
Two Novel Species of Talaromyces Discovered in a Karst Cave in the Satun UNESCO Global Geopark of Southern Thailand

 , , ,
, , , 

Abstract
:1. Introduction
2. Materials and Methods
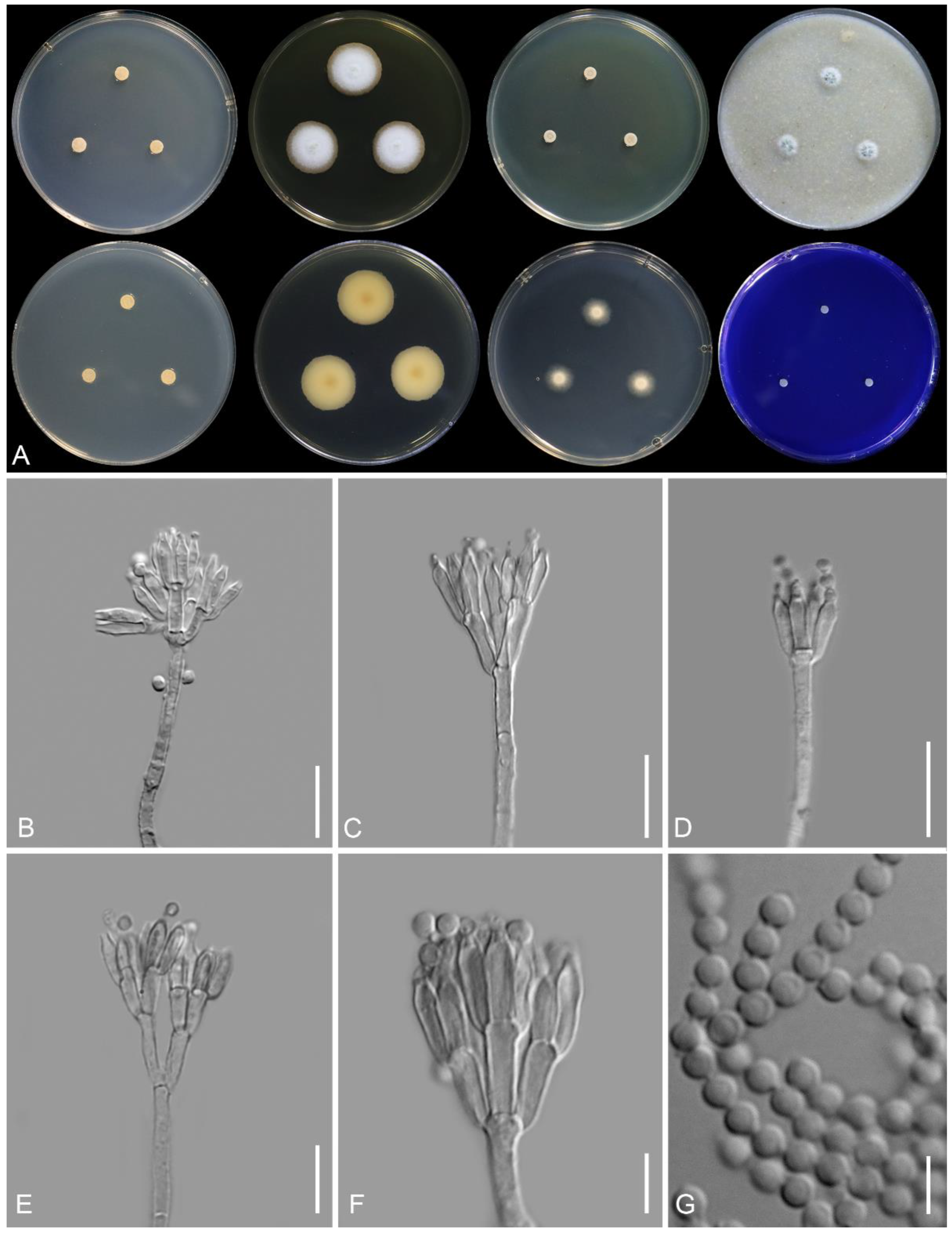
2.1. Collection, Isolation, and Morphology
2.2. DNA Extraction, PCR Amplification, and Phylogenetic Analyses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecular Locus | Primer Name | Direction | Reference | Amplification Profile | ||
|---|---|---|---|---|---|---|
| Denature | Repeat Step | Extension | ||||
| Internal transcribed spacers (ITS) | ITS1 | Forward | [17] | 94 °C (5 min) | 35 cycles, 94 °C (45 s), 55 °C (45 s), 72 °C (60 s) | 72 °C (7 min) |
| ITS5 | ||||||
| ITS4 | Reverse | |||||
| β-tubulin (BenA) | Bt2a | Forward | [18] | 94 °C (10 min) | 35 cycles, 94 °C (30 s), 57 °C (30 s), 72 °C (30 s) | 72 °C (10 min) |
| Bt2b | Reverse | |||||
| Calmodulin (CaM) | cmd5 | Forward | [19] | 94 °C (3 min) | 30 cycles, 94 °C (1 min), 57 °C (1 min), 72 °C (1 min) | 72 °C (10 min) |
| cmd6 | Reverse | |||||
| RNA polymerase II (RPB2) | 5F2 | Forward | [20] | 94 °C (3 min) | 34 cycles, 94 °C (1 min), 54 °C (1 min), 72 °C (1.30 min) | 72 °C (8 min) |
| 7cR | Reverse | |||||
| Taxon | Original Strain Number | GenBank Accession Number | |||
|---|---|---|---|---|---|
| ITS | BenA | CaM | RPB2 | ||
| Talaromyces aerius | CBS 140611 T | KU866647 | KU866835 | KU866731 | KU866991 |
| T. affinitatimellis | CBS 143840 T | LT906543 | LT906552 | LT906549 | LT906546 |
| T. africanus | CBS 147340 T = DTO 179-C5 | OK339610 | OK338782 | OK338808 | OK338833 |
| T. albisclerotius | CBS 141839 T = DTO 340-G5 | MN864276 | MN863345 | MN863322 | MN863334 |
| T. albobiverticillius | CBS 133440 T | HQ605705 | KF114778 | KJ885258 | KM023310 |
| T. amyrossmaniae | NFCCI 1919 T | MH909062 | MH909064 | MH909068 | MH909066 |
| T. assiutensis | CBS 147.78 T | JN899323 | KJ865720 | KJ885260 | KM023305 |
| T. atroroseus | CBS 133442 T | KF114747 | KF114789 | KJ775418 | KM023288 |
| T. austrocalifornicus | CBS 644.95 T | JN899357 | KJ865732 | KJ885261 | MN969147 |
| T. basipetosporus | CBS 143836 T = FMR 9720 | LT906542 | LT906563 | - | LT906545 |
| T. brasiliensis | URM 7618 T | MF278323 | LT855560 | LT855563 | MN969198 |
| T. calidominioluteus | CBS 147313 T = DTO 052-G3 | OK339612 | OK338786 | OK338817 | OK338837 |
| T. catalonicus | CBS 143039 T = FMR 16441 | LT899793 | LT898318 | LT899775 | LT899811 |
| T. chongqingensis | CS26-67 T | MZ358001 | MZ361343 | MZ361350 | MZ361357 |
| T. clemensii | PPRI 26753 T | MK951940 | MK951833 | MK951906 | MN418451 |
| T. convolutus | CBS 100537 T | JN899330 | KF114773 | MN969316 | JN121414 |
| T. diversus | CBS 320.48 T | KJ865740 | KJ865723 | KJ885268 | KM023285 |
| T. erythromellis | CBS 644.80 T | JN899383 | HQ156945 | KJ885270 | KM023290 |
| T. gaditanus | CBS 169.81 T = DTO 228-B8 | MH861318 | OK338775 | OK338802 | OK338827 |
| T. germanicus | CBS 147314 T = DTO 055-D1 | OK339619 | OK338799 | OK338812 | OK338845 |
| T. guatemalensis | CCF 6215 T | MN322789 | MN329687 | MN329688 | MN329689 |
| T. halophytorum | KACC 48127 T | MH725786 | MH729367 | MK111426 | MK111427 |
| T. heiheensis | HMAS 248789 T = CGMCC 3.18012 | KX447526 | KX447525 | KX447532 | KX447529 |
| T. minioluteus | CBS 642.68 T | JN899346 | MN969409 | KJ885273 | JF417443 |
| T. minnesotensis | CBS 142381 T | LT558966 | LT559083 | LT795604 | LT795605 |
| T. pernambucoensis | URM 6894 T | LR535947 | LR535945 | LR535946 | LR535948 |
| T. phuphaphetensis | TBRC 16281 T | ON692803 | ON706960 | ON706962 | ON706964 |
| T. resinae | CBS 324.83 T = IMI 080450 | MT079858 | MN969442 | MT066184 | MN969221 |
| T. rubrifaciens | CGMCC 3.17658 T | KR855658 | KR855648 | KR855653 | KR855663 |
| T. samsonii | CBS 137.84 T = DTO 304-C3 = DTO 169-G6 | MH861709 | OK338798 | OK338824 | OK338844 |
| T. satunensis | TBRC 16246 T | ON692804 | ON706961 | ON706963 | - |
| T. solicola | DAOM 241015 T | FJ160264 | GU385731 | KJ885279 | KM023295 |
| T. speluncarum | CBS 143844 T = FMR 16671 | LT985890 | LT985901 | LT985906 | LT985911 |
| T. subericola | CBS 144322 T = FMR 15656 | LT985888 | LT985899 | LT985904 | LT985909 |
| T. systylus | BAFCcult 3419 T | KP026917 | KR233838 | KR233837 | - |
| T. trachyspermus | CBS 373.48 T = IMI 040043 | JN899354 | KF114803 | KJ885281 | JF417432 |
| T. ucrainicus | CBS 162.67 T = FRR 3462 | JN899394 | KF114771 | KJ885282 | KM023289 |
| T. udagawae | CBS 579.72 T = IMI 197482 | JN899350 | KF114796 | KX961260 | MN969148 |
| T. flavus | CBS 310.38 T | JN899360 | JX494302 | KF741949 | JF417426 |
3. Results
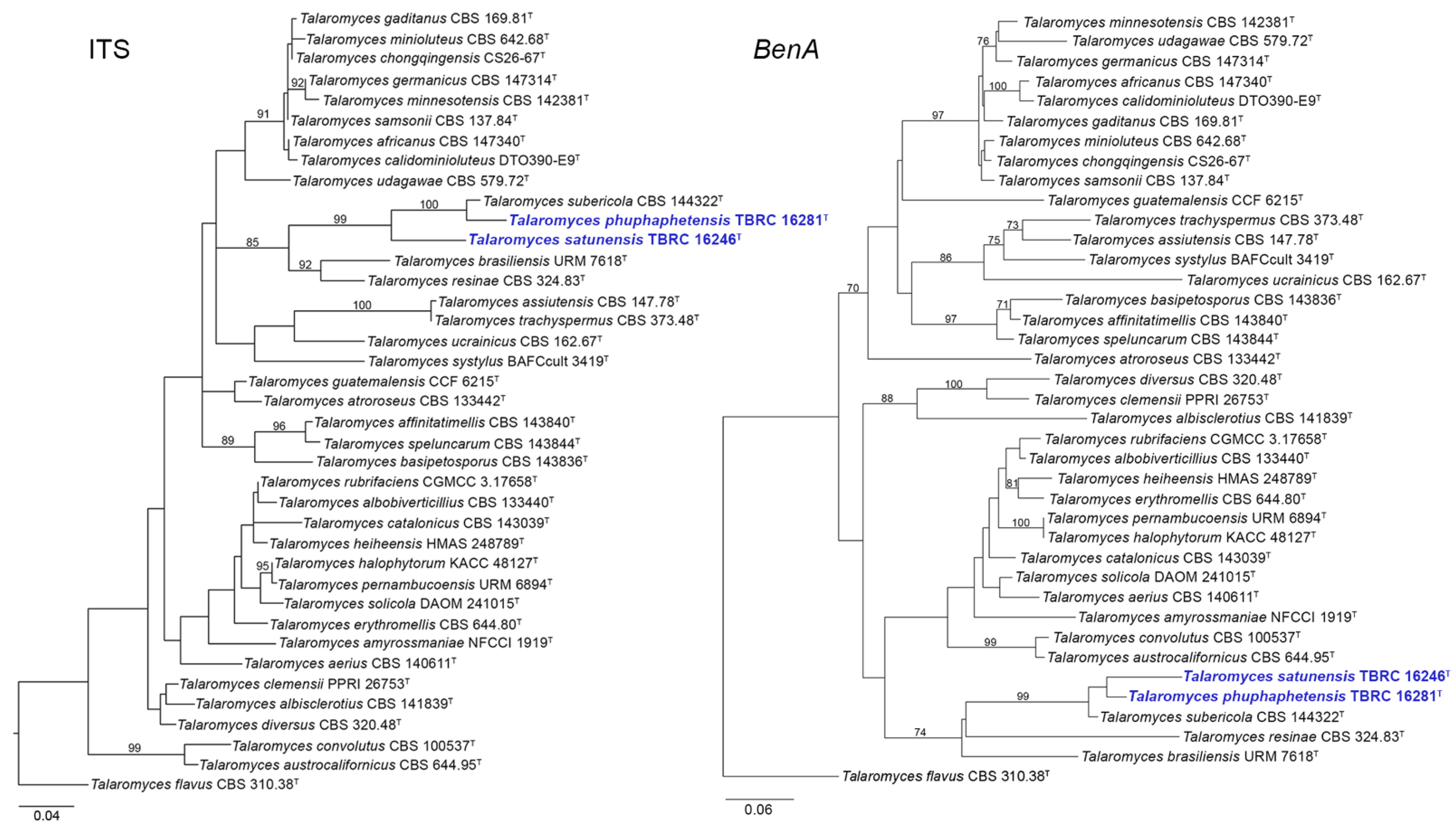
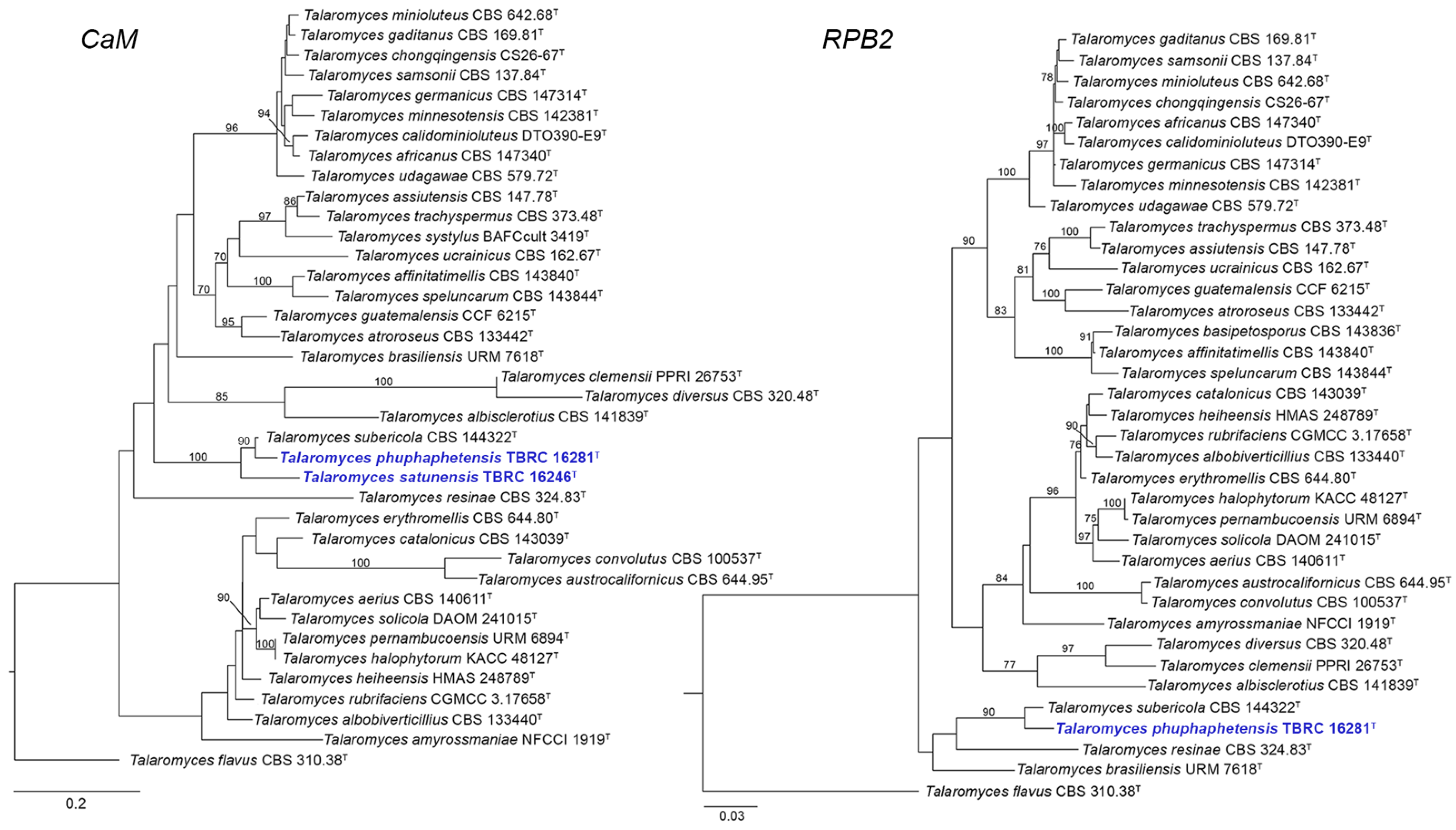
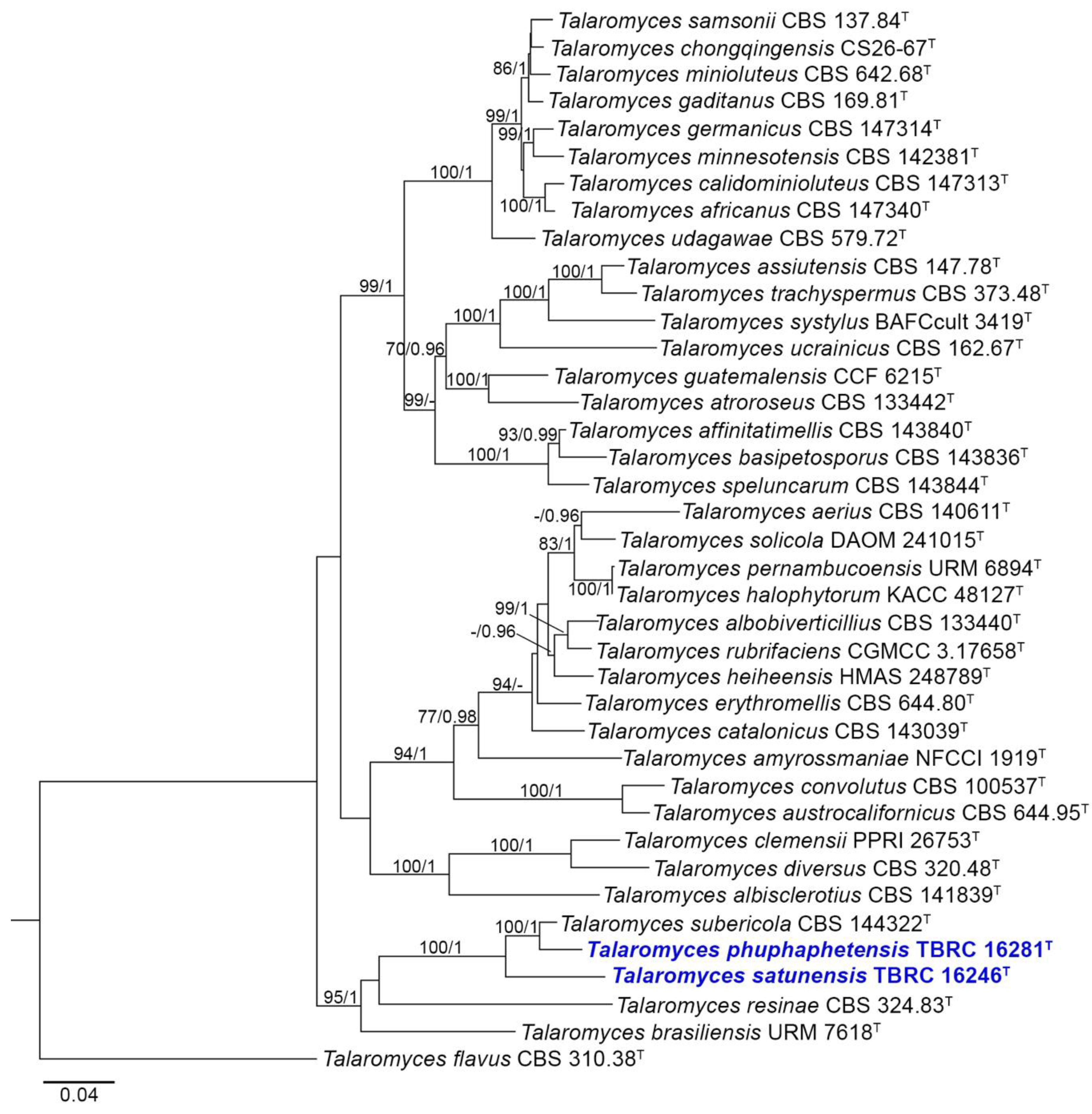
3.1. Phylogenetic Analysis
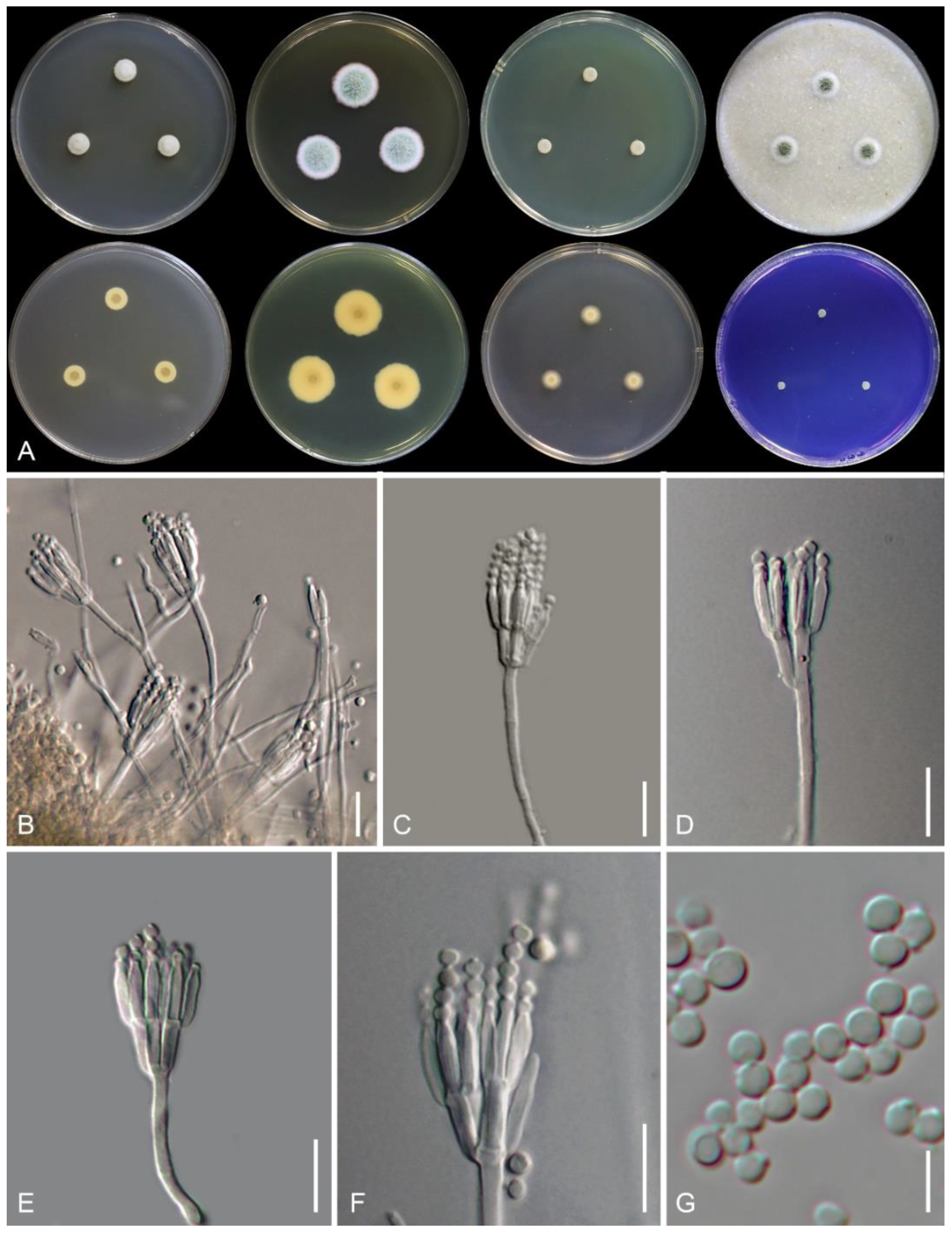
3.2. Taxonomy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benjamin, C.R. Ascocarps of Aspergillus and Penicillium. Mycologia 1955, 47, 669–687. [Google Scholar] [CrossRef]
- Guevara-Suarez, M.; Sutton, D.; Gené, J.; García, D.; Wiederhold, N.; Guarro, J.; Cano-Lira, J.F. Four new species of Talaromyces from clinical sources. Mycoses 2017, 60, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.C.; Chen, K.; Qin, W.T.; Zhuang, W.Y. Talaromyces heiheensis and T. mangshanicus, two new species from China. Mycol. Prog. 2017, 16, 73–81. [Google Scholar] [CrossRef]
- Barbosa, R.N.; Bezerra, J.D.P.; Souza-Motta, C.M.; Frisvad, J.C.; Samson, R.A.; Oliveira, N.T.; Houbraken, J. New Penicillium and Talaromyces species from honey, pollen and nests of stingless bees. Antonie Van Leeuwenhoek 2018, 111, 1883–1912. [Google Scholar] [CrossRef] [PubMed]
- Rajeshkumar, K.C.; Yilmaz, N.; Marathe, S.D.; Seifert, K.A. Morphology and multigene phylogeny of Talaromyces amyrossmaniae, a new synnematous species belonging to the section Trachyspermi from India. MycoKeys 2019, 45, 41–56. [Google Scholar] [CrossRef]
- You, Y.H.; Aktaruzzaman, M.; Heo, I.; Park, J.M.; Hong, J.W.; Hong, S.B. Talaromyces halophytorum sp. nov. isolated from roots of Limonium tetragonum in Korea. Mycobiology 2020, 48, 133–138. [Google Scholar] [CrossRef]
- Rodríguez-Andrade, E.; Stchigel, A.M.; Guarro, J.; Cano-Lira, J.F. Fungal diversity of deteriorated sparkling wine and cork stoppers in Catalonia, Spain. Microorganisms 2020, 8, 12. [Google Scholar] [CrossRef]
- Yordkayhun, S.; Wattanasen, K.; Thungprue, N. Geophysical investigation of the karst geosites in Satun UNESCO Global Geopark, Thailand: Implication for sinkhole hazard assessment. Geosci. J. 2022, 26, 249–266. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.Z.; Liu, S.J.; Cai, L. Culturable mycobiota from Karst caves in China, with descriptions of 20 new species. Persoonia 2017, 39, 1–31. [Google Scholar] [CrossRef]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.B.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 2nd ed.; Methuen: London, UK, 1967; pp. 1–243. [Google Scholar]
- Sri-indrasutdhi, V.; Boonyuen, N.; Suetrong, S.; Chuaseeharonnachai, C.; Sivichai, S.; Gareth, J.E.B. Wood-inhabiting freshwater fungi from Thailand: Ascothailandia grenadoidia gen. et sp. nov., Canalisporium grenadoidia sp. nov. with a key to Canalisporium species (Sordariomycetes, Ascomycota). Mycoscience 2010, 51, 411–420. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, A.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable, and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phyloge-netic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenet-ics. In PCR Protocols: A Guide to Methods and Applications; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Hong, S.B.; Cho, H.S.; Shin, H.D.; Frisvad, J.C.; Samson, R.A. Novel Neosartorya species isolated from soil in Korea. Int. J. Syst. Evol. Microbiol. 2006, 56, 477–486. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Qi, Z.T.; Kong, H.Z. A new species of Penicillium. Acta Mycologica Sin. 1982, 1, 103–105. [Google Scholar]
- Zhang, Z.K.; Wang, X.C.; Zhuang, W.Y.; Cheng, X.H.; Zhao, P. New species of Talaromyces (Fungi) isolated from soil in southwestern China. Biology 2021, 10, 745. [Google Scholar] [CrossRef]
- Stošić, S.; Ristić, D.; Gašić, K.; Starović, M.; Grbić, M.L.; Vukojević, J.; Živković, S. Talaromyces minioluteus: New postharvest fungal pathogen in Serbia. Plant Dis. 2020, 104, 656–667. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.R.; Xu, M.Y.; Kong, W.L.; Wu, F.; Zhang, Y.; Xie, X.L.; Li, D.W.; Wu, X.Q. Fine identification and classification of a novel beneficial Talaromyces fungal species from masson pine phizosphere soil. J. Fungi 2022, 8, 155. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, N.; Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of the genus Talaromyces. Stud. Mycol. 2014, 78, 175–341. [Google Scholar] [CrossRef] [PubMed]
- Yaguchi, T.; Someya, A.; Udagawa, S.-I. A reappraisal of intrageneric classification of Talaromyces based on the ubiquinone systems. Mycoscience 1996, 37, 55–60. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Yilmaz, N.; Thrane, U.; Rasmussen, K.B.; Houbraken, J.; Samson, R.A. Talaromyces atroroseus, a new species efficiently producing industrially relevant red pigments. PLoS ONE 2013, 8, e84102. [Google Scholar] [CrossRef]
- Lagashetti, A.; Dufossé, L.; Singh, S.K.; Singh, P. Fungal pigments and their prospects in different industries. Microorganisms 2019, 7, 604. [Google Scholar] [CrossRef]
- Morales-Oyervides, L.; Oliveira, J.; Sousa-Gallagher, M.; Méndez-Zavala, A.; Montañez, J.C. Assessment of the dyeing properties of the pigments produced by Talaromyces spp. J. Fungi 2017, 3, 38. [Google Scholar] [CrossRef]





| Microscopic Characters | T. brasiliensis [4] | T. phuphaphetensis (This Study) | T. resinae [21] | T. satunensis (This Study) | T. subericola [7] | ||
|---|---|---|---|---|---|---|---|
| On MEA | Conidiophore | stipes (μm) | 20–50 × 2.5–4 | 15–60 × 2.5–3 | N/A | 20–290 × 2–3.2 | N/A |
| branching | biverticillate | mostly biverticillate, monoverticillate | mostly biverticillate, monoverticillate, terverticillate | ||||
| ornamentation | smooth | finely tuberculate | tuberculate | ||||
| Metulae | size (μm) | 8–11 × 2.5–3.5 | 5–9 × 1.5–3 | 5.5–10 × 2–3.3 | |||
| per verticil | 5–6 | 2–6 | 2–5 | ||||
| Phialides | size (μm) | 7–11 (−14) × 2–3 | 7–9.5 × 2–3 | 6–9 × 2–3.2 | |||
| per metula | 3–4 | 3–5 | 2–5 | ||||
| Conidia | size (μm) | 2–3 | 2.0–3.5 | 2.5–3 | |||
| shape | globose | Globose to sub-globose | globose to sub-globose | ||||
| ornamentation | finely roughened | smooth | smooth | ||||
| On CZ | Conidiophore | stipes (μm) | N/A | 30 − 100 (–120) × 2 − 3 | 40 − 60 (−80) × 3 − 4 | 25 − 135 × 2 − 3.5 | N/A |
| branching | biverticillate, monoverticillate | most biverticillate, monoverticillate symmetric | biverticillate, monoverticillate | ||||
| ornamentation | tuberculate | smooth | tuberculate | ||||
| Metulae | size (μm) | 6 − 8 × 2.5 − 3 | 6–8 (–12) × 2.5–3.5 | 5−9 × 2−3.5 | |||
| per verticil | 2–4 | N/A | 2–3 | ||||
| Phialides | size (μm) | 5 − 11 × 2 − 3 | 6–8 (−12) × 2 − 3 | 6 − 9 × 2 − 3 | |||
| per metula | 3–4 | N/A | 3–5 | ||||
| Conidia | size (μm) | 3–4 | (3−) 3.5 − 4.5 (−5) | 3–4 | |||
| shape | globose to sub-globose | globose to sub-globose | globose to sub-globose | ||||
| ornamentation | smooth | tuberculate | smooth | ||||
| On CYA * | Conidiophore | stipes (μm) | N/A | 20 − 70 × 2 − 3 | N/A | 35 − 85 × 2 − 2.5 | 30 − 45 × 2 − 3 |
| branching | biverticillate, monoverticillate | biverticillate, monoverticillate | biverticillate | ||||
| ornamentation | finely tuberculate | finely tuberculate | smooth | ||||
| Metulae | size (μm) | 8 − 12 × 2 − 3 | 6 − 9 × 2 − 2.5 | 12 − 20 × 2 − 3 | |||
| per verticil | 2–6 | 2–4 | 2–3 | ||||
| Phialides | size (μm) | 8 − 11 × 2 − 3 | 6.5 − 10.5 × 2 − 2.5 | 7 − 10 × 2 − 3 | |||
| per metula | 3–5 | 3–4 | 2–4 | ||||
| Conidia | size (μm) | 2–2.5 | 2–3 | 3 | |||
| shape | globose to sub-globose | globose to sub-globose | ellipsoidal to globose | ||||
| ornamentation | smooth | smooth | smooth-walled but verruculose with age | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nuankaew, S.; Chuaseeharonnachai, C.; Preedanon, S.; Somrithipol, S.; Saengkaewsuk, S.; Kwantong, P.; Phookongchai, S.; Srikitikulchai, P.; Kobmoo, N.; Wang, X.-C.; et al. Two Novel Species of Talaromyces Discovered in a Karst Cave in the Satun UNESCO Global Geopark of Southern Thailand. J. Fungi 2022, 8, 825. https://doi.org/10.3390/jof8080825
Nuankaew S, Chuaseeharonnachai C, Preedanon S, Somrithipol S, Saengkaewsuk S, Kwantong P, Phookongchai S, Srikitikulchai P, Kobmoo N, Wang X-C, et al. Two Novel Species of Talaromyces Discovered in a Karst Cave in the Satun UNESCO Global Geopark of Southern Thailand. Journal of Fungi. 2022; 8(8):825. https://doi.org/10.3390/jof8080825
Chicago/Turabian StyleNuankaew, Salilaporn, Charuwan Chuaseeharonnachai, Sita Preedanon, Sayanh Somrithipol, Supicha Saengkaewsuk, Papichaya Kwantong, Sarinya Phookongchai, Prasert Srikitikulchai, Noppol Kobmoo, Xin-Cun Wang, and et al. 2022. "Two Novel Species of Talaromyces Discovered in a Karst Cave in the Satun UNESCO Global Geopark of Southern Thailand" Journal of Fungi 8, no. 8: 825. https://doi.org/10.3390/jof8080825