
Synergistic Interaction between the Entomopathogenic Fungus Akanthomyces attenuatus (Zare & Gams) and the Botanical Insecticide Matrine against Megalurothrips usitatus (Bagrall)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Rearing of Akanthomyces attenuatus
2.2. Fungal Culture and Preparations of Conidial Suspension
2.3. Matrine
2.4. Influence of Matrine on Biological Characteristics of Akanthomyces attenuatus
2.5. Bioassay Studies
2.5.1. Bioassay 1: Efficacy of Akanthomyces attenuatus Isolate SCAUDCL-53 against Megalurothrips usitatus
2.5.2. Bioassay 2: Efficacy of Matrine against Megalurothrips usitatus
2.5.3. Bioassay 3: Efficacy of Single or Joint Treatments of Akanthomyces attenuatus and Matrine against Megalurothrips usitatus
2.6. Enzymatic Activity of Megalurothrips usitatus in Response to Single or Joint Treatments of Akanthomyces attenuatus and Matrine
2.6.1. Insect Treatment and Sample Preparation
2.6.2. Total Protein Assay
2.6.3. Enzyme Activity Assays
2.7. Data Analysis
3. Results
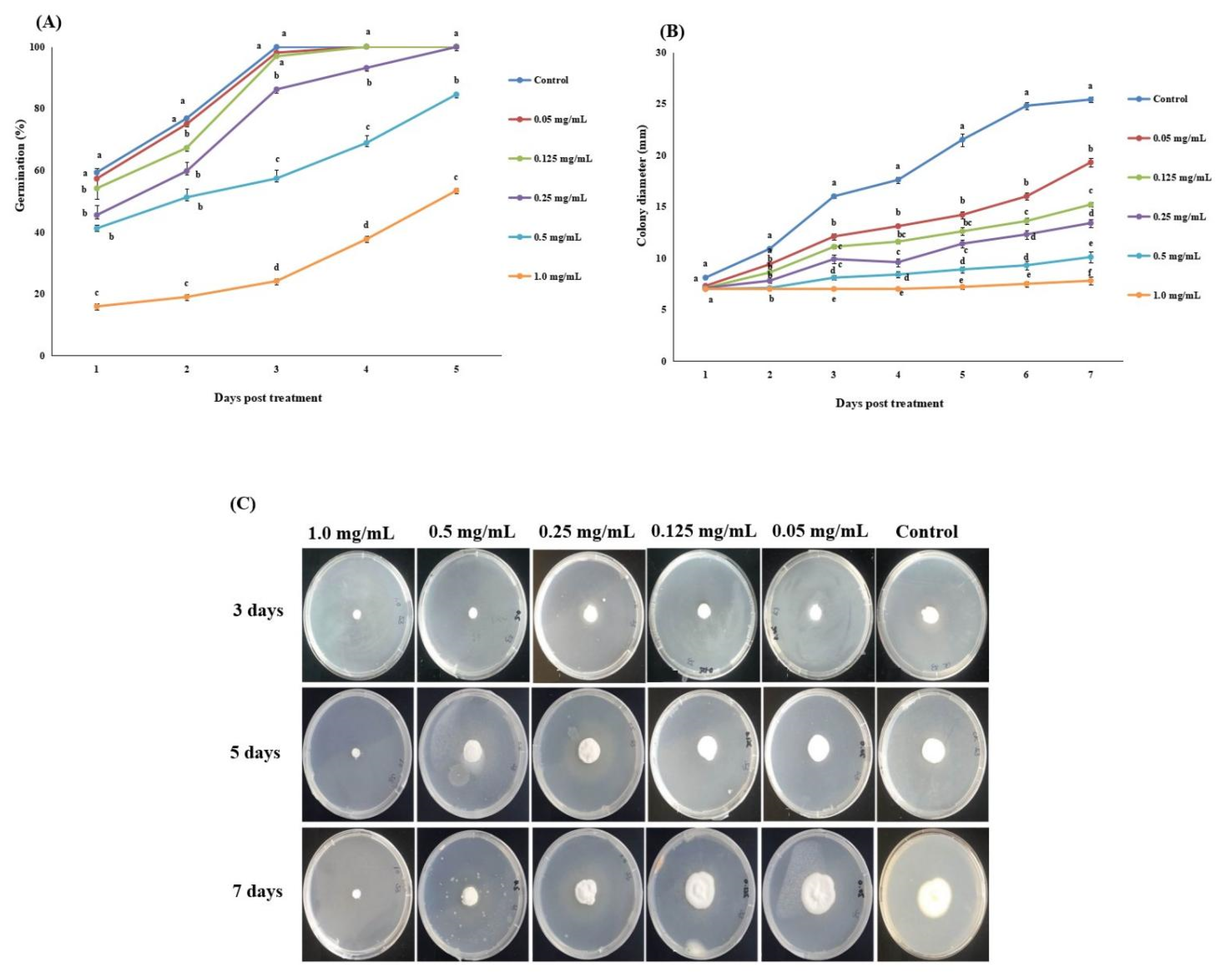
3.1. Influence of Matrine on Biological Characteristics of Akanthomyces attenuatus
3.2. Bioassay Studies
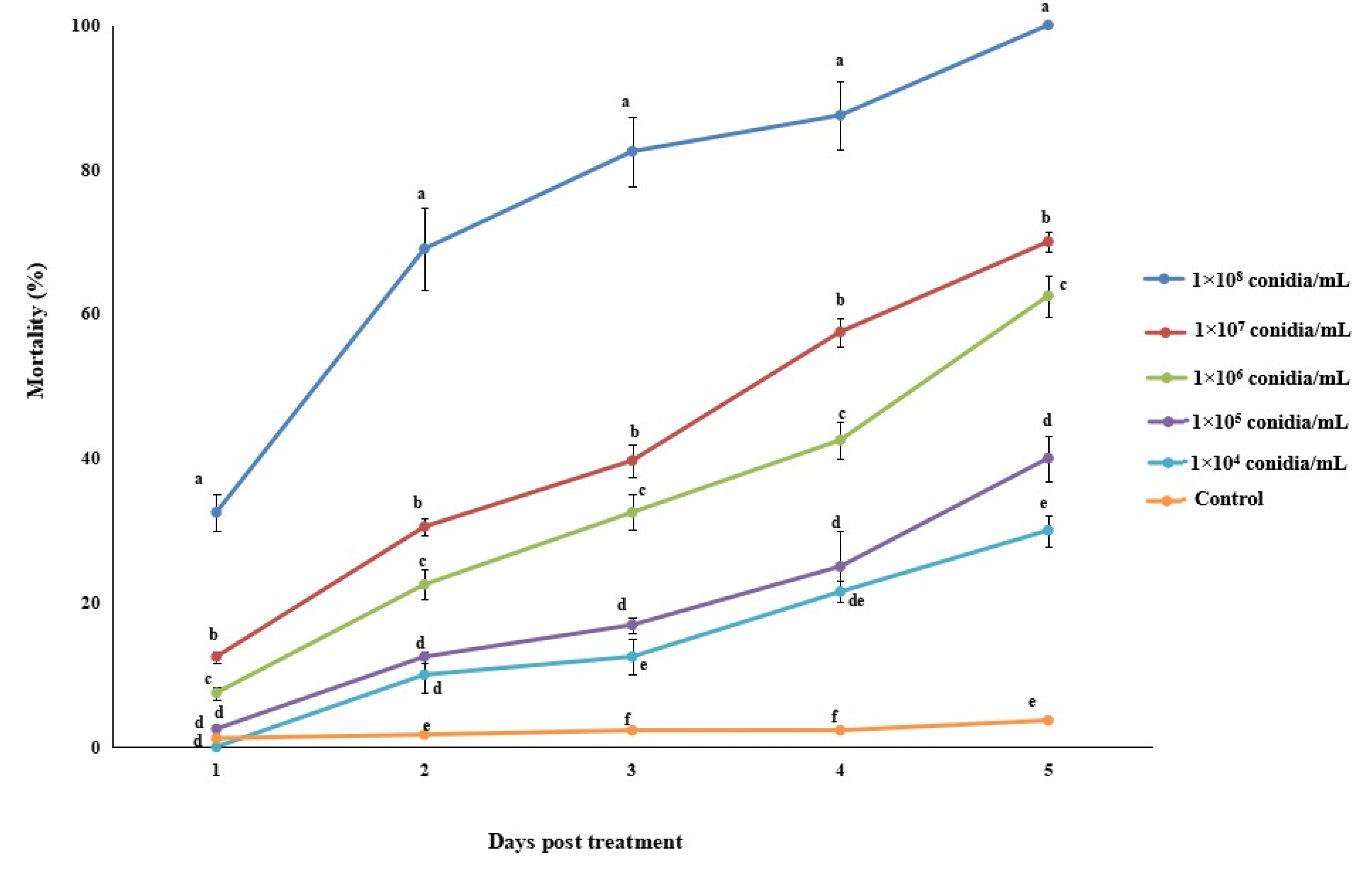
3.2.1. Bioassay 1: Efficacy of Akanthomyces attenuatus against Megalurothrips usitatus
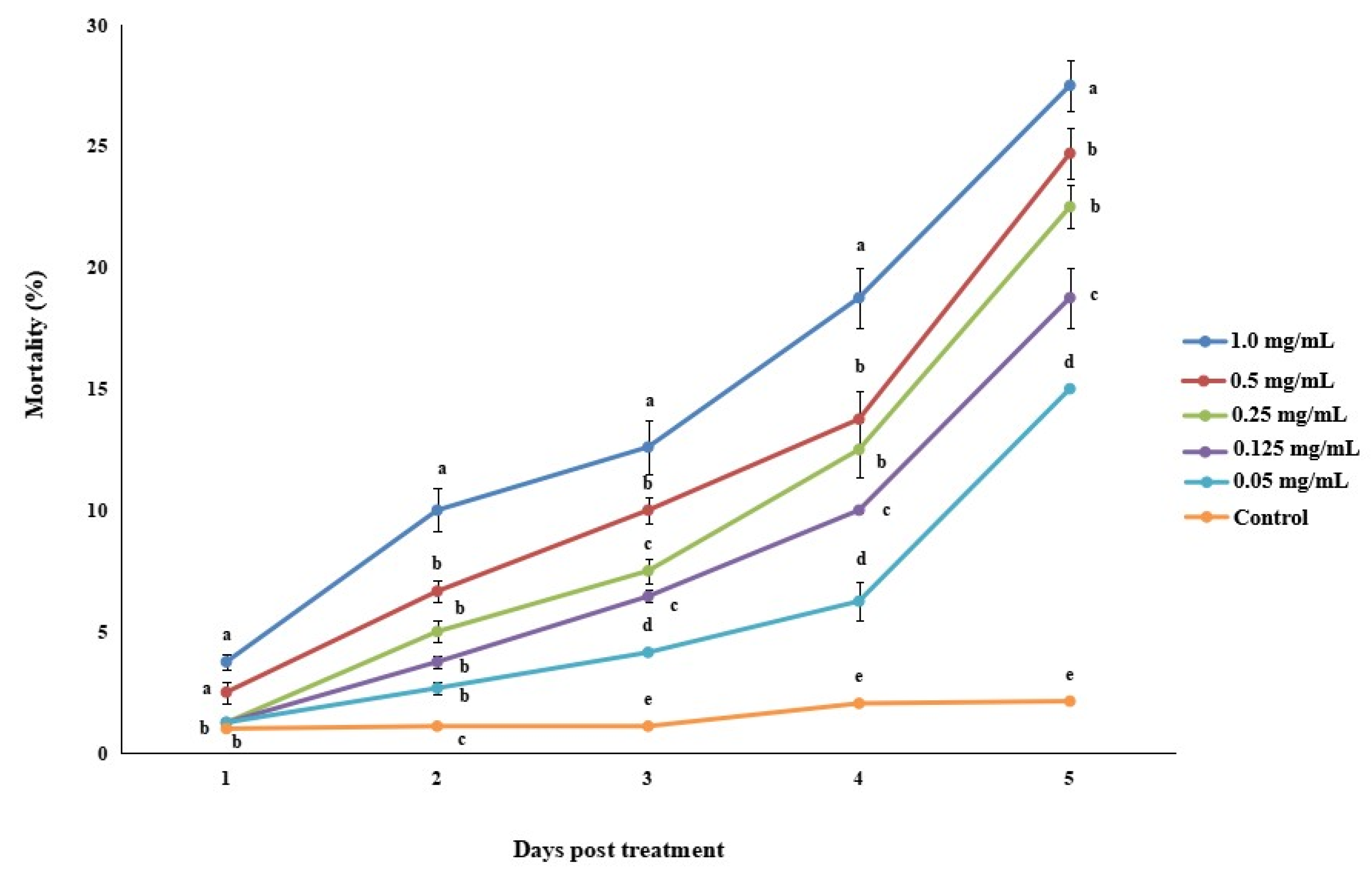
3.2.2. Bioassay 2: Efficacy of Matrine against Megalurothrips usitatus
3.2.3. Bioassay 3: Efficacy of Single or Joint Treatments of Akanthomyces attenuatus and Matrine against Megalurothrips usitatus
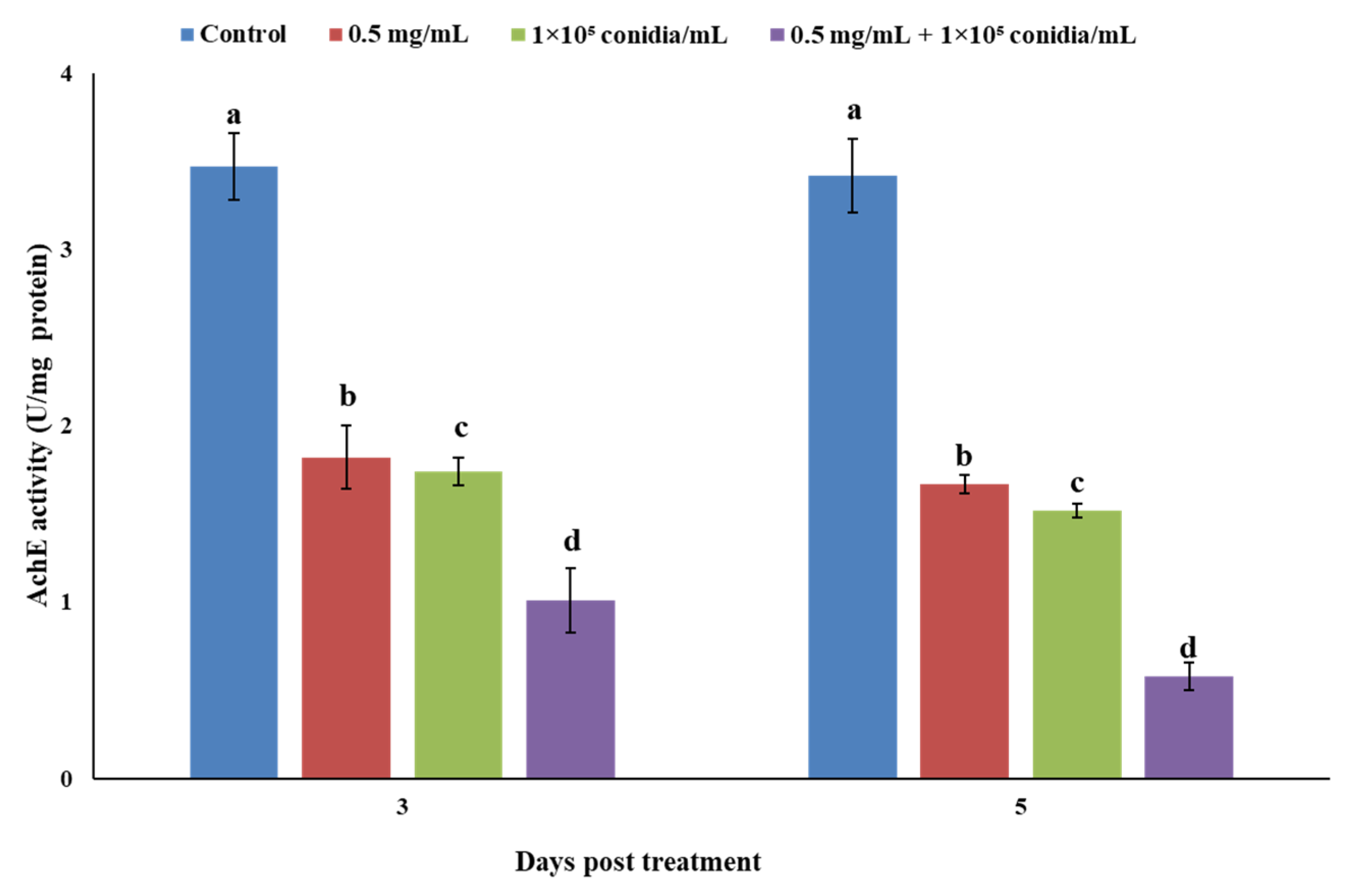
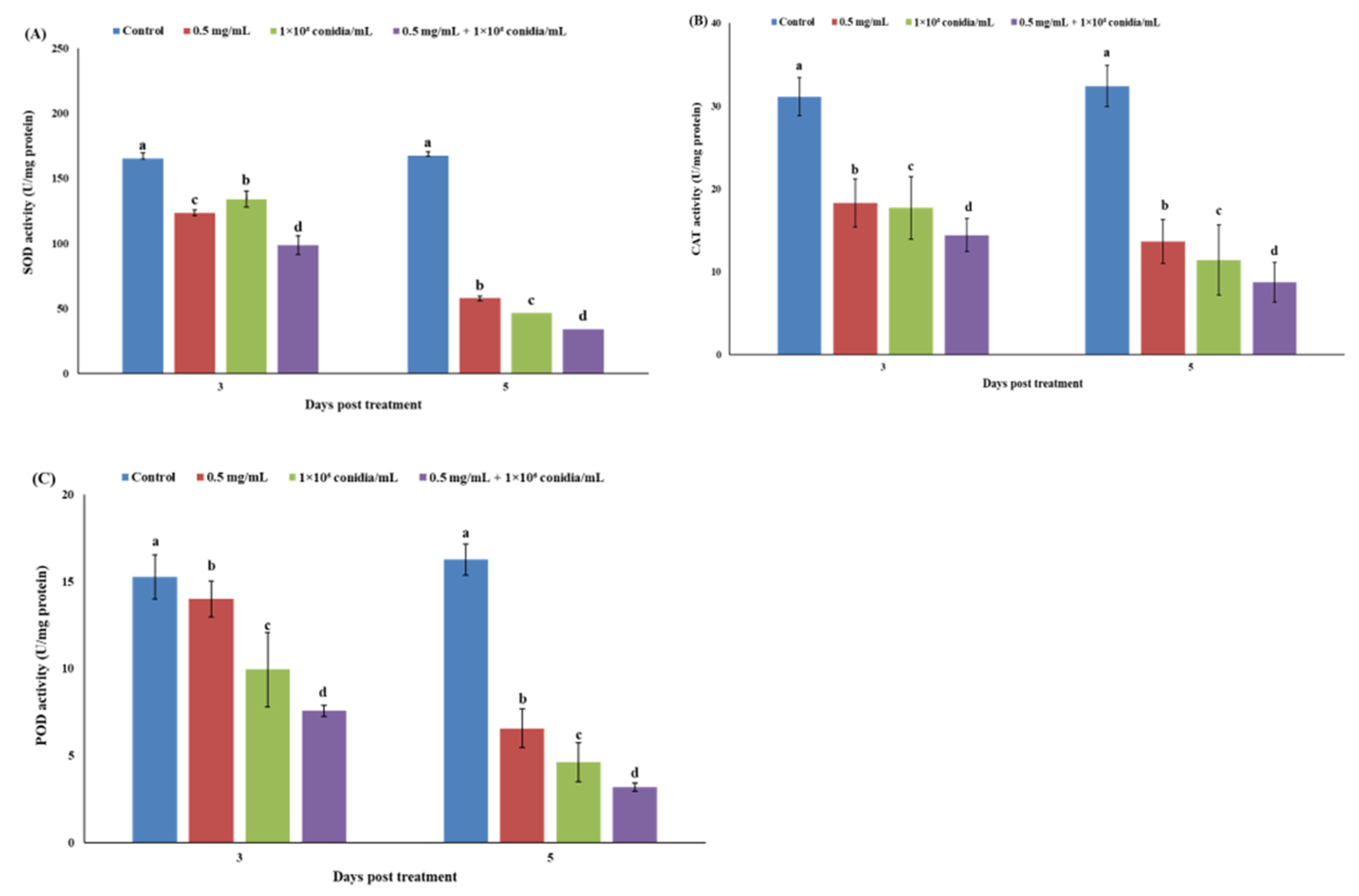
3.3. Enzymatic Response of Megalurothrips usitatus to Individual or Combined Treatments of Akanthomyces attenuatus and Matrine
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mound, L.A.; Walker, A.K. Thysanoptera as tropical tramps: New records from New Zealand and the Pacific. N. Z. Entomol. 1987, 9, 70–85. [Google Scholar] [CrossRef]
- Tang, L.D.; Yan, K.L.; Fu, B.L. The life table parameters of Megalurothrips usitatus (Thysanopera: Thripidea) on four leguminous crops. Fla. Entomol. 2015, 98, 620–625. [Google Scholar] [CrossRef]
- Yang, B.; Du, C.L.; Ali, S.; Wu, J.H. Molecular characterization and virulence of fungal isolates against the bean flower thrips, Megalurothrips usitatus Bagnall (Thysanopera: Thripidea). Egypt. J. Biol. Pest Cont. 2020, 30, 50. [Google Scholar] [CrossRef]
- Immaraju, J.A.; Paine, T.D.; Bethke, J.A.; Robb, K.L.; Newman, J.P. Western flower thrips (Thysanopera: Thripidea) resistance to insecticides in coastal California greenhouses. J. Econ. Entomol. 1992, 85, 9–14. [Google Scholar] [CrossRef]
- Skinner, M.; Gouli, S.; Frank, C.E.; Parker, B.L.; Kim, J.S. Management of Frankliniella occidentalis (Thysanopera: Thripidea) with granular formulations of entomopathogenic fungi. Biol. Cont. 2012, 63, 246–252. [Google Scholar] [CrossRef]
- Wright, S.P.; Filotas, M.J.; Sanderson, J.P. Comparative efficacy of emulsifiableoil, wettable-powder, and unformulated-powder preparations of Beauveria bassiana against the melon aphid Aphi gossypii. Biocon. Sci. Technol. 2016, 26, 894–914. [Google Scholar] [CrossRef]
- Du, C.; Yang, B.; Wu, J.H.; Ali, S. Identification and virulence characterization of two Akanthomyces attenuatus isolates against Megalurothrips usitatus (Thysanopera: Thripidea). Insects. 2019, 10, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, L.X.; Henderson, G. Antifeedant activity and acute and residual toxicity of alkaloids from Sophora flavescens (Leguminosae) against formosan subterranean termites (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2007, 100, 866–870. [Google Scholar] [CrossRef]
- Liu, Z.L.; Goh, S.H.; Ho, S.H. Screening of Chinese medicinal herbs for bioactivity against Sitophilus zeamais Motschulsky and Tribolium castaneum (Herbst). J. Stored Prod. Res. 2007, 43, 290–296. [Google Scholar] [CrossRef]
- Cheng, X.; Ye, J.; He, H.; Liu, Z.; Xu, C.; Wu, B. Synthesis, characterization and in vitro biological evaluation of two matrine derivatives. Sci. Rep. 2018, 8, 15686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakr, E.M.; Soliman, Z.R.; Hassan, M.F.; Tawadrous, S.S.D. Biological activity of the organic pesticide Baicao No.1 against the red spider mite Tetranychus urticae Koch. Acarines. 2012, 6, 35–39. [Google Scholar]
- Wu, J.H.; Yu, X.; Wang, X.S.; Tang, L.D.; Ali, S. Matrine enhances the pathogenicity of Beauveria brongniartii against Spodoptera litura (Lepidoptera: Noctuidae). Front. Microbiol. 2019, 10, 1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, I.C.; Kim, J.; Kim, H.M.; Kim, D.I.; Kim, S.G.; Kim, S.S.; Jang, C. Evaluation of toxicity of plant extract made by neem and matrine against main pests and natural enemies. Kor. J. Appl. Entomol. 2009, 48, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Zanardi, O.Z.; Ribeiro, L.D.P.; Ansante, T.F.; Santos, M.S.; Bordini, G.P.; Yamamotoa, P.T. Bioactivity of a matrine-based biopesticide against four pest species of agricultural importance. Crop Prot. 2015, 67, 160–167. [Google Scholar] [CrossRef]
- Li, S.G.; Hau, R.M.; Lin, H.F.; Cao, H.Q.; Hu, J.; Hu, P. Control effects of four biological pesticides and two chemical pesticides and their mixtures against mixed population of Nilaparvata lugens and Sogatella furcifera. Chin. Bull. Entomol. 2010, 47, 768–772. [Google Scholar]
- Dubovskiy, I.M.; Slyamova, N.D.; Kryukovi, N.D.; Yaroslavtseva, O.N.; Levchenko, M.V.; Belgibaeva, A.B.; Adilkhanzy, A.; Glupov, V.V. The activity of nonspecific esterases and glutathione-S-transferase in Locusta migratoria larvae infected with the fungus Metarhizium anisopliae (Ascomycota, Hypocreales). Entomol. Rev. 2012, 92, 27–31. [Google Scholar] [CrossRef]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Cao, G.C.; Li, Y.; Tu, X.B.; Wang, G.J.; Nong, X.Q.; Whitman, D.W.; Zhang, Z. Biochemical basis of synergism between pathogenic fungus Metarhizium anisopliae and insecticide chlorantraniliprole in Locusta migratoria (Meyen). Sci. Rep. 2016, 6, 28424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zibaee, A.; Bandani, A.R.; Tork, M. Effect of the entomopathogenic fungus, Beauveria bassiana, and its secondary metabolite on detoxifying enzyme activities and acetylcholinesterase (AChE) of the Sunn pest, Eurygaster integriceps (Heteroptera: Scutellaridae). Biocontrol Sci. Technol. 2009, 19, 485–498. [Google Scholar] [CrossRef]
- Xing, J.; Liang, P.; Gao, X.W. Effects of sublethal concentrations of chlorantraniliprole on insecticide susceptibility and detoxifying enzyme activity in Plutella xylostella. Chin. J. Pestic. Sci. 2011, 13, 464–470. [Google Scholar]
- Terriere, L.C. Induction of detoxication enzymes in insects. Annu. Rev. Entomol. 1984, 29, 71–88. [Google Scholar] [CrossRef]
- Ali, S.; Zhang, C.; Wang, Z.Q.; Wang, X.M.; Wu, J.H.; Cuthbertson, A.G.S.; Qiu, B.L. Toxicological and biochemical basis of synergism between the entomopathogenic fungus Lecanicillium muscarium and the insecticide matrine against Bemisia tabaci (Gennadius). Sci. Rep. 2017, 7, 46558. [Google Scholar] [CrossRef]
- Espinosa, P.J.; Contreras, J.; Quinto, V.; Grávalos, C.; Fernández, E.; Bielza, P. Metabolic mechanisms of insecticide resistance in the western flower thrips, Frankliniella occidentalis (Pergande). Pest Manag. Sci. 2005, 61, 1009–1015. [Google Scholar] [CrossRef] [PubMed]
- Lazreg, F.; Haung, Z.; Ali, S.; Ren, S.X. Effect of Lecanicillium muscarium on Eretmocerus sp. nr. furuhashii (Hymenoptera: Aphelinidae), a parasitoid of Bemisia tabaci (Hemiptera: Aleyrodidae). J. Pest Sci. 2009, 82, 27–32. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ellman, G.L.; Coutney, K.D.; Andre, V.; Featherstone, R.M. A new and rapid calorimetric determination of acetylcholinestrase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Beers, R.F.; Sizer, I.W. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar] [CrossRef]
- Shannon, L.M.; Kay, E.; Lew, J.Y. Peroxidase isozymes from horseradish roots. I. Isolation and physical properties. J. Biol. Chem. 1966, 241, 2166–2172. [Google Scholar] [CrossRef]
- Xu, D.; Ali, S.; Huang, Z. Insecticidal activity influence of 20-Hydroxyecdysone on the pathogenicity of Isaria fumosorosea against Plutella xylostella. Biol. Control. 2011, 56, 239–244. [Google Scholar] [CrossRef]
- Wu, J.; Li, J.; Zhang, C.; Yu, X.; Cuthbertson, A.G.S.; Ali, S. Biological impact and enzyme activities of Spodoptera litura (Lepidoptera: Noctuidae) in response to synergistic action of matrine and Beauveria brongniartii. Front. Physiol. 2020, 11, 584405. [Google Scholar] [CrossRef]
- Nakajyo, S.; Shimizu, K.; Kometani, A.; Suzuki, A.; Ozaki, H.; Urakawa, N. On the inhibitory mechanism of bassianolide, a cyclodepsipeptide, in acetylcholine-induced contraction in guinea pig Taenia coli. Jpn. J. Pharmacol. 1983, 33, 573–582. [Google Scholar] [CrossRef]
- Furlong, M.J.; Groden, E. Evaluation of synergistic interactions between the Colorado potato beetle (Coleoptera: Chrysomelidae) pathogen Beauveria bassiana and the insecticides, imidacloprid, and cyromazine. J. Econ. Entomol. 2001, 94, 344–356. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.Q.; Orozco, R.; Wijerante, E.M.K.; Espinosa-Artiles, P.; Lesile Gunatilaka, A.A.; Stock, S.P. Biosynthesis of the cyclooligomer depsipeptide bassianolide, an insecticidal virulence factor of Beauveria bassiana. Fungal Genet. Biol. 2009, 46, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Bourne, Y.; Sharpless, K.B.; Taylor, P.; Marchot, P. Steric and dynamic parameters influencing in situ cyclo additions to form triazole inhibitors with crystalline acetylcholinestrase. J. Amer. Chem. Soc. 2016, 138, 1611–1621. [Google Scholar] [CrossRef]
- Liu, L.J.; Alam, M.S.; Hirata, K.; Matsuda, K.; Ozoe, Y. Actions of quinolizidine alkaloids on Periplanta americana nicotinic acetylcholine receptors. Pest Manag. Sci. 2008, 64, 1222–1228. [Google Scholar] [CrossRef]
- Ding, S.Y.; Li, H.Y.; Li, X.F.; Zhang, Z.Y. Effects of two kinds of transgenic poplar on protective enzymes system in the midgut of larvae of American white moth. J. For. Res. Jpn. 2001, 12, 119–122. [Google Scholar]
- Felton, G.W.; Summers, C.B. Antioxidant systems in insects. Arch. Insect Biochem. 1995, 29, 187–197. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Megalurothrips usitatus Mortality (%) at Different Time Intervals | |||||
|---|---|---|---|---|---|---|
| 1d | 2d | 3d | 4d | 5d | ||
| Matrine (mg/mL) | 0.50 | 2.5 ± 0.36 d | 6.67 ± 0.67 g | 10.23 ± 0.45 g | 13.75 ± 1.24 i | 24.70 ± 1.89 h |
| 0.25 | 1.25 ± 0.14 e | 5.12 ± 0.23 g | 7.53 ± 0.35 h | 12.51 ± 1.21 jk | 22.51 ± 1.23 i | |
| Akanthomyces attenuatus (conidia/mL) | 1 × 107 | 12.54 ± 0.89 b | 30.36 ± 1.49 b | 39.67 ± 4.41 d | 57.48 ± 1.61 h | 70.03 ± 1.34 d |
| 1 × 106 | 7.51 ± 1.67 d | 22.48 ± 1.21 c | 32.51 ± 3.21 d | 42.47± 2.09 ij | 62.48 ± 1.37 ef | |
| 1 × 105 | 2.47 ± 0.17 d | 9.98 ± 0.23 fg | 12.56 ± 1.67 f | 25.07± 1.67 j | 40.12 ± 2.89 g | |
| Matrine (mg/mL) + Akanthomyces attenuatus (conidia/mL) | 0.50 + 1 × 107 | 17.23 * ± 2.33 a (9.59;17.19) | 36.67 * ± 1.67 a (25.49;21.00) | 51.34 * ± 3.14 a (35.56;3.80) | 72.33 * ± 3.12 b (53.63;5.54) | 89.67 * ± 2.88 a (75.09; 4.83) |
| 0.25 + 1 × 107 | 12.67 ± 1.67 b (8.16;0.02) | 30 * ± 2.89 b (16.00;12.24) | 46.03 * ± 2.12 b (26.39;14.56) | 67.67 * ± 6.11 bc (42.66;14.65) | 85.00 ± 2.89 ab (71.25;1.43) | |
| 0.50 + 1 × 106 | 8.33 ± 0.33 c (6.55;0.33) | 21.67 ± 3.33 c (14.44;1.27) | 39.67 * ± 2.12 c (36.39;2.41) | 69.67 *± 3.13 bc (50.37;7.38) | 86.67 *± 5.14 ab (56.01;16.77) | |
| 0.25 + 1 × 106 | 8.33 ± 1.67 c (6.55;0.48) | 11.67 ± 1.12 e (12.89;0.11) | 26.67 ± 1.67 e (21.98;1.01) | 58.33 *± 2.89 d (49.68;11.50) | 81.33 *± 3.12 b (70.93;25.96) | |
| 0.50 + 1 × 105 | 5 ± 1.33 cd (4.93;0.00) | 16.67 ± 3.33 d (15.94;0.03) | 32.33 *± 1.12 d (21.25;5.77) | 66.67 *± 2.89 c (35.27;27.94) | 78.33 *± 2.12 c (54.82;10.08) | |
| 0.25 + 1 × 105 | 3.33 ± 1.67 d (3.71;0.04) | 11.67 * ± 2.89 e (6.55;4.01) | 25.00 * ± 5.00 c (15.97;5.11) | 56.67 *± 4.33 d (34.37;14.46) | 66.67 *± 3.14 e (53.5;8.24) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.; Yang, B.; Zhang, X.; Cuthbertson, A.G.S.; Ali, S. Synergistic Interaction between the Entomopathogenic Fungus Akanthomyces attenuatus (Zare & Gams) and the Botanical Insecticide Matrine against Megalurothrips usitatus (Bagrall). J. Fungi 2021, 7, 536. https://doi.org/10.3390/jof7070536
Wu J, Yang B, Zhang X, Cuthbertson AGS, Ali S. Synergistic Interaction between the Entomopathogenic Fungus Akanthomyces attenuatus (Zare & Gams) and the Botanical Insecticide Matrine against Megalurothrips usitatus (Bagrall). Journal of Fungi. 2021; 7(7):536. https://doi.org/10.3390/jof7070536
Chicago/Turabian StyleWu, Jianhui, Bo Yang, Xiaochen Zhang, Andrew G. S. Cuthbertson, and Shaukat Ali. 2021. "Synergistic Interaction between the Entomopathogenic Fungus Akanthomyces attenuatus (Zare & Gams) and the Botanical Insecticide Matrine against Megalurothrips usitatus (Bagrall)" Journal of Fungi 7, no. 7: 536. https://doi.org/10.3390/jof7070536