
Soil Mycobiome Is Shaped by Vegetation and Microhabitats: A Regional-Scale Study in Southeastern Brazil

 ,
,  , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
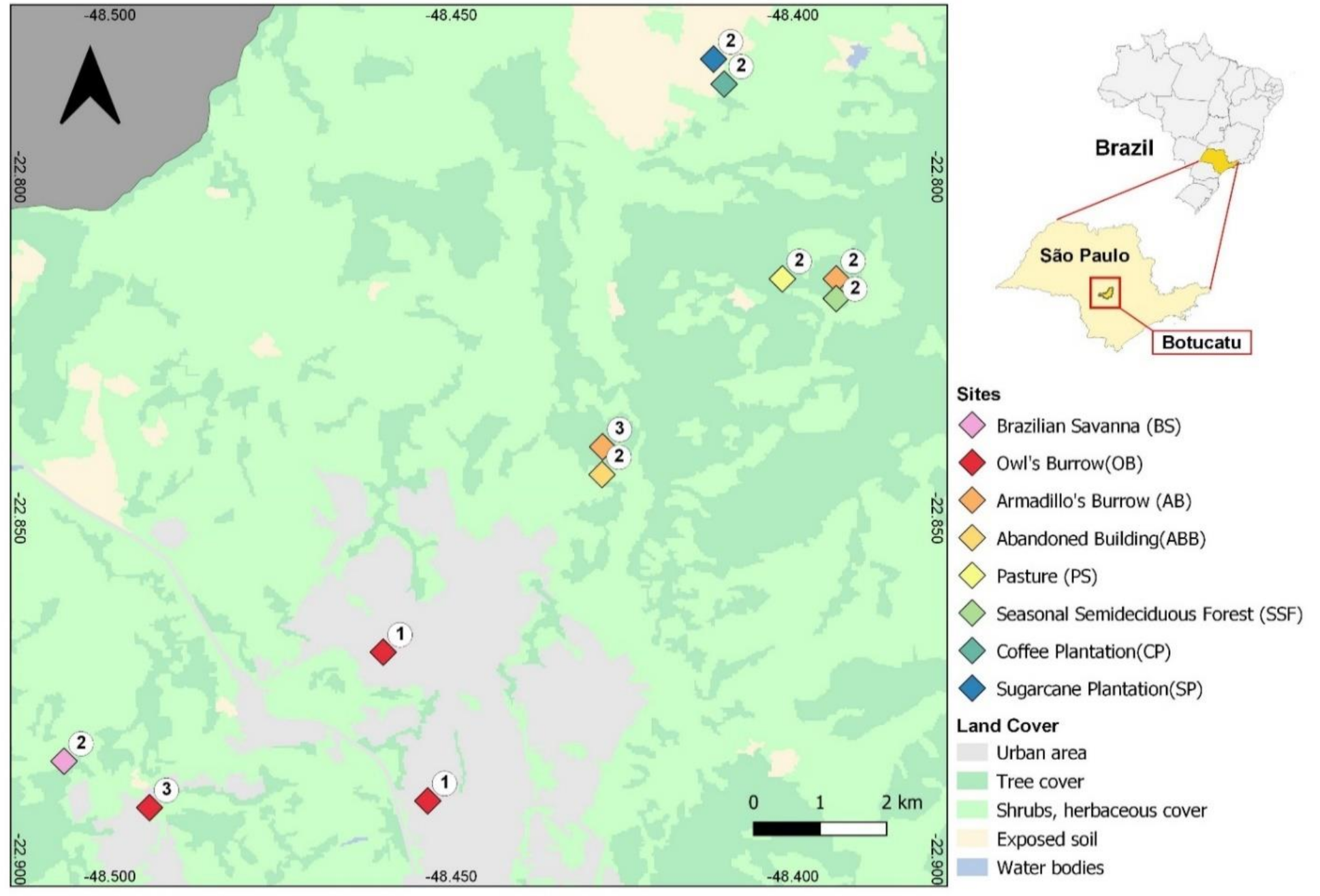
2.1. Study Area and Sampling
2.2. DNA Extraction and Sample Processing
2.3. Data Processing
2.4. Statistical Analysis
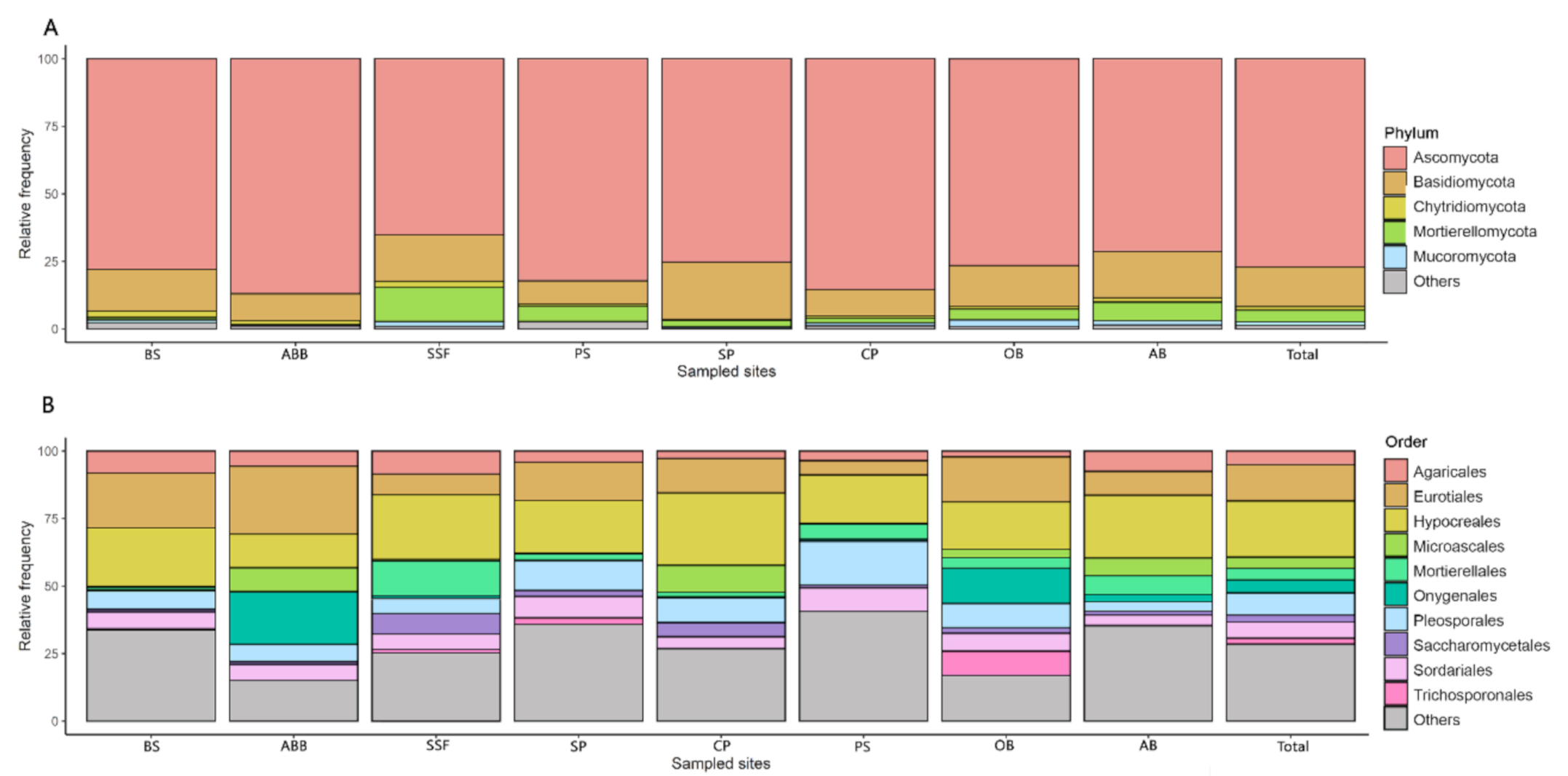
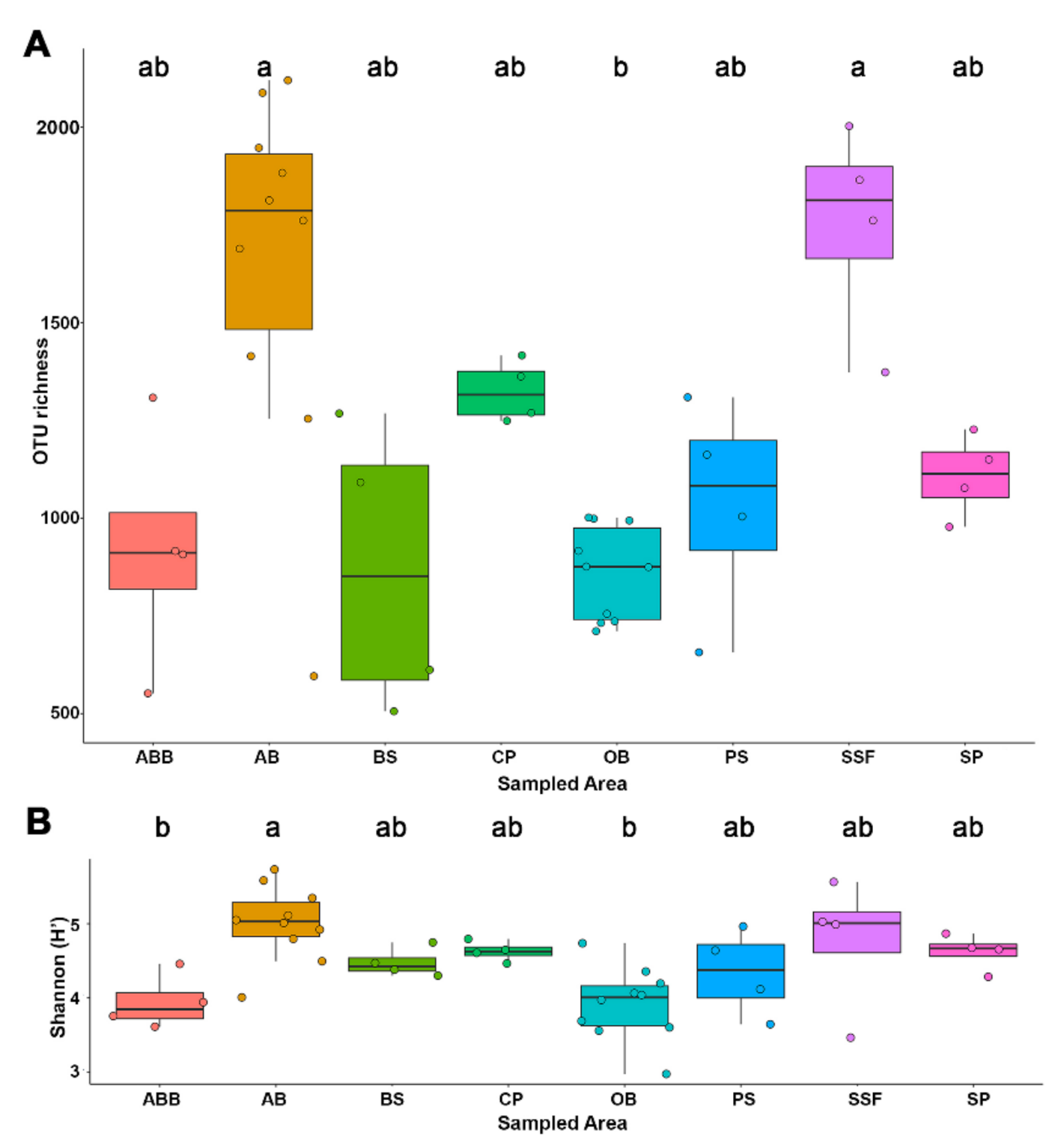
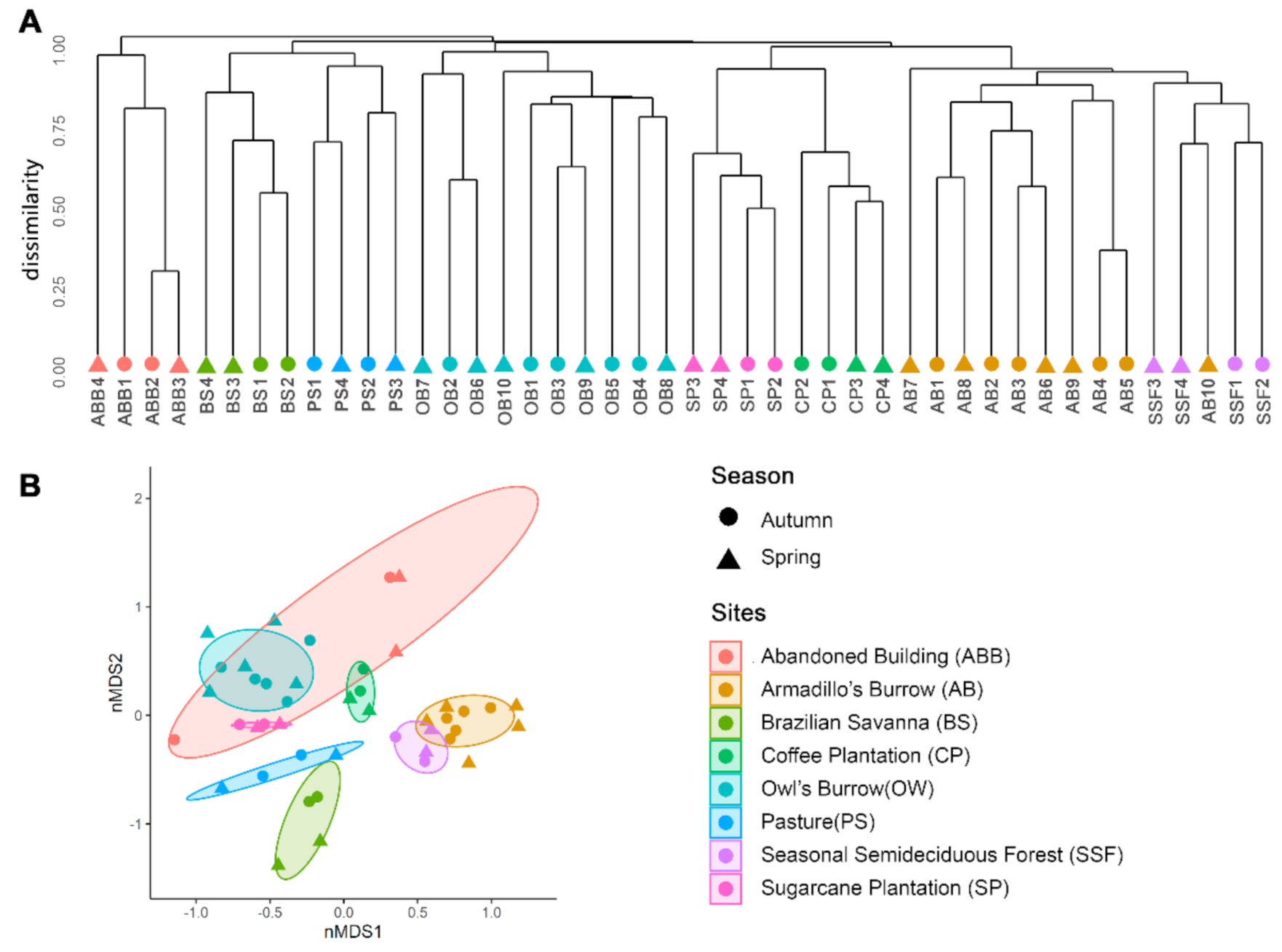
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heitman, J.; Howlett, B.; Crous, P.W.; Stukenbrock, E.; James, T.; Gow, N.A.R. The Fungal Kingdom; American Society for Microbiology: Washington, DC, USA, 2017; ISBN 978-1-55581-957-6. [Google Scholar]
- Hawksworth, D.L.; Lücking, R. Fungal Diversity Revisited: 2.2 to 3.8 Million Species. In The Fungal Kingdom; Heitman, H., Crous, S., James, G., Eds.; American Society of Microbiology: Washington, DC, USA, 2017; pp. 79–95. ISBN 978-1-55581-957-6. [Google Scholar]
- Lee Taylor, D.; Sinsabaugh, R.L. The Soil Fungi. In Soil Microbiology, Ecology and Biochemistry; Elsevier: Amsterdam, The Netherlands, 2015; pp. 77–109. ISBN 978-0-12-415955-6. [Google Scholar]
- Naranjo-Ortiz, M.A.; Gabaldón, T. Fungal Evolution: Major Ecological Adaptations and Evolutionary Transitions. Biol. Rev. 2019, 94, 1443–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawksworth, D.L. The Fungal Dimension of Biodiversity: Magnitude, Significance, and Conservation. Mycol. Res. 1991, 95, 641–655. [Google Scholar] [CrossRef]
- Blackwell, M. The Fungi: 1, 2, 3 … 5.1 Million Species? Am. J. Bot. 2011, 98, 426–438. [Google Scholar] [CrossRef]
- Taylor, D.L.; Hollingsworth, T.N.; McFarland, J.W.; Lennon, N.J.; Nusbaum, C.; Ruess, R.W. A First Comprehensive Census of Fungi in Soil Reveals Both Hyperdiversity and Fine-Scale Niche Partitioning. Ecol. Monogr. 2014, 84, 3–20. [Google Scholar] [CrossRef]
- Lücking, R.; Aime, M.C.; Robbertse, B.; Miller, A.N.; Aoki, T.; Ariyawansa, H.A.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Fungal Taxonomy and Sequence-Based Nomenclature. Nat. Microbiol. 2021, 6, 540–548. [Google Scholar] [CrossRef]
- O’Brien, H.E.; Parrent, J.L.; Jackson, J.A.; Moncalvo, J.-M.; Vilgalys, R. Fungal Community Analysis by Large-Scale Sequencing of Environmental Samples. AEM 2005, 71, 5544–5550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastida, F.; Eldridge, D.J.; García, C.; Kenny Png, G.; Bardgett, R.D.; Delgado-Baquerizo, M. Soil Microbial Diversity–Biomass Relationships Are Driven by Soil Carbon Content across Global Biomes. ISME J. 2021, 15, 2081–2091. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, H.; Tian, P.; Yao, X.; Sun, H.; Wang, Q.; Delgado-Baquerizo, M. Decoupled Diversity Patterns in Bacteria and Fungi across Continental Forest Ecosystems. Soil Biol. Biochem. 2020, 144, 107763. [Google Scholar] [CrossRef]
- Buée, M.; Reich, M.; Murat, C.; Morin, E.; Nilsson, R.H.; Uroz, S.; Martin, F. 454 Pyrosequencing Analyses of Forest Soils Reveal an Unexpectedly High Fungal Diversity: Research. New Phytol. 2009, 184, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Orgiazzi, A.; Bianciotto, V.; Bonfante, P.; Daghino, S.; Ghignone, S.; Lazzari, A.; Lumini, E.; Mello, A.; Napoli, C.; Perotto, S.; et al. 454 Pyrosequencing Analysis of Fungal Assemblages from Geographically Distant, Disparate Soils Reveals Spatial Patterning and a Core Mycobiome. Diversity 2013, 5, 73–98. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global Diversity and Geography of Soil Fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [Green Version]
- Peay, K.G.; Kennedy, P.G.; Talbot, J.M. Dimensions of Biodiversity in the Earth Mycobiome. Nat. Rev. Microbiol. 2016, 14, 434–447. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.H.; Anslan, S.; Bahram, M.; Wurzbacher, C.; Baldrian, P.; Tedersoo, L. Mycobiome Diversity: High-Throughput Sequencing and Identification of Fungi. Nat. Rev. Microbiol. 2019, 17, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Větrovský, T.; Kohout, P.; Kopecký, M.; Machac, A.; Man, M.; Bahnmann, B.D.; Brabcová, V.; Choi, J.; Meszárošová, L.; Human, Z.R.; et al. A Meta-Analysis of Global Fungal Distribution Reveals Climate-Driven Patterns. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Amend, A.S.; Seifert, K.A.; Samson, R.; Bruns, T.D. Indoor Fungal Composition Is Geographically Patterned and More Diverse in Temperate Zones than in the Tropics. Proc. Natl. Acad. Sci. USA 2010, 107, 13748–13753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finlay, B.J. Global Dispersal of Free-Living Microbial Eukaryote Species. Science 2002, 296, 1061–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemergut, D.R.; Schmidt, S.K.; Fukami, T.; O’Neill, S.P.; Bilinski, T.M.; Stanish, L.F.; Knelman, J.E.; Darcy, J.L.; Lynch, R.C.; Wickey, P.; et al. Patterns and Processes of Microbial Community Assembly. Microbiol. Mol. Biol. Rev. 2013, 77, 342–356. [Google Scholar] [CrossRef] [Green Version]
- Geml, J. Soil Fungal Communities Reflect Aspect-Driven Environmental Structuring and Vegetation Types in a Pannonian Forest Landscape. Fungal Ecol. 2019, 39, 63–79. [Google Scholar] [CrossRef]
- Parker, S.S. Buried Treasure: Soil Biodiversity and Conservation. Biodivers. Conserv. 2010, 19, 3743–3756. [Google Scholar] [CrossRef]
- Targulian, V.O.; Arnold, R.W.; Miller, B.A.; Brevik, E.C. Pedosphere. In Encyclopedia of Ecology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 162–168. ISBN 978-0-444-64130-4. [Google Scholar]
- Corlett, R.T. The Anthropocene Concept in Ecology and Conservation. Trends Ecol. Evol. 2015, 30, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Pärtel, M.; Öpik, M.; Moora, M.; Tedersoo, L.; Szava-Kovats, R.; Rosendahl, S.; Rillig, M.C.; Lekberg, Y.; Kreft, H.; Helgason, T.; et al. Historical Biome Distribution and Recent Human Disturbance Shape the Diversity of Arbuscular Mycorrhizal Fungi. New Phytol. 2017, 216, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated Modern Human–Induced Species Losses: Entering the Sixth Mass Extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [Green Version]
- McGill, B.J.; Dornelas, M.; Gotelli, N.J.; Magurran, A.E. Fifteen Forms of Biodiversity Trend in the Anthropocene. Trends Ecol. Evol. 2015, 30, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Seddon, N.; Mace, G.M.; Naeem, S.; Tobias, J.A.; Pigot, A.L.; Cavanagh, R.; Mouillot, D.; Vause, J.; Walpole, M. Biodiversity in the Anthropocene: Prospects and Policy. Proc. R. Soc. B 2016, 283, 20162094. [Google Scholar] [CrossRef] [Green Version]
- Guerra, C.A.; Rosa, I.M.D.; Valentini, E.; Wolf, F.; Filipponi, F.; Karger, D.N.; Nguyen Xuan, A.; Mathieu, J.; Lavelle, P.; Eisenhauer, N. Global Vulnerability of Soil Ecosystems to Erosion. Landsc. Ecol. 2020, 35, 823–842. [Google Scholar] [CrossRef] [Green Version]
- Tibbett, M.; Fraser, T.D.; Duddigan, S. Identifying Potential Threats to Soil Biodiversity. PeerJ 2020, 8, e9271. [Google Scholar] [CrossRef]
- Gehring, C.A.; Wolf, J.E.; Theimer, T.C. Terrestrial Vertebrates Promote Arbuscular Mycorrhizal Fungal Diversity and Inoculum Potential in a Rain Forest Soil. Ecol. Lett. 2002, 5, 540–548. [Google Scholar] [CrossRef] [Green Version]
- Clarke, L.J.; Weyrich, L.S.; Cooper, A. Reintroduction of Locally Extinct Vertebrates Impacts Arid Soil Fungal Communities. Mol. Ecol. 2015, 24, 3194–3205. [Google Scholar] [CrossRef]
- Dundas, S.J.; Hopkins, A.J.M.; Ruthrof, K.X.; Tay, N.E.; Burgess, T.I.; Hardy, G.E.S.J.; Fleming, P.A. Digging Mammals Contribute to Rhizosphere Fungal Community Composition and Seedling Growth. Biodivers. Conserv. 2018, 27, 3071–3086. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth. BioScience 2001, 51, 933. [Google Scholar] [CrossRef]
- Mueller, J.C.; Kuhl, H.; Boerno, S.; Tella, J.L.; Carrete, M.; Kempenaers, B. Evolution of Genomic Variation in the Burrowing Owl in Response to Recent Colonization of Urban Areas. Proc. R. Soc. B 2018, 285, 20180206. [Google Scholar] [CrossRef]
- Levey, D.J.; Duncan, R.S.; Levins, C.F. Use of Dung as a Tool by Burrowing Owls. Nature 2004, 431, 39. [Google Scholar] [CrossRef]
- Vizcaíno, S.F.; Loughry, W.J. (Eds.) The Biology of the Xenarthra; University Press of Florida: Gainesville, FL, USA, 2008; ISBN 978-0-8130-3165-1. [Google Scholar]
- Soibelzon, E. Using Paleoclimate and the Fossil Record to Explain Past and Present Distributions of Armadillos (Xenarthra, Dasypodidae). J. Mammal. Evol 2019, 26, 61–70. [Google Scholar] [CrossRef]
- Loughry, W.J.; Perez-Heydrich, C.; McDonough, C.M.; Oli, M.K. Population Dynamics and Range Expansion in Nine-Banded Armadillos. PLoS ONE 2013, 8, e68311. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Gastélum, L.; Romero-Olivares, A.L.; Escalante, A.E.; Rocha-Olivares, A.; Brizuela, C.; Riquelme, M. Impact of Seasonal Changes on Fungal Diversity of a Semi-Arid Ecosystem Revealed by 454 Pyrosequencing. FEMS Microbiol. Ecol. 2015, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouse, J.W., Jr.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with Erts. NASA Spec. Publ. 1974, 351, 309. [Google Scholar]
- Pettorelli, N.; Ryan, S.; Mueller, T.; Bunnefeld, N.; Jedrzejewska, B.; Lima, M.; Kausrud, K. The Normalized Difference Vegetation Index (NDVI): Unforeseen Successes in Animal Ecology. Clim. Res. 2011, 46, 15–27. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.-M.; Tucker, C.J.; Stenseth, N.C. Using the Satellite-Derived NDVI to Assess Ecological Responses to Environmental Change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef]
- Myneni, R.B.; Keeling, C.D.; Tucker, C.J.; Asrar, G.; Nemani, R.R. Increased Plant Growth in the Northern High Latitudes from 1981 to 1991. Nature 1997, 386, 698–702. [Google Scholar] [CrossRef]
- Congedo, L. Semi-Automatic Classification Plugin User Manual. 2014. Available online: https://www.researchgate.net/publication/265031337_Semi-Automatic_Classification_Plugin_User_Manual (accessed on 17 July 2021).
- Taylor, D.L.; Walters, W.A.; Lennon, N.J.; Bochicchio, J.; Krohn, A.; Caporaso, J.G.; Pennanen, T. Accurate Estimation of Fungal Diversity and Abundance through Improved Lineage-Specific Primers Optimized for Illumina Amplicon Sequencing. Appl. Environ. Microbiol. 2016, 82, 7217–7226. [Google Scholar] [CrossRef] [Green Version]
- Alvarado, P.; Teixeira, M.d.M.; Andrews, L.; Fernandez, A.; Santander, G.; Doyle, A.; Perez, M.; Yegres, F.; Barker, B.M. Detection of Coccidioides Posadasii from Xerophytic Environments in Venezuela Reveals Risk of Naturally Acquired Coccidioidomycosis Infections. Emerg. Microbes Infect. 2018, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Palmer, J.M.; Jusino, M.A.; Banik, M.T.; Lindner, D.L. Non-Biological Synthetic Spike-in Controls and the AMPtk Software Pipeline Improve Mycobiome Data. PeerJ 2018, 6, e4925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and Clustering Orders of Magnitude Faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A Versatile Open Source Tool for Metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An Open Annotation Tool for Parsing Fungal Community Datasets by Ecological Guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Anderson, M.J. A New Method for Non-Parametric Multivariate Analysis of Variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Ellison, A.M. Princípios de Estatística Em Ecologia; Artmed Editora: Porto Alegre, Brazil, 2016. [Google Scholar]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Stevens, M.H.H.; Oksanen, M.J.; Suggests, M. The Vegan Package. Community Ecol. Package 2007, 10, 719. [Google Scholar]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and Extrapolation with Hill Numbers: A Framework for Sampling and Estimation in Species Diversity Studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Egidi, E.; Delgado-Baquerizo, M.; Plett, J.M.; Wang, J.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K. A Few Ascomycota Taxa Dominate Soil Fungal Communities Worldwide. Nat. Commun. 2019, 10, 2369. [Google Scholar] [CrossRef] [Green Version]
- Baldrian, P.; Kolařík, M.; Štursová, M.; Kopecký, J.; Valášková, V.; Větrovský, T.; Žifčáková, L.; Šnajdr, J.; Rídl, J.; Vlček, Č.; et al. Active and Total Microbial Communities in Forest Soil Are Largely Different and Highly Stratified during Decomposition. ISME J. 2012, 6, 248–258. [Google Scholar] [CrossRef] [Green Version]
- Pawlowska, T.E.; Taylor, J.W. Organization of Genetic Variation in Individuals of Arbuscular Mycorrhizal Fungi. Nature 2004, 427, 733–737. [Google Scholar] [CrossRef]
- Baldrian, P. Forest Microbiome: Diversity, Complexity and Dynamics. FEMS Microbiol. Rev. 2017, 41, 109–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prescott, C.E.; Grayston, S.J. Tree Species Influence on Microbial Communities in Litter and Soil: Current Knowledge and Research Needs. For. Ecol. Manag. 2013, 309, 19–27. [Google Scholar] [CrossRef]
- Wheeler, K.A.; Hurdman, B.F.; Pitt, J.I. Influence of PH on the Growth of Some Toxigenic Species of Aspergillus, Penicillium and Fusarium. Int. J. Food Microbiol. 1991, 12, 141–149. [Google Scholar] [CrossRef]
- Nevarez, L.; Vasseur, V.; Le Madec, A.; Le Bras, M.A.; Coroller, L.; Leguérinel, I.; Barbier, G. Physiological Traits of Penicillium Glabrum Strain LCP 08.5568, a Filamentous Fungus Isolated from Bottled Aromatised Mineral Water. Int. J. Food Microbiol. 2009, 130, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Lauber, C.L.; Strickland, M.S.; Bradford, M.A.; Fierer, N. The Influence of Soil Properties on the Structure of Bacterial and Fungal Communities across Land-Use Types. Soil Biol. Biochem. 2008, 40, 2407–2415. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil Bacterial and Fungal Communities across a PH Gradient in an Arable Soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Furley, P.A. The Nature and Diversity of Neotropical Savanna Vegetation with Particular Reference to the Brazilian Cerrados. Glob. Ecol. Biogeogr. 1999, 8, 223–241. [Google Scholar] [CrossRef]
- Silva, N.A.P.d.; Frizzas, M.R.; Oliveira, C.M.d. Seasonality in Insect Abundance in the “Cerrado” of Goiás State, Brazil. Rev. Bras. Entomol. 2011, 55, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Vellend, M. Conceptual Synthesis in Community Ecology. Q. Rev. Biol. 2010, 85, 183–206. [Google Scholar] [CrossRef] [Green Version]
- Huston, M. A General Hypothesis of Species Diversity. Am. Nat. 1979, 113, 81–101. [Google Scholar] [CrossRef]
- Ettema, C.H.; Wardle, D.A. Spatial Soil Ecology. Trends Ecol. Evol. 2002, 17, 177–183. [Google Scholar] [CrossRef]
- Saetre, P. Spatial Patterns of Ground Vegetation, Soil Microbial Biomass and Activity in a Mixed Spruce-Birch Stand. Ecography 1999, 22, 183–192. [Google Scholar] [CrossRef]
- Vukicevich, E.; Lowery, T.; Bowen, P.; Úrbez-Torres, J.R.; Hart, M. Cover Crops to Increase Soil Microbial Diversity and Mitigate Decline in Perennial Agriculture. A Review. Agron. Sustain. Dev. 2016, 36, 48. [Google Scholar] [CrossRef] [Green Version]
- Cloutier, M.L.; Murrell, E.; Barbercheck, M.; Kaye, J.; Finney, D.; García-González, I.; Bruns, M.A. Fungal Community Shifts in Soils with Varied Cover Crop Treatments and Edaphic Properties. Sci. Rep. 2020, 10, 6198. [Google Scholar] [CrossRef]
- Wakelin, S.A.; Chu, G.; Broos, K.; Clarke, K.R.; Liang, Y.; McLaughlin, M.J. Structural and Functional Response of Soil Microbiota to Addition of Plant Substrate Are Moderated by Soil Cu Levels. Biol. Fertil. Soils 2010, 46, 333–342. [Google Scholar] [CrossRef]
- Fernández-Calviño, D.; Soler-Rovira, P.; Polo, A.; Díaz-Raviña, M.; Arias-Estévez, M.; Plaza, C. Enzyme Activities in Vineyard Soils Long-Term Treated with Copper-Based Fungicides. Soil Biol. Biochem. 2010, 42, 2119–2127. [Google Scholar] [CrossRef]
- Nunes, I.; Jacquiod, S.; Brejnrod, A.; Holm, P.E.; Johansen, A.; Brandt, K.K.; Priemé, A.; Sørensen, S.J. Coping with Copper: Legacy Effect of Copper on Potential Activity of Soil Bacteria Following a Century of Exposure. FEMS Microbiol. Ecol. 2016, 92. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Kirkby, C.A.; Schmutter, D.; Bissett, A.; Kirkegaard, J.A.; Richardson, A.E. Network Analysis Reveals Functional Redundancy and Keystone Taxa amongst Bacterial and Fungal Communities during Organic Matter Decomposition in an Arable Soil. Soil Biol. Biochem. 2016, 97, 188–198. [Google Scholar] [CrossRef]
- Sharma, R.; Rajak, R.C. Keratinophilic Fungi: Nature’s Keratin Degrading Machines! Resonance 2003, 8, 28–40. [Google Scholar] [CrossRef]
- Nordén, J.; Penttilä, R.; Siitonen, J.; Tomppo, E.; Ovaskainen, O. Specialist Species of Wood-Inhabiting Fungi Struggle While Generalists Thrive in Fragmented Boreal Forests. J. Ecol. 2013, 101, 701–712. [Google Scholar] [CrossRef]
- York, M.; Rosenberg, D.; Sturm, K. Diet and Food-Niche Breadth of Burrowing Owls (Athene Cunicularia) in the Imperial Valley, California. West. North Am. Nat. 2002, 62, 280–287. [Google Scholar]
- Poulin, R.G.; Todd, L.D.; Dohms, K.M.; Brigham, R.M.; Wellicome, T.I. Factors Associated with Nest- and Roost-Burrow Selection by Burrowing Owls (Athene Cunicularia) on the Canadian Prairies. Can. J. Zool. 2005, 83, 1373–1380. [Google Scholar] [CrossRef] [Green Version]
- Ayarbe, J.P.; Kieft, T.L. Mammal Mounds Stimulate Microbial Activity in a Semiarid Shrubland. Ecology 2000, 81, 1150–1154. [Google Scholar] [CrossRef]
- Rinaldi, A.C.; Comandini, O.; Kuyper, T.W. Ectomycorrhizal Fungal Diversity: Seperating the Wheat from the Chaff. Fungal Divers. 2008, 33, 1–45. [Google Scholar]
- Ryberg, M.; Matheny, P.B. Asynchronous Origins of Ectomycorrhizal Clades of Agaricales. Proc. R. Soc. B Biol. Sci. 2012, 279, 2003–2011. [Google Scholar] [CrossRef] [Green Version]
- Guarro, J.; Summerbell, R.C.; Samson, R.A. Libero Ajello Medical Mycology Collection Onygenales: The Dermatophytes, Dimorphics, and Keratin Degraders in Their Evolutionary Context; Studies in Mycology; Centraalbureau voor Schimmelcultures: Utrecht, The Netherlands, 2002; ISBN 907035148X. [Google Scholar]
- Scott, J.A.; Untereiner, W.A. Determination of Keratin Degradation by Fungi Using Keratin Azure. Med. Mycol. 2004, 42, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Voříšková, J.; Baldrian, P. Fungal Community on Decomposing Leaf Litter Undergoes Rapid Successional Changes. ISME J. 2013, 7, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Mašínová, T.; Bahnmann, B.D.; Větrovský, T.; Tomšovský, M.; Merunková, K.; Baldrian, P. Drivers of Yeast Community Composition in the Litter and Soil of a Temperate Forest. FEMS Microbiol. Ecol. 2017, 93, fiw223. [Google Scholar] [CrossRef]
- Chagas-Neto, T.C.; Chaves, G.M.; Colombo, A.L. Update on the Genus Trichosporon. Mycopathologia 2008, 166, 121. [Google Scholar] [CrossRef] [PubMed]
- Colombo, A.L.; Padovan, A.C.B.; Chaves, G.M. Current Knowledge of Trichosporon Spp. and Trichosporonosis. Clin. Microbiol. Rev. 2011, 24, 682–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
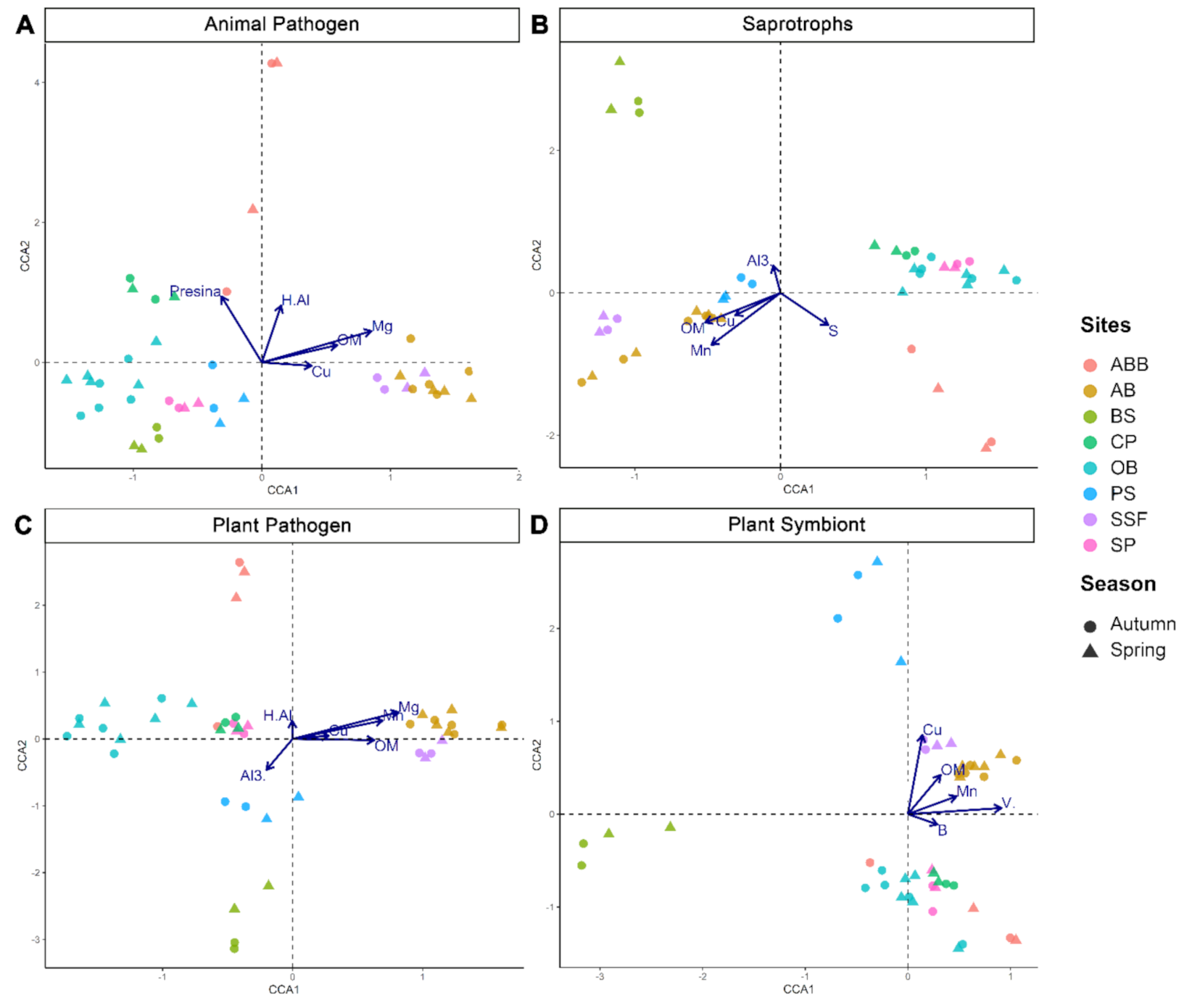{kind=link}
| Pairwise | ANOVA * and Kruskall–Wallis * | PERMANOVA * | ||||||
|---|---|---|---|---|---|---|---|---|
| Environment 1 | Environment 2 | H’ | S | Fungal Communities | Animal Pathogen | Plant Pathogen | Plant Symbiont | Saprotroph |
| Armadillo’s Burrow | Coffee Plantation | |||||||
| Armadillo’s Burrow | Abandoned Building | * | * | * | * | |||
| Armadillo’s Burrow | Brazilian Savana | * | * | |||||
| Armadillo’s Burrow | Owl’s Burrow | * | * | * | * | * | * | * |
| Armadillo’s Burrow | Pasture | * | * | |||||
| Armadillo’s Burrow | Seasonal Semi-deciduous Forest | * | ||||||
| Armadillo’s Burrow | Sugarcane Plantation | * | * | |||||
| Owl’s Burrow | Coffee Plantation | * | * | |||||
| Owl’s Burrow | Abandoned Building | |||||||
| Owl’s Burrow | Brazilian Savana | * | ||||||
| Owl’s Burrow | Pasture | * | ||||||
| Owl’s Burrow | Seasonal Semi-deciduous Forest | * | * | |||||
| Owl’s Burrow | Sugarcane Plantation | * | * | |||||
| Coffee Plantation | Abandoned Building | |||||||
| Coffee Plantation | Brazilian Savana | |||||||
| Coffee Plantation | Seasonal Semi-deciduous Forest | |||||||
| Coffee Plantation | Sugarcane Plantation | |||||||
| Coffee Plantation | Pasture | |||||||
| Abandoned Building | Brazilian Savana | |||||||
| Abandoned Building | Seasonal Semi-deciduous Forest | |||||||
| Abandoned Building | Sugarcane Plantation | |||||||
| Abandoned Building | Pasture | |||||||
| Brazilian Savana | Seasonal Semi-deciduous Forest | |||||||
| Brazilian Savana | Sugarcane Plantation | |||||||
| Brazilian Savana | Pasture | |||||||
| Seasonal Semideciduous Forest | Sugarcane Plantation | |||||||
| Seasonal Semideciduous Forest | Pasture | |||||||
| Sugarcane Plantation | Pasture | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamauchi, D.H.; Garcia Garces, H.; Teixeira, M.d.M.; Rodrigues, G.F.B.; Ullmann, L.S.; Garcia Garces, A.; Hebeler-Barbosa, F.; Bagagli, E. Soil Mycobiome Is Shaped by Vegetation and Microhabitats: A Regional-Scale Study in Southeastern Brazil. J. Fungi 2021, 7, 587. https://doi.org/10.3390/jof7080587
Yamauchi DH, Garcia Garces H, Teixeira MdM, Rodrigues GFB, Ullmann LS, Garcia Garces A, Hebeler-Barbosa F, Bagagli E. Soil Mycobiome Is Shaped by Vegetation and Microhabitats: A Regional-Scale Study in Southeastern Brazil. Journal of Fungi. 2021; 7(8):587. https://doi.org/10.3390/jof7080587
Chicago/Turabian StyleYamauchi, Danielle Hamae, Hans Garcia Garces, Marcus de Melo Teixeira, Gabriel Fellipe Barros Rodrigues, Leila Sabrina Ullmann, Adalberto Garcia Garces, Flavia Hebeler-Barbosa, and Eduardo Bagagli. 2021. "Soil Mycobiome Is Shaped by Vegetation and Microhabitats: A Regional-Scale Study in Southeastern Brazil" Journal of Fungi 7, no. 8: 587. https://doi.org/10.3390/jof7080587