
A First Expression, Purification and Characterization of Endo-β-1,3-Glucanase from Penicillium expansum
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Preparation of Culture Medium and Reagent
2.3. Construction and Transformation of Recombinant Plasmid pET-30a-PeBgl
2.4. Expression and Purification of PeBgl1 Protein
2.5. Determination of PeBgl1 Enzyme Activity
2.6. Study of the Enzymatic Characteristics of PeBgl1
2.6.1. Substrate Specificity of PeBgl1
2.6.2. Optimum Reaction Temperature and pH of PeBgl1
2.6.3. Effect of Na2HPO4-NaH2PO4 Buffer and NaCl Buffer Concentration in Substrate Solution on the Activity of Recombinant Enzyme
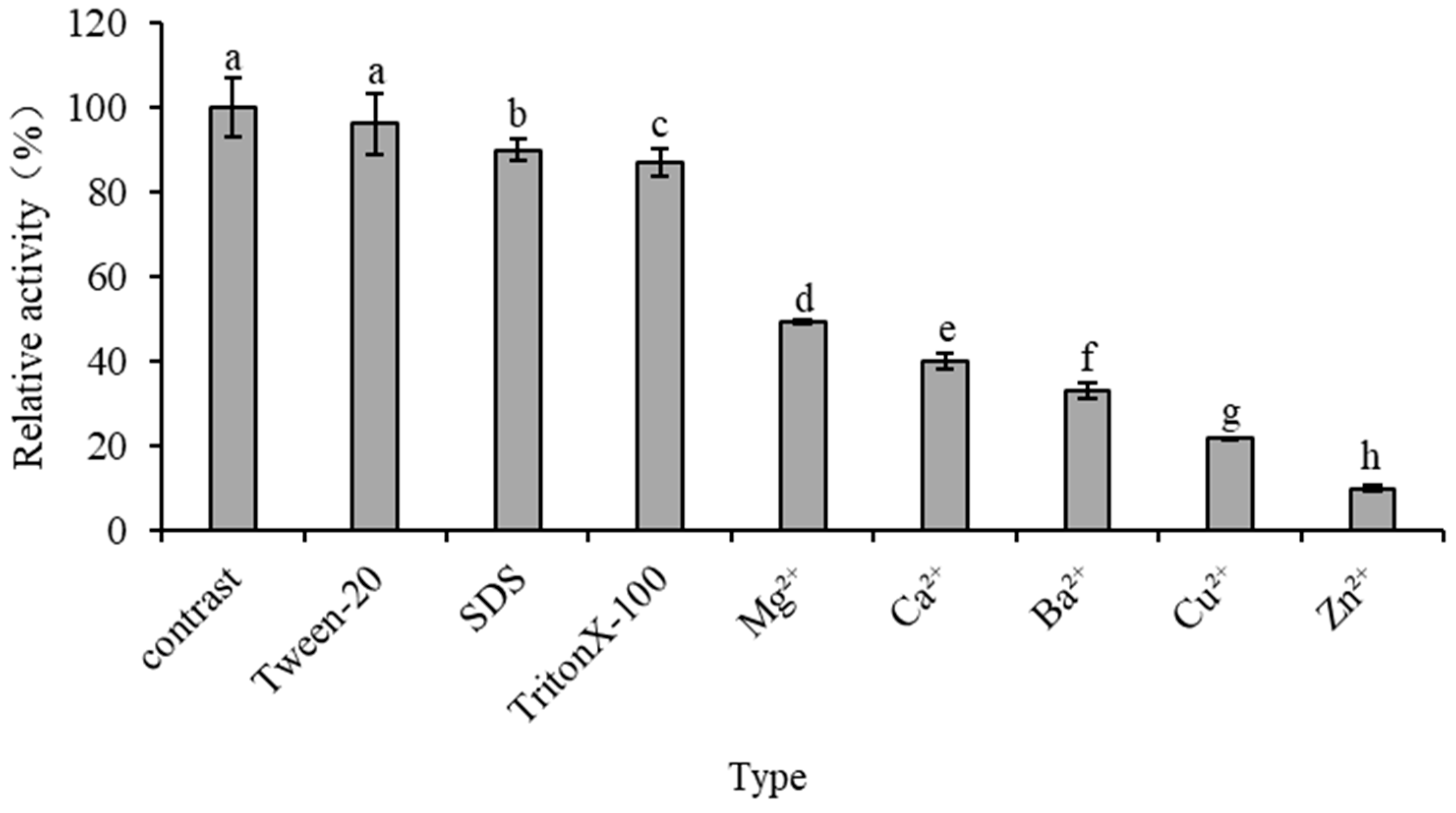
2.6.4. Effects of Metal Ions and Surfactants on Recombinant Enzyme Activity
3. Results and Discussion
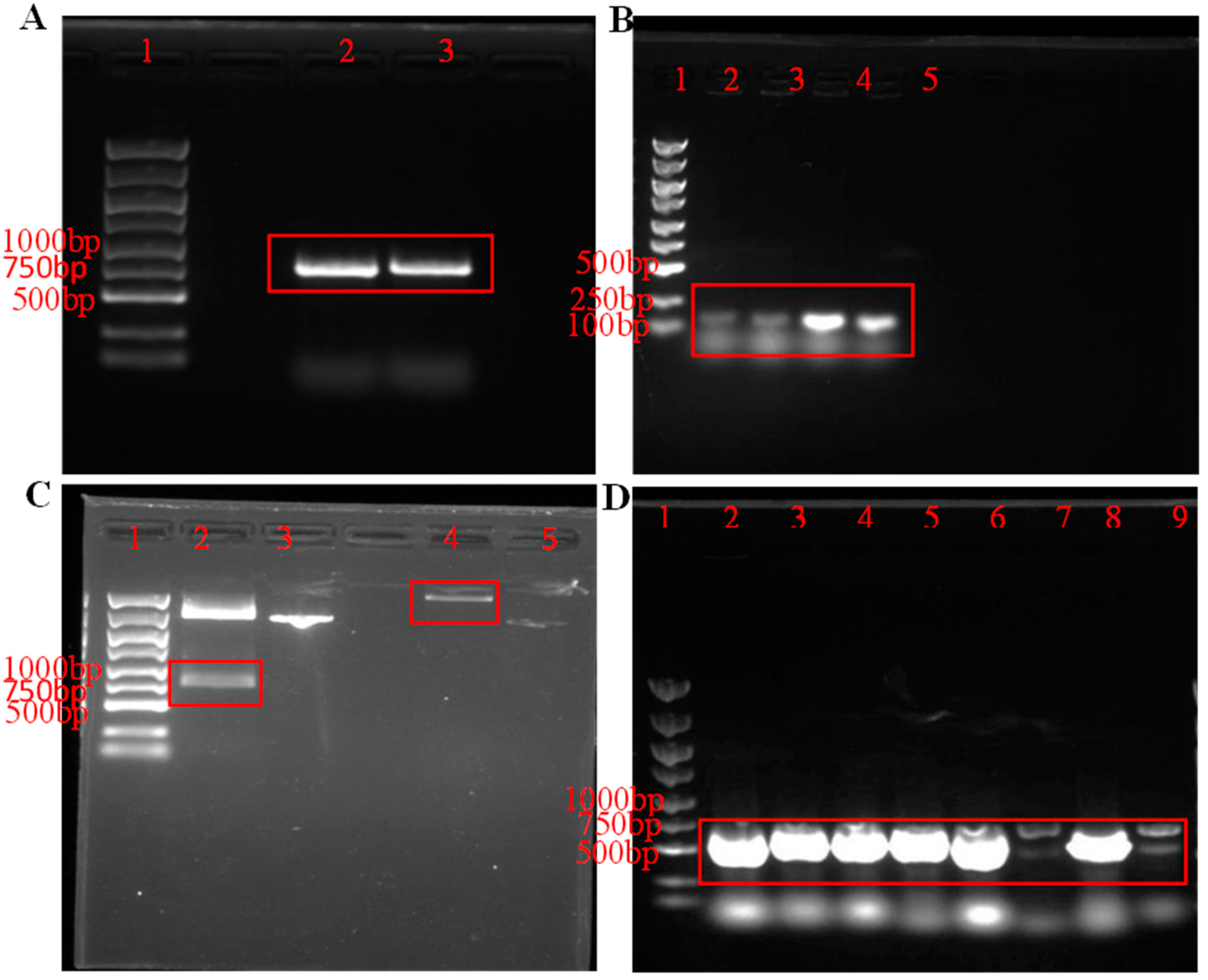
3.1. Construction of Recombinant Plasmid pET-30a-PeBgl1-Rosetta
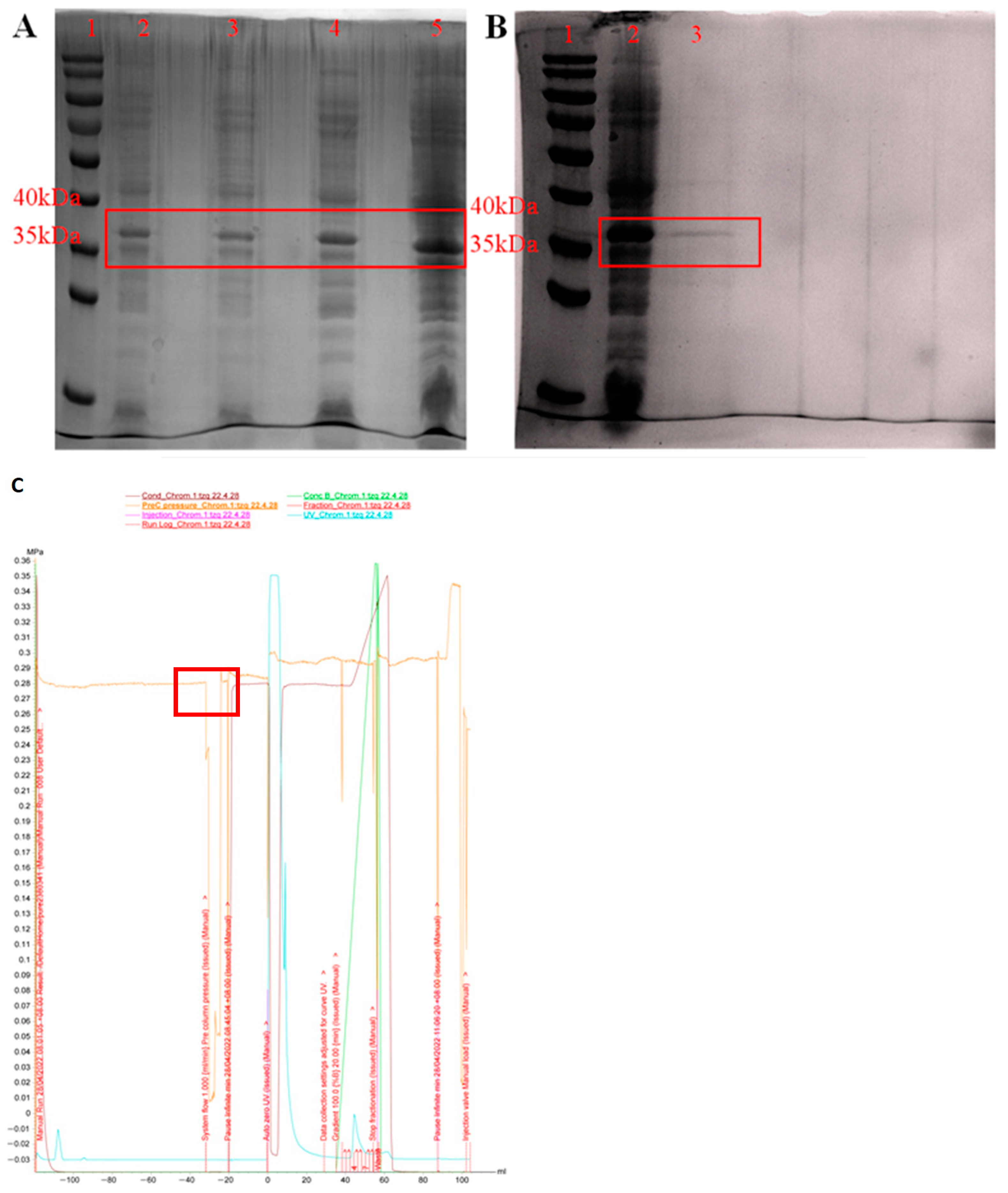
3.2. Expression and Purification of PeBgl1 Protein
3.3. Study of PeBgl1 Substrate Specificity
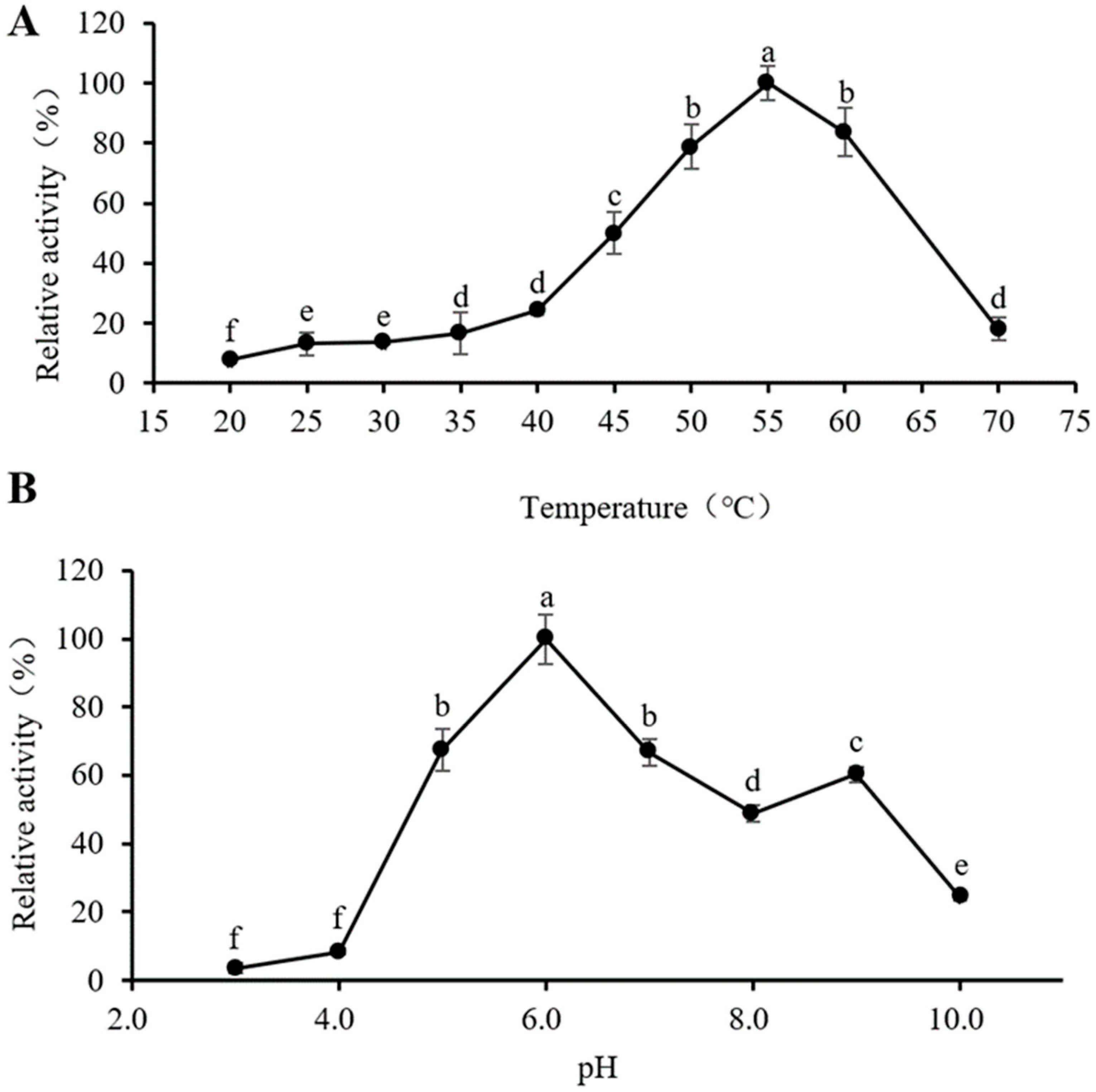
3.4. Optimum Reaction Temperature and pH of PeBgl1
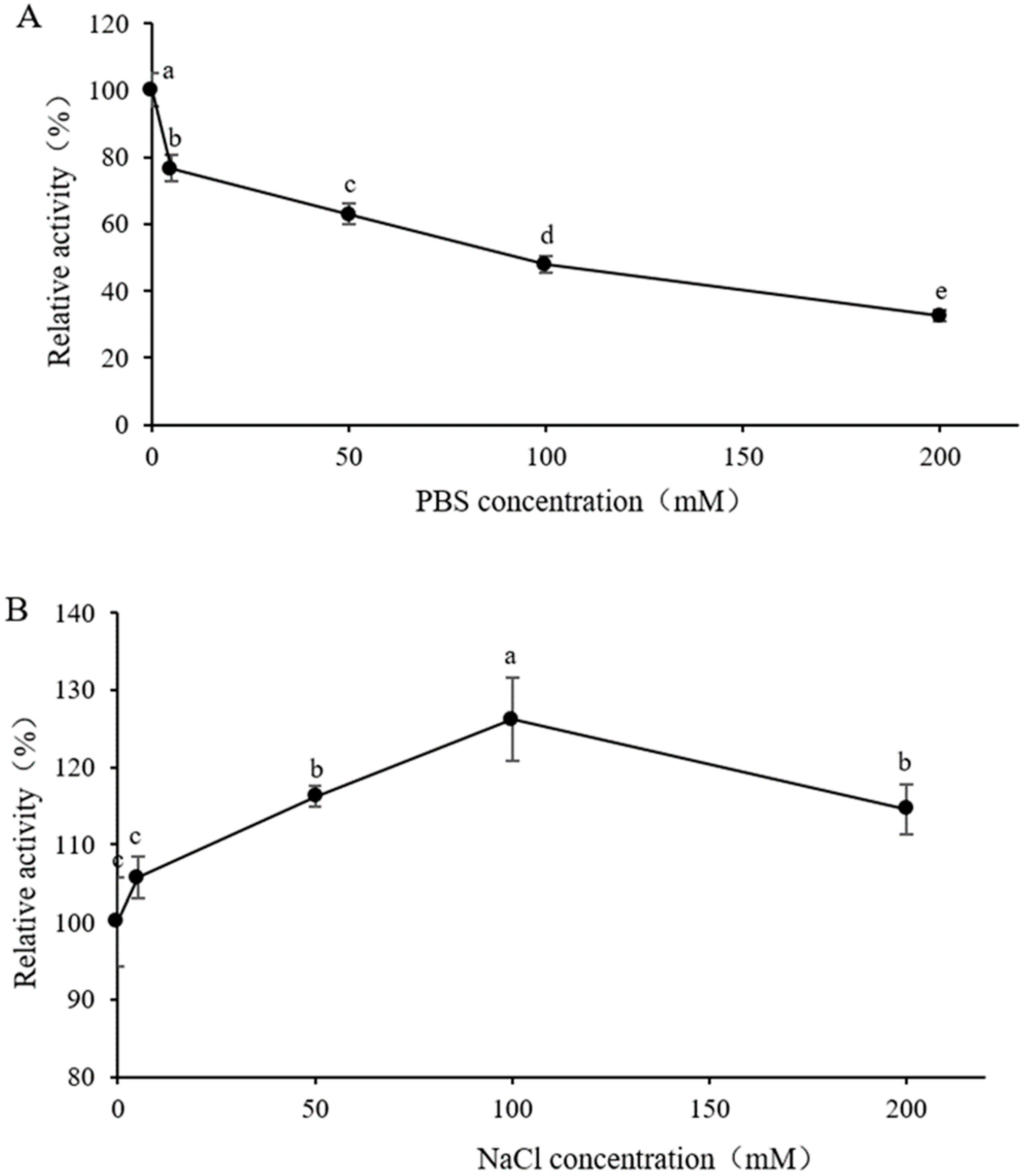
3.5. Effect of Buffer Concentration in Substrate Solution on Enzyme Activity
3.6. Effect of Metal Ions and Surfactants on Enzyme Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mészáros, Z.; Nekvasilová, P.; Bojarová, P.; Křen, V.; Slámová, K. Reprint of: Advanced glycosidases as ingenious biosynthetic instruments. Biotechnol. Adv. 2021, 51, 107820. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.-M.; Chen, J.; Shi, Y.-P. α-Glucosidase immobilization on chitosan-modified cellulose filter paper: Preparation, property and application. Int. J. Biol. Macromol. 2019, 122, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Mitsudome, T.; Xu, J.; Nagata, Y.; Masuda, A.; Iiyama, K.; Morokuma, D.; Li, Z.; Mon, H.; Lee, J.M.; Kusakabe, T. Expression, purification, and characterization of endo-β-N-acetylglucosaminidase H using baculovirus-mediated silkworm protein expression system. Appl. Biochem. Biotechnol. 2014, 172, 3978–3988. [Google Scholar] [CrossRef] [PubMed]
- Mhiri, S.; Bouanane-Darenfed, A.; Jemli, S.; Neifar, S.; Ameri, R.; Mezghani, M.; Bouacem, K.; Jaouadi, B.; Bejar, S. A thermophilic and thermostable xylanase from Caldicoprobacter algeriensis: Recombinant expression, characterization and application in paper biobleaching. Int. J. Biol. Macromol. 2020, 164, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Saldarriaga-Hernández, S.; Velasco-Ayala, C.; Flores, P.L.-I.; de Jesús Rostro-Alanis, M.; Parra-Saldivar, R.; Iqbal, H.M.N.; Carrillo-Nieves, D. Biotransformation of lignocellulosic biomass into industrially relevant products with the aid of fungi-derived lignocellulolytic enzymes. Int. J. Biol. Macromol. 2020, 161, 1099–1116. [Google Scholar] [CrossRef]
- Kalyani, D.C.; Reichenbach, T.; Aspeborg, H.; Divne, C. A homodimeric bacterial exo-β-1, 3-glucanase derived from moose rumen microbiome shows a structural framework similar to yeast exo-β-1, 3-glucanases. Enzym. Microb. Technol. 2021, 143, 109723. [Google Scholar] [CrossRef]
- Martin, K.; McDougall, B.M.; McIlroy, S.; Jayus; Chen, J.; Seviour, R.J. Biochemistry and molecular biology of exocellular fungal β-(1, 3)-and β-(1, 6)-glucanases. FEMS Microbiol. Rev. 2007, 31, 168–192. [Google Scholar] [CrossRef]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Extraction, structure and biofunctional activities of laminarin from brown algae. Int. J. Food Sci. Technol. 2015, 50, 24–31. [Google Scholar] [CrossRef]
- Taylor, P.R.; Tsoni, S.V.; Willment, J.A.; Dennehy, K.M.; Rosas, M.; Findon, H.; Haynes, K.; Steele, C.; Botto, M.; Gordon, S. Dectin-1 is required for β-glucan recognition and control of fungal infection. Nat. Immunol. 2007, 8, 31–38. [Google Scholar] [CrossRef]
- Chao, Q.; Ding, Y.; Chen, Z.-H.; Xiang, M.-H.; Wang, N.; Gao, X.-D. Recent Progress in Chemo-Enzymatic Methods for the Synthesis of N-Glycans. Front. Chem. 2020, 8, 513. [Google Scholar] [CrossRef]
- Usoltseva, R.V.; Belik, A.A.; Kusaykin, M.I.; Malyarenko, O.S.; Zvyagintseva, T.N.; Ermakova, S.P. Laminarans and 1,3-β-D-glucanases. Int. J. Biol. Macromol. 2020, 163, 1010–1025. [Google Scholar] [CrossRef]
- Balasubramanian, V.; Vashisht, D.; Cletus, J.; Sakthivel, N. Plant β-1, 3-glucanases: Their biological functions and transgenic expression against phytopathogenic fungi. Biotechnol. Lett. 2012, 34, 1983–1990. [Google Scholar] [CrossRef] [PubMed]
- Aimanianda, V.; Simenel, C.; Garnaud, C.; Clavaud, C.; Tada, R.; Barbin, L.; Mouyna, I.; Heddergott, C.; Popolo, L.; Ohya, Y. The dual activity responsible for the elongation and branching of β-(1, 3)-glucan in the fungal cell wall. MBio 2017, 8, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Kasahara, N.; Aida, K.; Tanaka, H. Three N-terminal domains of beta-1, 3-glucanase A1 are involved in binding to insoluble beta-1, 3-glucan. J. Bacteriol. 1992, 174, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kim, Y. Enhancement of β-1,3-Glucanase Activity by Sequential δ-Sequence Mediated Integration in Saccharomyces cerevisiae. J. Life Sci. 2014, 24, 1046–1054. [Google Scholar] [CrossRef]
- Kim, M.J.; Tamano, K.; Machida, M.; Kim, S.-K.; Kim, Y.; Nam, S.-W. Optimization for Production of Exo-β-1,3-glucanase (Laminarinase) from Aspergillus oryzae in Saccharomyces cerevisiae. Korean Soc. Biotechnol. Bioeng. J. 2011, 26, 427–432. [Google Scholar] [CrossRef]
- Gao, M.-J.; Yan, J.-J.; Zhao, Y.; Zhu, L.; Yang, G.-S.; Zhan, X.-B. Expression of a thermostable β-1,3-glucanase from Trichoderma harzianum in Pichia pastoris and use in oligoglucosides hydrolysis. Process Biochem. 2021, 107, 74–82. [Google Scholar] [CrossRef]
- Lu, Z.; Rämgård, C.; Ergenlioğlu, İ.; Sandin, L.; Hammar, H.; Andersson, H.; King, K.; Inman, A.R.; Hao, M.; Bulone, V.; et al. Multiple enzymatic approaches to hydrolysis of fungal β-glucans by the soil bacterium Chitinophaga Pinensis. FEBS J. 2023, 290, 2909–2922. [Google Scholar] [CrossRef]
- Abdelmoteleb, A.; Troncoso-Rojas, R.; Gonzalez-Soto, T.; Gonzalez-Mendoza, D. Antifungical Activity of Autochthonous Bacillus subtilis Isolated from Prosopis juliflora against Phytopathogenic Fungi. Mycobiology 2017, 45, 385–391. [Google Scholar] [CrossRef]
- Montoya Espinoza, W.J.; Nolasco Cárdenas, O.P.; Acuña Payano, R.K.; Gutiérrez, A.I.F. Expresión de β-1,3-Glucanasas de Beauveria bassiana en cultivo con extracto de los fitopatógenos Peronospora variabilis y Fusarium oxysporum. Sci. Agropecu. 2016, 7, 253–257. [Google Scholar] [CrossRef]
- Woo, C.B.; Kang, H.N.; Lee, S.B. Molecular cloning and anti-fungal effect of endo-beta-1,3-glucanase from Thermotoga maritima. Food Sci. Biotechnol. 2014, 23, 1243–1246. [Google Scholar] [CrossRef]
- Ma, J.; Qin, Z.; Zhou, P.; Wang, R.; Yan, Q.; Jiang, Z.; Yang, S. Structural insights into the substrate recognition and catalytic mechanism of a fungal glycoside hydrolase family 81 β-1, 3-glucanase. Enzym. Microb. Technol. 2022, 153, 109948. [Google Scholar] [CrossRef]
- Lee, Y.; Lee, J.-H.; Shim, W.-B.; Elvitigala, D.A.S.; Zoysa, M.D.; Lee, S.-J.; Heo, S.-J.; Lee, J.; Kang, D.-H.; Oh, C. Molecular cloning, over-expression and enzymatic characterization of an endo-acting β-1, 3-glucanase from marine bacterium Mesoflavibacter zeaxanthinifaciens S86 in Escherichia coli. Ocean Sci. J. 2014, 49, 425–432. [Google Scholar] [CrossRef]
- Almirall, M.; Francesch, M.; Perez-Vendrell, A.M.; Brufau, J.; Esteve-Garcia, E. The differences in intestinal viscosity produced by barley and β-glucanase alter digesta enzyme activities and ileal nutrient digestibilities more in broiler chicks than in cocks. J. Nutr. 1995, 125, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, P. Revisiting the Cellulosimicrobium cellulans yeast-lytic β-1, 3-glucanases toolbox: A review. Microb. Cell Factories 2006, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nagarajkumar, M.; Bhaskaran, R.; Velazhahan, R. Involvement of secondary metabolites and extracellular lytic enzymes produced by Pseudomonas fluorescens in inhibition of Rhizoctonia solani, the rice sheath blight pathogen. Microbiol. Res. 2004, 159, 73–81. [Google Scholar] [CrossRef]
- McGrath, C.E.; Wilson, D.B. Characterization of a Thermobifida fusca β-1, 3-glucanase (Lam81A) with a potential role in plant biomass degradation. Biochemistry 2006, 45, 14094–14100. [Google Scholar] [CrossRef]
- Scafati, V.; Troilo, F.; Ponziani, S.; Giovannoni, M.; Scortica, A.; Pontiggia, D.; Angelucci, F.; Di Matteo, A.; Mattei, B.; Benedetti, M. Characterization of two 1,3-β-glucan-modifying enzymes from Penicillium sumatraense reveals new insights into 1,3-β-glucan metabolism of fungal saprotrophs. Biotechnol. Biofuels Bioprod. 2022, 15, 138. [Google Scholar] [CrossRef]
- Wang, K.; Zheng, X.; Su, Y.; Lu, Y.; Yang, Q.; Shi, Y.; Lanhuang, B.; Zhang, X.; Zhao, L.; Godana, E.; et al. A glycoside hydrolase superfamily gene plays a major role in Penicillium expansum growth and pathogenicity in apples. Postharvest Biol. Technol. 2023, 198, 112228. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Fernández, F.J.; Vega, M.C. Choose a suitable expression host: A survey of available protein production platforms. Adv. Technol. Protein Complex Prod. Charact. 2016, 896, 15–24. [Google Scholar] [CrossRef]
- Meng, L.; Liu, Y.; Yin, X.; Zhou, H.; Wu, J.; Wu, M.; Yang, L. Effects of His-tag on catalytic activity and enantioselectivity of recombinant transaminases. Appl. Biochem. Biotechnol. 2020, 190, 880–895. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, Y.; Yuan, Q. Study the effect of His-tag on chondroitinase ABC I based on characterization of enzyme. Int. J. Biol. Macromol. 2015, 78, 96–101. [Google Scholar] [CrossRef]
- Woestenenk, E.A.; Hammarström, M.; van den Berg, S.; Härd, T.; Berglund, H. His tag effect on solubility of human proteins produced in Escherichia coli: A comparison between four expression vectors. J. Struct. Funct. Genom. 2004, 5, 217–229. [Google Scholar] [CrossRef]
- Klose, J.; Wendt, N.; Kubald, S.; Krause, E.; Fechner, K.; Beyermann, M.; Bienert, M.; Rudolph, R.; Rothemund, S. Hexa-histidin tag position influences disulfide structure but not binding behavior of in vitro folded N-terminal domain of rat corticotropin-releasing factor receptor type 2a. Protein Sci. 2004, 13, 2470–2475. [Google Scholar] [CrossRef] [PubMed]
- Majorek, K.A.; Kuhn, M.L.; Chruszcz, M.; Anderson, W.F.; Minor, W. Double trouble—Buffer selection and H is-tag presence may be responsible for nonreproducibility of biomedical experiments. Protein Sci. 2014, 23, 1359–1368. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Huang, Z. Effect of his-tag on expression, purification, and structure of zinc finger protein, ZNF191 (243-368). Bioinorg. Chem. Appl. 2016, 2016, 8206854. [Google Scholar] [CrossRef]
- Jin, J.; Hjerrild, K.A.; Silk, S.E.; Brown, R.E.; Labbé, G.M.; Marshall, J.M.; Wright, K.E.; Bezemer, S.; Clemmensen, S.B.; Biswas, S. Accelerating the clinical development of protein-based vaccines for malaria by efficient purification using a four amino acid C-terminal ‘C-tag’. Int. J. Parasitol. 2017, 47, 435–446. [Google Scholar] [CrossRef]
- Khairil Anuar, I.N.A.; Banerjee, A.; Keeble, A.H.; Carella, A.; Nikov, G.I.; Howarth, M. Spy&Go purification of SpyTag-proteins using pseudo-SpyCatcher to access an oligomerization toolbox. Nat. Commun. 2019, 10, 1734. [Google Scholar] [CrossRef]
- Liu, G.; Lin, R.; Yan, H.; Hu, Q.; Liu, L.; Weng, Y. PCR Amplification and Sequence Analysis of Apicoplast rpoB and TufA Genes of Two Eimeria spp.in Rabbits. J. Heilongjiang August First Land Reclam. Univ. 2014, 26, 40–43. [Google Scholar] [CrossRef]
- Mierendorf, R.; Yeager, K.; Novy, R. The pET system: Your choice for expression. Innovations 1994, 1, 1–3. [Google Scholar]
- Lü, Y.F.; Yang, J.L.; Qin, Y.; Wang, Y.L. Prokaryotic expression and polyclonal antibody preparation of human polyamine modulated factor-1. Biotechnol. Bull. 2016, 26, 521–525. [Google Scholar] [CrossRef]
- Bhatwa, A.; Wang, W.; Hassan, Y.I.; Abraham, N.; Li, X.-Z.; Zhou, T. Challenges associated with the formation of recombinant protein inclusion bodies in Escherichia coli and strategies to address them for industrial applications. Front. Bioeng. Biotechnol. 2021, 9, 630551. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhang, X.-M.; Yin, Y.-R.; Li, W.-J. Cloning, expression and characterization of a novel GH5 exo/endoglucanase of Thermobifida halotolerans YIM 90462T by genome mining. J. Biosci. Bioeng. 2015, 120, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Shibuya, H.; Jones, J.T. Molecular and biochemical characterization of an endo-β-1, 3-glucanase from the pinewood nematode Bursaphelenchus xylophilus acquired by horizontal gene transfer from bacteria. Biochem. J. 2005, 389, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Shu, C.-H.; Xu, C.-J.; Lin, E.-S. Production, purification and partial characterization of a novel endo-β-1, 3-glucanase from Agaricus brasiliensis. Process Biochem. 2006, 41, 1229–1233. [Google Scholar] [CrossRef]
- Ferrareze, P.A.G.; Correa, A.P.F.; Brandelli, A. Purification and characterization of a keratinolytic protease produced by probiotic Bacillus subtilis. Biocatal. Agric. Biotechnol. 2016, 7, 102–109. [Google Scholar] [CrossRef]
- Wei, Y.D.; Li, Y.; Deng, C.; Wu, S.H.; Huang, C.J.; Yi, Y. Expression and biochemical characterization of a multifunctional glycosidase from the thermophilic Bacillus licheniformis SR01. J. Gen. Appl. Microbiol. 2017, 63, 259–265. [Google Scholar] [CrossRef]
- Yi, P.; Yan, Q.; Jiang, Z.; Wang, L. A first glycoside hydrolase family 50 endo-β-1, 3-d-glucanase from Pseudomonas aeruginosa. Enzym. Microb. Technol. 2018, 108, 34–41. [Google Scholar] [CrossRef]
- Fuchs, K.-P.; Zverlov, V.V.; Velikodvorskaya, G.A.; Lottspeich, F.; Schwarz, W.H. Lic16A of Clostridium thermocellum, a non-cellulosomal, highly complex endo-β-1, 3-glucanase bound to the outer cell surface. Microbiology 2003, 149, 1021–1031. [Google Scholar] [CrossRef]
- Li, J.; Xu, X.; Shi, P.; Liu, B.; Zhang, Y.; Zhang, W. Overexpression and characterization of a novel endo-β-1, 3 (4)-glucanase from thermophilic fungus Humicola insolens Y1. Protein Expr. Purif. 2017, 138, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Brandelli, A.; Sala, L.; Kalil, S.J. Microbial enzymes for bioconversion of poultry waste into added-value products. Food Res. Int. 2015, 73, 3–12. [Google Scholar] [CrossRef]
- Krishnan, B.; Srivastava, S.S.; Sankeshi, V.; Garg, R.; Srivastava, S.; Sankaranarayanan, R.; Sharma, Y. βγ-Crystallination endows a novel bacterial glycoside hydrolase 64 with Ca2+-dependent activity modulation. J. Bacteriol. 2019, 201, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Schlenzig, D.; Wermann, M.; Ramsbeck, D.; Moenke-Wedler, T.; Schilling, S. Expression, purification and initial characterization of human meprin β from Pichia pastoris. Protein Expr. Purif. 2015, 116, 75–81. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrates | Major Glycoside Linkage (s) | Relative Activity (%) |
|---|---|---|
| Laminarin | 1,3-β | 32.11 c |
| Laminaribiose | 1,3-β | 100 a |
| Laminarihexaose | 1,3-β | 64.74 b |
| CMC-Na | 1,4-β | 0.52 d |
| Gentiobiose | 1,6-β | 0 |
| Maltose | 1,4-α | 0 |
| Salicin | D-glucose | 0.66 d |
| Xylo-oligosaccharide | 1,4-β | 0 |
| Chitin | 1,4-β | 1.83 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, K.; Huai, S.; Tan, Z.; Ngea, G.L.N.; Godana, E.A.; Shi, J.; Yang, Q.; Zhang, X.; Zhao, L.; Zhang, H. A First Expression, Purification and Characterization of Endo-β-1,3-Glucanase from Penicillium expansum. J. Fungi 2023, 9, 961. https://doi.org/10.3390/jof9100961
Wang K, Huai S, Tan Z, Ngea GLN, Godana EA, Shi J, Yang Q, Zhang X, Zhao L, Zhang H. A First Expression, Purification and Characterization of Endo-β-1,3-Glucanase from Penicillium expansum. Journal of Fungi. 2023; 9(10):961. https://doi.org/10.3390/jof9100961
Chicago/Turabian StyleWang, Kaili, Siyu Huai, Zhuqing Tan, Guillaume Legrand Ngolong Ngea, Esa Abiso Godana, Jun Shi, Qiya Yang, Xiaoyun Zhang, Lina Zhao, and Hongyin Zhang. 2023. "A First Expression, Purification and Characterization of Endo-β-1,3-Glucanase from Penicillium expansum" Journal of Fungi 9, no. 10: 961. https://doi.org/10.3390/jof9100961