doi.org/10.3390/land11030369
Cano E, Cano-Ortiz A, Fuentes JCP, Quinto-Canas R, Igbareyeh J, del Río S, Gomes CJP. Ecological and Syntaxonomic Analysis of Pinus halepensis Mill. in the Iberian Peninsula and Balearic Islands. Land. 2022; 11(3):369. https://doi.org/10.3390/land11030369
Chicago/Turabian StyleCano, Eusebio, Ana Cano-Ortiz, José Carlos Piñar Fuentes, Ricardo Quinto-Canas, Jehad Igbareyeh, Sara del Río, and Carlos José Pinto Gomes. 2022. "Ecological and Syntaxonomic Analysis of Pinus halepensis Mill. in the Iberian Peninsula and Balearic Islands" Land 11, no. 3: 369. https://doi.org/10.3390/land11030369