Proper Functions of Peroxisomes Are Vital for Pathogenesis of Citrus Brown Spot Disease Caused by Alternaria alternata

 ,
,  and
and 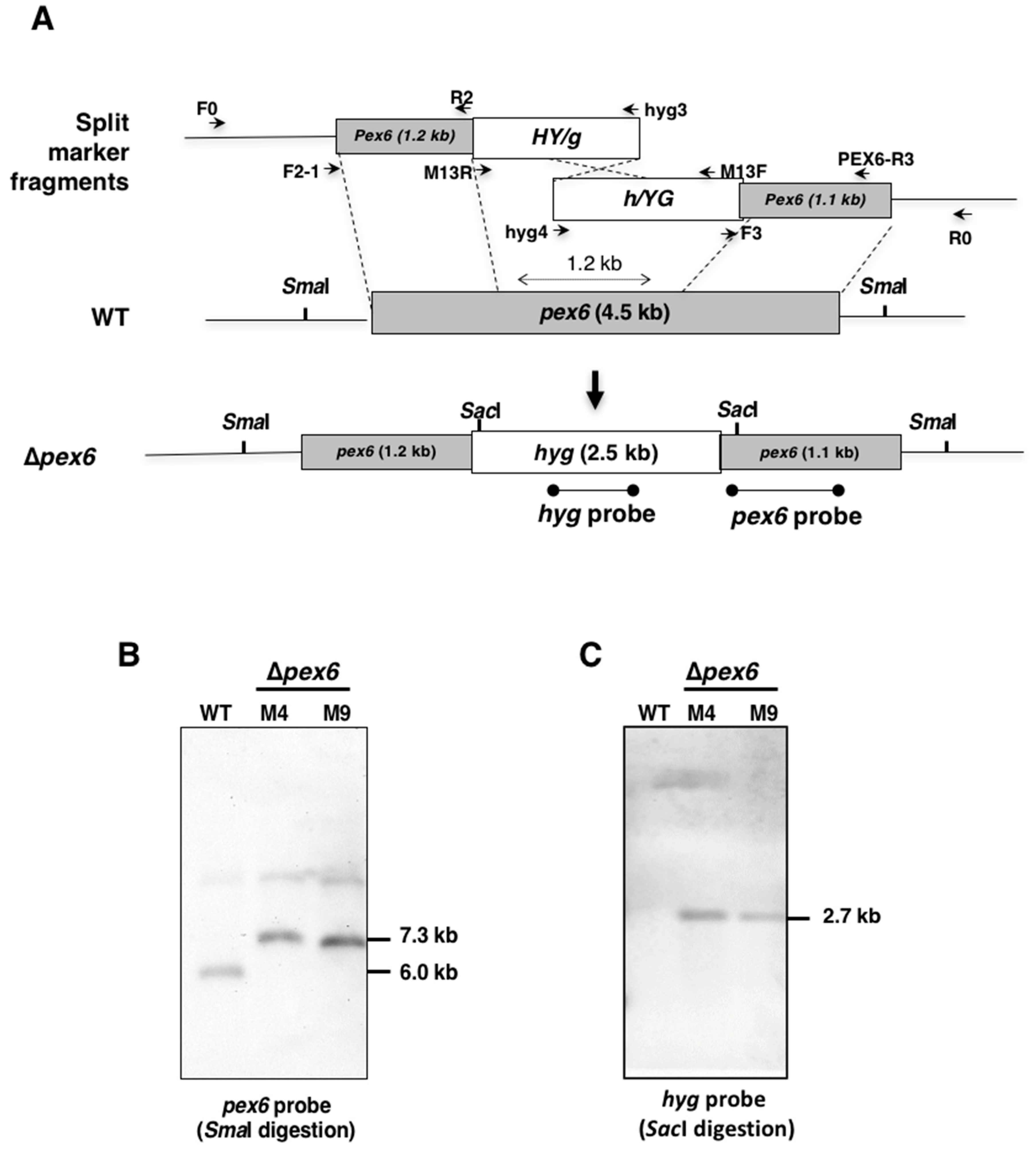{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture Conditions
2.2. Assays for Germination of Spores, Formation of Appressorium-Like Structures and Cell Viability
2.3. Sensitivity Tests
2.4. Molecular and Genetic Procedures
2.5. Virulence Assays
2.6. Purification and Analysis of Host-Selective Toxin
2.7. Microscopy
2.8. Statistical Analysis
3. Results
3.1. Identification and Mutation of Pex6 in the Tangerine Pathotype of A. alternata
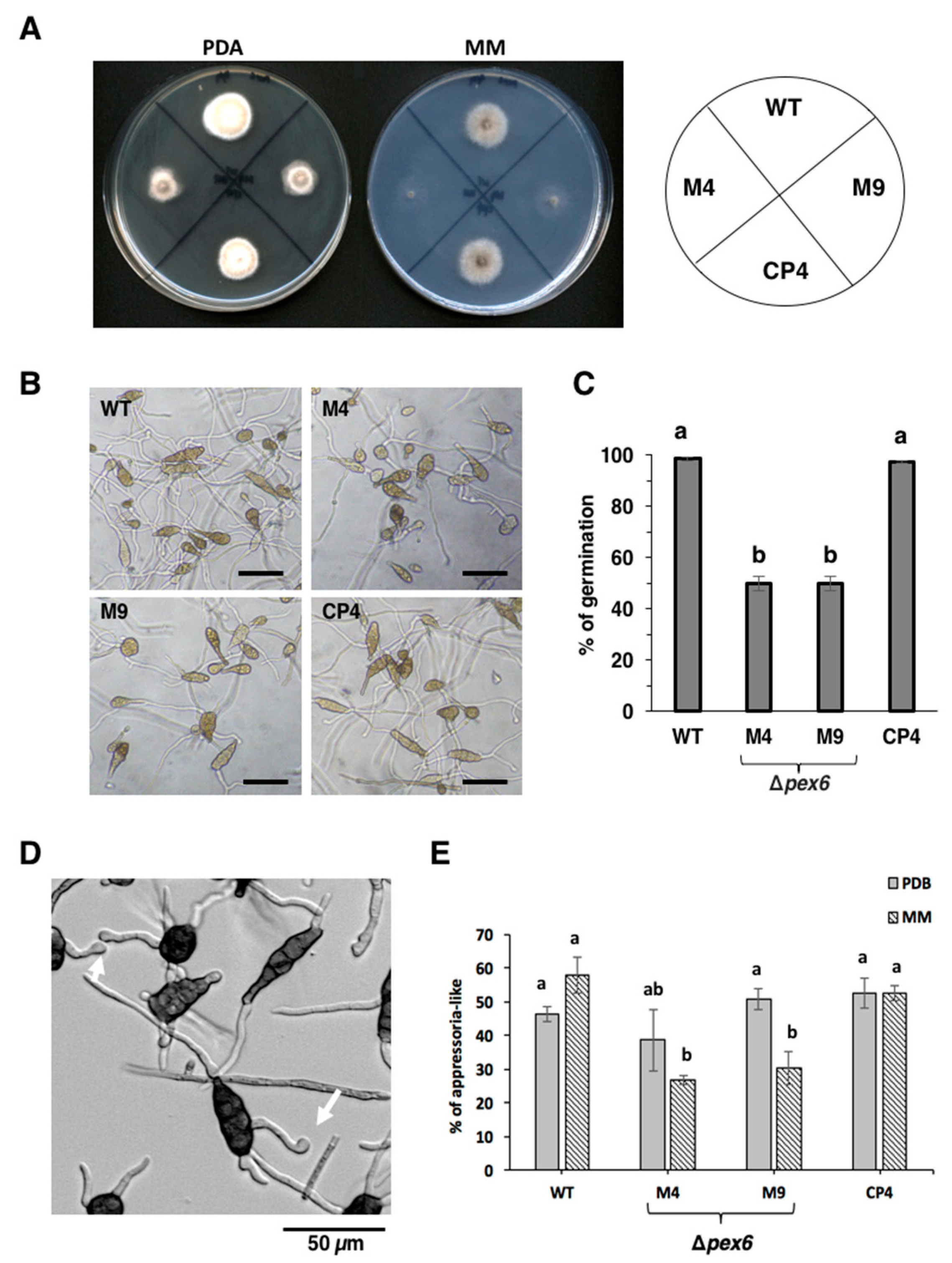
3.2. Pex6 is Required for Hyphal Growth and Conidia Germination
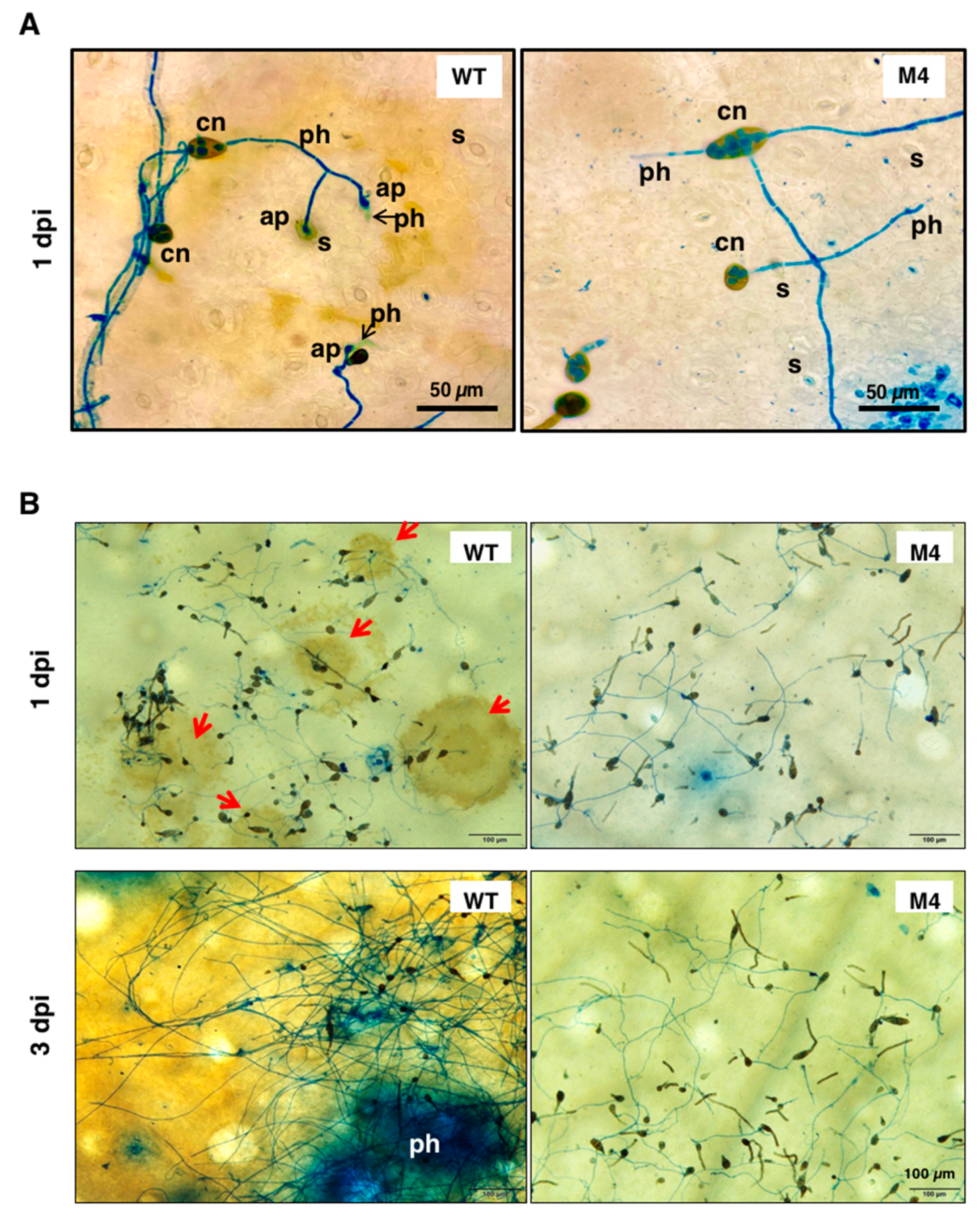
3.3. Pex6 Impacts the Formation of Appressorium-Like Structures
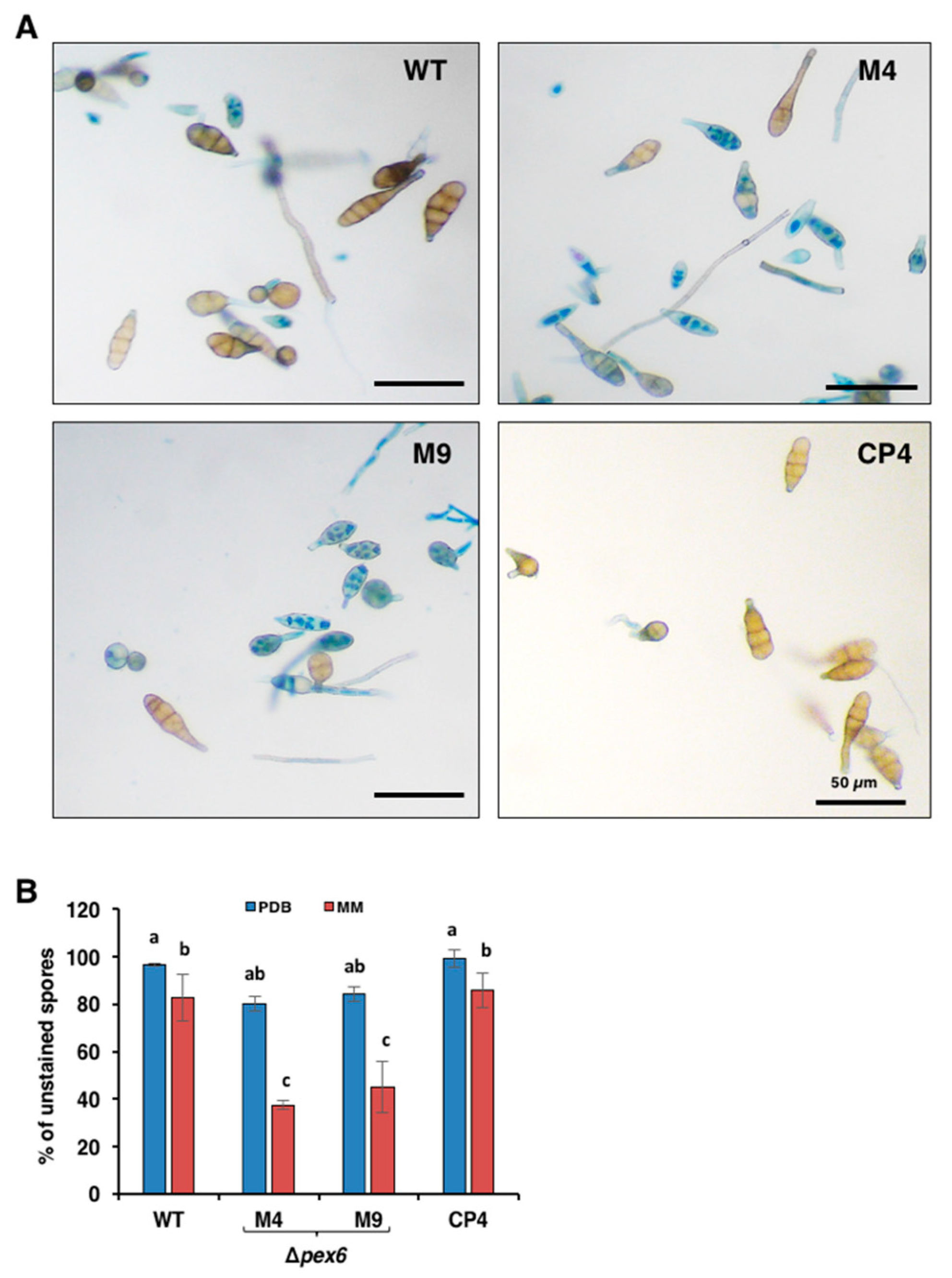
3.4. Deletion of Pex6 Leads to Low Conidia Viability
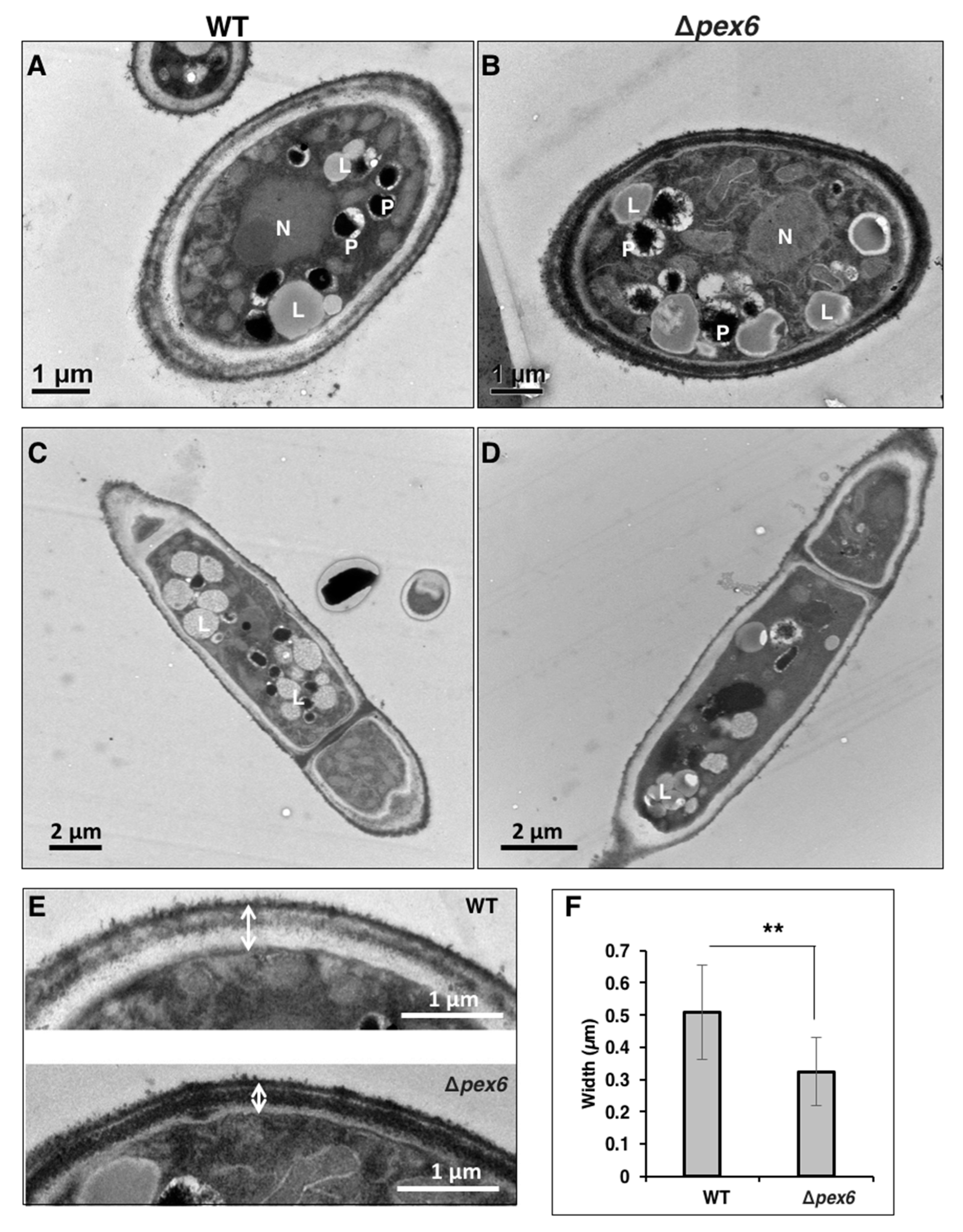
3.5. Pex6 is Involved in Lipid Metabolism and Maintenance of Cell-Wall Integrity
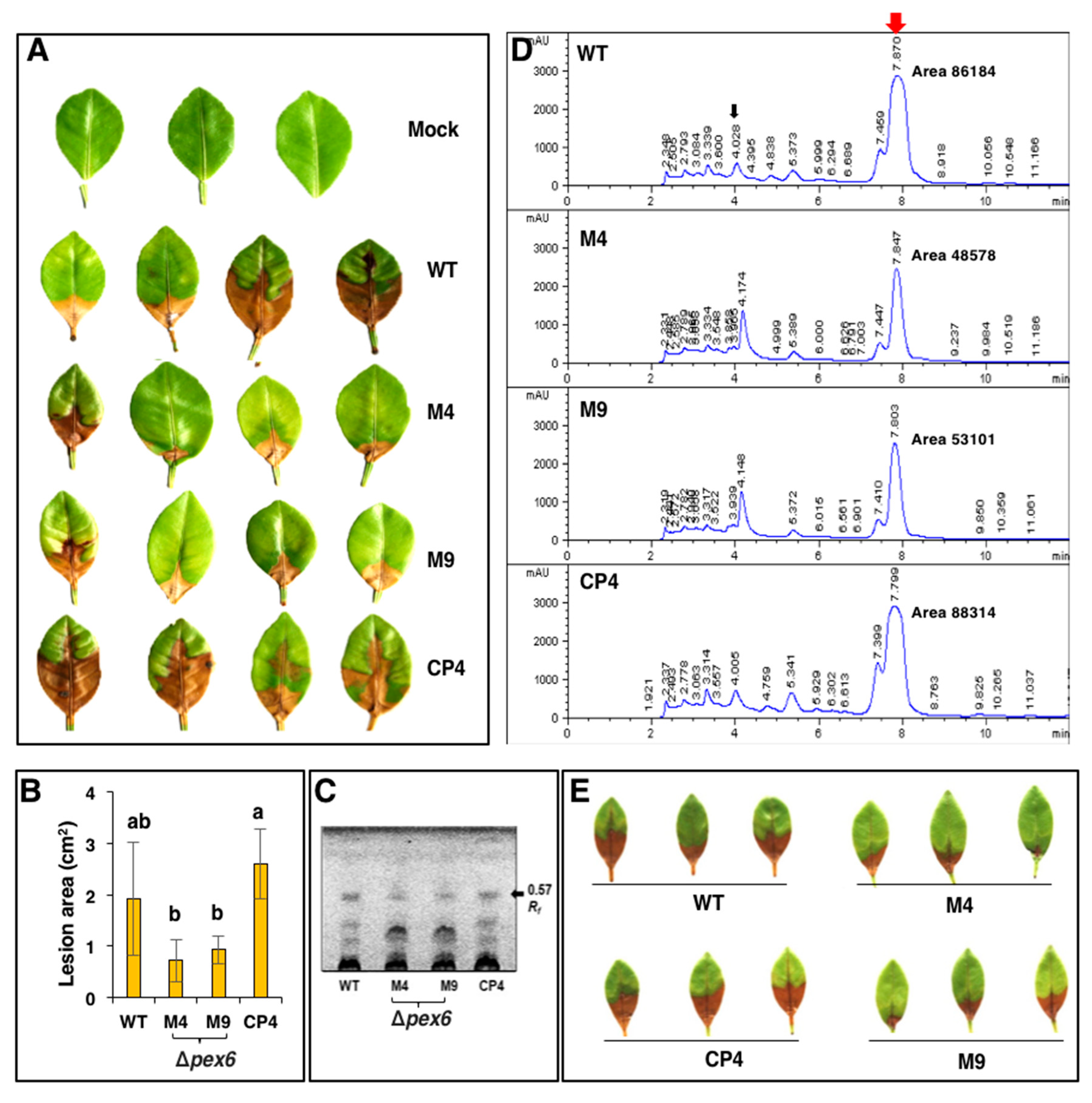
3.6. Pex6 is Required for Lesion Formation
3.7. Pex6 Deficiency Leads to Penetration Impairment
3.8. Pex6 Deficiency Impacts Toxin Production
3.9. Co-Inoculation of Δpex6 with Toxin Partially Restores Lesion Formation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- De Duve, C.; Baudhuin, P. Peroxisomes (microbodies and related particles). Physiol. Rev. 1966, 46, 323–357. [Google Scholar] [CrossRef]
- Titorenko, V.I.; Rachubinski, R.A. Dynamics of peroxisome assembly and function. Trend. Cell. Biol. 2001, 11, 22–29. [Google Scholar] [CrossRef]
- Wanders, R.J.; Ferdinandusse, S.; Brites, P.; Kemp, S. Peroxisomes, lipid metabolism and lipotoxicity. Biochim. Biophys. Acta 2010, 1801, 272–280. [Google Scholar] [CrossRef]
- Freitag, J.; Ast, J.; Linne, U.; Stehlik, T.; Martorana, D.; Bölker, M.; Sandrock, B. Peroxisomes contribute to biosynthesis of extracellular glycolipids in fungi. Mol. Microbiol. 2014, 93, 24–36. [Google Scholar] [CrossRef]
- Schrader, M.; Fahimi, H.D. Peroxisomes and oxidative stress. Biochim. Biophys. Acta 2006, 1763, 1755–1766. [Google Scholar] [CrossRef] [Green Version]
- Antonenkov, V.D.; Grunau, S.; Ohlmeier, S.; Hiltunen, J.K. Peroxisomes are oxidative organelles. Antioxid. Redox. Signal. 2010, 13, 525–537. [Google Scholar] [CrossRef]
- Nagan, N.; Zoeller, R.A. Plasmalogens: Biosynthesis and function. Prog. Lipid Res. 2001, 40, 199–229. [Google Scholar] [CrossRef]
- Wanders, R.J.; Waterham, H.R. Biochemistry of mammalian peroxisomes revisited. Annu. Rev. Biochem. 2006, 75, 295–332. [Google Scholar] [CrossRef]
- Boisson-Dernier, A.; Frietsch, S.; Kim, T.H.; Dizon, M.B.; Schroeder, J.I. The peroxin loss-of-function mutation abstinence by mutual consent disrupts male-female gametophyte recognition. Curr. Biol. 2008, 18, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Baker, A.; Bartel, B.; Linka, N.; Mullen, R.T.; Reumann, S.; Zolman, B.K. Plant Peroxisomes: Biogenesis and function. Plant Cell 2012, 24, 2279–2303. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, J.-I.; Yamaoka, S.; Matsuo, I.; Tsutsumi, N.; Kitamoto, K. A newly discovered function of peroxisomes: Involvement in biotin biosynthesis. Plant Signal. Behavior. 2012, 7, 1589–1593. [Google Scholar] [CrossRef] [Green Version]
- Titorenko, V.I.; Rachubinski, R.A. The peroxisome: Orchestrating important developmental decisions from inside the cell. J. Cell Biol. 2004, 164, 641–645. [Google Scholar] [CrossRef] [Green Version]
- Bhambra, G.K.; Wang, Z.-Y.; Soanes, D.M.; Wakley, G.E.; Talbot, N.J. Peroxisomal carnitine acetyl transferase is required for elaboration of penetration hyphae during plant infection by Magnaporthe grisea. Mol. Microbiol. 2006, 61, 46–60. [Google Scholar] [CrossRef]
- Weller, S.; Gould, S.J.; Valle, D. Peroxisome biogenesis disorders. Annu. Rev. Genomics Hum. Genet. 2003, 4, 165–211. [Google Scholar] [CrossRef]
- Peraza-Reyes, L.; Berteaux-Lecellier, V. Peroxisomes and sexual development in fungi. Front. Physiol. 2013, 4, 244. [Google Scholar] [CrossRef] [Green Version]
- Imazaki, A.; Tanaka, A.; Harimoto, Y.; Yamamoto, M.; Akimitsu, K.; Park, P.; Tsuge, T. Contribution of peroxisomes to secondary metabolism and pathogenicity in the fungal plant pathogen Alternaria alternata. Eukaryot. Cell 2010, 9, 682–694. [Google Scholar] [CrossRef] [Green Version]
- Meijer, W.H.; Gidijala, L.; Fekken, S.; Kiel, J.A.K.W.; van den Berg, M.A.; Lascaris, R.; Bovenberg, R.A.L.; van der Klei, I.J. Peroxisomes are required for efficient penicillin biosynthesis in Penicillium chrysogenum. Appl. Environ. Microbiol. 2010, 76, 5702–5709. [Google Scholar] [CrossRef] [Green Version]
- Bartoszewska, M.; Opaliński, Ł.; Veenhuis, M.; van der Klei, I.J. The significance of peroxisomes in secondary metabolite biosynthesis in filamentous fungi. Biotechnol. Lett. 2011, 33, 1921–1931. [Google Scholar] [CrossRef] [Green Version]
- Stehlik, T.; Sandrock, B.; Ast, J.; Freitag, J. Fungal peroxisomes as biosynthetic organelles. Curr. Opin. Cell Microbiol. 2014, 22, 8–14. [Google Scholar] [CrossRef] [PubMed]
- van der Klei, I.J.; Veenhuis, M. The versatility of peroxisome function in filamentous fungi. In: Peroxisomes and their key role in cellular signaling and metabolism. Subcell. Biochem. 2013, 69, 135–152. [Google Scholar]
- Gründlinger, M.; Yasmin, S.; Lechner, B.E.; Geley, S.; Schrettl, M.; Hynes, M.; Haas, H. Fungal siderophore biosynthesis is partially localized in peroxisomes. Mol. Microbiol. 2013, 88, 862–875. [Google Scholar] [CrossRef] [Green Version]
- Idnurm, A.; Giles, S.S.; Perfect, J.R.; Heitman, J. Peroxisome function regulates growth on glucose in the basidiomycete Cryptococcus neoformans. Eukaryot. Cell 2007, 6, 60–72. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, J.-I.; Kitamoto, K. Expanding functional repertoires of fungal peroxisomes: Contribution to growth and survival processes. Front. Physiol. 2013, 4, 177. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.-C.; Chen, C.-W.; Choo, C.Y.L.; Chen, Y.-K.; Yago, J.I.; Chung, K.-R. Biotin biosynthesis regulated by NADPH oxidases and lipid metabolism is required for infectivity in the citrus fungal pathogen Alternaria alternata. Microbiol. Res. 2020, in press. [Google Scholar] [CrossRef]
- Kimura, A.; Takano, Y.; Furusawa, I.; Okuno, T. Peroxisomal metabolic function is required for appressorium-mediated plant infection by Colletotrichum lagenarium. Plant Cell 2001, 13, 1945–1957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.-Y.; Soanes, D.M.; Kershaw, M.J.; Talbot, N.J. Functional analysis of lipid metabolism in Magnaporthe grisea reveals a requirement for peroxisomal fatty acid β-oxidation during appressorium-mediated plant infection. Mol. Plant–Microbe Interact. 2007, 20, 475–491. [Google Scholar] [CrossRef] [Green Version]
- Fujihara, N.; Sakaguchi, A.; Tanaka, S.; Fujii, S.; Tsuji, G.; Shiraishi, T.; O’Connell, R.; Kubo, Y. Peroxisome biogenesis factor PEX13 is required for appressorium-mediated plant infection by the anthracnose fungus Colletotrichum orbiculare. Mol. Plant–Microbe Interact. 2010, 23, 436–445. [Google Scholar] [CrossRef] [Green Version]
- Min, K.; Son, H.; Lee, J.; Choi, G.J.; Kim, J.-C.; Lee, Y.-W. Peroxisome function is required for virulence and survival of Fusarium graminearum. Mol. Plant–Microbe Interact. 2012, 25, 1617–1627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akimitsu, K.; Peever, T.L.; Timmer, L.W. Molecular, ecological and evolutionary approaches to understanding Alternaria diseases of citrus. Mol. Plant Pathol. 2003, 4, 435–446. [Google Scholar] [CrossRef]
- Kohmoto, K.; Itoh, Y.; Shimomura, N.; Kondoh, Y.; Otani, H.; Nishimura, S.; Nakatsuka, S. Isolation and biological activities of two host-specific toxins from tangerine pathotype of Alternaria alternata. Phytopathology 1993, 83, 495–502. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Masunaka, A.; Tsuge, T.; Yamamoto, M.; Ohtani, K.; Fukumoto, T.; Gomi, K.; Peever, T.L.; Akimitsu, K. Functional analysis of a multicopy host-selective ACT-toxin biosynthesis gene in the tangerine pathotype of Alternaria alternata using RNA silencing. Mol. Plant–Microbe Interact. 2008, 21, 1591–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, Y.; Masunaka, A.; Tsuge, T.; Yamamoto, M.; Ohtani, K.; Fukumoto, T.; Gomi, K.; Peever, T.L.; Tada, Y.; Ichimura, K.; et al. ACTTS3 encoding a polyketide synthase is essential for the biosynthesis of ACT-toxin and pathogenicity in the tangerine pathotype of Alternaria alternata. Mol. Plant–Microbe Interact. 2010, 23, 406–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Zhang, B.; Gai, Y.; Sun, X.; Chung, K.-R.; Li, H. Cell-wall-degrading enzymes required for virulence in the host selective toxin-producing necrotroph Alternaria alternata of citrus. Front. Microbiol. 2019, 10, 2514. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Chung, K.-R.; Li, H. The Alternaria alternata basal transcription factor II H subunit tfb5 required for DNA repair, toxin production, stress resistance and pathogenicity in citrus. Mol. Plant Pathol. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.-R. Stress response and pathogenicity of the necrotrophic fungal pathogen Alternaria alternata. Scientifica 2012. [Google Scholar] [CrossRef] [Green Version]
- Chung, K.-R. Reactive oxygen species in the citrus fungal pathogen Alternaria alternata: The roles of NADPH oxidase. Physiol. Mol. Plant Pathol. 2014, 88, 10–17. [Google Scholar] [CrossRef]
- Lin, C.-H.; Yang, S.L.; Chung, K.-R. The YAP1 homolog-mediated oxidative stress tolerance is crucial for pathogenicity of the necrotrophic fungus Alternaria alternata in citrus. Mol. Plant–Microbe Interact. 2009, 22, 942–952. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.L.; Lin, C.-H.; Chung, K.-R. Coordinate control of oxidative stress tolerance, vegetative growth, and fungal pathogenicity via the AP1-mediated pathway in the rough lemon pathotype of Alternaria alternata. Physiol. Mol. Plant Pathol. 2009, 74, 100–110. [Google Scholar] [CrossRef]
- Lin, C.-H.; Yang, S.L.; Chung, K.-R. Cellular responses required for oxidative stress tolerance, colonization and lesion formation by the necrotrophic fungus Alternaria alternata in citrus. Curr. Microbiol. 2011, 62, 807–815. [Google Scholar] [CrossRef]
- Chen, L.-H.; Lin, C.-H.; Chung, K.-R. Roles for SKN7 response regulator in stress resistance, conidiation and virulence in the citrus pathogen Alternaria alternata. Fungal Genet. Biol. 2012, 49, 802–813. [Google Scholar] [CrossRef]
- Yang, S.L.; Chung, K.-R. The NADPH oxidase-mediated production of H2O2 and resistance to oxidative stress in the necrotrophic pathogen Alternaria alternata of citrus. Mol. Plant Pathol. 2012, 13, 900–914. [Google Scholar] [CrossRef]
- Yang, S.L.; Chung, K.-R. Similar and distinct roles of NADPH oxidase components in the tangerine pathotype of Alternaria alternata. Mol. Plant Pathol. 2013, 14, 543–556. [Google Scholar] [CrossRef]
- Wang, P.-H.; Wu, P.-C.; Huang, R.; Chung, K.-R. The role of a nascent polypeptide-associated complex subunit alpha in siderophore biosynthesis, oxidative stress response and virulence in Alternaria alternata. Mol. Plant–Microbe Interact. 2020, 33, 668–679. [Google Scholar] [CrossRef]
- Chen, L.-H.; Lin, C.-H.; Chung, K.-R. A nonribosomal peptide synthetase mediates siderophore production and virulence in the citrus fungal pathogen Alternaria alternata. Mol. Plant Pathol. 2013, 14, 497–505. [Google Scholar] [CrossRef]
- Chen, L.-H.; Yang, S.L.; Chung, K.-R. Resistance to oxidative stress via regulating siderophore-mediated iron-acquisition by the citrus fungal pathogen Alternaria alternata. Microbiology 2014, 160, 970–979. [Google Scholar] [CrossRef]
- Chung, K.-R.; Wu, P.-C.; Chen, Y.-K.; Yago, J.I. The SreA repressor required for growth and suppression of siderophore biosynthesis, hydrogen peroxide resistance, cell wall integrity, and virulence in the phytopathogenic fungus Alternaria alternata. Fungal Genet. Biol. 2020, 139, 103384. [Google Scholar] [CrossRef]
- Lin, C.-H.; Chung, K.-R. Specialized and shared functions of the histidine kinase- and HOG1 MAP kinase-mediated signaling pathways in Alternaria alternata, a filamentous fungal pathogen of citrus. Fungal Genet. Biol. 2010, 47, 818–827. [Google Scholar] [CrossRef]
- Chung, K.-R. Mitogen-activated protein kinase signaling pathways of the tangerine pathotype of Alternaria alternata. MAP Kinase 2013, 2, e4. [Google Scholar] [CrossRef] [Green Version]
- Yu, P.-L.; Chen, L.-H.; Chung, K.-R. How the pathogenic fungus Alternaria alternata copes with stress via the response regulators SSK1 and SHO1. PLoS ONE 2016, 11, e0149153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.L.; Yu, P.-L.; Chung, K.-R. The glutathione peroxidase–mediated ROS resistance, fungicide sensitivity and cell wall construction in the citrus fungal pathogen Alternaria alternata. Environ. Microbiol. 2016, 18, 923–935. [Google Scholar] [CrossRef]
- Ma, H.; Wang, M.; Gai, Y.; Fu, H.; Ruan, R.; Chung, K.-R.; Li, H. Thioredoxin and glutaredoxin systems required for oxidative stress resistance, fungicide sensitivity and virulence of Alternaria alternata. Appl. Environ. Microbiol. 2018, 84, e00086-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.-H.; Tsai, H.-C.; Chung, K.-R. A major facilitator superfamily transporter-mediated resistance to oxidative stress and fungicides requires Yap1, Skn7, and MAP kinases in the citrus fungal pathogen Alternaria alternata. PLoS ONE 2017, 12, e0169103. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.-C.; Yu, P.-L.; Chen, L.-H.; Tsai, H.-C.; Chung, K.-R. A major facilitator superfamily transporter regulated by the stress-responsive transcription factor Yap1 is required for resistance to fungicides, xenobiotics, and oxidants and full virulence in Alternaria alternata. Front. Microbiol. 2018, 9, 2229. [Google Scholar] [CrossRef]
- Platta, H.W.; Erdmann, R. Peroxisomal dynamics. Trend. Cell Biol. 2007, 17, 474–484. [Google Scholar] [CrossRef]
- Smith, J.J.; Aitchison, J.D. Regulation of peroxisome dynamics. Curr. Opin. Cell Biol. 2009, 21, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Holroyd, C.; Erdmann, R. Protein translocation machineries of peroxisomes. FEBS Lett. 2001, 501, 6–10. [Google Scholar] [CrossRef]
- Platta, H.W.; Erdmann, R. The peroxisomal protein import machinery. FEBS Lett. 2007, 581, 2811–2819. [Google Scholar] [CrossRef]
- Chung, K.-R.; Lee, M.-H. Split marker-mediated transformation and targeted gene disruption in filamentous fungi. In Genetic Transformation System in Fungi; van den Berg, M., Maruthachalam, K., Eds.; Springer International Publishing: Cham, Switzerland, 2014; Volume 2, pp. 175–180. [Google Scholar]
- Wang, J.; Zhang, Z.; Wang, Y.; Li, L.; Chai, R.; Mao, X.; Jiang, H.; Qiu, H.; Du, X.; Lin, F.; et al. PTS1 peroxisomal import pathway plays shared and distinct roles to PTS2 pathway in development and pathogenicity of Magnaporthe oryzae. PLoS ONE 2013, 8, e5554. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.-Y.; Li, L.; Chai, R.-Y.; Qiu, H.-P.; Zhang, Z.; Wang, Y.-L.; Liu, X.-H.; Lin, F.-C.; Sun, G.-C. Pex13 and Pex14, the key components of the peroxisomal docking complex, are required for peroxisome formation, host infection and pathogenicity-related morphogenesis in Magnaporthe oryzae. Virulence 2019, 10, 292–314. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Pamplona, M.; Naqvi, N.I. Host invasion during rice-blast disease requires carnitine-dependent transport of peroxisomal acetyl-CoA. Mol. Microbiol. 2006, 61, 61–75. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, P.-C.; Chen, C.-W.; Choo, C.Y.L.; Chen, Y.-K.; Yago, J.I.; Chung, K.-R. Proper Functions of Peroxisomes Are Vital for Pathogenesis of Citrus Brown Spot Disease Caused by Alternaria alternata. J. Fungi 2020, 6, 248. https://doi.org/10.3390/jof6040248
Wu P-C, Chen C-W, Choo CYL, Chen Y-K, Yago JI, Chung K-R. Proper Functions of Peroxisomes Are Vital for Pathogenesis of Citrus Brown Spot Disease Caused by Alternaria alternata. Journal of Fungi. 2020; 6(4):248. https://doi.org/10.3390/jof6040248
Chicago/Turabian StyleWu, Pei-Ching, Chia-Wen Chen, Celine Yen Ling Choo, Yu-Kun Chen, Jonar I. Yago, and Kuang-Ren Chung. 2020. "Proper Functions of Peroxisomes Are Vital for Pathogenesis of Citrus Brown Spot Disease Caused by Alternaria alternata" Journal of Fungi 6, no. 4: 248. https://doi.org/10.3390/jof6040248