
Reconstitution of Polyketide-Derived Meroterpenoid Biosynthetic Pathway in Aspergillus oryzae


1
Laboratory of Natural Products Chemistry, Graduate School of Pharmaceutical Sciences, The University of Tokyo, Bunkyo-ku, Tokyo 113-0033, Japan
2
Collaborative Research Institute for Innovative Microbiology, The University of Tokyo, Yayoi 1-1-1, Bunkyo-ku, Tokyo 113-8657, Japan
*
Authors to whom correspondence should be addressed.
J. Fungi 2021, 7(6), 486; https://doi.org/10.3390/jof7060486
Submission received: 31 May 2021
/
Revised: 12 June 2021
/
Accepted: 14 June 2021
/
Published: 16 June 2021
(This article belongs to the Special Issue Aspergillus oryzae and related Koji molds)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The heterologous gene expression system with Aspergillus oryzae as the host is an effective method to investigate fungal secondary metabolite biosynthetic pathways for reconstruction to produce un-natural molecules due to its high productivity and genetic tractability. In this review, we focus on biosynthetic studies of fungal polyketide-derived meroterpenoids, a group of bioactive natural products, by means of the A. oryzae heterologous expression system. The heterologous expression methods and the biosynthetic reactions are described in detail for future prospects to create un-natural molecules via biosynthetic re-design.
1. Introduction
Aspergillus oryzae is a fungus that has been utilized for over 2000 years in the Japanese fermentation industry to yield sake, miso, and soy sauce, and for industrial enzyme production. A. oryzae is an important heterologous expression host for biosynthetic genes from Aspergillus species that produce numerous secondary metabolites, such as mycotoxins, aflatoxin, medicinal compounds, lovastatin, and pigments such as emodin [1,2,3], as well as other secondary metabolite-producing fungi. For investigations on their biosynthesis, the heterologous expression system is a valuable tool, as it provides information about their reactions based on the products accumulated in the transformants. The A. oryzae genome includes some genes encoding secondary metabolite biosynthetic enzymes, such as the non-ribosomal peptide synthetase, which is responsible for the production of toxic compounds, such as cyclopiazonic acid. However, most of these genes, including cyclopiazonic acid synthetases, are dysfunctional or not expressed in the A. oryzae strains used in fermented food, and many have been recognized as safe strains that do not produce any toxic compounds [4]. In addition, A. oryzae possesses a strong metabolic flux to supply precursors of polyketides, terpenoids, and peptides, as judged by the high yields of heterologous expression products. Furthermore, genetic manipulation methods, such as gene deletion or heterologous expression, strong promoters (e.g., PamyB and PenoA) and multicopy vectors [5,6], and the genetically tractable mutated strains A. oryzae M-2-3 and NSAR1, have been established [7,8]. The long-standing efforts of academic and industrial investigators have made this strain an excellent chassis for the heterologous expression of secondary metabolite genes. By using this strain, we can easily access new biosynthetic pathways and construct systems for the production of beneficial compounds.
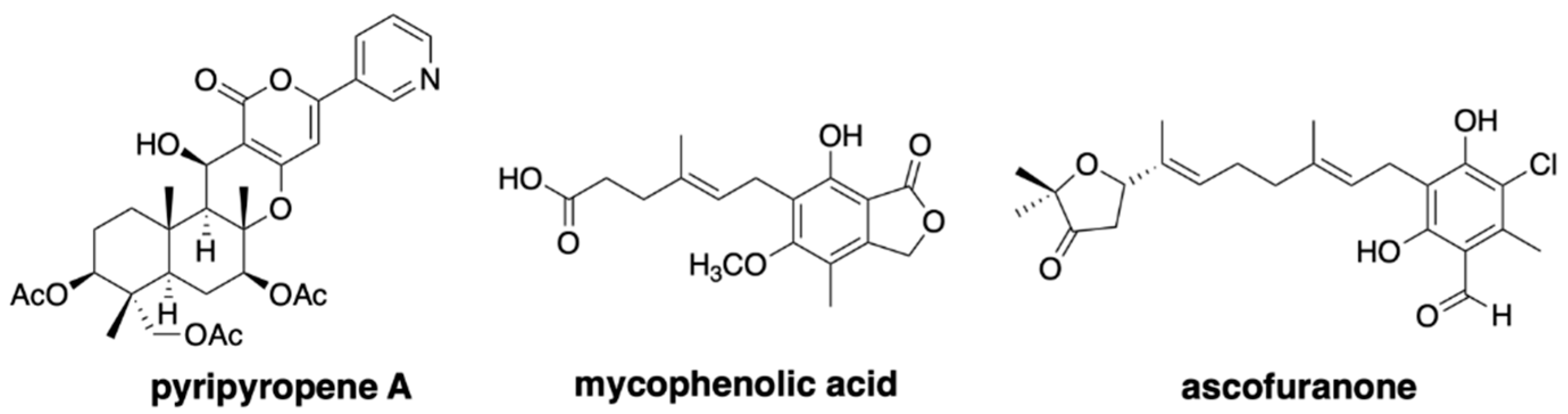
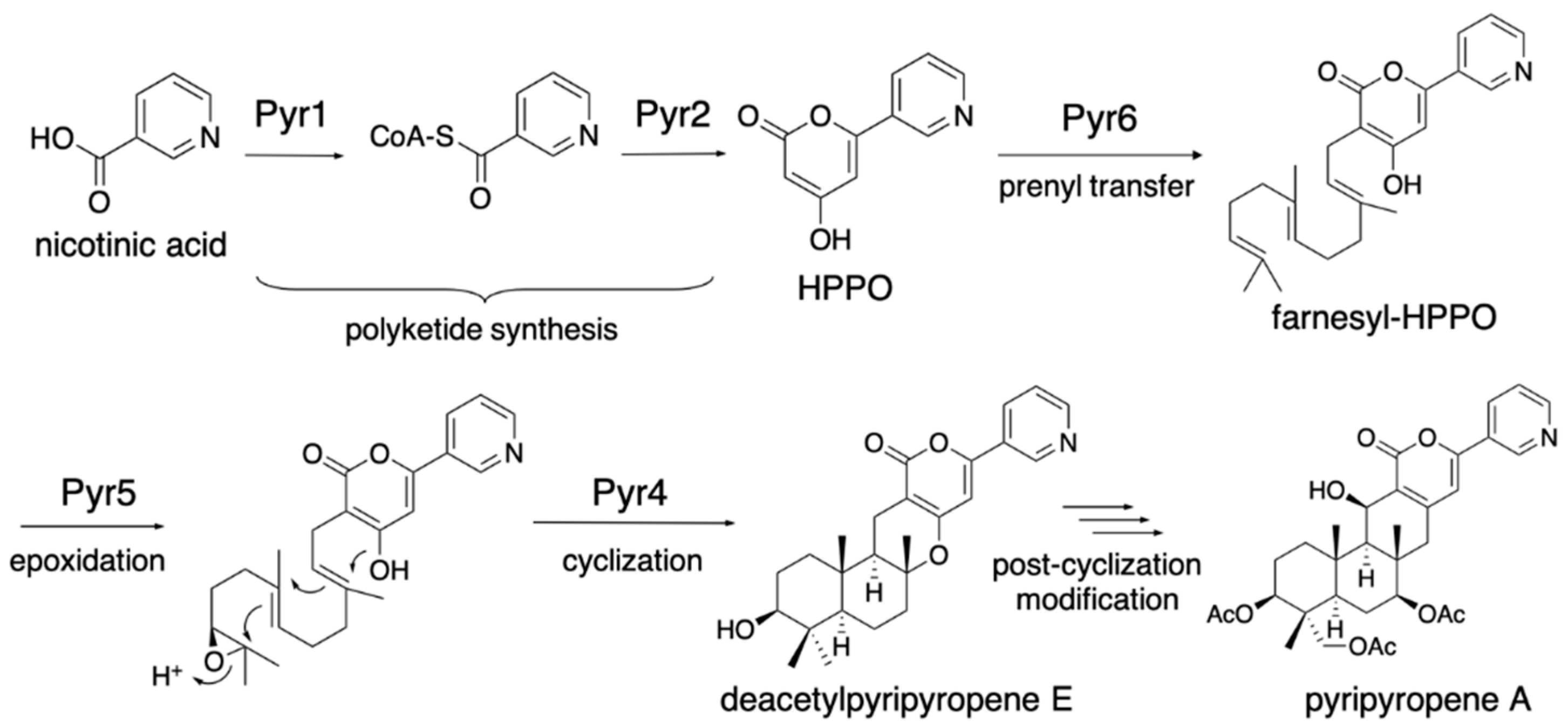
The fungal meroterpenoids are one of the groups of natural products that include various medicinally important compounds, such as the acyl-CoA:cholesterol acyltransferase inhibitor pyripyropene, the immunosuppressant mycophenolic acid, and the anti-trypanosomiasis ascofuranone (Figure 1) [9]. The biosynthesis of fungal meroterpenoids can be separated into five parts: polyketide synthesis, prenyl transfer, epoxidation, cyclization, and post-cyclization modification [10,11,12], as exemplified by pyripyropene A biosynthesis (Figure 2). By using the versatile A. oryzae host, which synthesizes polyketide and terpenoid compounds with high yields, we can create a fungal meroterpenoid production system. This method includes important benefits, in that we can exploit the novel biocatalysts by identifying the compounds accumulated in the heterologous expression system, and also re-engineer the biosynthetic pathway easily, by simply swapping one part of the biosynthetic machinery or feeding an un-natural substrate.
2. Pyripyropene A Biosynthetic Pathway
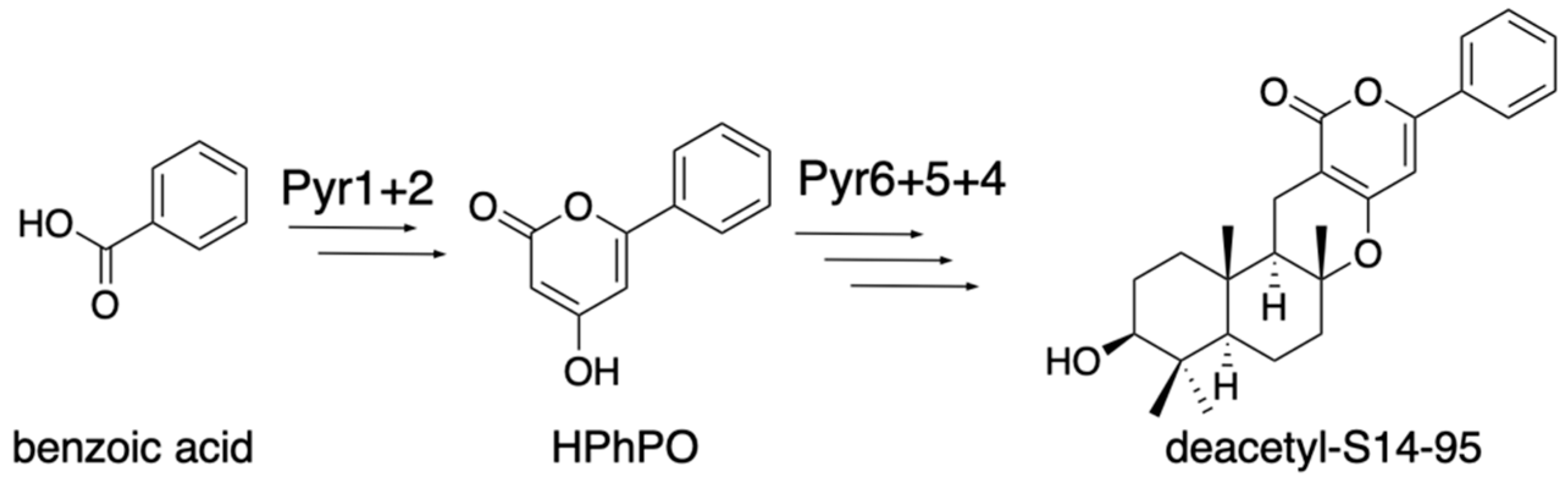
The earliest study of fungal meroterpenoid biosynthesis was the reconstitution of the early pyripyropene A biosynthetic pathway in the Aspergillus oryzae M-2-3 strain (ΔargB) [13]. The pyr2 (polyketide synthase, PKS) gene was cloned into the pTAex3 vector [14], and the pyr1 (CoA-ligase) gene was cloned into pPTRI [15], with the amyB promoter and terminator from pTAex3, to yield pTAex3-pyr2 and pPTRI-pyr1, respectively. Similarly, pyr5 (flavin-dependent monooxygenase, FMO) was cloned into pTAex3, and pyr4 (transmembrane meroterpenoid cyclase, CYC) and pyr6 (UbiA-type prenyltransferase, PT) were cloned into pPTRI with the amyB promoter and terminator, to yield pTAex3-pyr5 and pPTRI-pyr4+6, respectively. Two sets of vectors, pTAex3-pyr2 and pPTRI-pyr1 and pTAex3-pyr5 and pPTRI-pyr4+6, were introduced into Aspergillus oryzae M-2-3 [7]. The A. oryzae/pyr1+2 was fed nicotinic acid and produced 4-hydroxy-6-(3-pyridinyl)-2H-pyran-2-one (HPPO), while A. oryzae/pyr4+5+6 was fed HPPO and produced deacetylpyripyropene E. The structures of these products suggested that the following biosynthetic reactions, Pyr1 and Pyr2, produce HPPO from nicotinic acid, CoA, and malonyl-CoA; Pyr6 transfers the farnesyl group at C-3 to yield 3-farnesyl-HPPO; Pyr5 epoxidizes the terminal olefin of the farnesyl group; and Pyr4 facilitates the cyclization of the epoxidated prenyl chain via protonation to produce deacetylpyripyropene E (Figure 2). This was the first study that identified the function of the transmembrane meroterpenoid cyclase [16], and it paved the way toward investigations of the fungal meroterpenoid biosynthetic pathway. The A. oryzae/pyr1+2 strain was fed benzoic acid and generated 4-hydroxy-6-phenyl-2H-pyran-2-one (HpHpO), which was then fed to A. oryzae/pyr4+5+6 as a substrate to yield the un-natural meroterpenoid deacetyl-S14-95 (Figure 3) [13], illustrating the ease of mutasynthesis with this heterologous expression system.
3. 3,5-Dimethylorselinic Acid (DMOA)-Derived Meroterpenoid Biosynthetic Pathways
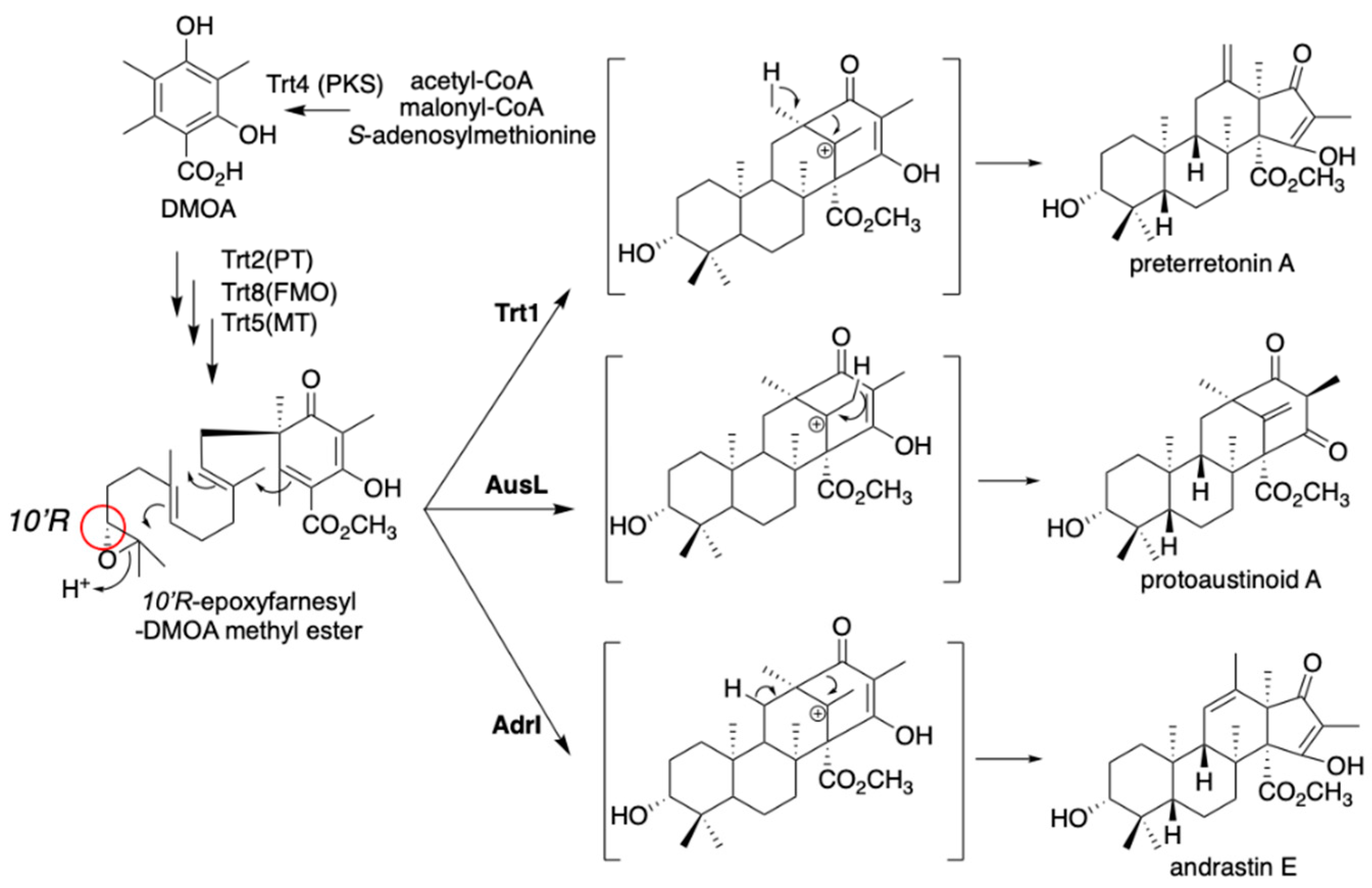
The fungal meroterpenoids derived from 3,5-dimethylorselinic acid (DMOA) exhibit structural diversity derived from various kinds of biosynthetic enzymes, including terpene cyclases and oxygenases [10,11,12]. In most of our expression studies, we employed A. oryzae NSAR1 [8] as the heterologous expression host. Since this strain possesses four auxotrophic mutations (ΔargB, adeA-, sC-, and niaD-), we can introduce four vectors harboring different auxotrophic markers, including argB (pTAex3) [14], adeA (pAdeA) [17], sC (pUSA) [18], and niaD (pUNA) [19]. By employing two vectors with the pyrithiamine resistance marker ptrA (pPTRI) [15] and the glufosinate resistance marker bar (pBAR) [20], we can introduce up to six vectors into a single expression host. Together, the strong amyB promoter on these vectors and the efficient metabolite efflux in the host facilitate the construction of a high-yield production system. By expressing the genes encoding Trt4 (PKS), Trt2 (PT), Trt8 (FMO), and Trt5 (methyltransferase) in A. oryzae NSAR1, we prepared a strain that produces 10’R-epoxyfarnesyl-3,5-DMOA methyl ester (Figure 4) [21,22]. With the additionally expressed trt1 (CYC) in the A. oryzae system, the strain produced preterretonin A (Figure 4). When we swapped trt1 with ausL, a homolog of trt1 classified in the same clade of the phylogenetic tree, the trt4285+ausL transformant produced protoaustinoid A, a compound with a differently cyclized terpene ring (Figure 4). Furthermore, replacing trt1 with adrI in the transformant also led to the production of another meroterpenoid, andrastin E [23]. These studies illustrate the usefulness of the heterologous gene expression system for metabolic engineering, in that we can create analogs by simply swapping one enzyme at a step in the biosynthetic reactions. A similar concept was also employed in the fungal indole diterpene combinatorial biosynthesis [24].
Recently, we produced novel meroterpenoids by introducing meroterpenoid cyclases from four different biosynthetic pathways into the A. oryzae host producing 10’R-epoxyfarnesyl-3,5-DMOA methyl ester, and obtained two novel mono-cyclized meroterpenoids [25]. It is intriguing that the four different meroterpenoid cyclases produced the same products from the non-native substrate.
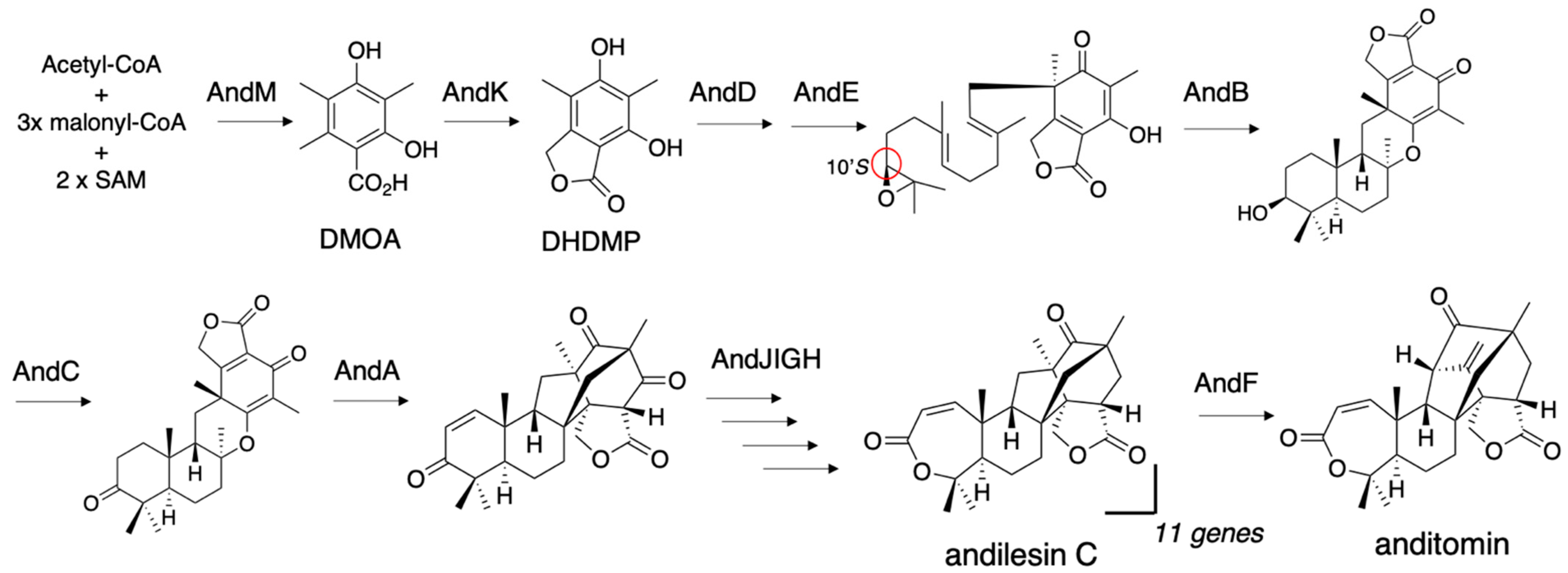
Anditomin is also a DMOA-derived meroterpenoid with the characteristic bicyclo[2.2.2]octane structure (Figure 5). In the anditomin biosynthetic study, we succeeded in the heterologous expression of 11 genes, andM(PKS)+K(P450 monooxygenase+hydrolase)+D(PT)+E(FMO)+B(CYC)+C(alcohol dehydrogenase)+A(α-ketoglutarate-dependent oxygenase)+J(Baeyer-Villiger monooxygenase)+I(ketoreductase)+G(acetyltransferase)+H(ene-reductase). To manage the number of vectors, two expression cassettes of PamyB-ORF-TamyB were connected with a linear vector by three-fragment in-fusion recombination [20]. First, we introduced pTAex3-andM, pUSA-andK+D, and pAdeA-andE+B into A. oryzae NSAR1, and then transformed the resultant strain with pUNA-andC+A and pPTRI-andJ+I to construct the nine-gene expression strain. Finally, pBARI-andG+H was transformed into the nine-gene transformant to yield the 11-gene expression strain, which produced andilesin C (Figure 5). pBARI was constructed by cloning the bar gene expression cassette with the gpd promoter from pBARGPE1 (FGSC) into pUC19. The transformants were selected with glufosinate, purified from a commercial herbicide [26]. To the best of our knowledge, the introduction of 11 genes into a single host is the largest number for A. oryzae heterologous expression. As we failed to introduce a 12th gene into the transformants, 11 genes might be the upper limit. The reaction of AndF was characterized through in vitro reaction by using andilesin C as a substrate. The characteristics of this biosynthesis are the generation of a 10S-epoxide, while the terretonin biosynthetic pathway generates a 10R-epoxide as a substrate for terpene cyclase, and the extensive terpene skeleton reconstruction by the α-ketoglutarate-dependent oxygenases AndA and AndF. The details of the catalysis by AndA have been extensively studied by a combination of X-ray crystal structural analysis and DFT calculations [27,28]. The generation of the 10S-epoxide in the Aspergillus expression system was also reported in the synthesis of the unique orthoester-bearing DMOA meroterpenoid novofumigatonin [29].
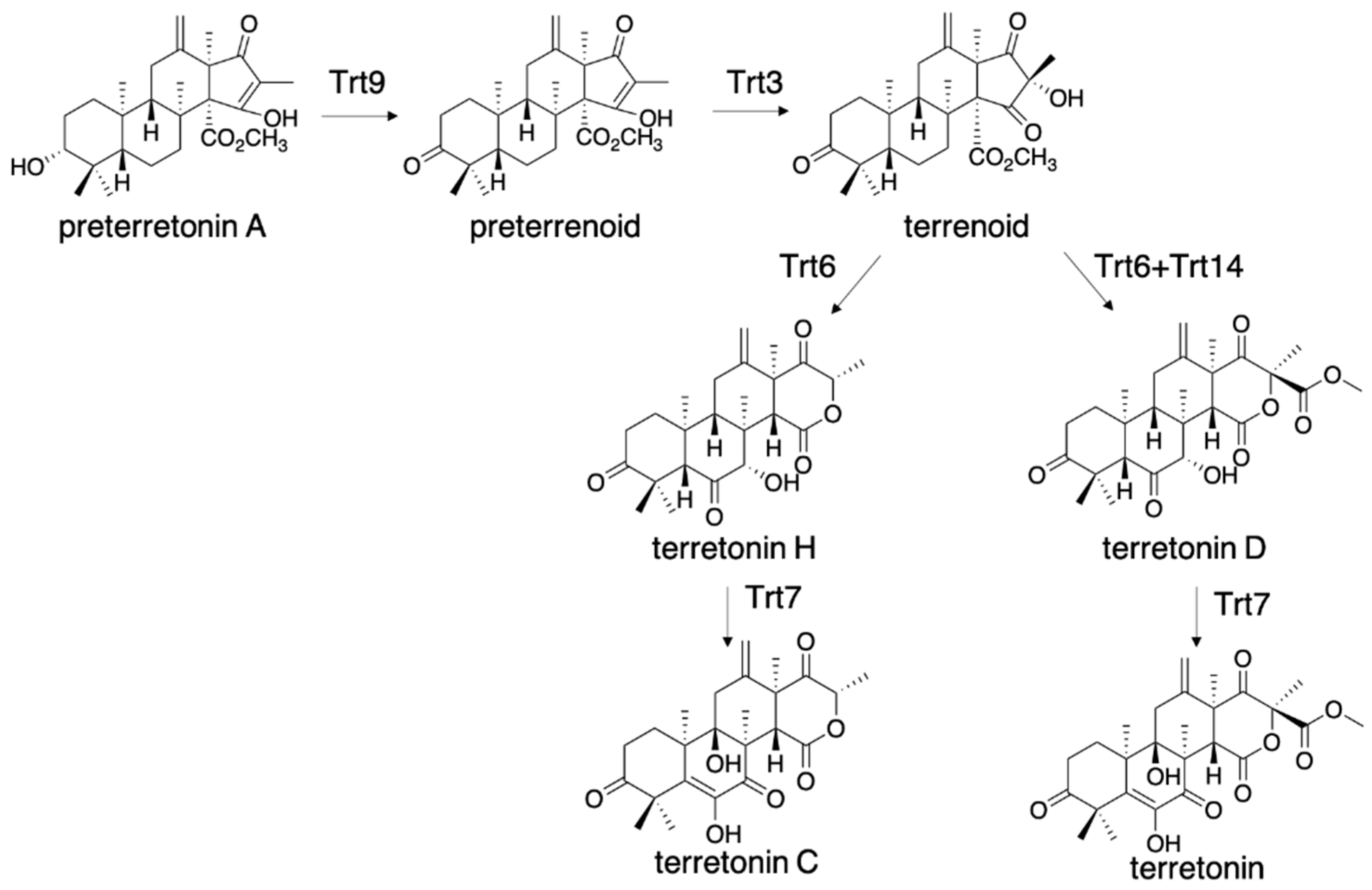
The D-ring of terretonin is highly oxygenated, and its biosynthesis is expected to include new biosynthetic enzymes. The expression of trt42851+trt9 (alcohol dehydrogenase) +trt3 (flavin monooxygenase) resulted in the production of terrenoid, which was further modified to terretonin H, with the additional expression of trt6 (P450 monooxygenase) (Figure 6) [30]. The membrane fraction expressing Trt6 was also prepared from A. oryzae/trt6 and used for the in vitro assay, indicating that Aspergillus gene expression system can also contribute to the preparation of membrane-bound enzymes that are only expressed in the eukaryotic host. Trt6 catalyzed C-7 hydroxylation, and the generated OH-7α formed a hydrogen bond with the carbonyl oxygen of the methyl ester. This hydrogen bond induces lactonization via the nucleophilic attack by OH-16 [30]. The resultant intermediate underwent retro-Claisen cleavage of the β-ketoester to the ring-expanded β-keto acid, and the following decarboxylation generated terretonin H. The additional expression of Trt14 (isomerase) in A. oryzae/trt42851936 yielded terretonin D, which bears a methyl ester group, and facilitated the D-ring forming reaction. The molecular basis of Trt14 catalysis was reported in 2017, according to its crystal structure complexed with intermediates [31].
4. Ascochlorin/Ascofuranone Biosynthetic Pathways
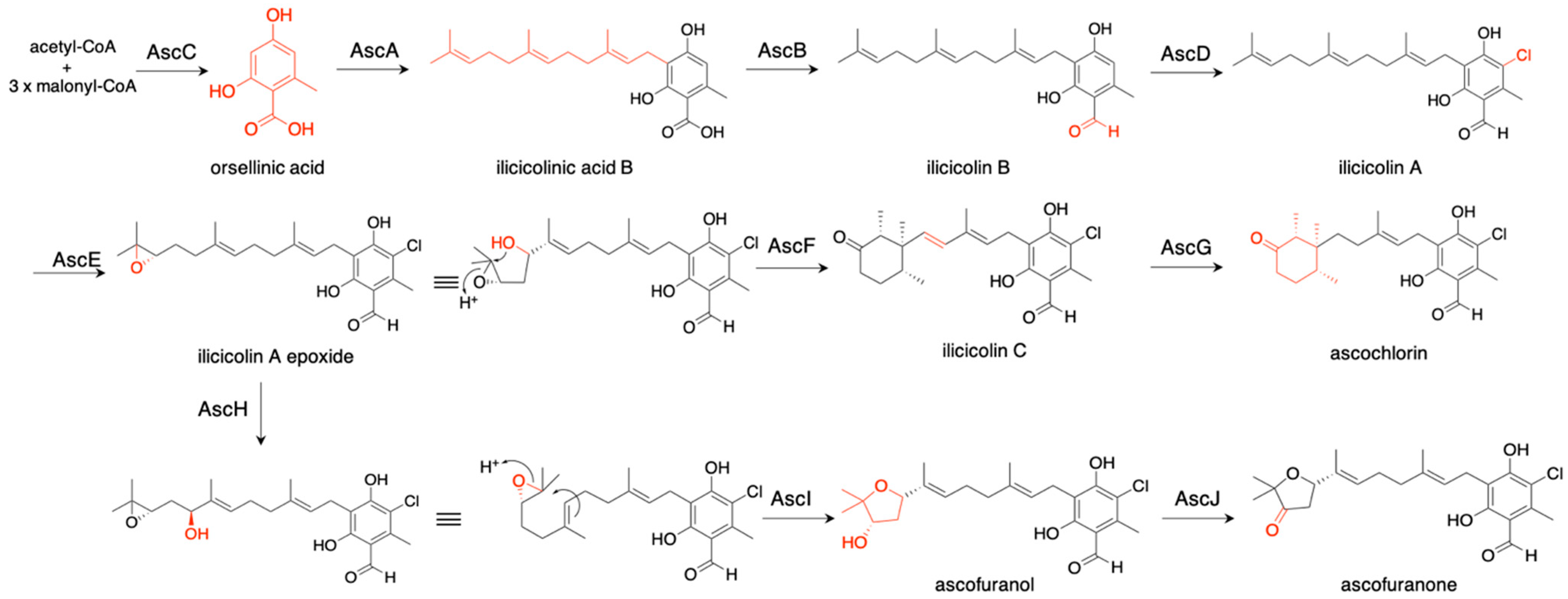
Acremonium egyptiacum produces two meroteropenoids, ascofuranone and ascochlorin, which are derived from the same intermediate, illicicolin A epoxide [32] (Figure 7). To investigate their biosynthetic pathways, we expressed ascC (polyketide synthase), ascA (prenyltransferase), and ascB (nonribosomal peptide synthetase-like reductase) in A. oryzae NSAR1, and the transformants produced illicicolin B with a yield of 0.71 mg/L. When we additionally expressed ascD (flavin-dependent halogenase), the transformant yielded a small amount of ilicicolin A (0.04 mg/L). The lower yield might be caused by the toxicity of the product against Aspergillus. Aspergillus soja was also employed as a heterologous expression host, and the proteins purified from this expression system were utilized for in vitro assays. The information from these experiments clarified the biosynthetic pathways of ascochlorin and ascofuranone, as shown in Figure 7. In ascochlorin biosynthesis, illicicolin A epoxide was protonated and cyclized into a (14S, 15R, 19R)-trimethylcyclohexanone ring structure by AscF to yield ilicicolin C, which was then oxidized by AscG (P450 oxidase) to produce ascochlorin. In ascofuranone biosynthesis, C-16 of illicicolin A epoxide was hydroxylated by AscH (P-450 oxygenase), and the product was cyclized into ascofuranol through 6-endo-tet cyclization by AscI. The ascofuranol was then oxidized by AscJ (NAD(P)-dependent dehydratase) to produce ascofuranone. AscI is a novel meroterpenoid cyclase sharing little similarity with known meroterpenoid cyclases, while AscF has similarity with Trt1 and known meroterpenoid cyclases [16,32,33]. The gene deletion of ascF resulted in the higher production of ascofuranone (501 mg/L) as compared with the wild-type strain (388 mg/L), illustrating the effectiveness of bioengineering to create a high-yield metabolite production system based on biosynthetic knowledge.
5. Diterpene Pyrone Biosynthetic Pathway
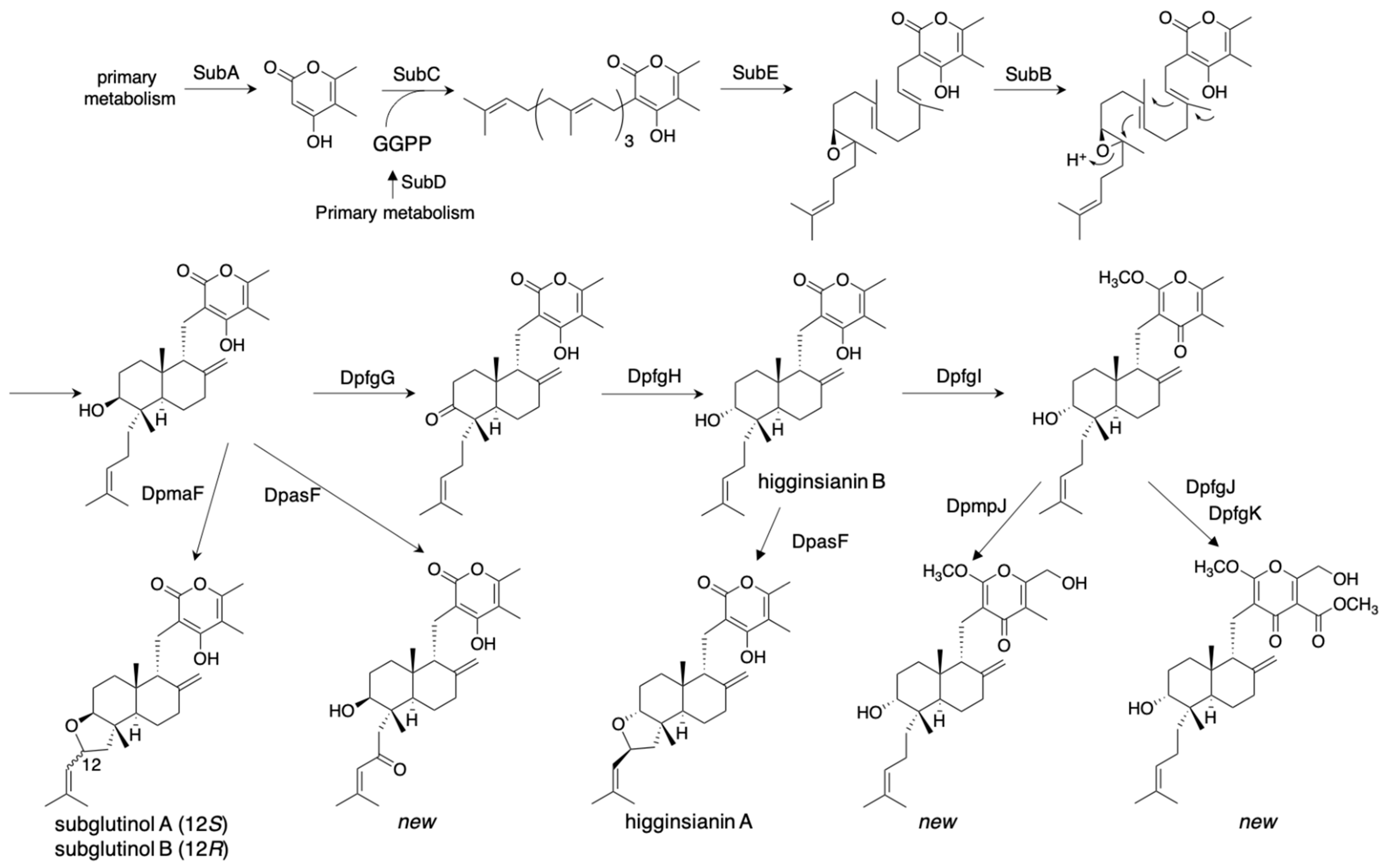
The most successful and thorough study on the combinatorial biosynthesis of fungal meroterpenoids was performed by Tsukada et al. [34]. They established ten biosynthetic pathways to yield 22 diterpene pyrones, including the immunosuppressive fungal meroterpenoid subglutinol A from Fusarium graminearum [35], and 15 novel analogs. They employed different combinations of the biosynthetic genes from five fungi isolated from a spider: F. graminearum, Macrophomina phaseolina, Colletotrichum higginsianum, Metarhizium anisopliae, and Arthrinium sacchari, in the A. oryzae NSAR1 heterologous expression system. The genes encoding SubA/DpasA (PKS), SubD/DpasD (geranylgeranylphosphate synthase), SubC/DpasC (PT), SubE/DpasE (FMO), and SubB/DpasB (CYC) from these five clusters produced the common diterpene pyrone intermediate with a high yield (87 mg/L). This intermediate structure was modified by the additional expression of DpmaF/DpasF/DpchF (flavin-dependent berberine bridge-like enzyme), to produce subglutinols A (89 mg/L) and B (6 mg/L) with their new analogs (Figure 8). Furthermore, they expressed DpfgG/DpmpG/DpchG and DpfgH/DpmpH/DpchH (NAD(P)-dependent reductases/dehydratases) to produce the 8-OH epimerized higginsianin B. DpfgI/DpmpI and DpfgK (methyltransferases), and DpmpJ and DpfgJ (P450 oxygenases) were employed to further diverge the pathway, which yielded a total of 22 diterpene pyrones (Figure 8). The well-designed cloning strategies enabled the authors to clone three or four gene expression cassettes into a single vector by two-step in-fusion cloning, and thus quickly construct diverse pathways. They reported a broad range of bioactivity tests, including anti-tumor, inhibition of insect innate immune system, anti-HIV, and anti-Alzheimer activities. Interestingly, the novel compounds produced via the un-natural pathways possess different bioactivities from the natural compounds. This study established the basis of synthetic biology to supply pharmacologically active compounds by heterologous expression systems.
6. Biosynthetic Pathways of Plant Meroterpenoids in A. oryzae
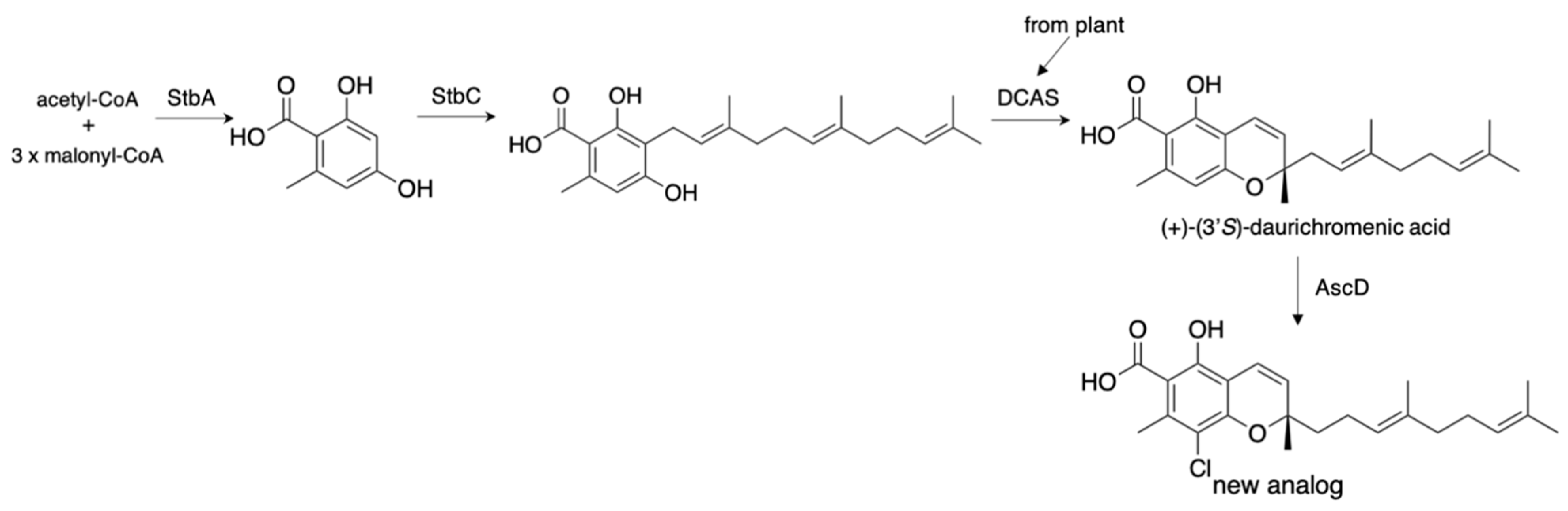
In addition to the fungal meroterpenoids, plant meroterpenoids can be produced in A. oryzae by expressing the plant enzyme with fungal meroterpenoid biosynthetic enzymes. In 2017, we successfully constructed the plant anti-HIV meroterpenoid daurichromenic acid (1.23 mg/L) by expressing DCAS (flavin-dependent oxidase) from the plant, Rhododendron dauricum, with StbA (PKS) and StbC (PT) from Stachybotrys bisbyi (Figure 9) [36,37]. Furthermore, the additional expression of AscD from the Fusarium sp. [33] yielded its un-natural halogenated analogs (2.06 mg/L). These results illustrate the potential of the Aspergillus expression system to produce secondary metabolites. The recent successful syntheses of basidiomycete and plant terpenes in A. oryzae further encourage the production of secondary metabolites from other organisms besides filamentous fungi [38,39].
7. Concluding Remarks
Aspergillus oryzae is a powerful heterologous expression host for the production of polyketides and terpenoids. The high flux of its primary metabolism and the ease of genetic manipulation provide opportunities to produce various meroterpenoids. This powerful microbial platform will contribute to the generation of pharmaceutically important, un-natural molecules with relatively easy genetic and metabolic engineering. The recent development of a genetic manipulation system based on CRISPR-Cas9 in A. oryzae will further expand this versatile expression system, to produce high yields of diverse bioactive molecules, through engineering its primary metabolism and constructing multi-gene expression systems [40,41].
Author Contributions
Conceptualization, writing—review, editing, critical revising, and funding acquisition, T.A. and I.A. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by a Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology, Japan (JSPS KAKENHI grant number JP16H06443, JP17H04763, JP19H04641, JP20H00490, JP20KK0173, and JP21H02636), the New Energy and Industrial Technology Development Organization (NEDO, Grant Number JPNP20011), UTEC-UTokyo FSI Research Grant Program, Kato Memorial Bioscience Foundation, and The Asahi Glass Foundation.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sanchez, J.F.; Somoza, A.D.; Keller, N.P.; Wang, C.C.C. Advances in Aspergillus secondary metabolite research in the post-genomic era. Nat. Prod. Rep. 2012, 29, 351–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lubertozzi, D.; Keasling, J.D. Developing Aspergillus as a host for heterologous expression. Biotechnol. Adv. 2009, 27, 53–75. [Google Scholar] [CrossRef] [PubMed]
- Pahirulzaman, K.A.K.; Williams, K.; Lazarus, C.M. A toolkit for heterologous expression of metabolic pathways in aspergillus oryzae. Methods Enzymol. 2012, 517, 241–260. [Google Scholar] [CrossRef]
- Barbesgaard, P.; Heldt-Hansen, H.P.; Diderichsen, B. On the safety of Aspergillus oryzae: A review. Appl. Microbiol. Biotechnol. 1992, 36, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Tada, S.; Gomi, K.; Kitamoto, K.; Kumagai, C.; Tamura, G.; Hara, S. Identification of the promoter region of the Taka-amylase A gene required for starch induction. Agric. Biol. Chem. 1991, 55, 1939–1941. [Google Scholar] [CrossRef]
- Tsuboi, H.; Koda, A.; Toda, T.; Minetoki, T.; Hirotsune, M.; Machida, M. Improvement of the Aspergillus oryzae enolase promoter (P-enoA) by the introduction of cis-element repeats. Biosci. Biotechnol. Biochem. 2005, 69, 206–208. [Google Scholar] [CrossRef] [Green Version]
- Gomi, K.; Iimura, Y.; Hara, S. Integrative Transformation of Aspergillus oryzae with a Plasmid Containing the Aspergillus nidulans argB gene. Agric. Biol. Chem. 1987, 51, 2549–2555. [Google Scholar] [CrossRef]
- Jin, F.J.; Maruyama, J.I.; Juvvadi, P.R.; Arioka, M.; Kitamoto, K. Development of a novel quadruple auxotrophic host transformation system by argB gene disruption using adeA gene and exploiting adenine auxotrophy in Aspergillus oryzae. FEMS Microbiol. Lett. 2004, 239, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Geris, R.; Simpson, T.J. Meroterpenoids produced by fungi. Nat. Prod. Rep. 2009, 26, 1063–1094. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, Y.; Abe, I. Biosynthesis of fungal meroterpenoids. Nat. Prod. Rep. 2016, 33, 26–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, Y.; Awakawa, T.; Mori, T.; Abe, I. Unusual chemistries in fungal meroterpenoid biosynthesis. Curr. Opin. Chem. Biol. 2016, 31, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Abe, I. Enzymology and biosynthesis of the orsellinic acid derived medicinal meroterpenoids. Curr. Opin. Biotechnol. 2021, 69, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Tokunaga, K.; Matsuda, Y.; Fujii, I.; Abe, I.; Ebizuka, Y.; Kushiro, T. Reconstitution of a fungal meroterpenoid biosynthesis reveals the involvement of a novel family of terpene cyclases. Nat. Chem. 2010, 2, 858–864. [Google Scholar] [CrossRef]
- Fujii, T.; Yamaoka, H.; Gomi, K.; Kitamoto, K.; Kumagai, C. Cloning and nucleotide sequence of the ribonuclease T1 gene (rntA) from Aspergillus oryzae and its expression in Saccharomyces cerevisiae and Aspergillus oryzae. Biosci. Biotechnol. Biochem. 1995, 59, 1869–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubodera, T.; Yamashita, N.; Nishimura, A. Pyrithiamine resistance gene (ptrA) of Aspergillus oryzae: Cloning, characterization and application as a dominant selectable marker for transformation. Biosci. Biotechnol. Biochem. 2000, 64, 1416–1421. [Google Scholar] [CrossRef] [PubMed]
- Barra, L.; Abe, I. Chemistry of fungal meroterpenoid cyclases. Nat. Prod. Rep. 2021, 38, 566–585. [Google Scholar] [CrossRef]
- Jin, F.J.; Maruyama, J.I.; Juvvadi, P.R.; Arioka, M.; Kitamoto, K. Adenine auxotrophic mutants of Aspergillus oryzae: Development of a novel transformation system with triple auxotrophic hosts. Biosci. Biotechnol. Biochem. 2004, 68, 656–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, O.; Nan, S.N.; Akao, T.; Tominaga, M.; Watanabe, H.; Satoh, T.; Enei, H.; Akita, O. dffA gene from Aspergillus oryzae encodes L-ornithine N5-oxygenase and is indispensable for deferriferrichrysin biosynthesis. J. Biosci. Bioeng. 2003, 95, 82–88. [Google Scholar] [CrossRef]
- Yamada, O.; Lee, B.R.; Gomi, K.; Iimura, Y. Cloning and functional analysis of the Aspergillus oryzae conidiation regulator gene brlA by its disruption and misscheduled expression. J. Biosci. Bioeng. 1999, 87, 424–429. [Google Scholar] [CrossRef]
- Matsuda, Y.; Wakimoto, T.; Mori, T.; Awakawa, T.; Abe, I. Complete biosynthetic pathway of anditomin: Nature’s sophisticated synthetic route to a complex fungal meroterpenoid. J. Am. Chem. Soc. 2014, 136, 15326–15336. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Tokunaga, K.; Radhakrishnan, E.K.; Fujii, I.; Abe, I.; Ebizuka, Y.; Kushiro, T. Identification of a key prenyltransferase involved in biosynthesis of the most abundant fungal meroterpenoids derived from 3,5-dimethylorsellinic acid. ChemBioChem 2012, 13, 1132–1135. [Google Scholar] [CrossRef]
- Matsuda, Y.; Awakawa, T.; Itoh, T.; Wakimoto, T.; Kushiro, T.; Fujii, I.; Ebizuka, Y.; Abe, I. Terretonin biosynthesis requires methylation as essential step for cyclization. ChemBioChem 2012, 13, 1738–1741. [Google Scholar] [CrossRef]
- Matsuda, Y.; Awakawa, T.; Abe, I. Reconstituted biosynthesis of fungal meroterpenoid andrastin A. Tetrahedron 2013, 69, 8199–8204. [Google Scholar] [CrossRef]
- Tang, M.C.; Lin, H.C.; Li, D.; Zou, Y.; Li, J.; Xu, W.; Cacho, R.A.; Hillenmeyer, M.E.; Garg, N.K.; Tang, Y. Discovery of unclustered fungal indole diterpene biosynthetic pathways through combinatorial pathway reassembly in engineered yeast. J. Am. Chem. Soc. 2015, 137, 13724–13727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Li, X.; Mori, T.; Awakawa, T.; Abe, I. Novel cyclohexyl meroterpenes produced by combinatorial biosynthesis. Chem. Pharm. Bull. 2021, 69, 444–446. [Google Scholar] [CrossRef] [PubMed]
- Chooi, Y.H.; Cacho, R.; Tang, Y. Identification of the viridicatumtoxin and griseofulvin gene clusters from Penicillium aethiopicum. Chem. Biol. 2010, 17, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, Y.; Mitsuhashi, T.; Matsuda, Y.; Senda, M.; Sato, H.; Yamazaki, M.; Uchiyama, M.; Senda, T.; Abe, I. Structural and computational bases for dramatic skeletal rearrangement in anditomin biosynthesis. J. Am. Chem. Soc. 2018, 140, 9743–9750. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Matsuda, Y.; Abe, I. Unique chemistry of non-heme iron enzymes in fungal biosynthetic pathways. Nat. Prod. Rep. 2018, 35, 633–645. [Google Scholar] [CrossRef]
- Matsuda, Y.; Bai, T.; Phippen, C.B.W.; Nødvig, C.S.; Kjærbølling, I.; Vesth, T.C.; Andersen, M.R.; Mortensen, U.H.; Gotfredsen, C.H.; Abe, I.; et al. Novofumigatonin biosynthesis involves a non-heme iron-dependent endoperoxide isomerase for orthoester formation. Nat. Commun. 2018, 9, 2587. [Google Scholar] [CrossRef]
- Matsuda, Y.; Iwabuchi, T.; Wakimoto, T.; Awakawa, T.; Abe, I. Uncovering the unusual D-ring construction in terretonin biosynthesis by collaboration of a multifunctional cytochrome P450 and a unique isomerase. J. Am. Chem. Soc. 2015, 137, 3393–3401. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; Iwabuchi, T.; Hoshino, S.; Wang, H.; Matsuda, Y.; Abe, I. Molecular basis for the unusual ring reconstruction in fungal meroterpenoid biogenesis. Nat. Chem. Biol. 2017, 13, 1066–1073. [Google Scholar] [CrossRef]
- Araki, Y.; Awakawa, T.; Matsuzaki, M.; Cho, R.; Matsuda, Y.; Hoshino, S.; Shinohara, Y.; Yamamoto, M.; Kido, Y.; Inaoka, D.K.; et al. Complete biosynthetic pathways of ascofuranone and ascochlorin in Acremonium egyptiacum. Proc. Natl. Acad. Sci. USA 2019, 116, 8269–8274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, Z.; Awakawa, T.; Wang, D.; Hu, Y.; Abe, I. Multidomain P450 epoxidase and a terpene cyclase from the ascochlorin biosynthetic pathway in Fusarium sp. Org. Lett. 2019, 21, 2330–2334. [Google Scholar] [CrossRef]
- Tsukada, K.; Shinki, S.; Kaneko, A.; Murakami, K.; Irie, K.; Murai, M.; Miyoshi, H.; Dan, S.; Kawaji, K.; Hayashi, H.; et al. Synthetic biology based construction of biological activity-related library of fungal decalin-containing diterpenoid pyrones. Nat. Commun. 2020, 11, 1830. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.C.; Lobkovsky, E.; Pliam, N.B.; Strobel, G.; Clardy, J. Subglutinols a and b: Immunosuppressive Compounds from the Endophytic Fungus Fusarium subglutinans. J. Org. Chem. 1995, 60, 7076–7077. [Google Scholar] [CrossRef]
- Li, C.; Matsuda, Y.; Gao, H.; Hu, D.; Yao, X.S.; Abe, I. Biosynthesis of LL-Z1272β: Discovery of a new member of NRPS-like enzymes for aryl-aldehyde formation. ChemBioChem 2016, 17, 904–907. [Google Scholar] [CrossRef]
- Okada, M.; Saito, K.; Wong, C.P.; Li, C.; Wang, D.; Iijima, M.; Taura, F.; Kurosaki, F.; Awakawa, T.; Abe, I. Combinatorial biosynthesis of (+)-daurichromenic acid and its halogenated analogue. Org. Lett. 2017, 19, 3183–3186. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Minami, A.; Ozaki, T.; Wu, J.; Kawagishi, H.; Maruyama, J.I.; Oikawa, H. Efficient reconstitution of basidiomycota diterpene erinacine gene cluster in ascomycota host Aspergillus oryzae based on genomic DNA sequences. J. Am. Chem. Soc. 2019, 141, 15519–15523. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Liu, J.; Du, Y.; Pei, X.; Li, M. Aspergillus oryzae biosynthetic platform for de novo iridoid production. J. Agric. Food Chem. 2021, 69, 2501–2511. [Google Scholar] [CrossRef] [PubMed]
- Katayama, T.; Tanaka, Y.; Okabe, T.; Nakamura, H.; Fujii, W.; Kitamoto, K.; Maruyama, J. Development of a genome editing technique using the CRISPR/Cas9 system in the industrial filamentous fungus Aspergillus oryzae. Biotechnol. Lett. 2016, 38, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Katayama, T.; Nakamura, H.; Zhang, Y.; Pascal, A.; Fujii, W.; Maruyama, J. Forced recycling of an AMA1-based genome-editing plasmid allows for efficient multiple gene deletion/integration in the industrial filamentous fungus Aspergillus oryzae. Appl. Environ. Microbiol. 2019, 85, e01896-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Representative bioactive fungal meroterpenoids.

Figure 2.
Biosynthetic pathway of pyripyropene A.

Figure 3.
Un-natural product synthesis from benzoic acid with Pyr12456.

Figure 4.
Diversification of DMOA-derived meroterpenoids with Trt1, AusL, and AdrI. C-10’R is highlighted with a red circle.
Figure 4.
Diversification of DMOA-derived meroterpenoids with Trt1, AusL, and AdrI. C-10’R is highlighted with a red circle.

Figure 5.
The 11-step anditomin biosynthetic pathway reconstructed in the A. oryzae heterologous expression system and the AndF reaction. C-10’S is highlighted with a red circle.
Figure 5.
The 11-step anditomin biosynthetic pathway reconstructed in the A. oryzae heterologous expression system and the AndF reaction. C-10’S is highlighted with a red circle.

Figure 6.
Biosynthesis of terretonin H and terretonin D from preterretonin A.

Figure 7.
Biosynthetic pathways of ascochlorin and ascofuranone. The red fonts indicate the structural change after each biosynthetic reaction.
Figure 7.
Biosynthetic pathways of ascochlorin and ascofuranone. The red fonts indicate the structural change after each biosynthetic reaction.

Figure 8.
Combinatorial biosynthesis of diterpenoid pyrones in A. oryzae.

Figure 9.
Production of plant meroterpenoids in A. oryzae.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Awakawa, T.; Abe, I. Reconstitution of Polyketide-Derived Meroterpenoid Biosynthetic Pathway in Aspergillus oryzae. J. Fungi 2021, 7, 486. https://doi.org/10.3390/jof7060486
AMA Style
Awakawa T, Abe I. Reconstitution of Polyketide-Derived Meroterpenoid Biosynthetic Pathway in Aspergillus oryzae. Journal of Fungi. 2021; 7(6):486. https://doi.org/10.3390/jof7060486
Chicago/Turabian StyleAwakawa, Takayoshi, and Ikuro Abe. 2021. "Reconstitution of Polyketide-Derived Meroterpenoid Biosynthetic Pathway in Aspergillus oryzae" Journal of Fungi 7, no. 6: 486. https://doi.org/10.3390/jof7060486
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.