
Isolation and Molecular Characterization of the Romaine Lettuce Phylloplane Mycobiome
by
 ,
,
Danny Haelewaters
1,* ,
,
Hector Urbina
1,2,
Samuel Brown
1,
Shannon Newerth-Henson
1 and
M. Catherine Aime
1,* 1
Department of Botany and Plant Pathology, Purdue University, West Lafayette, IN 47907, USA
2
Division of Plant Industry, Florida Department of Agriculture and Consumer Services, Gainesville, FL 32608, USA
*
Authors to whom correspondence should be addressed.
J. Fungi 2021, 7(4), 277; https://doi.org/10.3390/jof7040277
Submission received: 9 March 2021
/
Revised: 27 March 2021
/
Accepted: 5 April 2021
/
Published: 7 April 2021
(This article belongs to the Section Fungal Evolution, Biodiversity and Systematics)
Abstract
:Romaine lettuce (Lactuca sativa) is an important staple of American agriculture. Unlike many vegetables, romaine lettuce is typically consumed raw. Phylloplane microbes occur naturally on plant leaves; consumption of uncooked leaves includes consumption of phylloplane microbes. Despite this fact, the microbes that naturally occur on produce such as romaine lettuce are for the most part uncharacterized. In this study, we conducted culture-based studies of the fungal romaine lettuce phylloplane community from organic and conventionally grown samples. In addition to an enumeration of all such microbes, we define and provide a discussion of the genera that form the “core” romaine lettuce mycobiome, which represent 85.5% of all obtained isolates: Alternaria, Aureobasidium, Cladosporium, Filobasidium, Naganishia, Papiliotrema, Rhodotorula, Sampaiozyma, Sporobolomyces, Symmetrospora and Vishniacozyma. We highlight the need for additional mycological expertise in that 23% of species in these core genera appear to be new to science and resolve some taxonomic issues we encountered during our work with new combinations for Aureobasidium bupleuri and Curvibasidium nothofagi. Finally, our work lays the ground for future studies that seek to understand the effect these communities may have on preventing or facilitating establishment of exogenous microbes, such as food spoilage microbes and plant or human pathogens.
1. Introduction
Microorganisms—bacteria and fungi, including yeasts—are ubiquitous components of the food supply. The rise in organic food production has resulted in, among other things, an increase in naturally occurring microbial populations on produce. Whereas the majority of microbiota on fresh produce are likely to be harmless or even beneficial, many species are responsible for food spoilage and some can cause serious diseases in humans [1]. For example, the Fusarium fujikuroi Nirenberg species complex (FFSC) encompasses over fifty phylogenetic species, a number of which were found to be among the principal fungal contaminants of maize grains from rural commodity markets in Kenya [2]. These species are causal agents for ear rot of maize and all of them produce fumonisins and related mycotoxins that are carcinogenic in humans and animals [3,4].
The effect that natural plant microbial communities may have on exogenous (=introduced) microbes (such as plant or human pathogens) is poorly understood, to the extent that basic data are lacking as to whether microbial interactions may inhibit or facilitate pathogen spread. In addition, for most produce, essential basic data on naturally occurring microbial communities are missing.
Romaine lettuce (Lactuca sativa L.) is one of the most commonly used vegetable crops in the USA, with a rapidly expanding market [5]. The romaine lettuce leaf surface, or phylloplane, like that of all plant leaves, is home to innumerable microbes—fungal and bacterial. However, because lettuce leaves are nearly always consumed raw rather than cooked, the microbes on the lettuce phylloplane are often ingested by the consumer. Many outbreaks of human pathogenic bacteria, such as shiga-toxin producing Escherichia coli, are associated with romaine lettuce; it has been among the fresh produce items most implicated in food-borne diseases between 1973 and 1997 [6]. Escherichia coli strain O157:H7 is the most common bacterial causal agent of the outbreaks associated with romaine lettuce and other leafy green vegetables [7,8,9,10]. Sources of E. coli outbreaks on leafy greens such as lettuce may be contaminated soil, irrigated water and fecal animal deposits [11,12,13]. During the mid-2018 outbreak, 210 people from 36 states were infected with E. coli O157:H7; 96 were hospitalized. This E. coli strain can cause bloody diarrhea, stomach cramps, vomiting and kidney failure. In some cases, the gut poisoning can be fatal; following the mid-2018 outbreak, five people died [14].
Despite this, our knowledge about natural fungal species associated with the phylloplane of romaine lettuce is still limited, yet this knowledge is necessary for understanding how this naturally occurring flora interacts with introduced human pathogens on the phylloplane. In a large-scale effort to characterize the fungal microbiota of romaine lettuce from a public health perspective, we want to answer the following questions: (i) What species are typically part of the fungal community on romaine lettuce leaves? (ii) What drives changes in fungal communities over time? (iii) Which romaine lettuce community type—organic versus conventional—is most vulnerable to establishment by E. coli? Resolving these questions is critical to understanding the ability of human-pathogenic bacteria to enter and persist in these romaine lettuce fungal communities as well to developing a reasonable strategy for mitigating against future outbreaks of E. coli.
Toward that goal, in this paper, we investigate the fungal communities present on the phylloplane of commercially grown romaine lettuce obtained bought in grocery stores in the USA, as well as lettuce heads grown hydroponically. We combined culturing techniques and a Sanger sequencing approach to compare communities between conventionally grown and organic samples.
2. Materials and Methods
2.1. Collection of Samples and Fungal Isolation
Lettuce heads were obtained from commercial vendors in Indiana (Indianapolis, Lafayette, West Lafayette), Illinois (Champaign, Chicago), Virginia (Springfield) and Washington, D.C. between April and December, 2016. In addition, we obtained three hydroponically grown romaine lettuce heads from an indoor environmentally controlled facility based in Cleveland, Ohio. Lettuce heads were stored at 4 °C overnight and then immediately processed. In total, we examined 42 lettuce heads—grown either conventionally (n = 25), organically (n = 14), or hydroponically (n = 3). External leaves were discarded prior to random sampling of leaves from the exterior towards the center of each lettuce head. A sample of 25 g from different leaves was obtained from each lettuce plant and blended in 225 mL of sterile 100 µM phosphate buffer (5.4 g/L monosodium phosphate + 8.7 g/L disodium phosphate). From each sample, a 1 mL aliquot was plated in serial dilutions up to 10-3 on each of the following media to recover different groups of fungi (yeasts, filamentous and slow-growing): (i) Yeast Malt extract Agar (YMA), (ii) Potato Dextrose Agar (PDA) and (iii) Rose Bengal Agar (RBA), all with 2% agar and supplemented with 50 µg/mL chloramphenicol and 100 µg/mL ampicillin (all BD, Franklin Lakes, NJ, USA) to inhibit bacterial growth.
From each lettuce sample, up to 12 colonies were chosen for subculturing. Selection of colonies was based on morphology; colonies with differing morphologies on each plate were selected; additional colonies with similar morphologies were selected secondarily to increase the likelihood of obtaining morphologically similar but phylogenetically different species. Axenic strains obtained after several rounds of subculturing are maintained on PDA slants at 4 °C and in 15% (for filamentous strains) or 40% (for yeast-like strains) glycerol at −80 °C in the culture collection of M.C. Aime (MCA), Department of Botany and Plant Pathology, Purdue University, West Lafayette, IN, USA.
2.2. Identification of Fungal Isolates
DNA from axenic cultures was extracted using the Wizard Genomic DNA Purification Kit (Promega, Madison, Wisconsin) or amplified directly using a colony PCR method [15]. We amplified the internal transcribed spacer region of the ribosomal DNA (ITS) for all pure strains. Primer combinations used were ITS1f/ITS4 and ITS1f/ITS4B [16,17]. Amplifications were performed on a pro S Mastercycler (Eppendorf, Hauppauge, New York) in 25 reactions containing 12.5 μL of 2× MyTaq Mix (Bioline, Swedesboro, New Jersey), 9.5 µL of ddH2O and 1.0 µL of DNA. Cycling conditions included an initial denaturation at 94 °C for 5 min; followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 50 °C for 45 s, extension at 72 °C for 45 s; and a final extension step at 72 °C for 7 min. PCR products were visualized on 1% agarose gel using 1× TAE buffer and SYBR GelRed. Purification and sequencing were outsourced to Genewiz (Plainfield, NJ, USA). Sequence reads were assembled, trimmed and edited in Sequencher 5.0 (Gene Codes Corporation, Ann Arbor, Michigan). All forward and reverse identical reads were collapsed to separate unique sequences. Generated sequences were deposited to NCBI GenBank (accession numbers in Supplementary Table S1). All ITS sequences were subjected to a nucleotide BLAST search for initial identification to genus level (http://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 7 April 2021). Tentative generic IDs were confirmed by morphology of originating cultures.
2.3. Statistical Analyses
To determine whether treatment (conventional, organic, hydroponic) had a significant effect on the presence of fungal isolates, we performed a one-way ANOVA with α = 0.05. Because the three-treatment dataset was biased (30 isolates from hydroponic samples versus 147 from conventional samples and 154 from organic samples), we also analyzed the effect of treatment only considering fungal isolates from conventional and organic lettuce samples. We calculated the sample variances for both conventional and organic isolates (25.36 and 26.11, respectively) and then performed a two-tailed t test with equal variance. We also analyzed the effect of treatment (conventional, organic) on the number of obtained isolates as well as on the number of species they represent, based on isolates of the “core” community alone (fide [18])—including representatives of the genera Alternaria Nees; Aureobasidium Viala & G. Boyer; Cladosporium Link; Filobasidium L.S. Olive; Naganishia Goto; Papiliotrema J.P. Samp., M. Weiss & R. Bauer; Rhodotorula F.C. Harrison; Sampaiozyma Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout; Sporobolomyces Kluyver & C.B. Niel; Symmetrospora Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout; and Vishniacozyma Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout. All statistical analyses were done in Excel 2016 (Microsoft, Redmond, WA, USA) using the XLMiner Analysis ToolPak App (Frontline Systems Inc., Incline Village, NV, USA) downloaded from the Microsoft Office store.
2.4. Phylogenetic Analyses
In many genera, a simple BLAST query will not be sufficient for accurate species identification. This is because no single sequence similarity threshold will be able to distinguish intraspecific from interspecific variability in all fungi [19], especially yeasts [20]. For most genera in the core community, we constructed a dataset of ex-type sequences of most similar taxa downloaded from GenBank and supplemented with our newly generated sequences (strains and GenBank accession numbers in Supplementary Table S1). Alignments were constructed using MUSCLE v. 3.7 [21], available on the CIPRES Science Gateway v. 3.3 [22]. After alignment of the ITS datasets, partial SSU and partial LSU were removed by looking for the motifs 5′-ATCATTA-3′ (3′ end of SSU) and 5′-TGACCT-3′ (5′ start of LSU) and deleting downstream and upstream sequence data, respectively [23]. Phylogenetic relationships were inferred by maximum likelihood (ML). We used the command-line version of IQ-TREE [24,25]. Appropriate models of nucleotide substitution were selected according to the corrected Akaike Information Criterion (AICc) through ModelFinder [26] (Table 1). Ultrafast bootstrapping was done with 1000 replicates [27]. Final trees with ML bootstrap support values (BS) were visualized in FigTree (http://tree.bio.ed.ac.uk/software/figtree/, accessed on 7 April 2021) and edited in Adobe Illustrator 24.1.1 (San Jose, CA, USA).
3. Results
3.1. Culture-Based Community from Romaine Lettuce
A total of 331 fungal isolates were obtained from 42 romaine lettuce samples; 154 isolates were obtained from 14 organic lettuce samples, 147 from 25 conventional samples and 30 from three hydroponic samples. ITS barcode sequences were generated for 249 of these isolates, representing 59 species in 28 genera (Appendix A, Table A1). At the level of phylum, the majority of isolates were Basidiomycota (63.5%), followed by Ascomycota (36.1%) and Mucoromycota (0.4%, a single isolate). The subphylum that was most represented among lettuce isolates was Pezizomycotina (35.0%), followed by Agaricomycotina (32.5%) and Pucciniomycotina (30.1%). The other subphyla in our dataset were represented by one to three isolates: Mucoromycotina, Saccharomycotina, Ustilaginomycotina. In terms of growth, yeast-like fungi were most represented with 170 isolates (68%), compared to 79 isolates of filamentous fungi (32%). Filamentous fungi were distributed in Pezizomycotina (77 isolates), Agaricomycotina (1) and Mucoromycotina (1). The yeast-like fungi were distributed in Agaricomycotina (80 isolates), Pucciniomycotina (75), Pezizomycotina (10), Saccharomycotina (3) and Ustilaginomycotina (2).
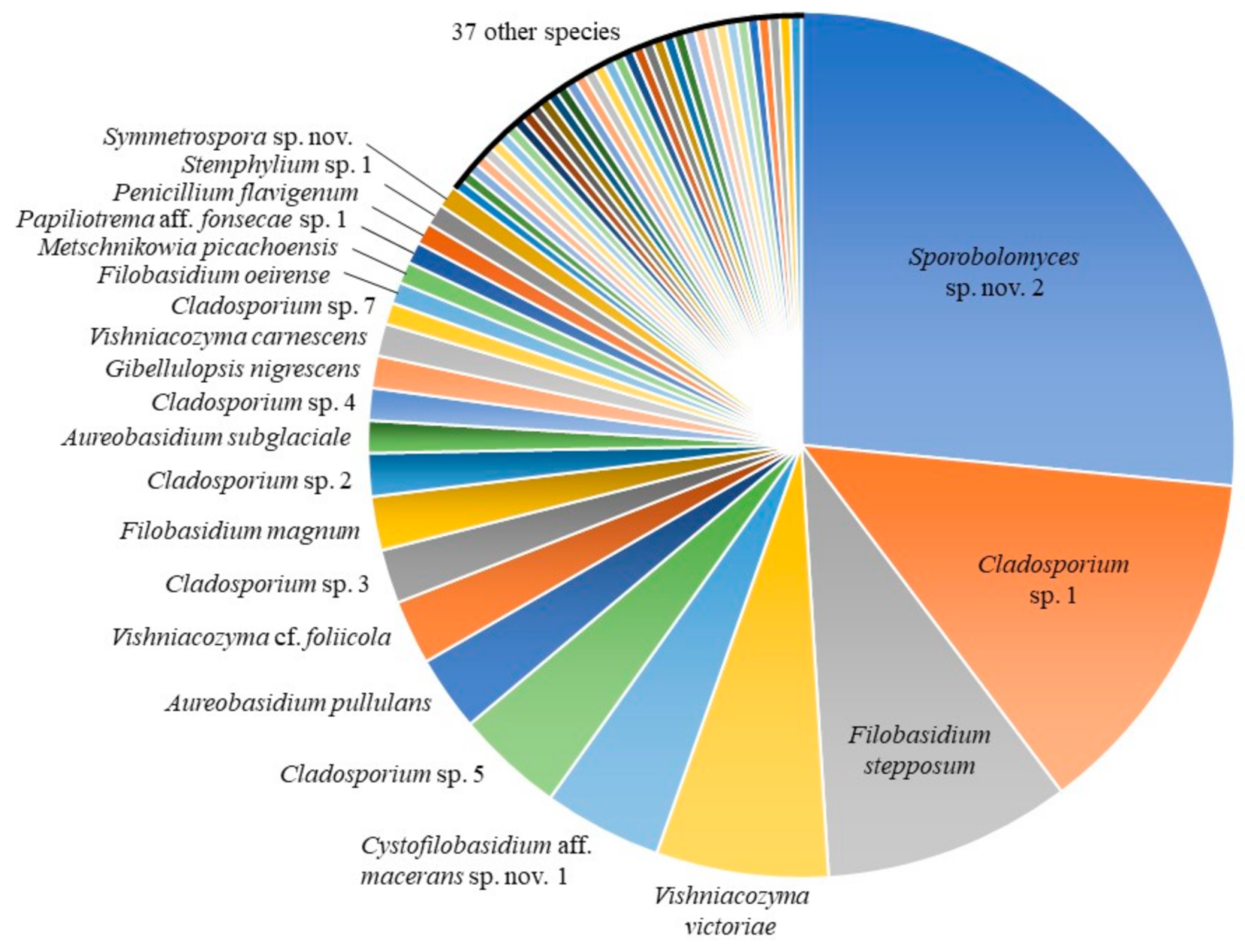
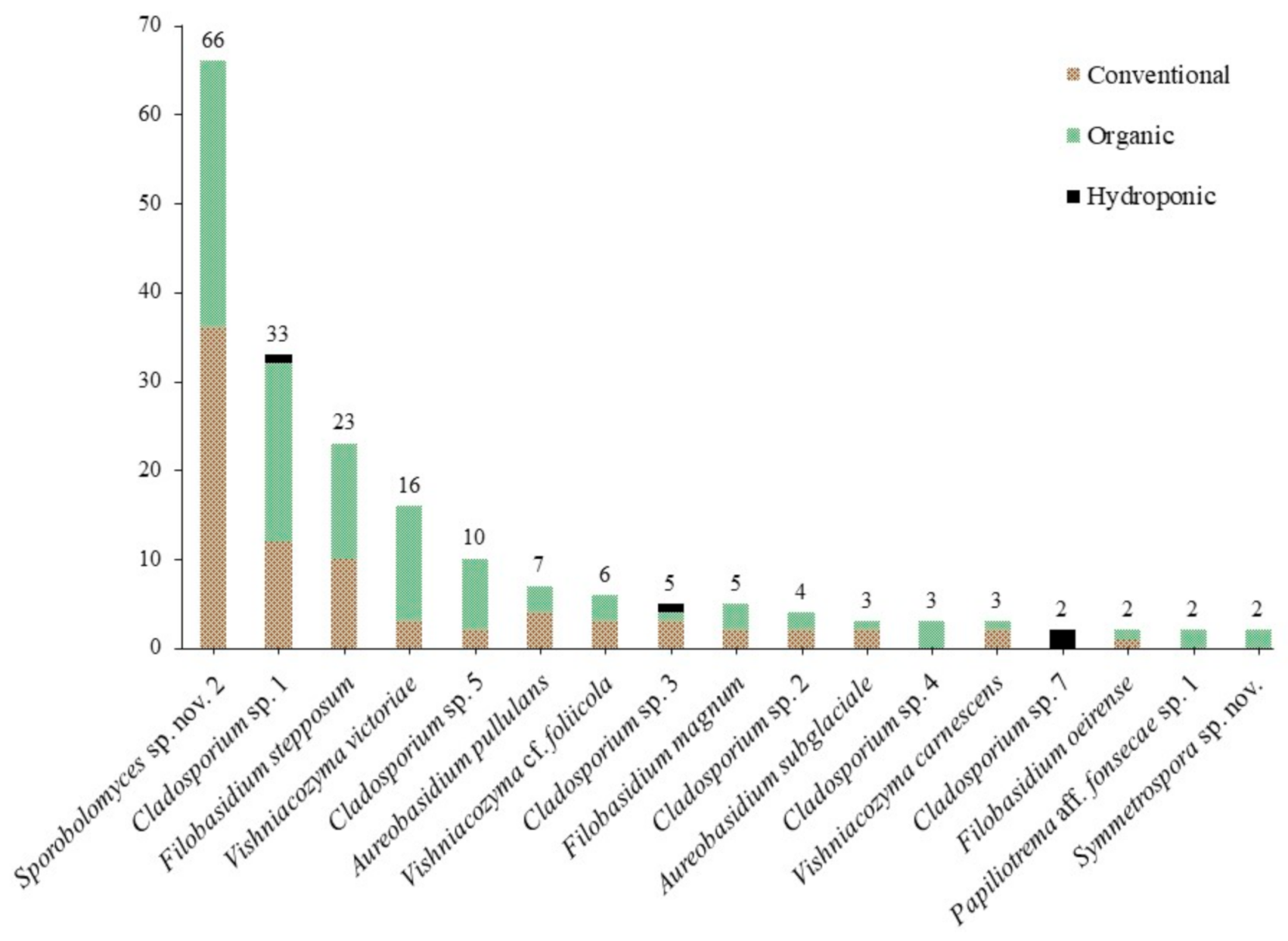
The most abundant taxa in our culture-based community belonged to the genus Sporobolomyces (26.9%), followed by Cladosporium (23.7%), Filobasidium (13.3%), Vishniacozyma (10.8%), Cystofilobasidium Oberw. & Bandoni (4.8%) and Aureobasidium (4.0%) (Table 2). Fifty-nine species were identified using the ITS barcode region (Figure 1). The highest number of isolates obtained for a single species was 66: an undescribed species of the red-yeast genus Sporobolomyces (Pucciniomycotina, Microbotryomycetes), with 36 isolates from 16 conventional lettuce samples and 30 from 11 organic samples (Figure 2). We obtained isolates of this species from 27 of 42 lettuce samples (64.3%). Five other species were abundant, with 10 or more isolates: 33 isolates of Cladosporium sp. 1 and 10 isolates of Cladosporium sp. 5 (Pezizomycotina, Dothideomycetes); 23 isolates of Filobasidium stepposum (Golubev & J.P. Samp.) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout; 16 isolates of Vishniacozyma victoriae (M.J. Montes, Belloch, Galiana, M.D. García, C. Andrés, S. Ferrer, Torr.-Rodr. & J. Guinea) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout; and 11 isolates of Cystofilobasidium aff. macerans J.P. Samp. sp. nov. 1 (Agaricomycotina, Tremellomycetes). Between two and seven isolates were obtained for sixteen species and 37 species were found only once (one isolate each).
3.2. Statistical Analyses
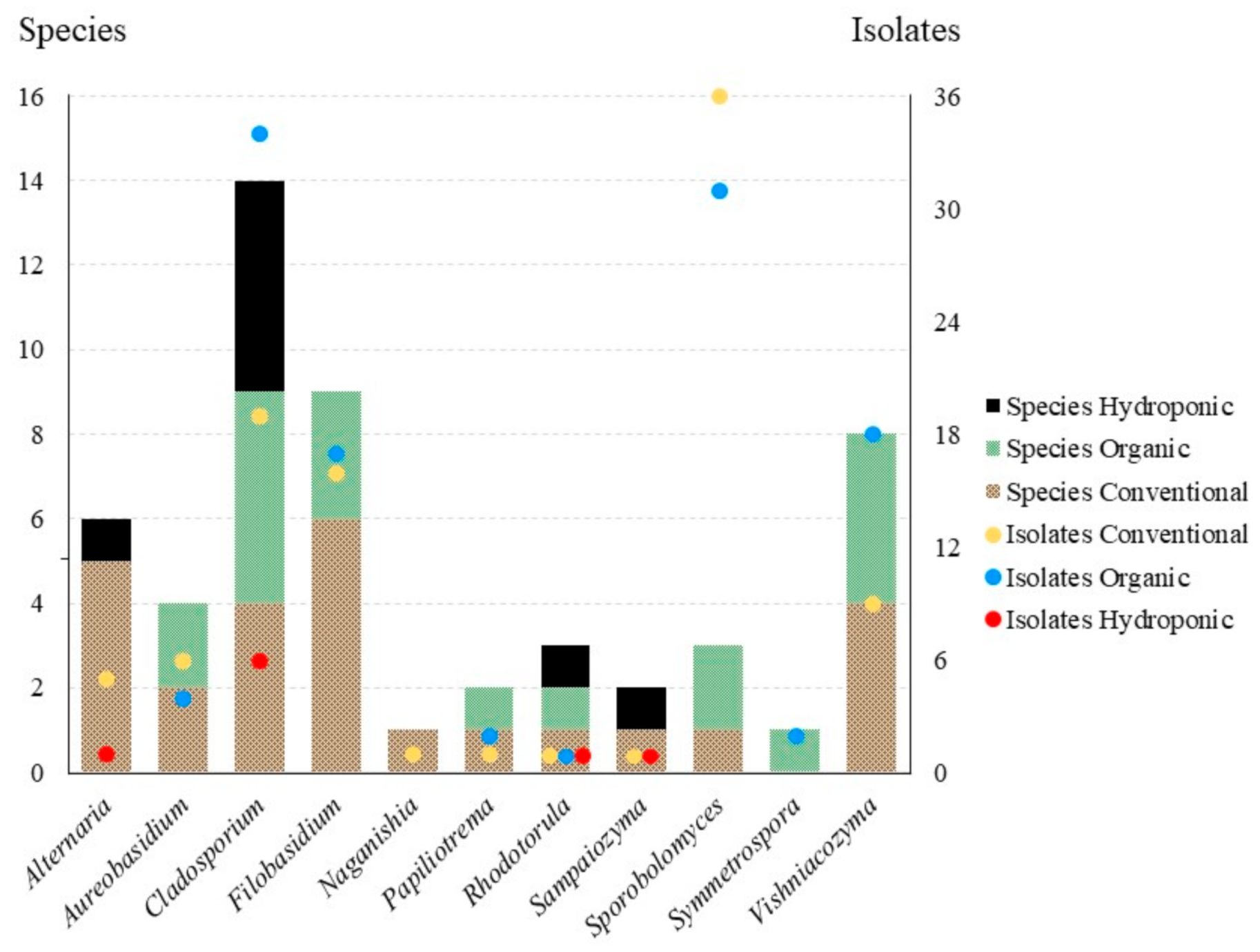
We obtained a total of 110 isolates representing 34 species from conventional lettuce samples, 128 isolates and 33 species from organic lettuce samples and eleven isolates and ten species from the hydroponic samples. Effect of treatment was significant among all samples (three treatments, F(2,174) = 3.9067, p = 0.0219), but it was not significant when only isolates from conventional and organic lettuce samples were compared (to treatments, t(116) = 0.3266, p = 0.7445). Of the 11 genera in the core community, 10 were obtained from conventional lettuce samples (95 isolates, 26 species), eight from organic samples (109 isolates, 19 species) and four from hydroponic samples (nine isolates, eight species) (Figure 3). For the core community, the effect of treatment on both the number of isolates (2 treatments, t(20) = 0.2473, p = 0.8072) and the number of species (t(20) = 0.8051, p = 0.4302) was not significant.
3.3. Taxonomy
3.3.1. Aureobasidium bupleuri (Bills) Haelew. & Aime, comb. nov.
MycoBank MB835676.
Basionym: Kabatiella bupleuri Bills, Mycologia 104 (4): 966 (2012).
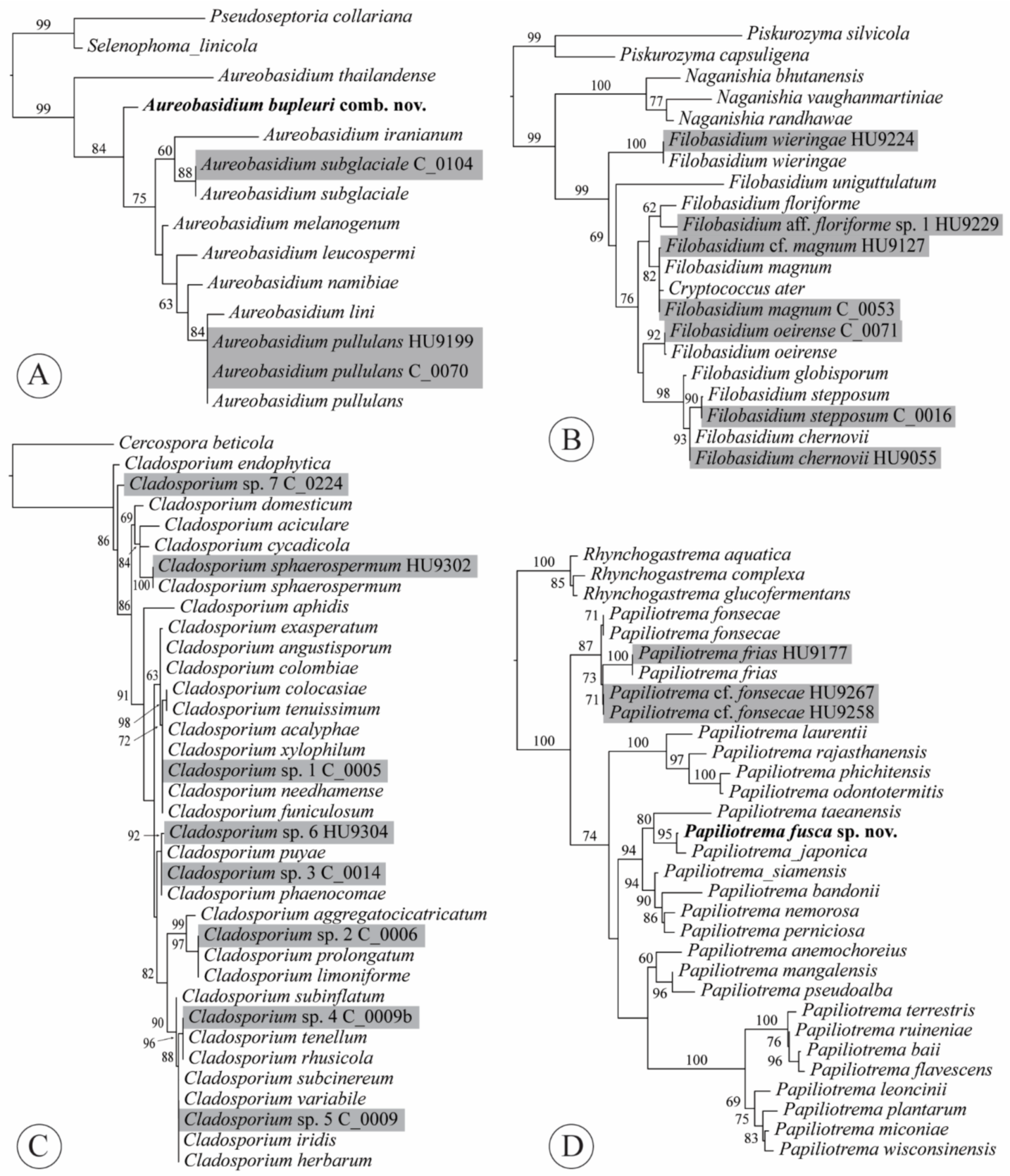
Notes: The combination was based on phylogenetic evidence. Taxonomy of the black yeasts has been traditionally confusing, with characters that are usually important in the classification of asexual morphs being phylogenetically uninformative in Dothideales. In our ITS tree, the ex-type sequence of K. bupleuri (GenBank acc. no. NR_121524 [28]) was nested within Aureobasidium (Figure 4A). The same result was found based on an ITS phylogeny, as well as an rpb2 phylogeny that both included ex-type sequences of K. bupleuri [29]. Both analyses also included the type species of Kabatiella, K. microsticta Bubák, which was retrieved within Aureobasidium with high support.
3.3.2. Curvibasidium nothofagi (C. Ramírez & A.E. González) Haelew. & Aime, comb. nov.
MycoBank MB835657.
Basionym: Rhodotorula nothofagi C. Ramírez and A.E. González, Mycopathologia 91 (3): 171 (1985).
Notes: The combination was made based on phylogenetic evidence. Kurtzman et al. [30] presented a phylogeny with sequences of the large subunit (LSU) ribosomal RNA gene that retrieved R. nothofagi in the Curvibasidium Samp. & Golubev clade [31] together with Curvibasidium cygneicollum J.P. Samp. and C. pallidicorallinum Golubev, Fell & N.W. Golubev with high support. In addition, blasting the ex-type ITS sequence (GenBank acc. no. AB038096, CBS:8166, JCM:9304; T. Nagahama et al. unpubl.) against other sequences from type materials, resulted in C. cygneicollum (NR_111077, CBS:4551 [32]) and C. pallidicorallinum (KY102982, CBS:9091 [20]) with the highest percentages of identity (98.86% and 99.48%, respectively).
3.3.3. Filobasidium magnum (Lodder & Kreger-van Rij) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout, in Liu et al., Stud. Mycol. 81: 118 (2015)
Basionym: Cryptococcus laurentii var. magnus Lodder & Kreger-van Rij, in Lodder and Kreger-van Rij, The yeasts: a taxonomic study [Edn 1] (Amsterdam): 670 (1952).
Synonyms: Cryptococcus ater (Castell. ex W.B. Cooke) Phaff & Fell, in Lodder, The yeasts: a taxonomic study, 2nd edn (Amsterdam): 1120 (1970), syn. nov.
Cryptococcus laurentii f. ater Castell. ex W.B. Cooke, Mycopath. Mycol. Appl. 30: 351 (1966), syn. nov.
Cryptococcus magnus (Lodder & Kreger-van Rij) Baptist and Kurtzman, Mycologia 68(6): 1200 (1977) [1976].
Notes: Cryptococcus laurentii (Kuff.) C.E. Skinner was combined as Papiliotrema laurentii (Kuff.) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout by Liu et al. [33] but C. ater [= C. laurentii f. ater] represented another species. The new synonymy was proposed based on sequence data and phylogenetic evidence. The ex-type ITS sequences of C. ater (GenBank acc. no. NR_144813; P. Gujjari and J. Zhou unpubl.) and F. magnum (NR_130655 [34]) were 98.91% identical, indicating a single species. Fell et al. [34] presented two phylogenies, one based on ITS, the other based on LSU, that both found C. ater and C. magnus to be a single species, which was confirmed by our analysis (Figure 4B). However, the synonymy was not formalized until now. Or it was incorrectly implemented; C. ater and C. laurentii f. ater were listed as synonyms of P. laurentii [35].
3.3.4. Papiliotrema fusca J.P. Samp., J. Inácio, Fonseca and Fell ex Haelew., sp. nov.
MycoBank MB836093
For description see: Int. J. Syst. Evol. Microbiol. 54 (3): 989 (2004).
Holotype: PYCC 5690, permanently preserved in a metabolically inactive state at the Portuguese Yeast Culture Collection. Ex-type culture: CBS 9648.
Synonyms: Auriculibuller fuscus J.P. Samp., J. Inácio, Fonseca & Fell, Int. J. Syst. Evol. Microbiol. 54 (3): 989 (2004), nom. inval., Art. 40.6 (Shenzhen).
Papiliotrema fusca (J.P. Samp., J. Inácio, Fonseca & Fell) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout, Stud. Mycol. 81: 126 (2015), nom. inval., Art. 40.6 (Shenzhen).
4. Discussion
Previous culture-based studies have found that the phylloplane of crops is dominated by species of Acremonium Link, Alternaria, Aureobasidium, Cladosporium (Ascomycota, all filamentous), Cryptococcus Vuill., Rhodotorula and Sporobolomyces (Basidiomycota, all yeast-like) [18,36,37,38,39]. We note that according to pre-2015 generic concepts some of these genera were polyphyletic. Many of these taxonomic problems are now resolved based on multi-locus phylogenies and new genera were introduced to accommodate orphaned species, including Filobasidium, Naganishia, Papiliotrema, Vishniacozyma (Tremellomycetes), Sampaiozyma (Microbotryomycetes) and Symmetrospora (Cystobasidiomycetes) [31,33]. Under these modern concepts, our results are consistent with previous work; the 213 isolates of Alternaria, Aureobasidium, Cladosporium, “Cryptococcus” [as Filobasidium, Naganishia, Papiliotrema and Vishniacozyma], Rhodotorula [in part as Sampaiozyma] and Sporobolomyces [in part as Symmetrospora] represent 85.5% of total isolates and, as a result, can be considered the core community [fide 18] (Figure 3). Of these isolates, the yeast-like taxa dominate (64.8% of the core community, while only 55.4% of the total culture-based dataset). We will discuss these genera below.
4.1. Alternaria
We obtained six isolates of Alternaria (Dothideomycetes, Pleosporales, Pleosporaceae), each representing a different species—five isolates were from conventional lettuce samples, one was from a hydroponic sample. Alternaria is a large genus of filamentous fungi with about 366 accepted species [40]. Species are commonly found on a wide range of foods, mostly under the names A. alternariae (Cooke) Woudenb. & Crous and A. tenuis Nees [syn. of Alternaria alternata (Fr.) Keissl.]. Several species are major pathogens of fresh fruit, eggplants, peppers and fresh vegetables [1,41].
The utility of ITS as a barcode for separating species in this genus was questioned earlier [42,43]. We have encountered this issue during our work with Alternaria isolates from romaine lettuce. For example, the ITS sequence of isolate HU9315 from a hydroponic lettuce sample was 100% similar to both ex-type sequences of A. angustiovoidea E.G. Simmons and A. destruens E.G. Simmons. We went one step further and compared ex-type sequences for the glyceraldehyde 3-phosphate dehydrogenase (gpd) gene to find that A. angustiovoidea (GenBank acc. no. JQ646315 [44]) and A. destruens (AY278812 [45]) can be separated from each other with that marker, with 1.04% nt differences between the two species. Since our study was based on ITS alone, we did not assign names to our isolates. Instead, we referred to them as Alternaria spp. 1–6 (Appendix A, Table A1). They are most close to A. angustiovoidea; A. arbusti E.G. Simmons; A. californica E.G. Simmons & S.T. Koike; A. conjuncta E.G. Simmons; A. dactylidicola Thambug., Camporesi & K.D. Hyde; A. destruens; A. eureka E.G. Simmons; A. oregonensis E.G. Simmons; A. prasonis E.G. Simmons; and A. ventricose R.G. Roberts. To our knowledge, none of these have been reported on lettuce or any member of Asteraceae before. We note that artificial transmission of Alternaria to atypical hosts has been reported. For example, A. dauci (J.G. Kühn) J.W. Groves & Skolko, causing leaf blight on carrot (Daucus carota L., Apiaceae), has been shown to infect non-Apiaceae hosts including lettuce [46,47,48].
Of the potential species in our dataset, only A. arbusti has a wide host range, including species in the genera Hordeum L., Phleum L. and Triticum L. (Poaceae); Prunus L. and Pyrus L. (Rosaceae); Sesbania Scop. (Fabaceae); and Solanum L. (Solanaceae). However, it had not been reported from lettuce before [49,50,51,52]. These findings highlight the need for additional taxonomic work within Alternaria and research on the production of toxins and their host-selectiveness for each species, from a public health standpoint.
4.2. Aureobasidium
We obtained ten isolates of Aureobasidium (Dothideomycetes, Dothideales, Saccotheciaceae), six isolates from six conventional lettuce samples and four isolates from three organic samples. Seven isolates represented A. pullulans (de Bary & Löwenthal) G. Arnaud, based on 100% similarity matches with the ITS sequence generated for the neotype [53]. Aureobasidium currently comprises 26 species, of which only A. pullulans is important in the food industry [1,40,54]. The black yeast-like A. pullulans is a ubiquitous saprobe, especially found on cold-stored and frozen foods as well as fresh produce. It was isolated from a wide range of vegetables but not Cucurbitaceae [55]: green beans, mustard greens, turnip greens, asparagus, broccoli, cabbage, chinese cabbage, cauliflower, kale, lettuce, okra and radish. Aureobasidium pullulans has applications in biological control [56], e.g., against postharvest Penicillium rot in apple and citrus [57,58] and fire blight caused by Erwinia amylovora (Burril) Winslowfire et al. in pome fruit trees [59].
The other species in our dataset, represented by three isolates, was A. subglaciale (Zalar, de Hoog & Gunde-Cim.) Zalar, de Hoog & Gunde-Cim. (Figure 4A, Appendix A, Table A1). The type of this species was isolated from subglacial ice from sea water in Norway. Given its psychrotolerant nature and exclusive presence in the Arctic [60], it is quite surprising to find three isolates of this species from three of our North American romaine lettuce samples—two from conventionally grown lettuce samples and one from organic lettuce. This may indicate that some treatments, such as refrigeration, may favor growth of microbes that are adapted for different climatic conditions and would otherwise not be able to compete on the phylloplane.
4.3. Cladosporium
Of 77 isolates of Dothideomycetes, 59 represented species of Cladosporium (Capnodiales, Cladosporiaceae). Those isolates were obtained from 28 of 42 lettuce samples (66.7%). Nineteen isolates were obtained from twelve conventional lettuce samples, 34 isolates from thirteen organic samples and six isolates from three hydroponic ones. Species of Cladosporium are among the most abundant fungi in outdoor and indoor air [61,62,63,64,65]; it is no surprise that the second-highest number of isolates from lettuce samples were Cladosporium. This genus currently includes 237 accepted species isolated from a range of substrates including plant material, soil, the air, food, building materials and clinical samples [40,65,66,67]. Species accepted in Cladosporium occur as pathogens on fresh fruit and vegetables, causing spoilage of strawberries, tomatoes and refrigerated cheese and meat. In other instances, they are merely colonizers [1,66].
Phylogenetic studies have shown that many morphologically circumscribed species comprise several cryptic species. Based on morphology, most species of Cladosporium can be assigned to one of three major species complexes—C. cladosporioides (Fresen.) G.A. de Vries, C. herbarum (Pers.) Link and C. sphaerospermum Penz. [65,68,69,70]. Using molecular data has improved understanding of species limits; the most informative phylogenetic markers are the actin gene (actA) and translation elongation factor 1-α (tef1) [66]. The ITS alone only provides accuracy to species complex-level; ITS sequences are usually identical for species belonging to the same species complex [71].
Five clusters of ITS sequences from our isolates were identical to ex-type sequences of 3–18 different species within either the C. cladosporioides complex or the C. herbarum complex (Figure 4C, Appendix A, Table A1), supporting previous findings [71]. Cladosporium sp. 1 is part of the C. cladosporioides complex, whereas Cladosporium spp. 2–5 all belong in the C. herbarum complex. Two isolates of Cladosporium sp. 7 from a hydroponic lettuce sample shared 99.11% identity with the ex-type sequence of C. endophyticum Tibpromma & K.D. Hyde, an endophytic species isolated from leaves of Pandanus sp. in Thailand. Finally, the ITS sequence of isolate HU9302 from a hydroponic lettuce sample shared 100% identity with the ex-neotype sequence of C. sphaerospermum, representing the only species of Cladosporium that we could identify with certainty. This species has been reported from a wide range of food, but is apparently less common than C. cladosporioides [1].
4.4. Filobasidium
A total of 33 isolates representing six species of the yeast genus Filobasidium (Tremellomycetes, Filobasidiales, Filobasidiaceae) were obtained from 20 lettuce samples. Sixteen isolates were obtained from eleven conventional lettuce samples, 17 isolates from nine organic samples. Filobasidium currently contains 11 species isolates from leaves and florets of various plants, weathered inflorescence scapes of Yucca L., apples, grapes (including grape juice and must), Euploea Fabricius butterflies, sake moto, cider, breweries/wineries, cheeses, raw milk and forest soil [30,33,72,73,74,75,76]. In addition, F. globosporum Bandoni & Oberw. and F. sp. (as “elegans”) have been identified in the intestines of humans and mice [77,78], whereas F. uniguttulatum Kwon-Chun isolates have been obtained from a patient with meningitis and from cloaca and fresh dropping samples of feral pigeons [79,80].
Li et al. [76] primarily used LSU for species delimitation phylogenetic analyses, but these authors also discussed number of nucleotide mismatches among ITS sequences in their assessments of species limits. A ≥ 3% mismatch in the ITS for a certain isolate with an ex-type was considered as an indication of that isolate representing a separate species. Following the same strategy, our isolates represent F. chernovii (Á. Fonseca, Scorzetti & Fell) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout (one isolate); F. magnum (Lodder & Kreger-van Rij) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout (five isolates); F. oeirense (Á. Fonseca, Scorzetti & Fell) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout (two isolates); F. stepposum (Golubev & J.P. Samp.) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout (23 isolates) and F. wieringae (Á. Fonseca, Scorzetti & Fell) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout (one isolate) (Appendix A, Table A1). In addition, we isolated a single isolate (HU9229) of F. aff. floriforme L.S. Olive sp. 1. Compared to the ex-type sequence of F. floriforme, there were four mismatches and one gap (99.06% identity). Even though support was low, the evolutionary distance between the two isolates in our ML tree of Filobasidium ITS sequences (Figure 4B) showed that they are likely not the same species. All our isolates are the first ones from romaine lettuce. As most species appear to be non-pathogenic to humans and animals [30], the isolates we obtained may be merely environmental contaminants of the lettuce surface.
4.5. Naganishia
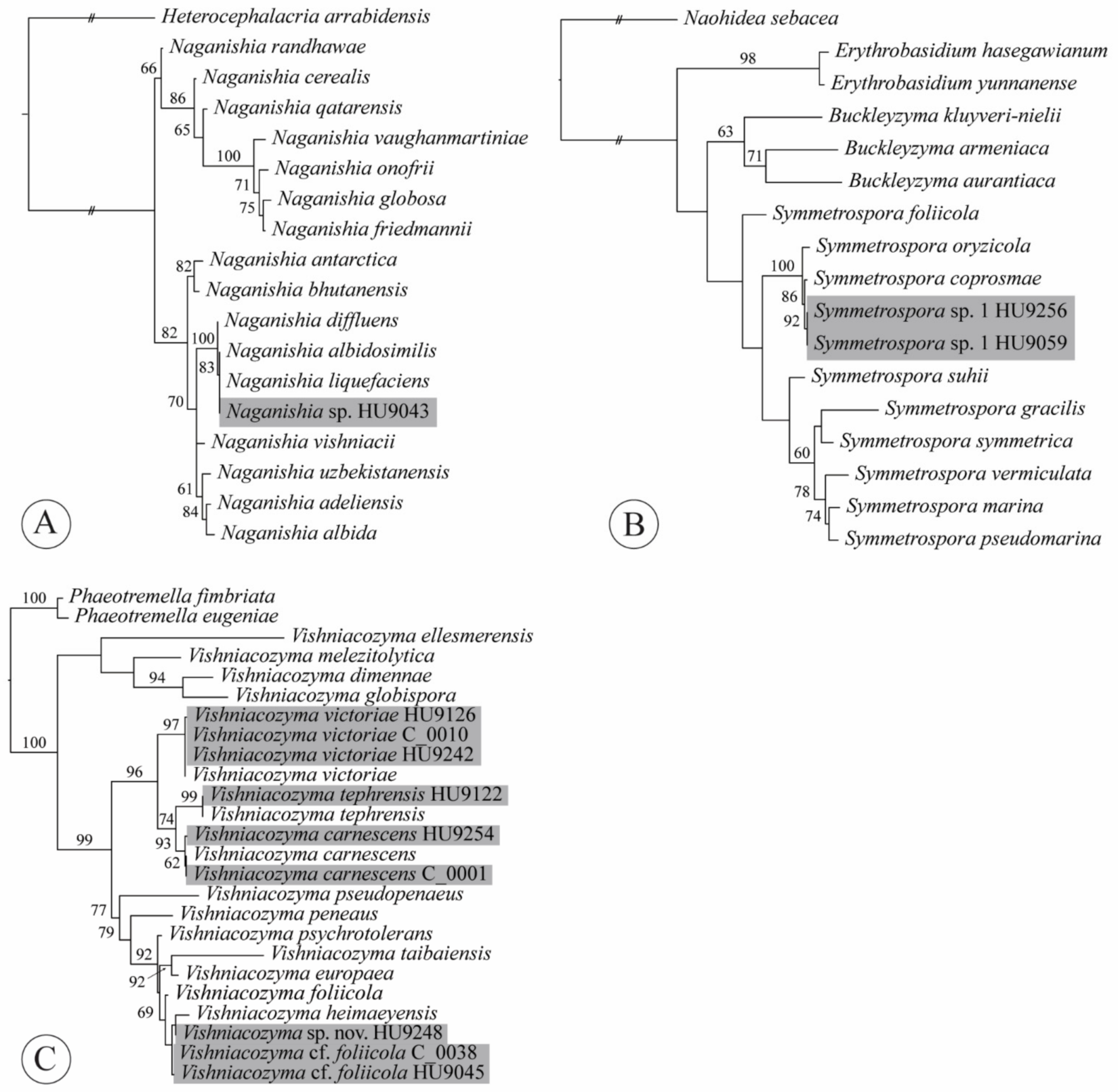
We isolated a single strain of Naganishia sp. (Tremellomycetes, Filobasidiales, Filobasidiaceae) from a conventional lettuce sample (Figure 5A, Appendix A, Table A1). The genus Naganishia was described to accommodate the yeast N. globosa Goto [81]. Liu et al. [33], based on an LSU phylogeny, synonymized 14 species that previously belonged to Cryptococcus (albidus clade (sensu [82])). Currently, 18 species of Naganishia are accepted [76,83,84,85]. Species in this genus have been isolated from air, water, soil, cold environments (glaciers, snow, cryoconite), plant surfaces, cheese, fermented cereals and a purification tank for polluted water [83,86]. Three species are reported from clinical samples: Naganishia diffluens (Zach) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout; N. globosa [syn. Cryptococcus saitoi Á. Fonseca, Scorzetti & Fell] and N. liquefaciens (Saito & M. Ota) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout [30]. Distinction among N. albidosimilis (Vishniac & Kurtzman) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout, N. diffluens and N. liquefaciens can be best achieved using LSU (D1/D2 loop region); the three species differ by 0–1 nt substitutions in the ITS. The sequence of our isolate HU9043 is 100% identical to both N. albidosimilis and N. liquefaciens. According to Kurtzman et al. [30], identifications of N. albidosimilis, N. diffluens and N. liquefaciens based solely on ITS sequences are best considered tentative. This is also visible from our ITS tree (Figure 5A), in which isolate HU9043 and the ex-type sequences of both N. albidosimilis and N. liquefaciens are placed together with high support, sister to N. diffluens. Neither of these species have thus far been found on leaves of lettuce.
4.6. Papiliotrema
We obtained three isolates of Papiliotrema (Tremellomycetes, Tremellales, Rhynchogastremaceae), two isolates of P. aff. fonsecae (V. de García, Zalar, Brizzio, Gunde-Cim. & Van Broock) Yurkov sp. 1 and one isolate of P. frias V. de García, Zalar, Brizzio, Gunde-Cim. & van Broock ex Yurkov (Appendix A, Table A1). Yeasts in the genus Papiliotrema have been isolated from inflorescences and leaf tissues of plants, trees, termite guts, subglacial ice and meltwater from glaciers, water samples and soil [87,88,89,90,91,92,93]. Interestingly, P. siamensis Suruss. & Limtong was found as both an epiphyte and an endophyte [91]. It appears that both ITS and LSU can be used for species delimitation in this genus; Pagani et al. [93] presented comparable nucleotide differences among species in the ITS and LSU regions. The phylogenetic reconstruction of an ITS dataset with our isolates and ex-type collections resolved all species described to date (Figure 4D). Isolate HU9177 obtained from a conventional lettuce sample was conspecific with P. frias. Isolates HU9258 and HU9267 (Figure 4D) are most similar with the ex-type sequence of P. fonsecae (99.76%, 1 nt different). However, they were retrieved as a sister clade to P. frias, but with considerable evolutionary distance. The distance between P. fonsecae isolates (EXF-4087 and ZM13F84) and our lettuce isolates was negligible, but it is likely that our isolates represent an undescribed species, which we here refer to as P. aff. fonsecae sp. 1. We note that we included a non-type sequence of P. fonsecae in this tree, because the ex-type sequence for P. fonsecae was very short and was retrieved in different positions without support in preliminary trees.
4.7. Rhodotorula
Rhodotorula (Microbotryomycetes, Sporidiobolales, Sporidiobolaceae) is one of two genera (Sporobolomyces being the other one) in which asexual red-pigmented yeasts were artificially placed based on morphology and physiology [94,95,96]. Both these genera are polyphyletic in their traditional sense [34,97,98] and representatives of Rhodotorula s.l. are retrieved in 17 different clades in Cystobasidiomycetes, Microbotryomycetes and Ustilaginomycetes [97,99]. Consequently, 27 species of Rhodotorula were recombined in 15 genera, leaving 19 accepted species in Rhodotorula emend. [31]. In our assessment of the genus, we found that R. nothofagi should be combined in Curvibasidium based on both ITS and LSU sequence data [20,30,32] (see Section 3.3).
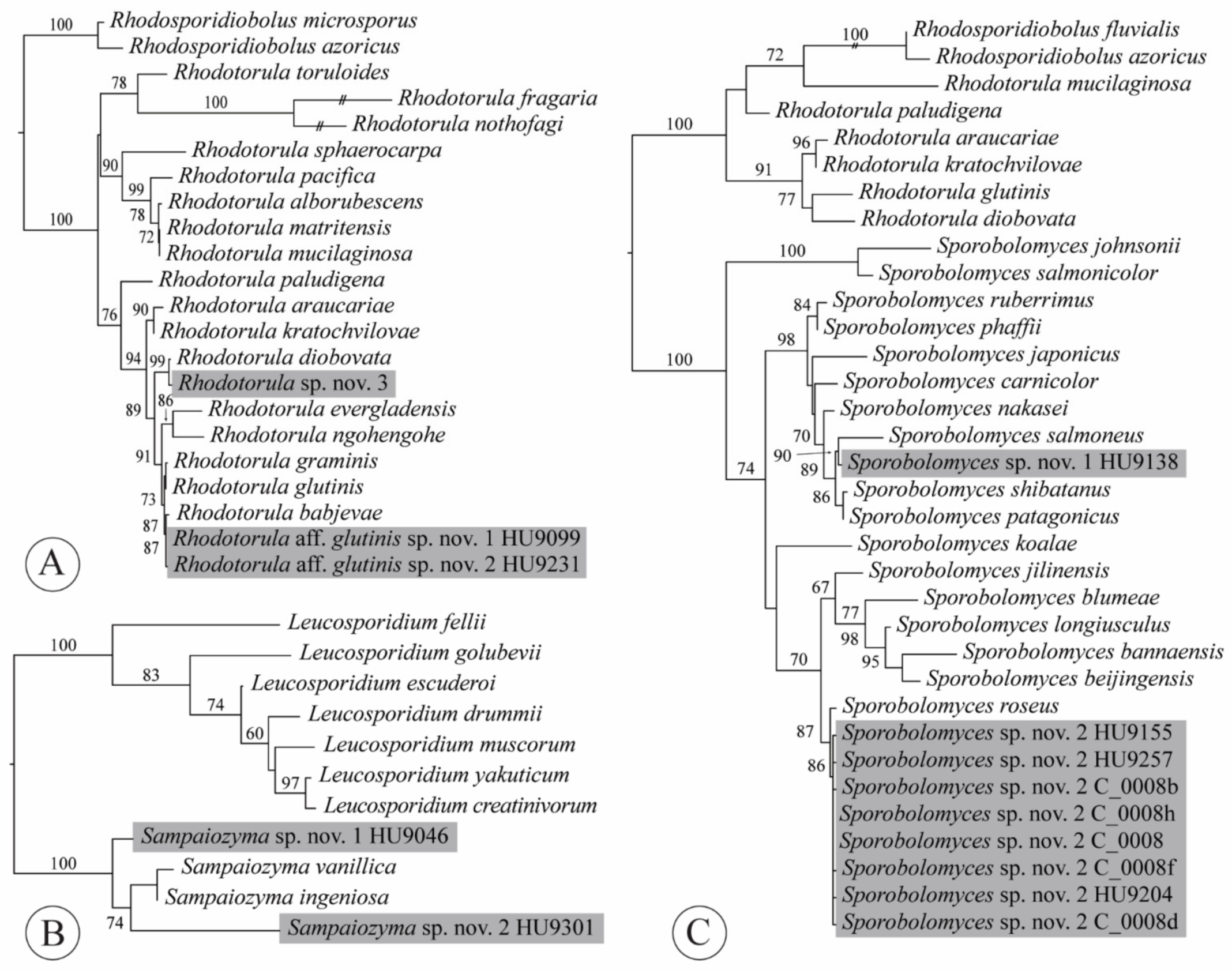
We obtained three isolates from three lettuce samples, each representative of another, potentially undescribed species (Appendix A, Table A1). Two isolates belonged in the R. glutinis sensu stricto group (Figure 6A), which was proposed by Gadanho and Sampaio [100] to accentuate the close phylogenetic relationships among R. babjevae (Golubev) Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout, R. glutinis (Fresen.) F.C. Harrison and R. graminis Di Menna. According to Kurtzman et al. [30], 1–3 mismatches were found among ITS sequences of these three species. Both isolates HU9099 and HU9231 were most similar to H. babjevae, with only two and three mismatches with the ex-type sequence for this species (GenBank acc. no. NR_077096 [101]), respectively. The ITS sequences of isolates HU9099 and HU9231 differed from each other in one nucleotide, which seems sufficient to recognize them as distinct species within the R. glutinis sensu stricto clade [sensu 30]. Our third isolate (HU9305), from a hydroponic lettuce sample, was most similar to R. diobovata (S.Y. Newell & I.L. Hunter) Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout, with three mismatches—thus, also representing an undescribed species.
Rhodotorula is a ubiquitous saprobic genus that is isolated from many different habitats, including in extreme conditions [102]. Gildemacher et al. [103] reported that Rhodotorula sp. suppresses other yeast species and increases russet formation on apples. A number of studies have mentioned biocontrol properties of Rhodotorula “glutinis” although their identifications may be erroneous because this species is rarely isolated compared to other morphologically similar species of red yeast [30]. Rhodotorula often occurs on the phylloplane and other types of plant substrates and species have been isolated from lettuce leaves in previous studies [104,105,106,107]. However, we are only aware of one study focusing on romaine lettuce [108]; red romaine lettuce (Lactuca sativa cv. “Outredgeous”) was grown in rooting pillows in a plant growth chamber on the International Space Station, harvested, brought back to earth and analyzed to genus-level. Of ten genera isolated from lettuce leaves and roots, Rhodotorula was the most common. Urbina and Aime [109] reported the urgency of formally characterizing undescribed species that are constantly consumed. This may be especially important for Rhodotorula species, some of which have been reported as emerging (opportunistic) pathogens for both humans and animals [102,110,111,112].
4.8. Sampaiozyma
Sampaiozyma (Microbotryomycetes incertae sedis) was one of the genera introduced by Wang et al. [31] to accommodate red-yeast species previously classified as Rhodotorula, isolated from stagnant water and leaf surfaces of pasture plants [113,114]: S. ingeniosa (Di Menna) Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout and S. vanillica (J.P. Samp.) Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout. Sampaiozyma is most closely related to Leucosporidium Fell, Statzell, I.L. Hunter & Phaff. Wang et al. [99] retrieved the genus—as the “vanillica clade”—as sister to Leucosporidium with high support in their ribosomal DNA and seven-locus phylogenies. Liu et al. [115] found high support for the sister relationship of Leucosporidium and Sampaiozyma. The ex-type ITS sequences of S. ingeniosa and S. vanillica only differ in three nucleotides. We obtained two isolates of Sampaiozyma, one from a conventional lettuce sample, the other from a hydroponic sample. The two isolates each represent an undescribed species (Figure 6B, Appendix A, Table A1). In the ITS sequence, isolate HU9046 had 13 mismatches compared to S. ingeniosa and 15 mismatches compared to S. vanillica. Isolate HU9301 differed from both species by 37 mismatches. Both lettuce isolates differed from each other by 39 mismatches in the ITS. Formal description of these is pending examination of morphological and physiological characteristics.
4.9. Sporobolomyces
Wang et al. [31] revised the red-yeast genus Sporobolomyces (Microbotryomycetes, Sporidiobolales, Sporidiobolaceae) because it was polyphyletic in the traditional sense, occurring in most of the yeast-forming classes of Pucciniomycotina, as well as in Agaricomycotina, Ustilaginomycotina and, most recently, in Pezizomycotina [97,99,116]. The authors proposed new combinations of 40 species in 16 different genera. The emended concept of Sporobolomyces included sixteen species. Six more species have been described by Lorenzini et al. [117] and Li et al. [76] bringing the number of currently accepted species to 22, although current estimates place the total number of species at 60 for Sporobolomyces [109]. The red-yeast genera in the Sporidiobolales produce lipid droplets full of carotenoid pigments, particularly β-carotene and torulene, which contribute to the rich pink to orange-red color of colonies [109,118,119]. These pigments, although poorly understood, seem to protect them against UV radiation and some provide antimicrobial, anticancer and antiaging activity [120,121]. Because of these characteristics, red-pigmented yeasts have gained interest from the pharmaceutical, cosmetics and biotechnology industries [122,123,124].
Sporobolomyces species are free-living and worldwide in distribution. They are reported from many different habitats—including freshwater and marine ecosystems, excrements of maize pests, fruit must, soil, buildings and air—although most commonly from the plant phylloplane [109,117,125,126,127,128,129,130,131]. Last [127] suggested an association between colony development and senescence, after observing that numbers of colonies of Sporobolomyces start to increase when cereal leaves reach their half-age. The promise of S. roseus Kluyver & C.B. Niel as a biocontrol against postharvest diseases of pome fruits has been shown by Janisiewicz et al. [132]. After yeast application to wounded apples, incidence of rots was reduced from 33% to 0% for Penicillium expansum Link (blue mold) and from 92% to 4% to Botrytis cinerea Pers. (gray mold). Important to note is that, in addition to the leaf age, humidity is also an important factor in the growth of S. roseus. Bashi and Fokkema [133] experimentally found that relative humidity of 65% decreased the population of colonies; Sporobolomyces can only use nutrients that are present on the leaves at relative humidity of at least 90%.
Sporobolomyces was the most abundantly recovered genus in our culture-based community, representing more than a quarter of all isolates (26.9%). Sixty-seven isolates were obtained from 27 of 42 lettuce samples (64.3%). Whereas the number of isolates was the highest of any genus isolated in this study, the number of species was only two. Both species appeared to be undescribed (Figure 6C, Appendix A, Table A1). Sporobolomyces sp. nov. 1 was represented by a single isolate (HU9138), obtained from an organic lettuce sample. It shares 98.82% identity with S. patagonicus Libkind, Van Broock & J.P. Samp. isolated from subsurface water in Argentina. Sporobolomyces sp. nov. 2 was detected on eleven organic lettuce samples (30 isolates) and sixteen conventional samples (36 isolates). Through a BLAST search in NCBI GenBank (100% identity hits), we found that this same yeast has a broad geographic distribution and is found from different habitats: on a wilting leaf of Parthenocissus sp. (Vitales, Vitaceae) in China [134], Prunus avium L. cherries (Rosales, Roaceae) in Spain [135], Vitis vinifera L. grapes (Vitales, Vitaceae) in Washington [136], glacier rocks in Antarctica [137], Quercus faginea Lam. leaves (Fagales, Fagaceae) in Portugal [20], Antirrhinum L. flowers (Lamiales, Plantaginaceae) in California, Septoria blasdalei-diseased leaves of Ceanothus arboreus Greene (Rosales, Rhamnaceae) in California [109] and guava juice in Egypt (Z.S.M. Soliman unpubl.).
4.10. Symmetrospora
From two organic lettuce samples, we obtained two isolates of an undescribed species of Symmetrospora (Cystobasidiomycetes incertae sedis, Symmetrosporaceae). Symmetrospora was introduced by Wang et al. [31] for species that were previously placed in the polyphyletic red-yeast genera Rhodotorula and Sporobolomyces of the “gracilis/marina clade” [34,97,99,138,139]. Haelewaters et al. [140] and Li et al. [76] recently proposed four new species and a new combination of Symmetrospora, bringing the total to 11 recognized species. Symmetrospora is known in North and South America, Europe, Asia and Australia and species in the genus have been isolated from a wide diversity of habitats—leaf surfaces, air, marine water, beetle guts and a sea sponge [76,125,128,140,141,142,143,144,145]. Our two isolates were previously reported as S. cf. coprosma [140]. These isolates, HU9059 and HU9256, are most similar (99.48% identity) to the ex-type sequence of S. coprosmae (Hamam. & Nakase) Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout (GenBank acc. no. NR_073317 [32]) but seem to represent a distinct, undescribed species based on the phylogenetic reconstruction of an ITS dataset (Figure 5B). Formal description of this species is pending the characterization of morphology and physiology [140,146].
4.11. Vishniacozyma
In our dataset, we obtained 27 isolates of Vishniacozyma (Tremellomycetes, Tremellales, Bulleribasidiaceae) from 18 lettuce samples, of which nine isolates were obtained from eight conventional lettuce samples and 18 from ten organic ones. Liu et al. [33], in their phylogenetic classification of Tremellomycetes, introduced Vishniacozyma to accommodate eleven species previously known as members of Bullera Derx and Cryptococcus. Currently, the genus Vishniacozyma contains sixteen species [33,76,147,148]. Species in the genus are isolated from different sources—including flowers and plant leaves, sediments and surface sediments of a retreating glacier and soil. According to Li et al. [76], 9 or more nucleotide differences (1.9% mismatches) among ITS sequences can be considered as indication for different taxa. A total of five species were represented in our dataset (Figure 5C, Appendix A, Table A1): Vishniacozyma carnescens (Verona & Luchetti) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout (three isolates); V. cf. foliicola Q.M. Wang & F.Y. Bai ex Yurkov (6 isolates); V. aff. heimaeyensis Vishniac ex Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout sp. nov. (one isolate); V. tephrensis Vishniac ex Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout (one isolate); and V. victoriae (M.J. Montes, Belloch, Galiana, M.D. García, C. Andrés, S. Ferrer, Torr.-Rodr. & J. Guinea) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout (16 isolates). Isolate HU9248 had seven mismatches with V. heimaeyensis in its ITS, which indicated that it represented a different species, which was confirmed by the evolutionary distance among both isolates revealed from the ITS phylogenetic analysis (Figure 5C). Our ITS sequences of C_0038 and HU9045 were 98.67–98.86 identical to the ex-type sequence of V. foliicola, marking 6–7 mismatches. The placement of these isolates in our ITS tree had no support, which is why we marked them as V. cf. foliicola, pending detailed study of morphological and physiological characteristics.
5. Conclusions
Lettuce leaves are nearly always consumed raw rather than cooked. As a result, the microbes that colonize the lettuce phylloplane are mostly ingested by the consumer. Despite this, our knowledge about natural fungal species associated with the phylloplane of romaine lettuce is still very limited, yet this knowledge is necessary for understanding how this naturally occurring flora interacts with introduced human pathogens on the phylloplane. Thus, characterizing these microbial communities is crucial from a public health point of view. In addition, the lettuce industry in the US is worth 6 billion USD annually [5], which adds a not to be underestimated economic factor to the balance. Both factors have led to an interest in the native mycota on the romaine lettuce phylloplane. Here, we characterized the culturable fungal microorganisms on the leaf surface of romaine lettuce using a culture-dependent study combined with molecular identification based on the fungal barcode [149]. We obtained 331 isolates from 42 romaine lettuce samples and generated barcode sequences (ITS) for 249 isolates, representing 59 species. Of these, 12 are undescribed species, all yeasts. This is no surprise; Kachalkin et al. [85] reported that “it becomes challenging for researchers to maintain an overview of the ever-growing diversity of undescribed yeasts”, because countless biodiversity studies report undescribed yeast species. Yeasts are isolated from all around the world and from a plethora of habitats and substrates [150]. Especially the phylloplane represent a popular substrate for (asymptomatic, presumably, saprobic) yeasts. Indeed, plant leaf surfaces represent one of the largest terrestrial microbial habitats; the estimated total surface area available for colonization is 200–640 million km2 [151,152]. Compared to filamentous fungi, yeasts appear to colonize plant leaf surfaces more actively [86,153].
The core fungal community on romaine lettuce includes 38 species in 11 genera (Alternaria, Aureobasidium, Cladosporium, Filobasidium, Naganishia, Papiliotrema, Rhodotorula, Sampaiozyma, Sporobolomyces, Symmetrospora and Vishniacozyma). Within this core community, the yeast-like taxa dominate (with 64.8% of isolates). Interestingly, management treatment (conventional vs. organic) has no effect on the number of fungal isolates (t(20) = 0.2473, p = 0.8072), nor on the number of species (t(20) = 0.8051, p = 0.4302). However, we emphasize that these results may be biased due to the nature of this study; we may not have cultured every species that was effectively present on lettuce leaves (due to selective media, slow-growing species, etc.). Treatments potentially driving changes in fungal communities are best studied using a high-throughput sequencing approach (D. Haelewaters and M.C. Aime, in preparation).
Based on over a decade of fieldwork in understudied habitats and regions, Urbina and Aime [109] estimated that a meager 16% of red yeast species in Sporidiobolales have been described to date. For one species of Sporidiobolales, an undescribed species of Sporobolomyces, we recovered 66 isolates from 27 romaine lettuce samples. This species represented 26.5% of our total isolates and 31.0% of our “core” culture-based community. An earlier study also reported Sporobolomyces to be a dominant portion of the lettuce phylloplane, with 35.5% to 63.0% representation of the total community depending on the cultivar [18]. It is important to formally characterize this undescribed species, which is presumably often consumed with romaine lettuce. Provided its dominance, this species should be considered a candidate for experimental work assessing interactions with human pathogens. To mitigate future outbreaks, it is crucial to understand the ability of E. coli to enter and persist in these phylloplane fungal communities as part of an outbreak control strategy.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/jof7040277/s1, Table S1: GenBank accession numbers of ex-type strains used in core community datasets.
Author Contributions
Conceptualization, D.H., H.U. and M.C.A.; methodology, D.H., H.U., S.B. and S.N.-H.; formal analysis, D.H.; data curation, D.H.; writing—original draft preparation, D.H.; writing—review and editing, D.H. and M.C.A.; visualization, D.H.; supervision, M.C.A.; funding acquisition, M.C.A. All authors have read and agreed to the published version of the manuscript.
Funding
D.H. and H.U. were supported by the Center for Food Safety Engineering at Purdue University funded by the US Department of Agriculture (USDA), Agricultural Research Service, under project 8072-42000-077-00D. This work was supported in part by the USDA National Institute of Food and Agriculture Hatch project 1010662 to M.C.A.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Results from BLAST searches of contigs (with multiple identical isolates) or individual isolates in tabular format. For each contig or isolate, the following are shown: species name, number of isolates (total and by treatment: C, conventional; O, organic; H, hydroponic) and best match information (species name, strain information, GenBank accession number, type status, percentage of similarity and reference in which the sequence was first published). Query coverage was between 75 and 100% for all BLAST searches.
Table A1.
Results from BLAST searches of contigs (with multiple identical isolates) or individual isolates in tabular format. For each contig or isolate, the following are shown: species name, number of isolates (total and by treatment: C, conventional; O, organic; H, hydroponic) and best match information (species name, strain information, GenBank accession number, type status, percentage of similarity and reference in which the sequence was first published). Query coverage was between 75 and 100% for all BLAST searches.
| Contig or Isolate | Species | Number of Isolates | Best Match | Strain | Sequence | Similarity [%] | Reference | |||
|---|---|---|---|---|---|---|---|---|---|---|
| All | C | O | H | |||||||
| C_0008 | Sporobolomyces sp. nov. 2 | 66 | 36 | 30 | Sporobolomyces roseus | CBS:7683T | KY105472 | 98.84 | [20] | |
| C_0005 | Cladosporium sp. 1 | 33 | 12 | 20 | 1 | Cladosporium xylophilum (a) | CBS:125997T | MH863875 | 100.00 | [20] |
| C_0016 | Filobasidium stepposum | 23 | 10 | 13 | Filobasidium stepposum | CBS:10265T | NR_111207 | 99.83 | [32] | |
| C_0010, HU9126, HU9242 | Vishniacozyma victoriae | 16 | 3 | 13 | Vishniacozyma victoriae | ATCC:MYA-305T | MH809977 | 99.81–100.00 | R.E. Rush et al. unpubl. | |
| C_0063 | Cystofilobasidium aff. macerans sp. nov. 1 | 11 | 7 | 4 | Cystofilobasidium macerans | CBS:10757T | KY103183 | 98.24 | [20] | |
| C_0009 | Cladosporium sp. 5 | 10 | 2 | 8 | Cladosporium iridis (b) | CBS:138.40T | EU167591 | 100.00 | [154] | |
| C_0070, HU9199 | Aureobasidium pullulans | 7 | 4 | 3 | Aureobasidium pullulans (c) | CBS 584.75T | KT693733 | 100.00 | [53] | |
| C_0038, HU9045 | Vishniacozyma cf. foliicola | 6 | 3 | 3 | Vishniacozyma foliicola | CBS:9920T | KY105821 | 98.67–98.86 | [20] | |
| C_0014 | Cladosporium sp. 3 | 5 | 3 | 1 | 1 | Cladosporium puyae (d) | CBS:274.80AT | NR_152298 | 100.00 | [69] |
| C_0053, HU9127 | Filobasidium magnum | 5 | 2 | 3 | Filobasidium magnum | CBS:140T | NR_130655 | 99.67–100.00 | [34] | |
| C_0006 | Cladosporium sp. 2 | 4 | 2 | 2 | Cladosporium limoniforme (d) | CBS:140484T | KT600397 | 100.00 | [69] | |
| C_0104 | Aureobasidium subglaciale | 3 | 2 | 1 | Aureobasidium subglaciale | CBS:123387T | KT693735 | 100.00 | [53] | |
| C_0009b | Cladosporium sp. 4 | 3 | 3 | Cladosporium rhusicola (d) | CBS:138.40T | NR_152299 | 100.00 | [69] | ||
| C_0003 | Gibellulopsis nigrescens | 3 | 2 | 1 | Gibellulopsis nigrescens | CBS:120949T | NR_149327 | 100.00 | [155] | |
| C_0001, HU9254 | Vishniacozyma carnescens | 3 | 2 | 1 | Vishniacozyma carnescens | CBS:973T | KY105817 | 99.84–100.00 | [20] | |
| C_0224 | Cladosporium sp. 7 | 2 | 2 | Cladosporium endophyticum | MFLUCC:17-0599T | MG646956 | 99.11 | [156] | ||
| C_0071 | Filobasidium oeirense | 2 | 1 | 1 | Filobasidium oeirense | CBS:8681T | KY103438 | 99.37 | [20] | |
| C_0215 | Metschnikowia picachoensis | 2 | 2 | Metschnikowia picachoensis | NRRL:Y-27607T | AY494780 | 100.00 | [157] | ||
| C_0205 | Papiliotrema aff. fonsecae sp. 1 | 2 | 2 | Papiliotrema fonsecae | EX:F-4087T | NR_119972 | 99.76 | [32] | ||
| C_0057 | Penicillium flavigenum | 2 | 1 | 1 | Penicillium flavigenum | CBS:419.89T | MH862182 | 100.00 | [158] | |
| C_0045 | Stemphylium sp. 1 | 2 | 2 | Stemphylium waikerieanum | VPRI:21969T | MK336832 | 100.00 | [70] | ||
| C_0048 | Symmetrospora sp. nov. | 2 | 2 | Symmetrospora coprosmae | CBS:7899T | NR_073317 | 99.48 | [101] | ||
| HU9265 | Acanthophysium sp. 1 | 1 | 1 | Acanthophysium bisporum | CBS:240.86T | MH861954 | 90.02 | [158] | ||
| HU9315 | Alternaria sp. 1 | 1 | 1 | Alternaria angustiovoidea (e) | CBS:195.86T | MH861939 | 100.00 | [158] | ||
| HU9181 | Alternaria sp. 2 | 1 | 1 | Alternaria ventricosa (e) | CBS:121546T | MH863116 | 100.00 | [158] | ||
| HU9087 | Alternaria sp. 3 | 1 | 1 | Alternaria dactylidicola | MFLUCC:15-0466T | NR_151852 | 99.01 | [159] | ||
| HU9178 | Alternaria sp. 4 | 1 | 1 | Alternaria conjuncta | CBS 196.86T | MH861940 | 99.67 | [158] | ||
| HU9194 | Alternaria sp. 5 | 1 | 1 | Alternaria eureka | CBS 193.86T | MH861937 | 98.78 | [158] | ||
| HU9226 | Alternaria sp. 6 | 1 | 1 | Alternaria arbusti (e) | CBS 596.93T | MH862447 | 100.00 | [158] | ||
| HU9303 | Beauveria cf. bassiana | 1 | 1 | Beauveria bassiana | ARSEF:1564T | NR_111594 | 98.91 | [32] | ||
| HU9220 | Botryotinia pelargonii | 1 | 1 | Botryotinia pelargonii | CBS:497.50T | AJ716290 | 100.00 | [160] | ||
| HU9201 | Bullera aff. unica sp. 1 | 1 | 1 | Bullera unica | CBS:8290T | NR_073256 | 99.62 | [32] | ||
| HU9228 | Bullera unica | 1 | 1 | Bullera unica | CBS:8290T | NR_073256 | 100.00 | [32] | ||
| HU9149 | Bulleromyces albus | 1 | 1 | Bulleromyces albus | CBS:500T | KY101811 | 100.00 | [20] | ||
| HU9304 | Cladosporium sp. 6 | 1 | 1 | Cladosporium puyae | CBS:274.80AT | NR_152298 | 99.82 | [69] | ||
| HU9302 | Cladosporium sphaerospermum | 1 | 1 | Cladosporium sphaerospermum | ATCC:11289T | AY361958 | 100.00 | [161] | ||
| HU9177 | Papiliotrema frias | 1 | 1 | Cryptococcus frias | CRUB 1250T | GU997162 | 100.00 | [89] | ||
| HU9106 | Cystofilobasidium infirmominiatum | 1 | 1 | Cystofilobasidium infirmominiatum | CBS:323T | NR_073232 | 100.00 | [32] | ||
| HU9025 | Epicoccum nigrum complex sp. 1 | 1 | 1 | Epicoccum layuense | CGMCC: 3.18362T | NR_158265 | 100.00 | [162] | ||
| HU9229 | Filobasidium aff. floriforme sp. 1 | 1 | 1 | Filobasidium floriforme | CBS:6241T | NR_119429 | 99.06 | [32] | ||
| HU9055 | Filobasidium chernovii | 1 | 1 | Filobasidium chernovii | CBS:8679T | KY103413 | 100.00 | [20] | ||
| HU9224 | Filobasidium wieringae | 1 | 1 | Filobasidium wieringae | CBS:1937T | NR_077105 | 100.00 | [20] | ||
| HU9215 | Holtermanniella sp. nov. 1 | 1 | 1 | Holtermanniella takashimae | CBS:11174T | NR_137721 | 99.42 | [163] | ||
| HU9066 | Leucosporidium yakuticum | 1 | 1 | Leucosporidium yakuticum | CBS:8621T | NR_155332 | 100.00 | [20] | ||
| HU9283 | Metschnikowia cf. rubicola | 1 | 1 | Metschnikowia rubicola | NRRL:Y-6064T | MG050888 | 99.64 | [164] | ||
| HU9309 | Moesziomyces aphidis | 1 | 1 | Moesziomyces aphidis | CBS:517.83T | NR_145336 | 100.00 | [165] | ||
| HU9011 | Mucor circinelloides f. circinelloides | 1 | 1 | Mucor circinelloides f. circinelloides | CBS:195.68T | HQ154604 | 99.12 | [166] | ||
| HU9043 | Naganishia sp. | 1 | 1 | Naganishia liquefaciens (f) | CBS:968T | NR_073220 | 100.00 | [32] | ||
| HU9061 | Ophiostoma sp. 1 | 1 | 1 | Ophiostoma rufum (g) | CBS:144871T | MH837040 | 100.00 | [167] | ||
| HU9060 | Penicillium sp. 1 | 1 | 1 | Penicillium hoeksii | CBS:137776T | NR_137913 | 99.63 | [168] | ||
| HU9099 | Rhodotorula aff. glutinis sp. nov. 1 | 1 | 1 | Rhodotorula babjevae | CBS:7808T | NR_077096 | 99.66 | [101] | ||
| HU9231 | Rhodotorula aff. glutinis sp. nov. 2 | 1 | 1 | Rhodotorula babjevae | CBS:7808T | NR_077096 | 99.49 | [101] | ||
| HU9305 | Rhodotorula sp. nov. 3 | 1 | 1 | Rhodotorula diobovata | CBS:6085T | KY104768 | 99.51 | [20] | ||
| HU9046 | Sampaiozyma sp. nov. 1 | 1 | 1 | Sampaiozyma ingeniosa | CBS:4240T | NR_111080 | 97.49 | [32] | ||
| HU9301 | Sampaiozyma sp. nov. 2 | 1 | 1 | Sampaiozyma vanillica | CBS:7404T | KY105310 | 93.94 | [20] | ||
| HU9138 | Sporobolomyces sp. nov. 1 | 1 | 1 | Sporobolomyces patagonicus | CBS:9657T | KY105521 | 98.82 | [20] | ||
| HU9240 | Tilletiopsis aff. pallescens sp. nov. 1 | 1 | 1 | Tilletiopsis pallescens | CBS 606.83T | NR_111216 | 98.49 | [32] | ||
| HU9248 | Vishniacozyma aff. heimaeyensis sp. nov. | 1 | 1 | Vishniacozyma heimaeyensis | CBS:8933T | NR_077070 | 98.63 | [32] | ||
| HU9122 | Vishniacozyma tephrensis | 1 | 1 | Vishniacozyma tephrensis | CBS:8935T | NR_144812 | 100.00 | [169] | ||
Footnotes: (a) Ex-type sequences of 18 Cladosporium species (C. cladosporioides complex) with 100% similarity. (b) Ex-type sequences of 16 Cladosporium species (C. herbarum complex) with 100% similarity. (c) Ex-type sequence of Aureobasidium proteae (GenBank acc. no. KT693732, CBS:111973 [53]) is also 100% similar. (d) Ex-type sequences of 2–3 Cladosporium species (C. herbarum complex) with 100% similarity. (e) Ex-type sequences of 2 Alternaria species with 100% similarity. (f) Ex-type sequence of Naganishia albidosimilis (GenBank acc. no. NR_077113, JCM:8843 [32]) is also 100% similar. (g) Ex-type sequences of 8 Ophiostoma species with 100% similarity.
References
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage, 3rd ed.; Springer: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Kibe, E.N. Occurrence of Mycotoxigenic Fungi in Maize from Food Commodity Markets in Kenya. Master’s Thesis, Ghent University, Ghent, Belgium, 2015. [Google Scholar]
- Marasas, W.F.O. Fumonisins: History, world-wide occurrence and impact. In Fumonisins in Food; Jackson, L.S., DeVries, J.W., Bullerman, L.B., Eds.; Springer: New York, NY, USA, 1996; pp. 1–17. [Google Scholar] [CrossRef]
- Shephard, G.S. Risk assessment of aflatoxins in food in Africa. Food Addit. Contam. 2008, 25, 1246–1256. [Google Scholar] [CrossRef]
- USDA (United States Department of Agriculture). 2017 Census of Agriculture. Summary and State Data. Vol. 1. Geogr. Area Ser. 2019, 51, 1–711. [Google Scholar]
- Sivapalasingam, S.; Friedman, C.R.; Cohen, L.; Tauxe, R.V. Fresh produce: A growing cause of outbreaks of foodborne illness in the United States, 1973 through 1997. J. Food Prot. 2004, 67, 2342–2353. [Google Scholar] [CrossRef] [PubMed]
- Brandl, M.T. Fitness of human enteric pathogens on plants and implications for food safety. Annu. Rev. Phytopathol. 2006, 44, 367–392. [Google Scholar] [CrossRef] [Green Version]
- Rangel, J.M.; Sparling, P.H.; Crowe, C.; Griffin, P.M.; Swerdlow, D.L. Epidemiology of Escherichia coli O157: H7 outbreaks, united states, 1982–2002. Emerg. Infect. Dis. 2005, 11, 603. [Google Scholar] [CrossRef] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef]
- Slayton, R.B.; Turabelidze, G.; Bennett, S.D.; Schwensohn, C.A.; Yaffee, A.Q.; Khan, F.; Butler, C.; Trees, E.; Ayers, T.L.; Davis, M.L.; et al. Outbreak of shiga toxin-producing Escherichia coli (STEC) O157:H7 associated with romaine lettuce consumption, 2011. PLoS ONE 2013, 8, e55300. [Google Scholar] [CrossRef] [Green Version]
- Jeamsripong, S.; Chase, J.A.; Jay-Russell, M.T.; Buchanan, R.L.; Atwill, E.R. Experimental in-field transfer and survival of Escherichia coli from animal feces to romaine lettuce in Salinas Valley, California. Microorganisms 2019, 7, 408. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Mark Ibekwe, A.; Crowley, D.E.; Yang, C.-H. Persistence of Escherichia coli O157 and non-O157 strains in agricultural soils. Sci. Total Environ. 2014, 490, 822–829. [Google Scholar] [CrossRef]
- Sai, C.B.; Srinivasan, N.; Zachariah, J.K.; Dananjeyan, B. Experimentation on artificial inoculation studies for persistence of shiga-like toxin-producing Escherichia coli (E. coli O157) in agricultural soils and vegetables using real-time PCR. J. Food Biochem. 2019, 43, e13035. [Google Scholar] [CrossRef]
- CDC (Centers for Disease Control and Prevention). Multistate Outbreak of E. coli O157:H7 Infections Linked to Romaine Lettuce (Final Update). Available online: https://www.cdc.gov/ecoli/2018/o157h7-04-18/index.html (accessed on 7 March 2021).
- Albu, S.; Toome, M.; Aime, M.C. Violaceomyces palustris gen. et sp. nov. and a new monotypic lineage, Violaceomycetales ord. nov. in Ustilaginomycetes. Mycologia 2015, 107, 1193–1204. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Hunter, P.J.; Pink, D.A.; Bending, G.D. Cultivar-level genotype differences influence diversity and composition of lettuce (Lactuca sp.) phyllosphere fungal communities. Fungal Ecol. 2015, 17, 183–186. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [Green Version]
- Vu, D.; Groenewald, M.; Szöke, S.; Cardinali, G.; Eberhardt, U.; Stielow, B.; de Vries, M.; Verkleij, G.J.M.; Crous, P.W.; Boekhout, T.; et al. DNA barcoding analysis of more than 9000 yeast isolates contributes to quantitative thresholds for yeast species and genera delimitation. Stud. Mycol. 2016, 85, 91–105. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; Institute of Electrical and Electronics Engineers: Piscataway, NJ, USA, 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Baral, H.-O.; Weber, E.; Marson, G.; Quijada, L. A new connection between wood saprobism and beetle endosymbiosis: The rarely reported saprobic discomycete Tromeropsis is congeneric with the symbiotic yeast Symbiotaphrina (Symbiotaphrinales, Xylonomycetes) and two asexual morphs misplaced in Hyphozyma. Mycol. Prog. 2018, 17, 215–254. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2017, 35, 518–522. [Google Scholar] [CrossRef]
- Bills, G.F.; González Menéndez, V.; Platas, G. Kabatiella bupleuri sp. nov. (Dothideales), a pleomorphic epiphyte and endophyte of the Mediterranean plant Bupleurum gibraltarium (Apiaceae). Mycologia 2012, 104, 962–973. [Google Scholar] [CrossRef] [PubMed]
- Humphries, Z.; Seifert, K.A.; Hirooka, Y.; Visagie, C.M. A new family and genus in Dothideales for Aureobasidium-like species isolated from house dust. IMA Fungus 2017, 8, 299–315. [Google Scholar] [CrossRef] [Green Version]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T. The Yeasts: A Taxonomic Study, 5th ed.; Elsevier: London, UK, 2011; Volume 3. [Google Scholar]
- Wang, Q.-M.; Yurkov, A.M.; Göker, M.; Lumbsch, H.T.; Leavitt, S.D.; Groenewald, M.; Theelen, B.; Liu, X.-Z.; Boekhout, T.; Bai, F.-Y. Phylogenetic classification of yeasts and related taxa within Pucciniomycotina. Stud. Mycol. 2015, 81, 149–189. [Google Scholar] [CrossRef] [Green Version]
- Schoch, C.L.; Robbertse, B.; Robert, V.; Vu, D.; Cardinali, G.; Irinyi, L.; Meyer, W.; Nilsson, R.H.; Hughes, K.; Miller, A.N.; et al. Finding needles in haystacks: Linking scientific names, reference specimens and molecular data for Fungi. Database 2014, 2014, bau061. [Google Scholar] [CrossRef]
- Liu, X.-Z.; Wang, Q.-M.; Göker, M.; Groenewald, M.; Kachalkin, A.V.; Lumbsch, H.T.; Millanes, A.M.; Wedin, M.; Yurkov, A.M.; Boekhout, T.; et al. Towards an integrated phylogenetic classification of the Tremellomycetes. Stud. Mycol. 2015, 81, 85–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fell, J.W.; Boekhout, T.; Fonseca, A.; Scorzetti, G.; Statzell-Tallman, A. Biodiversity and systematics of basidiomycetous yeasts as determined by large-subunit rDNA D1/ D2 domain sequence analysis. Int. J. Syst. Evol. Microbiol. 2000, 50, 1351–1371. [Google Scholar] [CrossRef]
- Index Fungorum. Search Index Fungorum. Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 5 March 2021).
- Thompson, I.P.; Bailey, M.J.; Fenlon, J.S.; Fermor, T.R.; Lilley, A.K.; Lynch, J.M.; McCormack, P.J.; McQuilken, M.P.; Purdy, K.J.; Rainey, P.B.; et al. Quantitative and qualitative seasonal changes in the microbial community from the phyllosphere of sugar-beet (Beta vulgaris). Plant Soil 1993, 150, 177–191. [Google Scholar] [CrossRef]
- McCormack, P.J.; Wildman, H.G.; Jeffries, P. Production of antibacterial compounds by phylloplane-inhabiting yeasts and yeastlike fungi. Appl. Environ. Microbiol. 1994, 60, 927–931. [Google Scholar] [CrossRef] [Green Version]
- Andrews, J.H.; Harris, R.F. The ecology and biogeography of microorganisms on plant surfaces. Ann. Rev. Phytopathol. 2000, 38, 145–180. [Google Scholar] [CrossRef] [PubMed]
- Whipps, J.M.; Hand, P.; Pink, D.A.C.; Bending, G.D. Phyllosphere microbiology with special reference to diversity and plant genotype. J. Appl. Microbiol. 2008, 104, 1744–1755. [Google Scholar] [CrossRef] [Green Version]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Barth, M.; Hankinson, T.R.; Zhuang, H.; Breidt, F. Microbiological spoilage of fruits and vegetables. In Compendium of the Microbiological Spoilage of Foods and Beverages; Sperber, W., Doyle, M., Eds.; Springer: New York, NY, USA, 2009; pp. 135–183. [Google Scholar] [CrossRef]
- Pryor, B.; Michailides, T. Morphological, pathogenic, and molecular characterization of Alternaria isolates associated with alternaria late blight of pistachio. Phytopathology 2002, 92, 406–416. [Google Scholar] [CrossRef] [Green Version]
- Badotti, F.; de Oliveira, F.S.; Garcia, C.F.; Vaz, A.B.M.; Fonseca, P.L.C.; Nahum, L.A.; Oliveira, G.; Góes-Neto, A. Effectiveness of ITS and sub-regions as DNA barcode markers for the identification of Basidiomycota (Fungi). BMC Microbiol. 2017, 17, 42. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, D.P.; Gannibal, P.B.; Peever, T.L.; Pryor, B.M. The sections of Alternaria: Formalizing species-group concepts. Mycologia 2013, 105, 530–546. [Google Scholar] [CrossRef] [Green Version]
- Pryor, B.M.; Bigelow, D.M. Molecular characterization of Embellisia and Nimbya species and their relationship to Alternaria, Ulocladium and Stemphylium. Mycologia 2003, 95, 1141–1154. [Google Scholar] [CrossRef]
- Joly, P. Le genre Alternaria: Recherches Physiologiques, Biologiques et Systématiques; Lechevalier: Paris, France, 1964. [Google Scholar]
- Strandberg, J.O. Alternaria species that attack vegetable crops: Biology and options for diseases management. In Alternaria Biology, Plant Diseases and Metabolites; Chelkowski, J., Visconti, A., Eds.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1992; pp. 175–208. [Google Scholar]
- Boedo, C.; Benichou, S.; Berruyer, R.; Bersihand, S.; Dongo, A.; Simoneau, P.; Lecomte, M.; Briard, M.; Le Clerc, V.; Poupard, P. Evaluating aggressiveness and host range of Alternaria dauci in a controlled environment. Plant Pathol. 2012, 61, 63–75. [Google Scholar] [CrossRef]
- Anita, D.D.; Sridhar, K.R.; Bhat, R. Diversity of fungi associated with mangrove legume Sesbania bispinosa (Jacq.) W. Wight (Fabaceae). Livest. Res. Rural Dev. 2009, 21, 67. [Google Scholar]
- Varvas, T.; Kullman, B. First records of two ascomycetes on Phleum pratense in Estonia. Folia Cryptogam. Estonica 2012, 49, 73–76. [Google Scholar]
- Tymon, L.S.; Peever, T.L.; Johnson, D.A. Identification and enumeration of small-spored Alternaria species associated with potato in the US Northwest. Plant Dis. 2016, 100, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poursafar, A.; Ghosta, Y.; Javan-Nikkhah, M. Identification of Alternaria species from the section Infectoriae associated with wheat and barley black (sooty) head mold in Iran. Taxon. Biosyst. 2017, 9, 13–30. [Google Scholar]
- van Nieuwenhuijzen, E.J.; Houbraken, J.A.; Meijer, M.; Adan, O.C.; Samson, R.A. Aureobasidium melanogenum: A native of dark biofinishes on oil treated wood. Ant. Van Leeuwenhoek 2016, 109, 661–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Species Fungorum. Search Species Fungorum. Available online: http://www.speciesfungorum.org/Names/Names.asp (accessed on 5 March 2021).
- Webb, T.A.; Mundt, J.O. Molds on vegetables at the time of harvest. Appl. Environ. Microbiol. 1978, 35, 655–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, R.R.; Singh, D.; Singh, R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: A review. Biol. Control 2009, 50, 205–221. [Google Scholar] [CrossRef]
- Wilson, C.L.; Chalutz, E. Postharvest biocontrol of Penicillium rots of citrus with antagonistic yeasts and bacteria. Sci. Hortic. 1989, 40, 105–112. [Google Scholar] [CrossRef]
- Bencheqroun, S.K.; Baji, M.; Massart, S.; Labhilili, M.; El Jaafari, S.; Jijaki, M.H. In vitro and in situ study of postharvest apple blue mold biocontrol by Aureobasidium pullulans: Evidence for the involvement of competition for nutrients. Postharvest Biol. Technol. 2007, 46, 128–135. [Google Scholar] [CrossRef]
- Seibold, A.; Fried, A.; Kunz, S.; Moltmann, E.; Lange, E.; Jelkmann, W. Yeasts as antagonists against fireblight. EPPO Bull. 2004, 34, 389–390. [Google Scholar] [CrossRef]
- Gostinčar, C.; Ohm, R.A.; Kogej, T.; Sonjak, S.; Turk, M.; Zajc, J.; Zalar, P.; Grube, M.; Sun, H.; Han, J.; et al. Genome sequencing of four Aureobasidium pullulans varieties: Biotechnological potential, stress tolerance, and description of new species. BMC Genom. 2014, 15, 549. [Google Scholar] [CrossRef] [Green Version]
- Fradkin, A.; Tobin, R.S.; Tarlo, S.M.; Tucic-Porretta, M.; Malloch, D. Species identification of airborne molds and Its significance for the detection of indoor pollution. Int. J. Air Pollut. Control Hazard. Waste Manag. 1987, 37, 51–53. [Google Scholar] [CrossRef]
- Flannigan, B. Microorganisms in indoor air. In Microorganisms in Home and Indoor Work Environments: Diversity, Health Impacts, Investigation and Control, 2nd ed.; Flannigan, B., Samson, R., Miller, D., Eds.; CRC Press: Boca Raton, FL, USA, 2001; pp. 17–31. [Google Scholar] [CrossRef]
- Horner, W.E.; Worthan, A.G.; Morey, P.R. Air- and dustborne mycoflora in houses free of water damage and fungal growth. Appl. Environ. Microbiol. 2004, 70, 6394–6400. [Google Scholar] [CrossRef] [Green Version]
- Visagie, C.M.; Hirooka, Y.; Tanney, J.B.; Whitfield, E.; Mwange, K.; Meijer, M.; Amend, A.S.; Seifert, K.A.; Samson, R.A. Aspergillus, Penicillium and Talaromyces isolated from house dust samples collected around the world. Stud. Mycol. 2014, 78, 63–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bensch, K.; Groenewald, J.Z.; Meijer, M.; Dijksterhuis, J.; Jurjević, Ž.; Andersen, B.; Houbraken, J.; Crous, P.W.; Samson, R.A. Cladosporium species in indoor environments. Stud. Mycol. 2018, 89, 177–301. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Denis, M.; Gené, J.; Sutton, D.A.; Wiederhold, N.P.; Cano-Lira, J.F.; Guarro, J. New species of Cladosporium associated with human and animal infections. Persoonia 2016, 36, 281–298. [Google Scholar] [CrossRef] [Green Version]
- Sandoval-Denis, M.; Sutton, D.A.; Martin-Vicente, A.; Cano-Lira, J.F.; Wiederhold, N.; Guarro, J.; Gené, J. Cladosporium species recovered from clinical samples in the United States. J. Clin. Microbiol. 2015, 53, 2990–3000. [Google Scholar] [CrossRef] [Green Version]
- Bensch, K.; Braun, U.; Groenewald, J.Z.; Crous, P.W. The genus Cladosporium. Stud. Mycol. 2012, 72, 1–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bensch, K.; Groenewald, J.Z.; Braun, U.; Dijksterhuis, J.; de Jesús Yáñez-Morales, M.; Crous, P.W. Common but different: The expanding realm of Cladosporium. Stud. Mycol. 2015, 82, 23–74. [Google Scholar] [CrossRef] [Green Version]
- Marin-Felix, Y.; Hernández-Restrepo, M.; Iturrieta-González, I.; García, D.; Gené, J.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Quaedvlieg, W.; Schumacher, R.K.; et al. Genera of phytopathogenic fungi: GOPHY 3. Stud. Mycol. 2019, 94, 1–124. [Google Scholar] [CrossRef]
- Bensch, K.; Groenewald, J.Z.; Dijksterhuis, J.; Starink-Willemse, M.; Andersen, B.; Summerell, B.A.; Shin, H.D.; Dugan, F.M.; Schroers, H.J.; Braun, U.; et al. Species and ecological diversity within the Cladosporium cladosporioides complex (Davidiellaceae, Capnodiales). Stud. Mycol. 2010, 67, 1–94. [Google Scholar] [CrossRef]
- Lavoie, K.; Touchette, M.; St-Gelais, D.; Labrie, S. Characterization of the fungal microflora in raw milk and specialty cheeses of province of Quebec. Dairy Sci. Technol. 2012, 92, 455–468. [Google Scholar] [CrossRef] [Green Version]
- Merín, M.G.; Mendoza, L.M.; Morata de Ambrosini, V.I. Pectinolytic yeasts from viticultural and enological environments: Novel finding of Filobasidium capsuligenum producing pectinases. J. Basic Microbiol. 2014, 54, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Šuranská, H.; Raspor, P.; Uroić, K.; Golić, N.; Kos, B.; Mihajlović, S.; Begović, J.; Šušković, J.; Topisirović, L.; Čadež, N. Characterisation of the yeast and mould biota in traditional white pickled cheeses by culture-dependent and independent molecular techniques. Folia Microbiol. 2016, 61, 455–463. [Google Scholar] [CrossRef]
- Lin, W.R.; PH, W.; Hsieh, S.Y.; Tsai, C.H.; Hsiao, S.C. Yeasts associated with Euploea butterflies. Mycosphere 2018, 9, 149–154. [Google Scholar] [CrossRef]
- Li, A.-H.; Yuan, F.-X.; Groenewald, M.; Bensch, K.; Yurkov, A.M.; Li, K.; Han, P.-J.; Guo, L.-D.; Aime, M.C.; Sampaio, J.P.; et al. Diversity and phylogeny of basidiomycetous yeasts from plant leaves and soil: Proposal of two new orders, three new families, eight new genera and one hundred and seven new species. Stud. Mycol. 2020, 96, 17–140. [Google Scholar] [CrossRef]
- Scupham, A.J.; Presley, L.L.; Wei, B.; Bent, E.; Griffith, N.; McPherson, M.; Zhu, F.; Oluwadara, O.; Rao, N.; Braun, J.; et al. Abundant and diverse fungal microbiota in the murine intestine. Appl. Environ. Microbiol. 2006, 72, 793–801. [Google Scholar] [CrossRef] [Green Version]
- Gouba, N.; Raoult, D.; Drancourt, M. Eukaryote culturomics of the gut reveals new species. PLoS ONE 2014, 9, e106994. [Google Scholar] [CrossRef]
- Mattsson, R.; Haemig, P.D.; Olsen, B. Feral pigeons as carriers of Cryptococcus laurentii, Cryptococcus uniguttulatus and Debaryomyces hansenii. Med. Mycol. 1999, 37, 367–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, W.; Liao, W.; Hagen, F.; Theelen, B.; Shi, W.; Meis, J.F.; Boekhout, T. Meningitis caused by Filobasidium uniguttulatum: Case report and overview of the literature. Mycoses 2012, 55, 105–109. [Google Scholar] [CrossRef]
- Goto, S. On a new yeast genus Naganishia. J. Ferment. Tech. 1963, 41, 459–462. [Google Scholar]
- Fonseca, Á.; Scorzetti, G.; Fell, J.W. Diversity in the yeast Cryptococcus albidus and related species as revealed by ribosomal DNA sequence analysis. Can. J. Microbiol. 2000, 46, 7–27. [Google Scholar] [CrossRef]
- Fotedar, R.; Kolecka, A.; Boekhout, T.; Fell, J.W.; Anand, A.; Al Malaki, A.; Zeyara, A.; Al Marri, M. Naganishia qatarensis sp. nov., a novel basidiomycetous yeast species from a hypersaline marine environment in Qatar. Int. J. Syst. Evol. Microbiol. 2018, 68, 2924–2929. [Google Scholar] [CrossRef]
- Crous, P.W.; Carnegie, A.J.; Wingfield, M.J.; Sharma, R.; Mughini, G.; Noordeloos, M.E.; Santini, A.; Shouche, Y.S.; Bezerra, J.D.P.; Dima, B.; et al. Fungal Planet description sheets: 868–950. Persoonia 2019, 42, 291–473. [Google Scholar] [CrossRef] [PubMed]
- Kachalkin, A.V.; Turchetti, B.; Inácio, J.; Carvalho, C.; Mašínová, T.; Pontes, A.; Röhl, O.; Glushakova, A.M.; Akulov, A.; Baldrian, P.; et al. Rare and undersampled dimorphic basidiomycetes. Mycol. Prog. 2019, 18, 945–971. [Google Scholar] [CrossRef]
- Fonseca, Á.; Inácio, J. Phylloplane yeasts. In Biodiversity and Ecophysiology of Yeasts; Péter, G., Rosa, C., Eds.; Springer: Berlin, Germany, 2006; pp. 263–301. [Google Scholar] [CrossRef]
- Sampaio, J.P.; Weiß, M.; Gadanho, M.; Bauer, R. New taxa in the Tremellales: Bulleribasidium oberjochense gen. et sp. nov., Papiliotrema bandonii gen. et sp. nov. and Fibulobasidium murrhardtense sp. nov. Mycologia 2002, 94, 873–887. [Google Scholar] [CrossRef]
- Crestani, J.; Landell, M.F.; Faganello, J.; Vainstein, M.H.; Vishniac, H.S.; Valente, P. Cryptococcus terrestris sp. nov., a tremellaceous, anamorphic yeast phylogenetically related to Cryptococcus flavescens. Int. J. Syst. Evol. Microbiol. 2009, 59, 631–636. [Google Scholar] [CrossRef] [Green Version]
- De Garcia, V.; Zalar, P.; Brizzio, S.; Gunde-Cimerman, N.; Van Broock, M. Cryptococcus species (Tremellales) from glacial biomes in the southern (Patagonia) and northern (Svalbard) hemispheres. FEMS Microbiol. Ecol. 2012, 82, 523–539. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-Paim, K.; Ferreira, T.B.; Andrade-Silva, L.; Mora, D.J.; Springer, D.J.; Heitman, J.; Fonseca, F.M.; Matos, D.; Melhem, M.S.; Silva-Vergara, M.L. Phylogenetic analysis of phenotypically characterized Cryptococcus laurentii isolates reveals high frequency of cryptic species. PLoS ONE 2014, 9, e108633. [Google Scholar] [CrossRef]
- Surussawadee, J.; Khunnamwong, P.; Srisuk, N.; Limtong, S. Papiliotrema siamense f.a., sp. nov., a yeast species isolated from plant leaves. Int. J. Syst. Evol. Microbiol. 2014, 64, 3058–3062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handel, S.; Wang, T.; Yurkov, A.M.; König, H. Sugiyamaella mastotermitis sp. nov. and Papiliotrema odontotermitis f.a., sp. nov. from the gut of the termites Mastotermes darwiniensis and Odontotermes obesus. Int. J. Syst. Evol. Microbiol. 2016, 66, 4600–4608. [Google Scholar] [CrossRef]
- Pagani, D.M.; Brandao, L.R.; Santos, A.R.O.; Felix, C.R.; Ramos, J.P.; Broetto, L.; Scorzetti, G.; Fell, J.W.; Rosa, C.A.; Valente, P.; et al. Papiliotrema leoncinii sp. nov. and Papiliotrema miconiae sp. nov., two tremellaceous yeast species from Brazil. Int. J. Syst. Evol. Microbiol. 2016, 66, 1799–1806. [Google Scholar] [CrossRef] [Green Version]
- Boekhout, T. A revision of ballistoconidia-forming yeasts and fungi. Stud. Mycol. 1991, 33, 1–194. [Google Scholar]
- Hamamoto, M.; Boekhout, T.; Nakase, T. Chapter 156. Sporobolomyces Kluyver & van Niel (1924). In The Yeasts, a Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; Volume 3, pp. 1929–1990. [Google Scholar] [CrossRef]
- Sampaio, J.P. Chapter 155. Rhodotorula Harrison (1928). In The Yeasts, a Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; Volume 3, pp. 1873–1927. [Google Scholar] [CrossRef]
- Aime, M.C.; Matheny, P.B.; Henk, D.A.; Frieders, E.M.; Nilsson, R.H.; Piepenbring, M.; McLaughlin, D.J.; Szabo, L.J.; Begerow, D.; Sampaio, J.P.; et al. An overview of the higher-level classification of Pucciniomycotina based on combined analyses of nuclear large and small subunit rDNA sequences. Mycologia 2006, 98, 895–905. [Google Scholar] [CrossRef]
- Bauer, R.; Begerow, D.; Sampaio, J.P.; Weiβ, M.; Oberwinkler, F. The simple-septate basidiomycetes: A synopsis. Mycol. Prog. 2006, 5, 41–66. [Google Scholar] [CrossRef]
- Wang, Q.-M.; Groenewald, M.; Takashima, M.; Theelen, B.; Han, P.-J.; Liu, X.-Z.; Boekhout, T.; Bai, F.-Y. Phylogeny of yeasts and related filamentous fungi within Pucciniomycotina determined from multigene sequence analyses. Stud. Mycol. 2015, 81, 27–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadanho, M.; Sampaio, J.P. Polyphasic taxonomy of the basidiomycetous yeast genus Rhodotorula: Rh. glutinis sensu stricto and Rh. dairenensis comb. nov. FEMS Yeast Res. 2002, 2, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Scorzetti, G.; Fell, J.W.; Fonseca, A.; Statzell-Tallman, A. Systematics of basidiomycetous yeasts: A comparison of large subunit D1/D2 and internal transcribed spacer rDNA regions. FEMS Yeast Res. 2002, 2, 495–517. [Google Scholar] [CrossRef]
- Wirth, F.; Goldani, L.Z. Epidemiology of Rhodotorula: An emerging pathogen. Interdiscip. Perspect. Infect. Dis. 2012, 2012, 465–717. [Google Scholar] [CrossRef] [Green Version]
- Gildemacher, P.; Heijne, B.; Silvestri, M.; Houbraken, J.; Hoekstra, E.; Theelen, B.; Boekhout, T. Interactions between yeasts, fungicides and apple fruit russeting. FEMS Yeast Res. 2006, 6, 1149–1156. [Google Scholar] [CrossRef] [Green Version]
- Magnuson, J.A.; King, A.D.; Török, T. Microflora of partially processed lettuce. Appl. Environ. Microbiol. 1990, 56, 3851–3854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Török, T.; King, A.D. Comparative study on the identification of food-borne yeasts. Appl. Environ. Microbiol. 1991, 57, 1207–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Corato, U. Fungal population dynamics in ready-to-eat salads during a shelf-life in Italy. J. Agric. Sci. Technol. A 2012, 2, 569–576. [Google Scholar]
- Yafetto, L.; Ekloh, E.; Sarsah, B.; Amenumey, E.K.; Adator, E.H. Microbiological contamination of some fresh leafy vegetables sold in Cape Coast, Ghana. Ghana J. Sci. 2019, 60, 11–23. [Google Scholar] [CrossRef]
- Khodadad, C.L.; Hummerick, M.E.; Spencer, L.E.; Dixit, A.R.; Richards, J.T.; Romeyn, M.W.; Smith, T.M.; Wheeler, R.M.; Massa, G.D. Microbiological and nutritional analysis of lettuce crops grown on the International Space Station. Front. Plant Sci. 2020, 11, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbina, H.; Aime, M.C. A closer look at Sporidiobolales: Ubiquitous microbial community members of plant and food biospheres. Mycologia 2018, 110, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Tuon, F.F.; Costa, S.F. Rhodotorula infection. A systematic review of 128 cases from literature. Rev. Iberoam. Micol. 2008, 25, 135–140. [Google Scholar] [CrossRef]
- Alvarez-Perez, S.; Mateos, A.; Dominguez, L.; Martinez-Nevado, E.; Blanco, J.L.; Garcia, M.E. Isolation of Rhodotorula mucilaginosa from skin lesions in a Southern sea lion (Otaria flavescens): A case report. Vet. Med. 2010, 55, 297–301. [Google Scholar] [CrossRef] [Green Version]
- Ioannou, P.; Vamvoukaki, R.; Samonis, G. Rhodotorula species infections in humans: A systematic review. Mycoses 2019, 62, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Dì Menna, M.E. Torulopsis ingeniosa n. sp., from grass leaves. J. Gen. Microbiol. 1958, 19, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Sampaio, J.P. Utilization of low molecular weight lignin-related aromatic compounds for the selective isolation of yeasts: Rhodotorula vanillica, a new basidiomycetous yeast species. Syst. Appl. Microbiol. 1995, 17, 613–619. [Google Scholar] [CrossRef]
- Liu, X.-Z.; Groenewald, M.; Boekhout, T.; Bai, F.-Y. Heitmania gen. nov., a new yeast genus in Microbotryomycetes, and description of three novel species: Heitmania litseae sp. nov., Heitmania castanopsis sp. nov. and Heitmania elacocarpi sp. nov. Int. J. Syst. Evol. Microbiol. 2017, 67, 4534–4540. [Google Scholar] [CrossRef] [PubMed]
- Haelewaters, D.; Peterson, R.A.; Nevalainen, H.; Aime, M.C. Inopinatum lactosum gen. & comb. nov., the first pink-pigmented yeast in Leotiomycetes. Int. J. Syst. Evol. Microbiol. 2021. in review. [Google Scholar]
- Lorenzini, M.; Zapparoli, G.; Azzolini, M.; Carvalho, C.; Sampaio, J.P. Sporobolomyces agrorum sp. nov. and Sporobolomyces sucorum sp. nov., two novel basidiomycetous yeast species isolated from grape and apple must in Italy. Int. J. Syst. Evol. Microbiol. 2019, 69, 3385–3391. [Google Scholar] [CrossRef]
- Valadon, L.R.G. Carotenoids as additional taxonomic characters in fungi: A review. Trans. Brit. Mycol. Soc. 1976, 67, 1–15. [Google Scholar] [CrossRef]
- Ratledge, C. Microorganisms for lipids. Acta Biotechnol. 1991, 11, 429–438. [Google Scholar] [CrossRef]
- Davoli, P.; Mierau, V.; Weber, R.W. Carotenoids and fatty acids in red yeasts Sporobolomyces roseus and Rhodotorula glutinis. Appl. Biochem. Microbiol. 2004, 40, 460–465. [Google Scholar] [CrossRef]
- Manimala, M.R.A.; Murugesan, R. In vitro antioxidant and antimicrobial activity of carotenoid pigment extracted from Sporobolomyces sp. isolated from natural source. J. Appl. Nat. Sci. 2014, 6, 649–653. [Google Scholar] [CrossRef]
- Li, C.J.; Zhao, D.; Li, B.X.; Zhang, N.; Yan, J.Y.; Zou, H.T. Whole genome sequencing and comparative genomic analysis of oleaginous red yeast Sporobolomyces pararoseus NGR identifies candidate genes for biotechnological potential and ballistospores-shooting. BMC Genom. 2020, 21, 181. [Google Scholar] [CrossRef]
- Frengova, G.; Beshkova, M. Carotenoids from Rhodotorula and Phaffia: Yeasts of biotechnological importance. J. Ind. Microbiol. Biotechnol. 2009, 36, 163–180. [Google Scholar] [CrossRef] [PubMed]
- Mannazzu, I.; Landolfo, S.; da Silva, T.L.; Buzzini, P. Red yeasts and carotenoid production: Outlining a future for non-conventional yeasts of biotechnological interest. World J. Microbiol. Biotechnol. 2015, 31, 1665–1673. [Google Scholar] [CrossRef] [Green Version]
- Derx, H.G. Etude sur les Sporobolomycetes. Ann. Mycol. 1930, 28, 1–23. [Google Scholar]
- Tubaki, K. Studies on the Sporobolomycetaceae in Japan. III. On Sporobolomyces and Bullera. Nagaoa 1953, 3, 12–21. [Google Scholar]
- Last, F.T. Seasonal incidence of Sporobolomyces on cereal leaves. Trans. Brit. Mycol. Soc. 1955, 38, 221–239. [Google Scholar] [CrossRef]
- Hamamoto, M.; Nakase, T. Ballistosporous yeasts found on the surface of plant materials collected in New Zealand. 1. Six new species in the genus Sporobolomyces. Ant. Van Leeuwenhoek 1995, 67, 151–171. [Google Scholar] [CrossRef]
- Nakase, T. Expanding world of ballistosporous yeasts: Distribution in the phyllosphere, systematics and phylogeny. J. Gen. Appl. Microbiol. 2000, 46, 189–216. [Google Scholar] [CrossRef] [Green Version]
- Molnár, O.; Wuczkowski, M.; Prillinger, H. Yeast biodiversity in the guts of several pests on maize; comparison of three methods: Classical isolation, cloning and DGGE. Mycol. Prog. 2008, 7, 111–123. [Google Scholar] [CrossRef]
- Debode, J.; Van Hemelrijck, W.; Creemers, P.; Maes, M. Effect of fungicides on epiphytic yeasts associated with strawberry. Microbiol. Open 2013, 2, 482–491. [Google Scholar] [CrossRef]
- Janisiewicz, W.J.; Peterson, D.L.; Bors, R.; Sheu, J.J.; Jan, S.P.; Lee, H.T.; Yu, S.M.; Seastedt, T.R.; Coxwell, C.C.; Ojima, D.S.; et al. Control of storage decay of apples with Sporobolomyces roseus. Plant Dis. 1994, 78, 466–470. [Google Scholar] [CrossRef]
- Bashi, E.; Fokkema, N.J. Environmental factors limiting growth of Sporobolomyces roseus, an antagonist of Cochliobolus sativus, on wheat leaves. Trans. Brit. Mycol. Soc. 1977, 68, 17–25. [Google Scholar] [CrossRef]
- Bai, F.-Y.; Zhao, J.-H.; Takashima, M.; Jia, J.-H.; Boekhout, T.; Nakase, T. Reclassification of the Sporobolomyces roseus and Sporidiobolus pararoseus complexes, with the description of Sporobolomyces phaffii sp. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 2309–2314. [Google Scholar] [CrossRef]
- Serradilla, M.J.; del Carmen Villalobos, M.; Hernández, A.; Martín, A.; Lozano, M.; de Guía Córdoba, M. Study of microbiological quality of controlled atmosphere packaged ‘Ambrunés’ sweet cherries and subsequent shelf-life. Int. J. Food Microbiol. 2013, 166, 85–92. [Google Scholar] [CrossRef]
- Bourret, T.B.; Grove, G.G.; Vandemark, G.J.; Henick-Kling, T.; Glawe, D.A. Diversity and molecular determination of wild yeasts in a central Washington State vineyard. N. Am. Fungi 2013, 8, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Barahona, S.; Yuivar, Y.; Socias, G.; Alcaíno, J.; Cifuentes, V.; Baeza, M. Identification and characterization of yeasts isolated from sedimentary rocks of Union Glacier at the Antarctica. Extremophiles 2016, 20, 479–491. [Google Scholar] [CrossRef]
- Nakase, T.; Takematsu, A.; Yamada, Y. Molecular approaches to the taxonomy of ballistosporous yeasts based on the analysis of the partial nucleotide sequences of 18S ribosomal ribonucleic acids. J. Gen. Appl. Microbiol. 1993, 39, 107–134. [Google Scholar] [CrossRef] [Green Version]
- Boekhout, T.; Fonseca, A.; Sampaio, J.P.; Bandoni, R.J.; Fell, J.W.; Kwon-Chung, K.J. Chapter 100. Discussion of teleomorphic and anamorphic basidiomycetous yeasts. In The Yeasts, a Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; Volume 3, pp. 1339–1372. [Google Scholar] [CrossRef]
- Haelewaters, D.; Toome-Heller, M.; Albu, S.; Aime, M.C. Red yeasts from leaf surfaces and other habitats: Three new species and a new combination of Symmetrospora (Pucciniomycotina, Cystobasidiomycetes). Fungal Syst. Evol. 2020, 5, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Phaff, H.J.; Mrak, E.M.; Williams, O.B. Yeasts isolated from shrimp. Mycologia 1952, 44, 431–451. [Google Scholar] [CrossRef]
- Shivas, R.G.; de Miranda, L.R. Two new species of the genus Sporobolomyces and a new Rhodotorula species from leaf surfaces. Ant. Van Leeuwenhoek 1983, 49, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Nakase, T.; Suzuki, M. Bullera intermedia sp. nov. and Sporobolomyces oryzicola sp. nov. isolated from dead leaves of Oryza sativa. J. Gen. Appl. Microbiol. 1986, 32, 149–155. [Google Scholar] [CrossRef]
- Takashima, M.; Nakase, T. Four new species of the genus Sporobolomyces isolated from leaves in Thailand. Mycoscience 2000, 41, 357–369. [Google Scholar] [CrossRef]
- Wang, Q.-M.; Bai, F.-Y. Four new yeast species of the genus Sporobolomyces from plant leaves. FEMS Yeast Res. 2004, 4, 579–586. [Google Scholar] [CrossRef] [Green Version]
- Suh, S.-O.; Zhang, N.; Nguyen, N.; Gross, S.; Blackwell, M. Lab Manual for Yeast Study; Louisiana State University: Baton Rouge, LA, USA, 2008. [Google Scholar]
- Tsuji, M.; Tanabe, Y.; Vincent, W.F.; Uchida, M. Vishniacozyma ellesmerensis sp. nov., a psychrophilic yeast isolated from a retreating glacier in the Canadian High Arctic. Int. J. Syst. Evol. Microbiol. 2019, 69, 696–700. [Google Scholar] [CrossRef]
- Félix, C.R.; Andrade, D.A.; Almeida, J.H.; Navarro, H.M.; Fell, J.W.; Landell, M.F. Vishniacozyma alagoana sp. nov. a tremellomycetes yeast associated with plants from dry and rainfall tropical forests. Int. J. Syst. Evol. Microbiol. 2020, 70, 3449–3454. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Fungal Barcoding Consortium. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Péter, G.; Takashima, M.; Čadež, N. Yeast habitats: Different but global. In Yeasts in Natural Ecosystems: Ecology; Buzzini, P., Lachance, M.A., Yurkov, A., Eds.; Springer: Cham, Switzerland, 2017; pp. 39–71. [Google Scholar] [CrossRef]
- Morris, C.E. Phyllosphere. In Encyclopedia of Life Sciences; Nature Publishing Group: London, UK, 2001. [Google Scholar] [CrossRef]
- Morris, C.E.; Kinkel, L.L. Fifty years of phyllosphere microbiology: Significant contributions to research in related fields. In Phyllosphere Microbiology; Lindow, S.E., Hecht-Poinar, E.I., Elliott, V., Eds.; APS Press: St. Paul, MN, USA, 2002; pp. 365–375. [Google Scholar]
- Lindow, S.E.; Brandl, M.T. Microbiology of the phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef] [Green Version]
- Simon, U.K.; Groenewald, J.Z.; Crous, P.W. Cymadothea trifolii, an obligate biotrophic leaf parasite of Trifolium, belongs to Mycosphaerellaceae as shown by nuclear ribosomal DNA analyses. Persoonia 2009, 22, 49–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zare, R.; Gams, W.; Starink-Willemse, M.; Summerbell, R.C. Gibellulopsis, a suitable genus for Verticillium nigrescens, and Musicillium, a new genus for V. theobromae. Nova Hedwig. 2007, 85, 463–489. [Google Scholar] [CrossRef] [Green Version]
- Tibpromma, S.; Hyde, K.D.; Bhat, J.D.; Mortimer, P.E.; Xu, J.; Promputtha, I.; Doilom, M.; Yang, J.B.; Tang, A.M.; Karunarathna, S.C. Identification of endophytic fungi from leaves of Pandanaceae based on their morphotypes and DNA sequence data from southern Thailand. MycoKeys 2018, 33, 25–67. [Google Scholar] [CrossRef]
- Suh, S.-O.; Gibson, C.M.; Blackwell, M. Metschnikowia chrysoperlae sp. nov., Candida picachoensis sp. nov. and Candida pimensis sp. nov., isolated from the green lacewings Chrysoperla comanche and Chrysoperla carnea (Neuroptera: Chrysopidae). Int. J. Syst. Evol. Microbiol. 2004, 54, 1883–1890. [Google Scholar] [CrossRef] [PubMed]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Wanasinghe, D.N.; Phillips, A.J.L.; Camporesi, E.; Bulgakov, T.S.; Phukhamsakda, C.; Ariyawansa, H.A.; Goonasekara, I.D.; Phookamsak, R.; Dissanayake, A.; et al. Mycosphere notes 1–50: Grass (Poaceae) inhabiting Dothideomycetes. Mycosphere 2017, 8, 697–796. [Google Scholar] [CrossRef]
- Staats, M.; van Baarlen, P.; van Kan, J.A. Molecular phylogeny of the plant pathogenic genus Botrytis and the evolution of host specificity. Mol. Biol. Evol. 2005, 22, 333–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.G.; Managbanag, J.R.; Stamenova, E.K.; Jong, S.C. Comparative analysis of common indoor Cladosporium species based on molecular data and conidial characters. Mycotaxon 2004, 89, 441–451. [Google Scholar]
- Chen, Q.; Hou, L.W.; Duan, W.J.; Crous, P.W.; Cai, L. Didymellaceae revisited. Stud. Mycol. 2017, 87, 105–159. [Google Scholar] [CrossRef] [PubMed]
- Wuczkowski, M.; Passoth, V.; Turchetti, B.; Andersson, A.C.; Olstorpe, M.; Laitila, A.; Theelen, B.; van Broock, M.; Buzzini, P.; Prillinger, H.; et al. Description of Holtermanniella gen. nov., including Holtermanniella takashimae sp. nov. and four new combinations, and proposal of the order Holtermanniales to accommodate tremellomycetous yeasts of the Holtermannia clade. Int. J. Syst. Evol. Microbiol. 2011, 61, 680–689. [Google Scholar] [CrossRef] [Green Version]
- Kurtzman, C.P.; Robnett, C.J.; Basehoar, E.; Ward, T.J. Four new species of Metschnikowia and the transfer of seven Candida species to Metschnikowia and Clavispora as new combinations. Ant. Van Leeuwenhoek 2018, 111, 2017–2035. [Google Scholar] [CrossRef]
- Avis, T.J.; Caron, S.; Boekhout, T.; Hamelin, R.C.; Bélanger, R.R. Molecular and physiological analysis of the powdery mildew antagonist Pseudozyma flocculosa and related fungi. Phytopathology 2001, 91, 249–254. [Google Scholar] [CrossRef]
- Li, C.H.; Cervantes, M.; Springer, D.J.; Boekhout, T.; Ruiz-Vazquez, R.M.; Torres-Martinez, S.R.; Heitman, J.; Lee, S.C. Sporangiospore size dimorphism is linked to virulence of Mucor circinelloides. PLoS Path. 2011, 7, e1002086. [Google Scholar] [CrossRef]
- Jankowiak, R.; Strzałka, B.; Bilański, P.; Kacprzyk, M.; Wieczorek, P.; Linnakoski, R. Ophiostomatoid fungi associated with hardwood-infesting bark and ambrosia beetles in Poland: Taxonomic diversity and vector specificity. Fungal Ecol. 2019, 39, 152–167. [Google Scholar] [CrossRef]
- Houbraken, J.; Visagie, C.M.; Meijer, M.; Frisvad, J.C.; Busby, P.E.; Pitt, J.I.; Seifert, K.A.; Louis-Seize, G.; Demirel, R.; Yilmaz, N.; et al. A taxonomic and phylogenetic revision of Penicillium section Aspergilloides. Stud. Mycol. 2014, 78, 373–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.-M.; Boekhout, T.; Bai, F.-Y. Cryptococcus foliicola sp. nov. and Cryptococcus taibaiensis sp. nov., novel basidiomycetous yeast species from plant leaves. J. Gen. Appl. Microbiol. 2011, 57, 285–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Overview of the 59 species obtained from all lettuce samples analyzed in this study, by number of isolates. Only species with 2 or more isolates are labeled.
Figure 1.
Overview of the 59 species obtained from all lettuce samples analyzed in this study, by number of isolates. Only species with 2 or more isolates are labeled.

Figure 2.
Number of isolates by treatment (conventional, organic, hydroponic) for “core” community species with 2 or more isolates.
Figure 2.
Number of isolates by treatment (conventional, organic, hydroponic) for “core” community species with 2 or more isolates.

Figure 3.
Number of species (bars) and isolates (circles) by treatment (conventional, organic, hydroponic) for “core” community genera.
Figure 3.
Number of species (bars) and isolates (circles) by treatment (conventional, organic, hydroponic) for “core” community genera.

Figure 4.
Phylogenetic reconstructions based on ITS sequences for the following genera: (A), Aureobasidium; (B), Filobasidium; (C), Cladosporium; and (D), Papiliotrema. Topologies are the result of ML inference performed with IQ-TREE. For each node, the ML bootstrap (if ≥60) is presented above/below the branch leading to that node. Gray shading added for strains obtained from romaine lettuce samples during this study.
Figure 4.
Phylogenetic reconstructions based on ITS sequences for the following genera: (A), Aureobasidium; (B), Filobasidium; (C), Cladosporium; and (D), Papiliotrema. Topologies are the result of ML inference performed with IQ-TREE. For each node, the ML bootstrap (if ≥60) is presented above/below the branch leading to that node. Gray shading added for strains obtained from romaine lettuce samples during this study.

Figure 5.
Phylogenetic reconstructions based on ITS sequences for the following genera: (A), Naganishia; (B), Symmetrospora; and (C), Vishniacozyma. Topologies are the result of ML inference performed with IQ-TREE. For each node, the ML bootstrap (if ≥60) is presented above/below the branch leading to that node. Gray shading added for strains obtained from romaine lettuce samples during this study.
Figure 5.
Phylogenetic reconstructions based on ITS sequences for the following genera: (A), Naganishia; (B), Symmetrospora; and (C), Vishniacozyma. Topologies are the result of ML inference performed with IQ-TREE. For each node, the ML bootstrap (if ≥60) is presented above/below the branch leading to that node. Gray shading added for strains obtained from romaine lettuce samples during this study.

Figure 6.
Phylogenetic reconstructions based on ITS sequences for the following genera: (A) Rhodotorula; (B) Sampaiozyma; and (C) Sporobolomyces. Topologies are the result of ML inference performed with IQ-TREE. For each node, the ML bootstrap (if ≥60) is presented above/below the branch leading to that node. Gray shading added for strains obtained from romaine lettuce samples during this study.
Figure 6.
Phylogenetic reconstructions based on ITS sequences for the following genera: (A) Rhodotorula; (B) Sampaiozyma; and (C) Sporobolomyces. Topologies are the result of ML inference performed with IQ-TREE. For each node, the ML bootstrap (if ≥60) is presented above/below the branch leading to that node. Gray shading added for strains obtained from romaine lettuce samples during this study.

Table 1.
Features of sequence datasets and results of maximum likelihood (ML) inference by genus. Presented are the number of included isolates, number of total characters, number and percentage of parsimony-informative characters, the model of nucleotide substitution according to the corrected Akaike Information Criterion and the -lnL score of the best ML tree.
Table 1.
Features of sequence datasets and results of maximum likelihood (ML) inference by genus. Presented are the number of included isolates, number of total characters, number and percentage of parsimony-informative characters, the model of nucleotide substitution according to the corrected Akaike Information Criterion and the -lnL score of the best ML tree.
| Genus | Isolates | Characters | Informative | % Informative | Model | -lnL |
|---|---|---|---|---|---|---|
| Aureobasidium | 14 | 517 | 44 | 8.51 | SYM+I | 1317.537 |
| Cladosporium | 36 | 482 | 45 | 9.34 | SYM+R3 | 1530.217 |
| Filobasidium | 21 | 619 | 170 | 27.46 | TPM2u+F+R3 | 3027.598 |
| Naganishia | 18 | 588 | 67 | 11.39 | TIM2+F+R3 | 1906.687 |
| Papliotrema | 29 | 507 | 143 | 28.21 | GTR+F+I+G4 | 2833.792 |
| Rhodotorula | 21 | 572 | 139 | 24.30 | GTR+F+R3 | 2888.392 |
| Sampaiozyma | 11 | 553 | 63 | 11.39 | TIM3+F+I+G4 | 1541.096 |
| Sporobolomyces | 34 | 560 | 151 | 26.96 | TIM2+F+I+G4 | 3160.923 |
| Symmetrospora | 17 | 582 | 131 | 22.51 | GTR+F+G4 | 2778.027 |
| Vishniacozyma | 25 | 493 | 157 | 31.85 | TIM2+F+G4 | 2545.886 |
Table 2.
All genera obtained from romaine lettuce samples in this study, with number of species, number of total isolates and number of isolates by treatment (conventional, organic, hydroponic).
Table 2.
All genera obtained from romaine lettuce samples in this study, with number of species, number of total isolates and number of isolates by treatment (conventional, organic, hydroponic).
| Genus | Species | Total Isolates | Conventional | Organic | Hydroponic |
|---|---|---|---|---|---|
| Acanthophysium | 1 | 1 | 1 | ||
| Alternaria | 6 | 6 | 5 | 1 | |
| Aureobasidium | 2 | 10 | 6 | 4 | |
| Beauveria | 1 | 1 | 1 | ||
| Botryotinia | 1 | 1 | 1 | ||
| Bullera | 2 | 2 | 2 | ||
| Bulleromyces | 1 | 1 | 1 | ||
| Cladosporium | 8 | 59 | 19 | 34 | 6 |
| Cystofilobasidium | 2 | 12 | 7 | 5 | |
| Epicoccum | 1 | 1 | 1 | ||
| Filobasidium | 6 | 33 | 16 | 17 | |
| Gibellulopsis | 1 | 3 | 2 | 1 | |
| Holtermanniella | 1 | 1 | 1 | ||
| Leucosporidium | 1 | 1 | 1 | ||
| Metschnikowia | 2 | 3 | 1 | 2 | |
| Moesziomyces | 1 | 1 | 1 | ||
| Mucor | 1 | 1 | 1 | ||
| Naganishia | 1 | 1 | 1 | ||
| Ophiostoma | 1 | 1 | 1 | ||
| Papiliotrema | 2 | 3 | 1 | 2 | |
| Penicillium | 2 | 3 | 1 | 2 | |
| Rhodotorula | 3 | 3 | 1 | 1 | 1 |
| Sampaiozyma | 2 | 2 | 1 | 1 | |
| Sporobolomyces | 2 | 67 | 36 | 31 | |
| Stemphylium | 1 | 2 | 2 | ||
| Symmetrospora | 1 | 2 | 2 | ||
| Tilletiopsis | 1 | 1 | 1 | ||
| Vishniacozyma | 5 | 27 | 9 | 18 | |
| Total | 59 | 249 | 110 | 128 | 11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Haelewaters, D.; Urbina, H.; Brown, S.; Newerth-Henson, S.; Aime, M.C. Isolation and Molecular Characterization of the Romaine Lettuce Phylloplane Mycobiome. J. Fungi 2021, 7, 277. https://doi.org/10.3390/jof7040277
AMA Style
Haelewaters D, Urbina H, Brown S, Newerth-Henson S, Aime MC. Isolation and Molecular Characterization of the Romaine Lettuce Phylloplane Mycobiome. Journal of Fungi. 2021; 7(4):277. https://doi.org/10.3390/jof7040277
Chicago/Turabian StyleHaelewaters, Danny, Hector Urbina, Samuel Brown, Shannon Newerth-Henson, and M. Catherine Aime. 2021. "Isolation and Molecular Characterization of the Romaine Lettuce Phylloplane Mycobiome" Journal of Fungi 7, no. 4: 277. https://doi.org/10.3390/jof7040277
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.