
Nep1-like Proteins from Valsa mali Differentially Regulate Pathogen Virulence and Response to Abiotic Stresses


State Key Laboratory of Crop Stress Biology for Arid Areas, College of Plant Protection, Northwest A&F University, Yangling, Xianyang 712100, Shaanxi, China
*
Author to whom correspondence should be addressed.
J. Fungi 2021, 7(10), 830; https://doi.org/10.3390/jof7100830
Submission received: 13 July 2021
/
Revised: 27 September 2021
/
Accepted: 1 October 2021
/
Published: 4 October 2021
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Necrosis and ethylene-inducing peptide 1(Nep1)-like protein (NLP) is well known for its cytotoxicity and immunogenicity on dicotyledonous, and it has attracted large attention due to its gene expansion and functional diversification in numerous phytopathogens. Here, two NLP family proteins, VmNLP1 and VmNLP2, were identified in the pathogenic fungus Valsa mali. We showed that VmNLP2 but not VmNLP1 induced cell death when transiently expressed in Nicotiana benthamiana. VmNLP2 was also shown to induce cell death in apple leaves via the treatment of the Escherichia coli-produced recombinant protein. VmNLP1 and VmNLP2 transcripts were drastically induced at the early stage of V. mali infection, whereas only VmNLP2 was shown to be essential for pathogen virulence. We also found that VmNLP1 and VmNLP2 are required for maintaining the integrity of cell membranes, and they differentially contribute to V. mali tolerance to salt- and osmo-stresses. Notably, multiple sequence alignment revealed that the second histidine (H) among the conserved heptapeptide (GHRHDWE) of VmNLP2 is mutated to tyrosine (Y). When this tyrosine (Y) was substituted by histidine (H), the variant displayed enhanced cytotoxicity in N. benthamiana, as well as enhanced virulence on apple leaves, suggesting that the virulence role of VmNLP2 probably correlates to its cytotoxicity activity. We further showed that the peptide among VmNLP2, called nlp25 (VmNLP2), triggered strong immune response in Arabidopsis thaliana. This work demonstrates that NLPs from V. mali involve multiple biological roles, and shed new light on how intricately complex the functions of NLP might be.
1. Introduction
In plant–pathogen interactions, phytopathogens produce a variety of molecules, such as cell wall-degrading enzymes, effectors, and toxins, in order to establish effective infection on their host plants. Among them, effectors are usually produced to interfere with plant immunity by manipulating plant cellular processes [1,2]. On the contrary, plants can detect effectors, via either cell surface-localized or intracellular immune receptors, to activate immune responses and counteract pathogen infection [3,4]. Remarkably, there are quite a few molecules that can not only promote virulence, but also stimulate immune response on plants, of which, one of the most known examples represents the necrosis- and ethylene-inducing-like proteins (NLPs) [5,6].
Originally identified from culture filtrates of Fusarium oxysporum [7,8,9], NLPs are a superfamily of effector proteins widely distributed across bacteria, fungi and oomycetes. NLPs can be divided into three types, i.e., type 1, type 2 and type 3, which respectively contain two, four, and six conserved cysteine residues [8,9]. The typical bioactivity of NLPs is to cause cell death in dicotyledons [8,10]. Thus far, NLPs with cell death-inducing activity have been identified in various pathogens. Ottmann and colleagues have proven that NLPs can function as cytolytic toxins that promote the virulence of Pythium aphanidermatum [11]. Recently, the receptor for NLPs acting as toxins has been found to be plant sphingolipid glycosylinositol phosphorylceramides (GIPC) [12]. Notably, not all NLPs exhibit cell death-inducing activity in plants, and many characterized NLPs are noncytotoxic. For example, 5 out of 7 NLPs from Verticillium dahliae, 11 out of 19 NLPs from Phytophthora sojae, and all 10 tested NLPs from Hyaloperonospora arabidopsidis are not capable of inducing plant cell death [13,14,15]. Intriguingly, both cytotoxic and noncytotoxic NLPs can serve as immune elicitors. The cytotoxic NLPPcc from the phytopathogenic bacterium Pectobacterium carotovorum can activate the expression of immune marker genes HIN1 and PAD3 in tobacco [11]. The 10 noncytotoxic NLPs from H. arabidopsidis unexceptionally trigger immune responses when ectopically expressed in Arabidopsis [16]. Moreover, a conserved immunogenic peptide among NLPs has been identified, which can be recognized by the receptor-like protein RLP23 in Arabidopsis [17,18].
During the past two decades, increasing evidence shows that NLPs play essential roles in diverse plant–pathogen interactions. For instance, VdNLP1 and VdNLP2 from V. dahliae, both of which display cytotoxic activity, are induced during pathogen infection and are required for pathogen virulence [15]. VdNLP1 also affects pathogen vegetative growth and conidiospore production [15]. Besides, most HaNLP genes from H. arabidopsidis are up-regulated at the early infection stages, and the expression is accompanied by appressorium formation, suggesting that these HaNLP proteins may still be functioning in pathogen-plant attachment [13]. In addition, the NLPs from Phytophthora sojae are under strong positive selection, implying possible alternative roles in these proteins [14]. In spite of these, the function of noncytotoxic NLPs and cytotoxic NLPs outside pathogen–plant interactions remains largely undiscovered.
Apple Valsa canker caused by the ascomycete fungus Valsa mali, a weak parasitic pathogenic fungus, annually results in huge economic losses for the apple industry [19,20]. Though much efforts have been made to elucidate the infection mechanism of V. mali, the major virulence determinants of this fungus still remain largely puzzled. Much evidence has demonstrated that toxins play important roles during the V. mali infection of its apple host [21,22,23,24]. Considering that NLPs frequently function as cytolytic phytotoxins on dicotyledonous plants, we suspect that this member of proteins may play important roles in apple-V. mali pathosystem.
In this study, we successfully identified two NLP family proteins in V. mali, respectively named VmNLP1 and VmNLP2. We found that VmNLP2 but not VmNLP1 could induce cell death in the model plant Nicotiana benthamiana, and VmNLP2 can also cause cell death in the apple host. Both of these two genes were highly induced at early stage of V. mali infection. Target gene deletion revealed that only VmNLP2-deficient strain exhibited reduced virulence on apple host. Further analysis showed that VmNLP1 and VmNLP2 are differently involved in the maintenance of cell membrane integrity and pathogen tolerance to salt and osmotic stresses. Moreover, the immunogenic peptide among VmNLP2 displayed a potent activity to inducing immune response in A. thaliana, whereas the peptide among VmNLP1 showed a much weaker activity. Collectively, our results revealed that VmNLP2 is an essential virulence factor of V. mali, and we emphasized that the two NLP proteins from V. mali are functionally diversified in multifaceted biological roles, including their cytotoxic activity, contribution to pathogen virulence, and response to abiotic stresses.
2. Materials and Methods
2.1. Bioinformatics Analysis
The Nep1-like proteins from Valsa mali were identified by querying FoNep1 (Accession: AAC97382) from Fusarium oxysporum [25,26] against National Center for Biotechnology Information (NCBI) in V. mali genome [24]. Signal peptide prediction was performed by SignalP 4.1 [27]. Multi-sequence alignment was performed by Clustal W [28] with Blosum scoring matrix, and the parameters were set as follows: extending gap penalty (0.05), separation gap penalty (0.05), opening gap penalty (10), end gap penalty (10).
2.2. Strains and Plant Growth Conditions
The Valsa mali wild-type strain 03-8 and Sclerotinia sclerotiorum strain 1980 were stored in 20% glycerol at 4 °C. Escherichia coli strain DH5α used for plasmid construction was cultured on lysogeny broth (LB) medium at 37 °C, and Agrobacterium tumefaciens strain GV3101 used for agroinfiltration of plants was cultured on LB medium at 28 °C. Micropropagated apple (Malus domestica) rootstock plants GL-3 and Arabidopsis thaliana were cultured in an artificial climate chamber, 12 h/12 h light/dark at 22 °C. Nicotiana benthamiana was grown in an artificial greenhouse, 16 h/8 h light/dark at 22 °C.
2.3. Plasmid Construction
The open reading frames of VmNLP1, VmNLP2 or VmNLP2 without signal peptide were amplified from complementary DNA (cDNA) library of V. mali with gene-specific primers using Phanta Max Super-Fidelity DNA Polymerase (Vazyme, Nanjing, China). The hemagglutinin (HA)-tagged pCAMBIA1300 vector carrying kanamycin resistance gene was digested with KpnI and was subsequently ligated with the amplicons using ClonExpress II One-Step Cloning Kit (Vazyme, Nanjing, China). For generation of site-mutated proteins, the site-mutated fragments were amplified using mismatched base primers, which were subsequently cloned into pCAMBIA1300 vector using ClonExpress MultiS One-Step Cloning Kit (Vazyme, Nanjing, China). To generate gene complemented constructs, the PDL2 vector was linearized by XhoI digestion and further ligated with VmNLPs gene fragments that were amplified from V. mali genomic DNA containing their native promoters. The primers used for plasmid construction were listed in Supplementary Table S1. All PCR products were excised and purified from gel using Gel extraction Kit (OMEGA Biotech, Guangzhou, China). All constructs were sequence-verified by standard vector primers (Sangon Biotech, Shanghai, China).
2.4. A. tumefaciens-Mediated Transient Expression and Trypan Blue Staining
Four- to five-week-old N. benthamiana plants were used for agroinfiltration assays. The constructs were transformed into Agrobacterium tumefaciens GV3101 via electroporation. The colonies were selected by kanamycin (100 µg/mL), rifampicin (100 µg/mL), and confirmed by PCR. The individual correct transformant was cultured in LB medium at 28 °C with 220 rpm for 48 h. Cells were collected by centrifugation and were suspended in MES buffer (10 mM Magnesium chloride, 10 mM 2-(N-morpholino) ethane sulfonic acid (MES), 200 μM acetosyringone, pH 5.7). The suspended A. tumefaciens cells were incubated at room temperature for 3 h and were then mixed with P19 silencing suppressor. The final OD600 of suspensions were adjusted with MES buffer to 0.6. The suspensions were infiltrated into tobacco leaves using a needleless 1-mL syringe. Symptoms were observed 4–6 d post agroinfiltration. A. tumefaciens carrying INF1 and green fluorescent protein (GFP) were used as positive and negative control. Trypan blue staining was performed as described [29]. The experiment was repeated at least three times.
2.5. Western Blotting
N. benthamiana leaves transiently expressing indicated proteins were collected 36 h after infiltration. Lysing buffer (50 mM Tris, 150 mM NaCl, pH 7.5, 5 mM dithiothreitol (DTT), 0.5% TritonX-100, 1 mM phenylmethanesulfonyl fluoride (PMSF), 1% proteinase inhibitor cocktail (Sigma-Aldrich, St Louis, MO, USA)) was used to extracted total proteins. Ten minutes after centrifuge at 15,000× g (4 °C), the supernatant was collected to a new tube. The protein samples were boiled for 6 min in 2× sodium dodecyl sulfate (SDS) loading buffer and were then subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). Subsequently, the protein was transferred from gel to a polyvinylidene difluoride (PVDF) membrane under the transfer buffer (20 mM Tris, 150 mM glycine). The membrane was washed by Tris buffered saline (TBS), and TBST (TBS with 0.1% Tween 20) containing 5% nonfat dry milk was used to block the membrane at 50 rpm shaking for 2 h at room temperature. The membrane was then incubated overnight with mouse anti-HA (Abcam, Cambridge, UK, ab18181) monoclonal antibody and rinsed by TBST for three times, followed by staining with goat-anti mouse IgG (Abways, Shanghai, China) secondary antibody. Proteins were detected using ECL subtracted kit (GE Healthcare, RPN2235), as recommended by the manufacturer.
2.6. RNA Extraction and qRT-PCR Analysis
Total RNA from collected samples, including leaves of N. benthamiana and A. thaliana, apple twigs infected with V. mali, as well as mycelia from V. mali, was extracted using Quick RNA isolation Kit (Huayueyang, Beijing, China) according to the manufacturer’s instruction. RNA concentration and purity were quantified by NanoDrop Micro Photopeter (NanoDrop, Wilmington, DE, USA). The RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific, Waltham, MA, USA) was used for first-strand cDNA synthesis from 1 μg of total RNA, followed by quantitative reverse transcription-PCR (qRT-PCR) using Real Star Green Mixture (GenStar, Beijing, China) with specific primers. G6PDH in V. mali, Actin2 in A. thaliana or MdH in M. domestica were used as internal controls. Relative fold changes were calculated using the 2−ΔΔCT method [30].
2.7. Generation of Deletion Mutants, Complementation and Overexpression Transformants
To create deletion cassettes, the upstream and downstream flanking sequences amplified from V. mali genetic DNA were fused with NEO fragments amplified from vector pFL2, using double-joining PCR method [31]. The replacement fragment was transformed into protoplasts of V. mali via the polyethylene glycol (PEG)-mediated method [32]. The transformants were selected by G418-resistance followed by four types of PCR verifications to confirm the gene was knocked out. To obtain complementation transformants, pDL2 vector carrying VmNLPs or site-directed mutation variants (VmNLP1H127Y or VmNLP2Y137H) were transformed into protoplasts of VmNLPs deletion mutants. The hygromycin-resistance transformants were ascertained by PCR with primer pair VmNLP-5F/6R and sequencing. The overexpression transformants were achieved through fusing the fragments with the pDL2 vector containing the Magnaporthe grisea ribosomal protein 27 promoter, followed by transformation into the V. mali protoplasts. The transformants were filtered by hygromycin and total RNA was isolated to assess transcriptional levels of VmNLPs using qRT-PCR analysis. All primers used for gene knockout and overexpression are listed in Table S1.
2.8. Virulence and Phenotypic Analysis of Transformants
For virulence tests, fresh mycelial plugs collected from V. mali wild strain (WT), deletion mutants, complementation transformants and overexpression transformants were inoculated on leaves and twigs detached from Malus domestica Borkh. cv. Fuji [33]. Five millimeters mycelium plugs used in this assay were taken from the edge of growing colonies cultured on potato dextrose agar (PDA, 20% potato extract (w/v), 2% dextrose (w/v), 1.5% agar (w/v)) medium. Disease lesions were measured 2–3 days post-inoculation (dpi) for apple leaves and 4–5 dpi for apple twigs, followed by a data analysis. For vegetative growth detection, colony diameters of all strains cultured on PDA plates were calculated 2 days post-cultivation at 25 °C. For abiotic stress assays, PDA media were supplemented with congo red (CR, 400 μg/mL), sorbitol (1 M), sodium dodecyl sulfate (SDS, 0.01%) or KCl (0.3 M). Each experiment was conducted with three biological replicates, and all assays were repeated at least three times.
2.9. Expression and Purification of Recombinant VmNLP2 Protein
The coding sequence of VmNLP2 without signal peptide was amplified and cloned into pET28a vector (Novagen Inc., Madison, WI, USA) with NdeI and BamHI restriction sites, followed by transformation into E. coli BL21 (DE3). Cells were cultivated in LB medium at 37 °C until OD600 = 0.6~0.8. Subsequently, cells were introduced with 0.3 mM isopropyl-β-D-thiogalactopyrandoside (IPTG) for 24 h at 16 °C, and were then harvested by centrifugation at 5000× g. The collected bacteria cells were suspended in lysis buffer (20 mM sodium hydrogen phosphate, 300 mM NaCl, pH 7.4) containing 1 mM phenylmethanesulfonyl fluoride (PMSF), 1.98 mM β-mercaptoethanol and 1 mg ml−1 lysozyme followed by sonication. The supernatant of cells was collected via centrifugation at 4 °C and 10,000× g for 10 min and were subsequently loaded to Ni-NTA resin (Thermo Scientific, Waltham, MA, USA). VmNLP2 recombinant protein purification was finished by affinity chromatography according to the manufacturer’s instructions. The samples were subjected to silver staining using the Pierce™ Silver Stain for Mass Spectrometry Kit (Thermo Scientific, Waltham, MA, USA) following the manufacturer’s instructions, and were further analyzed by western blotting detection using anti-His monoclonal antibody (Abways, Shanghai, China).
2.10. Synthetic Peptides
Flg22 peptide was purchased from Genscript (Genscript Biotech Corporation, Nanjing, China; https://www.genscript.com.cn/) (accessed on 20 November 2020). nlp20 (VmNLP1) and nlp25 (VmNLP2) were synthesized by Sangon Biotech (Sangon Biotech, Shanghai, China; https://www.sangon.com/) (accessed on 17 August 2020). All peptides were dissolved by ultra-pure water to 500 μM for storage, and well diluted to 1 μM in each assay.
2.11. Measurement of Reactive Oxygen Burst
Reactive oxygen species (ROS) generation was detected as described [34]. Briefly, leaf discs were collected from A. thaliana seedlings and floated on 0.1 mL ddH2O overnight in a 96-well plate. Right before luminescence detection on a Varioskan LUX multimode microplate reader (Thermo Scientific), water was replaced with a 0.1 mL reaction solution (100 μM L-012 (Wako Chemical, Osaka, Japan), 20 μg mL−1 peroxidase (Solarbio, Beijing, China)), and 1 μM peptides. Eight biological replicates were used for each sample, and the experiments were repeated at least three times.
2.12. Inoculation Assays on A. thaliana
Five- to six-week-old A. thaliana seedlings were infiltrated with 1 μM peptides (nlp20 (VmNLP1), nlp25 (VmNLP2) or flg22) or ddH2O. Fresh plugs (diameter 5 mm) of S. sclerotiorum collected from PDA plates were inoculated on detached A. thaliana leaves 24 h post-infiltration. Inoculated leaves were incubated at a high humidity at 24 °C in 1/2 MS (Murashige & Skoog) medium (0.22% (w/v) MS basal medium (Phytotechnology, Lenexa, KS, USA)). The assay was performed at least three times.
3. Results
3.1. VmNLP2 but Not VmNLP1 Displays Cytotoxic Activity on N. benthamiana
To identify the NLP gene family in V. mali, a blast search of the V. mali genome [24] was performed with the FoNep1 from Fusarium oxysporum [25,26] in NCBI database. As a result, two different NLP homologues were identified and named VmNLP1 and VmNLP2, respectively (Table S2). Sequence analysis indicated that both VmNLP1 and VmNLP2 contain a predicted N-terminal signal peptide and two conserved cystine residues (Figure S1), for which they are classified as type 1 NLPs.
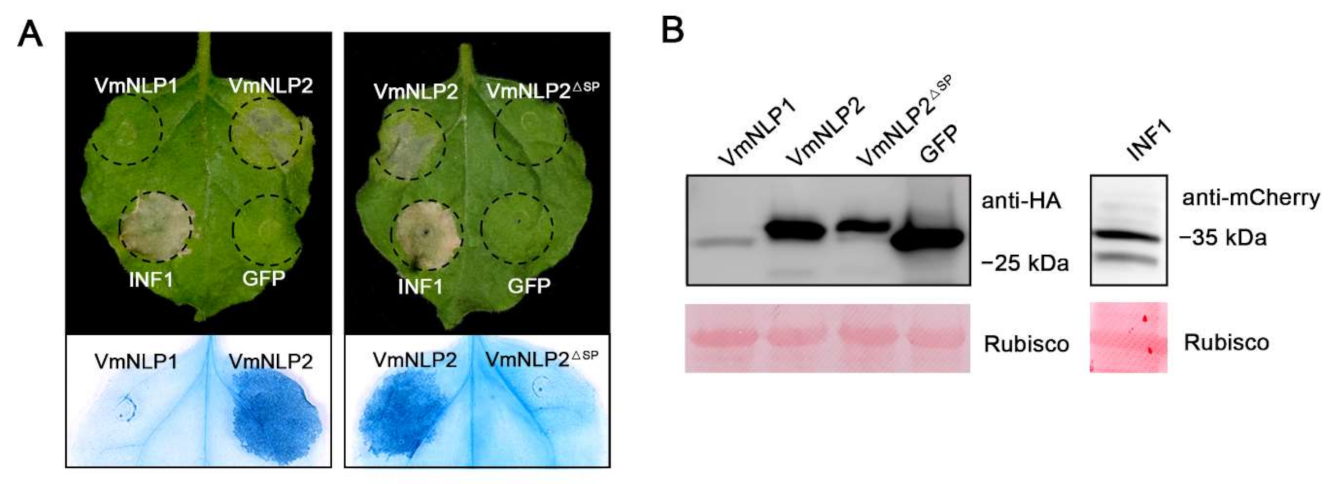
To test whether these two proteins exhibit cell death-inducing activity, hemagglutinin (HA)-tagged VmNLP1 and VmNLP2 were transiently expressed in N. benthamiana through agroinfiltration, with mCherry-tagged INF1 [35] and HA-tagged GFP used as positive and negative controls, respectively. As shown in Figure 1A, both VmNLP2 and INF1 induced visible cell death 5 days post agroinfiltration (dpa), whereas VmNLP1 and GFP control did not cause any necrosis symptoms. This indicates that VmNLP2 but not VmNLP1 displays cytotoxic activity in N. benthamiana. To test whether the signal peptide (SP) is needed for the cytotoxic activity of VmNLP2, VmNLP2 without SP (VmNLP2ΔSP) was transiently expressed in N. benthamiana. It showed that VmNLP2ΔSP cannot induce necrosis as full length VmNLP2 did (Figure 1A), suggesting that VmNLP2 requires its SP for cell death activation. Western blotting analysis showed that all proteins tested were successfully expressed in N. benthamiana (Figure 1B).
3.2. VmNLP2 Shows Cytotoxicity on Malus Domestica
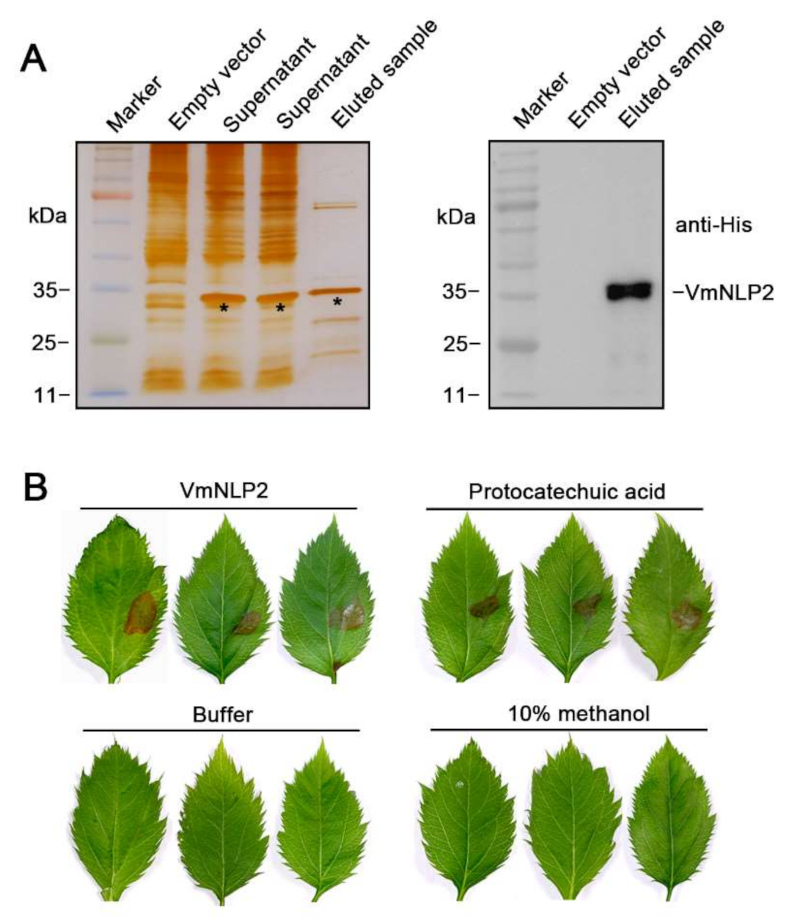
To examine whether VmNLP2 displays cytotoxic activity on its natural apple host, we firstly produced VmNLP2 recombinant protein in E. coli. N-terminal His-tagged VmNLP2 was verified through silver staining and western blotting detection (Figure 2A). Next, M. domestica leaves punctured by sterile syringes at the treatment site were dropped with 5 μM VmNLP2 purified protein, with buffer and 10% methanol used as negative controls, and 40 mM protocatechuic acid [22] used as a positive control. It illustrated that visible necrosis could be observed 24 h after treatment by VmNLP2 purified protein or protocatechuic acid, whereas neither buffer nor 10% methanol control caused any apparent symptoms (Figure 2B). Therefore, VmNLP2 exhibits cytotoxic activity on its apple host as well.
3.3. VmNLP2 Is Required for V. mali Virulence
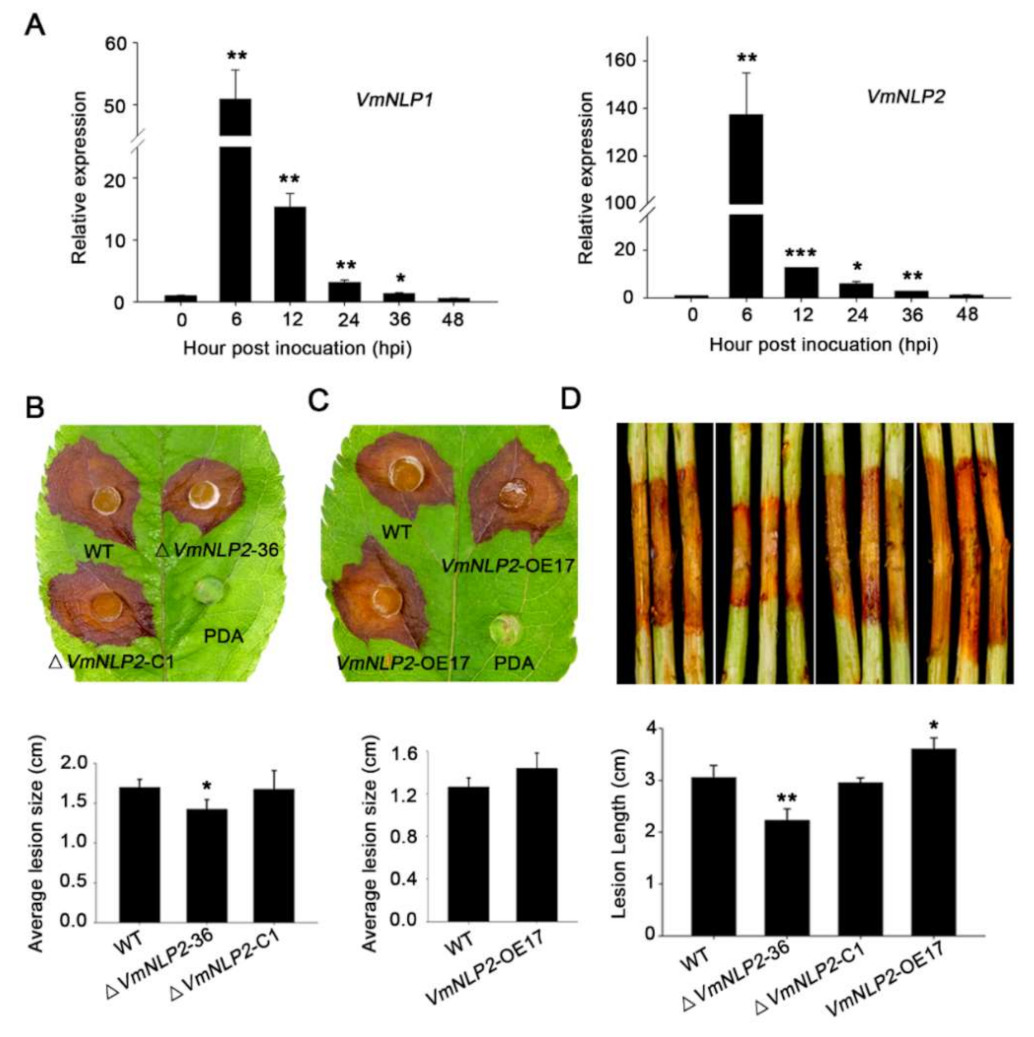
To determine the potential virulence roles of VmNLP1 and VmNLP2, we first analyzed their expression profiles during V. mali infection by qRT-PCR. As was shown, both VmNLP1 and VmNLP2 were markedly induced between 6 to 36 h post inoculation (hpi), and both of their transcripts peaked at 6 hpi (Figure 3A). This indicates that VmNLP1 and VmNLP2 are probably involved in the pathogenic process of V. mali, particularly during the early stage of pathogen infection.
We next tested whether VmNLP1 and VmNLP2 are required for pathogenicity of V. mali, for which, gene knockout mutants (Figure S2A) and corresponding complementation transformants (Figure S2B) of these two genes were generated via PEG-mediated protoplast transformation. Phenotypic analysis manifested that there was no obvious difference on filamentous growth in all deletion mutants (Figure S3A,B). Inoculation assays on apple host showed that ΔVmNLP1-10 and ΔVmNLP1-66 were as virulent as the WT strain (Figure S4A). Nevertheless, compared to WT strain, the virulence of ΔVmNLP2-36 was significantly impaired (Figure 3B). Moreover, by introducing VmNLP2 into ΔVmNLP2-36, the complementation transformant ΔVmNLP2-C1 caused comparable disease lesions to the WT strain (Figure 3B).
In order to further verify the above results, overexpression transformants were generated and their virulence were tested on detached apple leaves. The overexpression transformants of VmNLP1 (VmNLP1-OE6) and VmNLP2 (VmNLP2-OE17) were verified to show six-fold and eight-fold enhanced transcript levels, respectively (Supplementary Figure S5). Inoculation assays showed that lesion diameter caused by VmNLP1-OE6 was not conspicuously different from that caused by WT strain (Figure S4B). However, VmNLP2-OE17 caused larger disease lesion on apple leaves than that of the WT strain (Figure 3C). We also tested the virulence of these transformants on the detached twigs of apple. As expected, VmNLP1-deletion mutants did not result in apparent alterations in virulence compared to the WT strain (Figure S4C). Whereas, ΔVmNLP2-36 displayed markedly reduced lesion length compared with the WT strain, and the VmNLP2 complementation transformant was as virulent as the WT strain (Figure 3D). Consistently, the lesion length developed by VmNLP2-OE17 was greater than those that of the WT strain (Figure 3D). Taken together, these results demonstrated that VmNLP2 but not VmNLP1 is required for V. mali full virulence on the apple host.
3.4. VmNLP1 and VmNLP2 Differentially Contribute to Salt Tolerance, Osmotic and Membrane Stresses
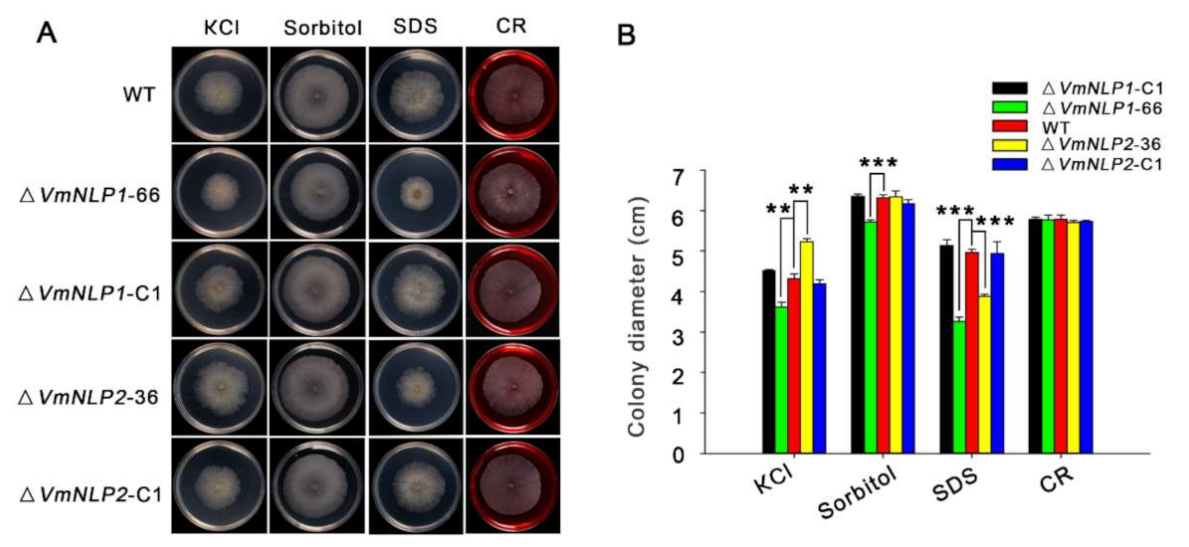
NLPs have also been demonstrated to occur in both pathogenic and non-pathogenic microorganisms [6,9], and it was assumed that potential function outside plant–pathogen interactions may exist for these proteins [10,15]. Therefore, apart from virulence, whether VmNLP1 and VmNLP2 are involved in V. mali responsiveness to other environmental factors such as abiotic stresses were subsequently tested. For this, V. mali WT strain, gene deletion mutants, and corresponding complementation transformants were cultured in PDA plants supplemented with 0.3 M KCl (salt stress), 1 M Sorbitol (osmotic stress), 0.01% SDS (cell membrane damaging agent), or 400 μg ml−1 CR (Congo red, cell wall inhibitor). We found that the vegetative growths of ΔVmNLP1-66 and ΔVmNLP2-36 were similar to that of the WT under the resistance to cell wall stress caused by CR. Interestingly, ΔVmNLP1-66 and ΔVmNLP2-36 displayed smaller vegetative growth under the treatment of SDS compared to the WT (Figure 4A), suggesting that both VmNLP1 and VmNLP2 participate in maintaining pathogen cell membrane integrity. In addition, under 1 M Sorbitol, the vegetative growth of ΔVmNLP1-66 but not ΔVmNLP2-36, was smaller than that of WT (Figure 4A), indicating that VmNLP1 also plays a protective role against osmo-tolerance. Unexpectedly, under 0.3 M KCl, the colony of ΔVmNLP1-66 was smaller, whereas ΔVmNLP2-36 colony was larger than that of the WT (Figure 4A), indicating that these two NLPs contrastingly regulate V. mali tolerance to salt stress. All the complement transformants showed similar tolerance levels to the WT under all tested abiotic stresses (Figure 4A,B). Collectively, VmNLP1 and VmNLP2 differentially participate in V. mali response to salt, osmotic, and membrane stresses.
3.5. The Second Histidine Residue among the Conserved Heptapeptide Is Mutated to Tyrosine in VmNLP2
The difference between VmNLP1 and VmNLP2 in multiple biological roles presented above prompted us to analyze their amino acid sequence. Fifteen fully studied NLPs derived from bacterial, fungal and oomycete species were selected for multiple sequence alignment. It revealed that several residues known to be essential for phytotoxic activity of NLPs, including K116, D117, as well as two cystine residues, are conserved in VmNLP1 and VmNLP2 (Figure S1). The characteristic heptapeptide motif “GHRHDWE” that are considered to be essential for necrosis-inducing activity of NLPs [9,11], are also conversed in almost all selected NLPs (Figure 5). Notably, however, for VmNLP2, the motif is “GHRYDWE”, in which the second histidine (H) among the heptapeptide is mutated to tyrosine (Y) (Figure 5), which may account for the distinct functions between VmNLP1 and VmNLP2.
3.6. Site-Directed Mutagenesis of Tyrosine to Histidine Promoted VmNLP2 Cytotoxicity and Virulence
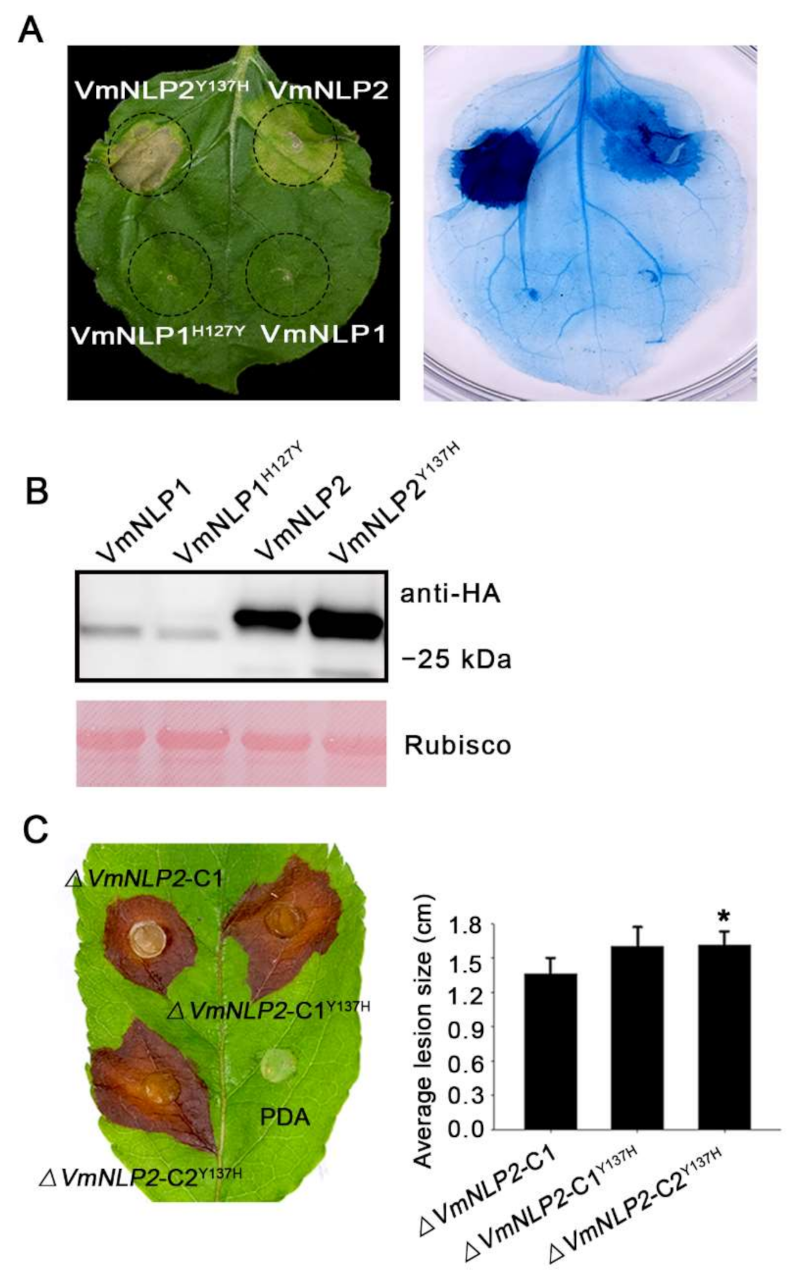
To assay whether the natural residue mutation among the heptapeptide of VmNLP2 affect its the cytotoxicity, constructs with this tyrosine substituted by histidine (VmNLP2Y137H) were generated. For comparison, we also replaced the Histidine among VmNLP1 with tyrosine (VmNLP1H127Y). When transiently expressed in N. benthamiana, neither VmNLP1 nor VmNLP1H127Y was able to induce plant cell death (Figure 6A). Interestingly, however, VmNLP2Y137H apparently exhibited enhanced cytotoxic activity than that of VmNLP2 (Figure 6A). All proteins expressed in N. benthamiana were verified by Western blotting (Figure 6B). Thus, this result suggests that the natural residue mutation among VmNLP2 can attenuate its cytotoxicity.
We next tested whether VmNLP1H127Y and VmNLP2Y137H can influence the virulence of V. mali. Site-directed mutation transformants ΔVmNLP1-CH127Y and ΔVmNLP2-CY137H were generated by introducing VmNLP1H127Y and VmNLP2Y137H fragments into the corresponding deletion mutants. All mutation transformants were ascertained by PCR analysis (Supplementary Figure S2B). Two of them were randomly selected for a virulence test. As was shown, ΔVmNLP1-CH127Y caused similar disease lesion as the complementation transformant ΔVmNLP1-C1 did, either on the detached apple twigs or leaves (Supplementary Figure S6A,B). However, compared with ΔVmNLP2-C1, ΔVmNLP2-CY137H showed an obvious increment in virulence when tested in detached apple leaves (Figure 6C). To be noted, ΔVmNLP2-CY137H showed no apparent change in virulence on detached twigs (Figure S7), which may be attributed to apple twigs being less sensitive. Altogether, the natural residue mutation among VmNLP2 can also attenuate its virulence, and VmNLP2 contribution to virulence is probably correlated to its cytotoxicity.
3.7. nlp25 among VmNLP2 Triggers Strong Immunity Response in A. thaliana
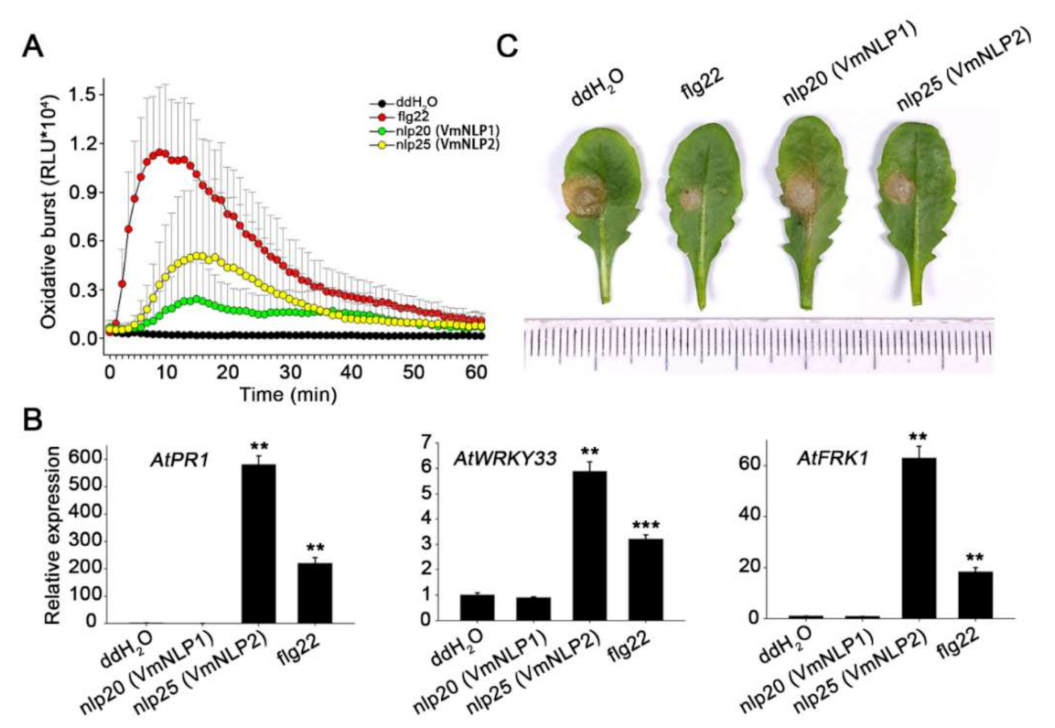
Since both cytotoxic and noncytotoxic NLPs possess a conserved immunogenic peptide [16,18], we next explored whether VmNLP1 and VmNLP2 contain such immunogenic peptides. Therefore, the peptides nlp20 derived from VmNLP1 and nlp25 derived from VmNLP2 (Figure S1) corresponding to nlp20 (NLPPp) [18] were synthesized. The peptides were subsequently used for measurement of immune responses, including ROS burst, expression of immune marker genes, and activation of disease resistance in Arabidopsis thaliana, a model plant that recognizes the nlp20 (NLPPp) peptide [17]. The well characterized flg22 peptide [43] was used as a positive control. The luminol-based chemiluminescence assay revealed that both nlp25 (VmNLP2) and nlp20 (VmNLP1) can elicit ROS burst in A. thaliana (Figure 7A). Notably, nlp20 (VmNLP1)-induced ROS level was not so strong as that of nlp25 (VmNLP2). We further treated with A. thaliana with 1 μΜ peptides and collected the samples 24 h later to analyze the expression of defense genes by qRT-PCR. As was shown in Figure 7B, the transcript levels of PR1, WRKY33 and FRK1 were markedly induced by treatment of nlp25 (VmNLP2), but not nlp20 (VmNLP1). In accordance, the disease lesions caused by the pathogenic fungus Sclerotinia sclerotiorum on Arabidopsis pre-treated with nlp25 (VmNLP2) but not nlp20 (VmNLP1) peptide were smaller than that of water control (Figure 7C). These results collectively indicate that nlp25 (VmNLP2) possesses a strong immunogenic activity, whereas nlp20 (VmNLP1) shows a much weaker and even no apparent immunogenic activity in A. thaliana.
We also tested whether the synthetic peptides exhibited immunogenic activities in N. benthamiana. As a result, none of them triggered apparent ROS bust (Supplementary Figure S8A), which is consistent with the fact that N. benthamiana lacks the RLP23 receptor for detection of nlp20 [17,18]. Most recently, two NLPs from Pythium oligandrum have been demonstrated to activate immune responses in N. benthamiana [44]. We next tested whether VmNLP recombinant proteins function similarly. As was shown, neither of them obviously induced ROS burst (Supplementary Figure S8A), nor did they activate the expression of immune marker genes like NbPR1, NbPR2 and NbPR4 in N. benthamiana (Supplementary Figure S8B). Thus, VmNLPs and the synthetic peptides therein display no immunogenic activity in the model plant N. benthamiana.
4. Discussion
The function of NLPs is varied due to the broad taxonomic distribution and rapid evolution [8,14]. In this study, we found that two NLPs from Valsa mali participate in a variety of biological roles and exhibit functional diversification. We have shown that only one of the two NLP proteins in V. mali, VmNLP2, possesses cytotoxic activity and contributes to virulence of V. mali. The non-cytotoxic VmNLP1 does not contribute to pathogenicity. However, we found that VmNLP1 positively regulates V. mali tolerance to salt, osmotic stress and cell membrane damaging agent. VmNLP2 was also shown to be involved in maintaining the integrity of cell membrane, but VmNLP1 and VmNLP2 played a distinct role in response to salt stress. Moreover, we showed that the immunogenic peptide nlp25 (VmNLP2) triggered strong immune responses in Arabidopsis thaliana, whereas nlp20 (VmNLP1) only elicited much weaker immune responses in A. thaliana.
Based on the ability to induce necrosis, NLPs can be divided into cytotoxic and non-cytotoxic NLPs. The cytotoxic NLPs can permeabilize plant plasma membranes of dicotyledons and lead to necrosis [7,11]. The non-cytotoxic NLPs are not capable of inducing cell necrosis, and their biological functions are largely mysterious [13,14]. Most necrotrophic phytopathogens have only one or two NLPs, and a majority of them, if not all, have been demonstrated to be cytotoxic. For example, Botrytis elliptica contains two NLPs which exhibit cytotoxic activity in many tested dicots [45], and two NLPs from Sclerotinia sclerotiorum can also induce necrosis when transiently expressed in tobacco leaves [36]. Here, we found that VmNLP2 but not VmNLP1 from the weakly parasitic V. mali exhibited cytotoxic activity in plants (Figure 1 and Figure 2). Actually, there are also some fungal species that contain necrotrophic phase encoding noncytotoxic NLPs on account of gene expansion, such as Verticillium dahlia [15] and Colletotrichum higginsianum [46]. It is worth noting that the mature protein of Botryis cinerea NLPs fused with the signal peptide of tobacco PR1a can trigger necrotic responses in N. benthamiana, whereas the BcNEP1 and BcNEP2 carrying their endogenous SP did not cause necrosis [37]. Likewise, it is possible that the inability of VmNLP1 to induce plant cell death may attribute to the potential inefficiency of its endogenous SP. Alternatively, the tertiary structure of VmNLP1 may not be conductive to its toxic function, similar to the case of HaNLP3, a typical noncytotoxic NLP from Hyaloperonospora arabidopsidis [13].
The transcripts of VmNLP1 and VmNLP2 were found to be highly induced at early stage of infection (Figure 3A), and we therefore hypothesized that these two genes may be involved in pathogenicity. Deletion of VmNLP2 reduced virulence on apple leaves and twigs (Figure 3B,D), however, the knocking out of VmNLP1 posed no apparent effect on pathogen virulence (Supplementary Figure S4A,C). In accordance, the overexpression of VmNLP2 in V. mali promoted pathogen infection in apple leaves (Figure 3C). This evidence together demonstrated that VmNLP2 is an important virulence factor of V. mali. This is similar to several characterized NLPs from other phytopathogens, which serve as virulence factors as well. For example, targeted deletion nip gene in E. carotovora subsp. carotovora compromised virulence in potato [42]; V. dahliae lacking either of the two cytotoxic NLPs showed significantly reduced virulence on tomato as well as on Arabidopsis plants [15]; overexpression of FoNep1 in the fungus Colletotrichum coccodes improved fungal virulence on Abutilon theophrasti [47]. Importantly, by multiple sequence alignment, we observed a residue mutation among the conserved heptapeptide motif of VmNLP2, in which the second histidine was mutated to tyrosine (Figure 5). We further found that the cytotoxic activity and virulence of VmNLP2 were simultaneously enhanced when the tyrosine residue was replaced with histidine residue (Figure 6), indicating a potential positive correlation between VmNLP2 cytotoxicity and its contribution to virulence. This result is reminiscent of NLPPcc from Pectobacterium carotovorum. It has been reported that the complementation of less cytotoxic NLPPcc variants in NLPPcc-deficient P. carotovorum strain resulted in correspondingly declined virulence [11]. These findings reinforce that NLPs can act as toxin-like virulence factors during the interaction of plant and pathogens.
Previous studies reveled that the large expansion of Nep1-like proteins family not only in oomycetes but also in fungi [6]. Thus, we were also interested in exploring the biological roles of VmNLPs apart from virulence and cytotoxic activity. In this study, we found that both VmNLP1 and VmNLP2 are involved in the salt tolerance and maintenance of cell membrane integrity of V. mali (Figure 4). In addition, VmNLP1 also participated in response to osmotic stress (Figure 4). Meanwhile, considering that NLPs are secreted proteins, we cannot exclude that VmNLP1 and VmNLP2 serve as ‘guards’ to differentially protect V. mali against abiotic stresses. A role of NLP in growth and conidiospore production has also been confirmed in the fungus V. dahlia [15]. Furthermore, the high expression of HaNLPs at an early infection stage led researchers to hypothesize it may play a role in zoospore attachment or primary contact between pathogen and host [13]. Our results here indicate that the two NLPs from V. mali are functionally diversified, not only in cytotoxicity and virulence, but also in regulation of pathogen responses to abiotic stresses.
In this study, we found that both the peptide nlp25 (VmNLP2) and nlp20 (VmNLP1) can elicit ROS burst in A. thaliana, however, ROS-triggered by nlp20 (VmNLP1) was much lower than that of nlp25 (VmNLP2) (Figure 7A). Additionally, nlp25 (VmNLP2) but not nlp20 (VmNLP1) activated the expression of defense-related genes and induced Arabidopsis resistance against Sclerotinia sclerotiorum (Figure 7B,C), though nlp20 (VmNLP1) is more similar to nlp20 derived from NLPPp [18]. This result further highlights the functional diversification of VmNLPs. We have also tested the response of apple leaves to these two peptides. Nevertheless, both nlp20 (VmNLP1) and nlp25 (VmNLP2) failed to trigger the expression of defense-related genes on apple, nor could they promote apple resistance to the V. mali infection (Supplementary Figure S9). It is possible that apple lacks a receptor like RLP23 from Arabidopsis [17] for detection of the immunogenic peptide among NLPs.
5. Conclusions
In summary, our study revealed that VmNLP2 is a cytotoxic NLP and it serves as an essential virulence factor of V. mali. Surprisingly, we showed that VmNLP1 and VmNLP2 play contrasting roles in V. mali response to cell membrane damaging agent, salt- and osmic-tolerance. The two NLPs in V. mali play functionally diversified roles in cytotoxicity, virulence, and regulation of pathogen tolerance to abiotic stresses, which offer a new understanding of the biological roles of the Nep1-like proteins.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/jof7100830/s1, Figure S1: Multiple sequence alignment of VmNLPs with its homologous sequences from other pathogen species using Clustal W. Figure S2: Validation of gene deletion and complementation mutants. Figure S3: Deletion of VmNLP1 or VmNLP2 posed no obvious influence on V. mali filamentous growth. Figure S4: Deletion of VmNLP1 showed no apparent influence on V. mali virulence. Figure S5: Relative transcript levels of VmNLP1 and VmNLP2 overexpression transformants. Figure S6: Substitution of the second histidine with tyrosine in the conserved heptapeptide among VmNLP1 did not affect virulence. Figure S7: Substitution of the tyrosine with histidine showed no apparent contribution to VmNLP2 virulence on apple twigs. Figure S8: VmNLP recombinant proteins and synthetic peptides are unable to trigger immune responses in N. benthamiana leaves. Figure S9: The peptide nlp20 (VmNLP1) and nlp25 (VmNLP2) are unable to trigger immune responses in apple leaves. Table S1: Primers used in this study. Table S2: Specification of VmNLP1 and VmNLP2.
Author Contributions
Conceptualization, L.H., J.L. and J.N.; methodology, J.L., J.N. and Y.C.; validation, J.L. and Y.C.; formal analysis, J.L. and J.N.; data curation, L.H. and J.L.; writing—original draft preparation, J.L.; writing—review and editing, L.H., J.N. and J.L.; visualization, J.L.; supervision, L.H.; project administration, L.H.; funding acquisition, L.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation-Xinjiang Joint Foundation of China (U1903206), and Major Scientific and Technological Projects of Shaanxi Province (2020zdzx03-03-01).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are available within the article and Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Boller, T.; He, S.Y. Innate immunity in plants: An arms race between pattern recognition receptors in plants and effectors in microbial pathogens. Science 2009, 324, 742–744. [Google Scholar] [CrossRef] [Green Version]
- De Jonge, R.; Bolton, M.D.; Thomma, B.P. How filamentous pathogens co-opt plants: The ins and outs of fungal effectors. Curr. Opin. Plant. Biol. 2011, 14, 400–406. [Google Scholar] [CrossRef]
- Toruño, T.Y.; Stergiopoulos, I.; Coaker, G. Plant-pathogen effectors: Cellular probes interfering with plant defenses in spatial and temporal manners. Annu. Rev. Phytopathol. 2016, 54, 419–441. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Feng, B.; Zhou, J.M.; Tang, D. Plant immune signaling: Advancing on two frontiers. J. Integr. Plant. Biol. 2020, 62, 2–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qutob, D.; Kemmerling, B.; Brunner, F.; Küfner, I.; Engelhardt, S.; Gust, A.A.; Luberacki, B.; Seitz, H.U.; Stahl, D.; Rauhut, T. Phytotoxicity and innate immune responses induced by Nep1-like proteins. Plant. Cell 2006, 18, 3721–3744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidl, M.F.; Van den Ackerveken, G. Activity and phylogenetics of the broadly occurring family of microbial Nep1-like proteins. Annu. Rev. Phytopathol. 2019, 57, 367–386. [Google Scholar] [CrossRef]
- Bailey, B.A. Purification of a protein from culture filtrates of Fusarium oxysporum that induces ethylene and necrosis in leaves of Erythroxylum coca. Phytopathology 1995, 85, 1250–1255. [Google Scholar] [CrossRef]
- Gijzen, M.; Nürnberger, T. Nep1-like proteins from plant pathogens: Recruitment and diversification of the NPP1 domain across taxa. Phytochemistry 2006, 67, 1800–1807. [Google Scholar] [CrossRef]
- Oome, S.; Van den Ackerveken, G. Comparative and functional analysis of the widely occurring family of Nep1-like proteins. Mol. Plant.-Microbe Interact. 2014, 27, 1081–1094. [Google Scholar] [CrossRef] [Green Version]
- Pemberton, C.L.; Salmond, G.P. The Nep1-like proteins—a growing family of microbial elicitors of plant necrosis. Mol. Plant. Pathol. 2004, 5, 353–359. [Google Scholar] [CrossRef]
- Ottmann, C.; Luberacki, B.; Küfner, I.; Koch, W.; Brunner, F.; Weyand, M.; Mattinen, L.; Pirhonen, M.; Anderluh, G.; Seitz, H.U. A common toxin fold mediates microbial attack and plant defense. Proc. Natl. Acad. Sci. USA 2009, 106, 10359–10364. [Google Scholar] [CrossRef] [Green Version]
- Lenarčič, T.; Albert, I.; Böhm, H.; Hodnik, V.; Pirc, K.; Zavec, A.B.; Podobnik, M.; Pahovnik, D.; Žagar, E.; Pruitt, R. Eudicot plant-specific sphingolipids determine host selectivity of microbial NLP cytolysins. Science 2017, 358, 1431–1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabral, A.; Oome, S.; Sander, N.; Küfner, I.; Nürnberger, T.; Van den Ackerveken, G. Nontoxic Nep1-like proteins of the downy mildew pathogen Hyaloperonospora arabidopsidis: Repression of necrosis-inducing activity by a surface-exposed region. Mol. Plant.-Microbe Interact. 2012, 25, 697–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, S.; Kong, G.; Qutob, D.; Yu, X.; Tang, J.; Kang, J.; Dai, T.; Wang, H.; Gijzen, M.; Wang, Y. The NLP toxin family in Phytophthora sojae includes rapidly evolving groups that lack necrosis-inducing activity. Mol. Plant.-Microbe Interact. 2012, 25, 896–909. [Google Scholar] [CrossRef] [Green Version]
- Santhanam, P.; van Esse, H.P.; Albert, I.; Faino, L.; Nürnberger, T.; Thomma, B.P. Evidence for functional diversification within a fungal NEP1-like protein family. Mol. Plant.-Microbe Interact. 2013, 26, 278–286. [Google Scholar] [CrossRef] [Green Version]
- Oome, S.; Raaymakers, T.M.; Cabral, A.; Samwel, S.; Böhm, H.; Albert, I.; Nürnberger, T.; Van den Ackerveken, G. Nep1-like proteins from three kingdoms of life act as a microbe-associated molecular pattern in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 16955–16960. [Google Scholar] [CrossRef] [Green Version]
- Albert, I.; Böhm, H.; Albert, M.; Feiler, C.E.; Imkampe, J.; Wallmeroth, N.; Brancato, C.; Raaymakers, T.M.; Oome, S.; Zhang, H. An RLP23–SOBIR1–BAK1 complex mediates NLP-triggered immunity. Nat. Plants 2015, 1, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Böhm, H.; Albert, I.; Oome, S.; Raaymakers, T.M.; Van den Ackerveken, G.; Nürnberger, T. A conserved peptide pattern from a widespread microbial virulence factor triggers pattern-induced immunity in Arabidopsis. PLoS Pathog. 2014, 10, e1004491. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Gao, X.; Du, Z.; Hu, Y.; Kang, Z.; Huang, L. Survey of apple Valsa canker in Weibei area of Shaanxi province. Acta Agric. Boreali-Occident. Sin. 2013, 1, 029. [Google Scholar]
- Wang, X.; Zang, R.; Yin, Z.; Kang, Z.; Huang, L. Delimiting cryptic pathogen species causing apple Valsa canker with multilocus data. Ecol. Evol. 2014, 4, 1369–1380. [Google Scholar] [CrossRef]
- Feng, Y.; Yin, Z.; Wu, Y.; Xu, L.; Du, H.; Wang, N.; Huang, L. LaeA controls virulence and secondary metabolism in apple canker pathogen Valsa mali. Front. Microbiol. 2020, 11, 2693. [Google Scholar] [CrossRef] [PubMed]
- Natsume, H.; Seto, H.; ōtake, N. Studies on apple canker disease. The necrotic toxins produced by Valsa ceratosperma. Agric. Biol. Chem. 1982, 46, 2101–2106. [Google Scholar] [CrossRef]
- Wang, C.; Li, C.; Li, B.; Li, G.; Dong, X.; Wang, G.; Zhang, Q. Toxins produced by Valsa mali var. mali and their relationship with pathogenicity. Toxins 2014, 6, 1139–1154. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Liu, H.; Li, Z.; Ke, X.; Dou, D.; Gao, X.; Song, N.; Dai, Q.; Wu, Y.; Xu, J.R. Genome sequence of Valsa canker pathogens uncovers a potential adaptation of colonization of woody bark. New Phytol. 2015, 208, 1202–1216. [Google Scholar] [CrossRef] [PubMed]
- Bailey, B.A.; Jennings, J.C.; Anderson, J.D. The 24-kDa protein from Fusarium oxysporum f. sp. erythroxyli: Occurrence in related fungi and the effect of growth medium on its production. Can. J. Microbiol. 1997, 43, 45–55. [Google Scholar] [CrossRef]
- Nelson, A. Sequence announcements. Plant. Mol. Biol. 1998, 38, 911. [Google Scholar] [CrossRef]
- Nielsen, H. Predicting secretory proteins with SignalP. In Protein Function Prediction; Springer: Berlin/Heidelberg, Germany, 2017; pp. 59–73. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Lukasik, E.; Gawehns, F.; Takken, F.L. The use of agroinfiltration for transient expression of plant resistance and fungal effector proteins in Nicotiana benthamiana leaves. In Plant Fungal Pathogens; Springer: Berlin/Heidelberg, Germany, 2012; pp. 61–74. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.-H.; Hamari, Z.; Han, K.-H.; Seo, J.-A.; Reyes-Domínguez, Y.; Scazzocchio, C. Double-joint PCR: A PCR-based molecular tool for gene manipulations in filamentous fungi. Fungal Genet. Biol. 2004, 41, 973–981. [Google Scholar] [CrossRef]
- Gao, J.; Li, Y.; Ke, X.; Kang, Z.; Huang, L. Development of genetic transformation system of Valsa mali of apple mediated by PEG. Wei Sheng Wu Xue Bao = Acta Microbiol. Sin. 2011, 51, 1194–1199. [Google Scholar]
- Wei, J.; Huang, L.; Gao, Z.; Ke, X.; Kang, Z. Laboratory evaluation methods of apple Valsa canker disease caused by Valsa ceratosperma sensu Kobayashi. Acta Phytopathologica Sin. 2010, 40, 14–20. [Google Scholar]
- Sang, Y.; Macho, A.P. Analysis of PAMP-triggered ROS burst in plant immunity. In Plant Pattern Recognition Receptors; Springer: Berlin/Heidelberg, Germany, 2017; pp. 143–153. [Google Scholar]
- Kamoun, S.; Van West, P.; Vleeshouwers, V.G.; De Groot, K.E.; Govers, F. Resistance of Nicotiana benthamiana to Phytophthora infestans is mediated by the recognition of the elicitor protein INF1. Plant. Cell 1998, 10, 1413–1425. [Google Scholar] [CrossRef] [Green Version]
- Dallal Bashi, Z.; Hegedus, D.D.; Buchwaldt, L.; Rimmer, S.R.; Borhan, M.H. Expression and regulation of Sclerotinia sclerotiorum necrosis and ethylene-inducing peptides (NEPs). Mol. Plant. Pathol. 2010, 11, 43–53. [Google Scholar] [CrossRef]
- Schouten, A.; Van Baarlen, P.; Van Kan, J.A. Phytotoxic Nep1-like proteins from the necrotrophic fungus Botrytis cinerea associate with membranes and the nucleus of plant cells. New Phytol. 2008, 177, 493–505. [Google Scholar] [CrossRef]
- Garcia, O.; Macedo, J.A.; Tibúrcio, R.; Zaparoli, G.; Rincones, J.; Bittencourt, L.M.; Ceita, G.O.; Micheli, F.; Gesteira, A.; Mariano, A.C. Characterization of necrosis and ethylene-inducing proteins (NEP) in the basidiomycete Moniliophthora perniciosa, the causal agent of witches’ broom in Theobroma cacao. Mycol. Res. 2007, 111, 443–455. [Google Scholar] [CrossRef]
- Kanneganti, T.-D.; Huitema, E.; Cakir, C.; Kamoun, S. Synergistic interactions of the plant cell death pathways induced by Phytophthora infestans Nep1-like protein PiNPP1. 1 and INF1 elicitin. Mol. Plant.-Microbe Interact. 2006, 19, 854–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qutob, D.; Kamoun, S.; Gijzen, M. Expression of a Phytophthora sojae necrosis-inducing protein occurs during transition from biotrophy to necrotrophy. Plant. J. 2002, 32, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Veit, S.; Wörle, J.M.; Nurnberger, T.; Koch, W.; Seitz, H.U. A novel protein elicitor (PaNie) from Pythium aphanidermatum induces multiple defense responses in carrot, Arabidopsis, and tobacco. Plant. Physiol. 2001, 127, 832–841. [Google Scholar] [CrossRef] [PubMed]
- Mattinen, L.; Tshuikina, M.; Mäe, A.; Pirhonen, M. Identification and characterization of Nip, necrosis-inducing virulence protein of Erwinia carotovora subsp.carotovora. Mol. Plant.-Microbe Interact. 2004, 17, 1366–1375. [Google Scholar] [CrossRef] [Green Version]
- Chinchilla, D.; Bauer, Z.; Regenass, M.; Boller, T.; Felix, G. The Arabidopsis receptor kinase FLS2 binds flg22 and determines the specificity of flagellin perception. Plant. Cell 2006, 18, 465–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.; Dong, X.; Li, J.; Wang, Y.; Cheng, Y.; Zhai, Y.; Li, X.; Wei, L.; Jing, M.; Dou, D. Type 2 Nep1-Like proteins from the biocontrol oomycete Pythium oligandrum suppress Phytophthora capsici infection in Solanaceous plants. J. Fungi 2021, 7, 496. [Google Scholar] [CrossRef]
- Staats, M.; Van Baarlen, P.; Schouten, A.; Van Kan, J.A. Functional analysis of NLP genes from Botrytis elliptica. Mol. Plant. Pathol. 2007, 8, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Kleemann, J.; Rincon-Rivera, L.J.; Takahara, H.; Neumann, U.; van Themaat, E.V.L.; van der Does, H.C.; Hacquard, S.; Stüber, K.; Will, I.; Schmalenbach, W. Sequential delivery of host-induced virulence effectors by appressoria and intracellular hyphae of the phytopathogen Colletotrichum higginsianum. PLoS Pathog. 2012, 8, e1002643. [Google Scholar] [CrossRef]
- Amsellem, Z.; Cohen, B.A.; Gressel, J. Engineering hypervirulence in a mycoherbicidal fungus for efficient weed control. Nat. Biotechnol. 2002, 20, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Transient expression of VmNLP2 induces cell death in N. benthamiana. (A) Representative N. benthamiana leaves showing cell death triggered by expressed proteins. VmNLP1, VmNLP2, VmNLP2 without signal peptide (VmNLP2ΔSP), GFP, and INF1 were transiently expressed in N. benthamiana. The leaves were photographed 5 d post agroinfiltration. Trypan blue staining was used to visualize the symptoms of cell death. (B) Immunoblot analysis on total proteins extracted from N. benthamiana with anti-HA or anti-mCherry antibodies. Rubisco protein was stained with Ponceau S as a loading control.
Figure 1.
Transient expression of VmNLP2 induces cell death in N. benthamiana. (A) Representative N. benthamiana leaves showing cell death triggered by expressed proteins. VmNLP1, VmNLP2, VmNLP2 without signal peptide (VmNLP2ΔSP), GFP, and INF1 were transiently expressed in N. benthamiana. The leaves were photographed 5 d post agroinfiltration. Trypan blue staining was used to visualize the symptoms of cell death. (B) Immunoblot analysis on total proteins extracted from N. benthamiana with anti-HA or anti-mCherry antibodies. Rubisco protein was stained with Ponceau S as a loading control.

Figure 2.
VmNLP2 recombinant protein displays cytotoxic activity on apple leaves. (A) Induced expression of VmNLP2 recombinant protein in E. coli. Silver-stained SDS-PAGE (left panel) and immunoblot analysis with anti-His antibody (right panel) were performed to detect VmNLP2 recombinant protein. Protein bands of interest were indicated by black asterisks. (B) Representative apple leaves showing cell death triggered by VmNLP2 purified protein. 5 μM VmNLP2 purified protein, buffer, 10% methanol and 40 mM protocatechuic acid were dropped on the needle-pricked area of apple leaves. Pictures were taken 2 days after treatment.
Figure 2.
VmNLP2 recombinant protein displays cytotoxic activity on apple leaves. (A) Induced expression of VmNLP2 recombinant protein in E. coli. Silver-stained SDS-PAGE (left panel) and immunoblot analysis with anti-His antibody (right panel) were performed to detect VmNLP2 recombinant protein. Protein bands of interest were indicated by black asterisks. (B) Representative apple leaves showing cell death triggered by VmNLP2 purified protein. 5 μM VmNLP2 purified protein, buffer, 10% methanol and 40 mM protocatechuic acid were dropped on the needle-pricked area of apple leaves. Pictures were taken 2 days after treatment.

Figure 3.
VmNLP2 is important for V. mali pathogenesis during infection on apple. (A) Expression profiles of VmNLP1 and VmNLP2 during V. mali infection of apple. Relative transcript levels of VmNLP1 and VmNLP2 at 0, 6, 12, 24, 36, and 48 hpi were determined by qRT-PCR. G6PDH gene in V. mali was used as an internal reference. Similar results were obtained in three independent experiments. Bars indicate standard deviations (SD) from three technical replicates. Asterisks indicate significant differences with 0 h based on Student’s t-test (*, p ≤ 0.05; **, p ≤ 0.01; ***, p ≤ 0.001). (B) Virulence tests of WT, VmNLP2 deletion mutant (ΔVmNLP2-36) and complementation transformant (ΔVmNLP2-C1). (C) Virulence tests of WT and VmNLP2 overexpression transformant (VmNLP2-OE17) on detached apple leaves. The experiment was repeated four times with six leaves per biological replicate. Representative photographs were taken at 2 dpi. Average lesion sizes (lesion diameter) were measured and the statistical analyses were performed by Student’s t-test (*, p ≤ 0.05). Bars indicate ±SD. (D) Virulence tests of WT, ΔVmNLP2-36, ΔVmNLP2-C1 and VmNLP2-OE17 on detached apple twigs. Similar results were obtained from at least three independent experiments. Representative photographs were taken 4 dpi. Lesion lengths were measured and the statistical analyses were performed by Student’s t-test (*, p ≤ 0.05; **, p ≤ 0.01). Bars indicate ±SD.
Figure 3.
VmNLP2 is important for V. mali pathogenesis during infection on apple. (A) Expression profiles of VmNLP1 and VmNLP2 during V. mali infection of apple. Relative transcript levels of VmNLP1 and VmNLP2 at 0, 6, 12, 24, 36, and 48 hpi were determined by qRT-PCR. G6PDH gene in V. mali was used as an internal reference. Similar results were obtained in three independent experiments. Bars indicate standard deviations (SD) from three technical replicates. Asterisks indicate significant differences with 0 h based on Student’s t-test (*, p ≤ 0.05; **, p ≤ 0.01; ***, p ≤ 0.001). (B) Virulence tests of WT, VmNLP2 deletion mutant (ΔVmNLP2-36) and complementation transformant (ΔVmNLP2-C1). (C) Virulence tests of WT and VmNLP2 overexpression transformant (VmNLP2-OE17) on detached apple leaves. The experiment was repeated four times with six leaves per biological replicate. Representative photographs were taken at 2 dpi. Average lesion sizes (lesion diameter) were measured and the statistical analyses were performed by Student’s t-test (*, p ≤ 0.05). Bars indicate ±SD. (D) Virulence tests of WT, ΔVmNLP2-36, ΔVmNLP2-C1 and VmNLP2-OE17 on detached apple twigs. Similar results were obtained from at least three independent experiments. Representative photographs were taken 4 dpi. Lesion lengths were measured and the statistical analyses were performed by Student’s t-test (*, p ≤ 0.05; **, p ≤ 0.01). Bars indicate ±SD.

Figure 4.
VmNLP1 and VmNLP2 differentially contribute to V. mali tolerance to salt, osmotic and membrane stresses. (A) Representative photographs showing the growth of all strains under abiotic stresses. WT, ΔVmNLP1-66, ΔVmNLP1-C1, ΔVmNLP2-36 and ΔVmNLP2-C1 were cultured on PDA medium and PDA supplemented with 0.3 M KCl, 1 M sorbitol, 0.01% sodium dodecyl sulfate (SDS) and 400 μg/mL Congo red (CR) at 25 °C for 3 days, respectively. The experiment was performed in triplicate, with five petri dishes in each repetition. (B) Colony diameter calculation for growth of the stains 3 days after inoculation on tested medium. Statistical analyses were performed by Student’s t-test (**, p ≤ 0.01; ***, p ≤ 0.001). Bars indicate ± SD of the mean of three replicates.
Figure 4.
VmNLP1 and VmNLP2 differentially contribute to V. mali tolerance to salt, osmotic and membrane stresses. (A) Representative photographs showing the growth of all strains under abiotic stresses. WT, ΔVmNLP1-66, ΔVmNLP1-C1, ΔVmNLP2-36 and ΔVmNLP2-C1 were cultured on PDA medium and PDA supplemented with 0.3 M KCl, 1 M sorbitol, 0.01% sodium dodecyl sulfate (SDS) and 400 μg/mL Congo red (CR) at 25 °C for 3 days, respectively. The experiment was performed in triplicate, with five petri dishes in each repetition. (B) Colony diameter calculation for growth of the stains 3 days after inoculation on tested medium. Statistical analyses were performed by Student’s t-test (**, p ≤ 0.01; ***, p ≤ 0.001). Bars indicate ± SD of the mean of three replicates.

Figure 5.
The second histidine residue among the conserved heptapeptide of VmNLP2 is mutated to tyrosine. A total of 17 NLPs, including VmNLP1 and VmNLP2 from Valsa mali, NLPpp from Parasitic phytophthora [18], FoNep1 from Fusarium oxysporum [25], SsNLP1 and SsNLP2 from Sclerotinia sclerotiorum [36], VdNLP1 and VdNLP2 from Verticillium dahlia [15], Bcnep1 and Bcnep2 from Botryis cinerea [37], MpNLP1 and MpNLP2 from Moniliophthora perniciosa [38], HaNLP3 from Hyaloperonospora arabidopsidis [13], PiNPP1.1 from Phytophthora infestans [39], PsojNIP from Phytophthora sojae [40], NLPpya from Pythium aphanidermatum [41], and NLPpcc from Pectobacterium carotovorum [42] were used for multiple sequence alignment by Clustal W. Part of the alignment results are shown. Conserved residues are shaded on a red background. Amino acid mutation among conserved heptapeptide is indicated by a black arrow.
Figure 5.
The second histidine residue among the conserved heptapeptide of VmNLP2 is mutated to tyrosine. A total of 17 NLPs, including VmNLP1 and VmNLP2 from Valsa mali, NLPpp from Parasitic phytophthora [18], FoNep1 from Fusarium oxysporum [25], SsNLP1 and SsNLP2 from Sclerotinia sclerotiorum [36], VdNLP1 and VdNLP2 from Verticillium dahlia [15], Bcnep1 and Bcnep2 from Botryis cinerea [37], MpNLP1 and MpNLP2 from Moniliophthora perniciosa [38], HaNLP3 from Hyaloperonospora arabidopsidis [13], PiNPP1.1 from Phytophthora infestans [39], PsojNIP from Phytophthora sojae [40], NLPpya from Pythium aphanidermatum [41], and NLPpcc from Pectobacterium carotovorum [42] were used for multiple sequence alignment by Clustal W. Part of the alignment results are shown. Conserved residues are shaded on a red background. Amino acid mutation among conserved heptapeptide is indicated by a black arrow.

Figure 6.
Substitution of the tyrosine with histidine promotes VmNLP2 cytotoxicity and contribution to virulence. (A) Representative N. benthamiana leaves showing cell death triggered by VmNLP2 and the mutation variant VmNLP2Y137H. VmNLP1, VmNLP2, VmNLP1 with introduced tyrosine substitution (VmNLP1H127Y) and VmNLP2 with introduced histidine substitution (VmNLP2Y137H) were transiently expressed in leaves. The cell death symptoms were assessed and photographed 4 days post-infiltration. Trypan blue staining was used to further visualize plant cell death. (B) Immunoblot analysis of proteins extracted from N. benthamiana transiently expressing indicated proteins. Rubisco protein was stained with Ponceau S as a loading control. (C) Virulence tests of complementation transformant (ΔVmNLP2-C1), or complementation transformant with introduced histidine substitution (ΔVmNLP2-C1Y137H, ΔVmNLP2-C2Y137H) on detached apple leaves. Similar results were obtained from three independent experiments. Photographs were taken at 2 dpi. Average lesions were calculated and the statistical analyses were performed by Student’s t-test (*, p ≤ 0.05). Bars indicate ± SD.
Figure 6.
Substitution of the tyrosine with histidine promotes VmNLP2 cytotoxicity and contribution to virulence. (A) Representative N. benthamiana leaves showing cell death triggered by VmNLP2 and the mutation variant VmNLP2Y137H. VmNLP1, VmNLP2, VmNLP1 with introduced tyrosine substitution (VmNLP1H127Y) and VmNLP2 with introduced histidine substitution (VmNLP2Y137H) were transiently expressed in leaves. The cell death symptoms were assessed and photographed 4 days post-infiltration. Trypan blue staining was used to further visualize plant cell death. (B) Immunoblot analysis of proteins extracted from N. benthamiana transiently expressing indicated proteins. Rubisco protein was stained with Ponceau S as a loading control. (C) Virulence tests of complementation transformant (ΔVmNLP2-C1), or complementation transformant with introduced histidine substitution (ΔVmNLP2-C1Y137H, ΔVmNLP2-C2Y137H) on detached apple leaves. Similar results were obtained from three independent experiments. Photographs were taken at 2 dpi. Average lesions were calculated and the statistical analyses were performed by Student’s t-test (*, p ≤ 0.05). Bars indicate ± SD.

Figure 7.
nlp25 peptide derived from VmNLP2 triggers plant immunity responses in A. thaliana. (A) Oxidative burst in A. thaliana treated with 1 μΜ peptides (nlp20 (VmNLP1), nlp25 (VmNLP2), flg22) or ddH2O. Values are showed as relative light units (RLU). (B) VmNLP2 activated transcript accumulation of defense related genes in A. thaliana. The leaves were infiltrated with 1 μΜ peptides (nlp20 (VmNLP1), nlp25 (VmNLP2), flg22) or ddH2O control. Gene expression was assessed 24 hpi by qRT-PCR. Actin2 was used as an endogenous reference. Means and SDs were calculated from three independent biological replicates. Significant differences based on student’s t-test are indicated by the asterisks (**, p ≤ 0.01; ***, p ≤ 0.001). (C) nlp25 (VmNLP2) promoted A. thaliana resistance against S. sclerotiorum. The leaves were infiltrated with 1μΜ synthetic peptide nlp20 (VmNLP1) or nlp25 (VmNLP2) 24 h before inoculation of S. sclerotiorum. The flg22 peptide and ddH2O were used as positive and negative control, respectively. The experiments were performed three times with similar results.
Figure 7.
nlp25 peptide derived from VmNLP2 triggers plant immunity responses in A. thaliana. (A) Oxidative burst in A. thaliana treated with 1 μΜ peptides (nlp20 (VmNLP1), nlp25 (VmNLP2), flg22) or ddH2O. Values are showed as relative light units (RLU). (B) VmNLP2 activated transcript accumulation of defense related genes in A. thaliana. The leaves were infiltrated with 1 μΜ peptides (nlp20 (VmNLP1), nlp25 (VmNLP2), flg22) or ddH2O control. Gene expression was assessed 24 hpi by qRT-PCR. Actin2 was used as an endogenous reference. Means and SDs were calculated from three independent biological replicates. Significant differences based on student’s t-test are indicated by the asterisks (**, p ≤ 0.01; ***, p ≤ 0.001). (C) nlp25 (VmNLP2) promoted A. thaliana resistance against S. sclerotiorum. The leaves were infiltrated with 1μΜ synthetic peptide nlp20 (VmNLP1) or nlp25 (VmNLP2) 24 h before inoculation of S. sclerotiorum. The flg22 peptide and ddH2O were used as positive and negative control, respectively. The experiments were performed three times with similar results.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, J.; Nie, J.; Chang, Y.; Huang, L. Nep1-like Proteins from Valsa mali Differentially Regulate Pathogen Virulence and Response to Abiotic Stresses. J. Fungi 2021, 7, 830. https://doi.org/10.3390/jof7100830
AMA Style
Liu J, Nie J, Chang Y, Huang L. Nep1-like Proteins from Valsa mali Differentially Regulate Pathogen Virulence and Response to Abiotic Stresses. Journal of Fungi. 2021; 7(10):830. https://doi.org/10.3390/jof7100830
Chicago/Turabian StyleLiu, Jianying, Jiajun Nie, Yali Chang, and Lili Huang. 2021. "Nep1-like Proteins from Valsa mali Differentially Regulate Pathogen Virulence and Response to Abiotic Stresses" Journal of Fungi 7, no. 10: 830. https://doi.org/10.3390/jof7100830
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.