Effect of Tidal Cycles on Bacterial Biofilm Formation and Biocorrosion of Stainless Steel AISI 316L

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Material
2.2. Experimental Set-Up and Procedure
2.3. Corrosion Testing
2.4. Morphological Surface Characterization
2.5. DNA Extraction and Denaturing Gradient Gel Electrophoresis (DGGE)
2.6. Bioinformatics and Statistical Analyses
3. Results
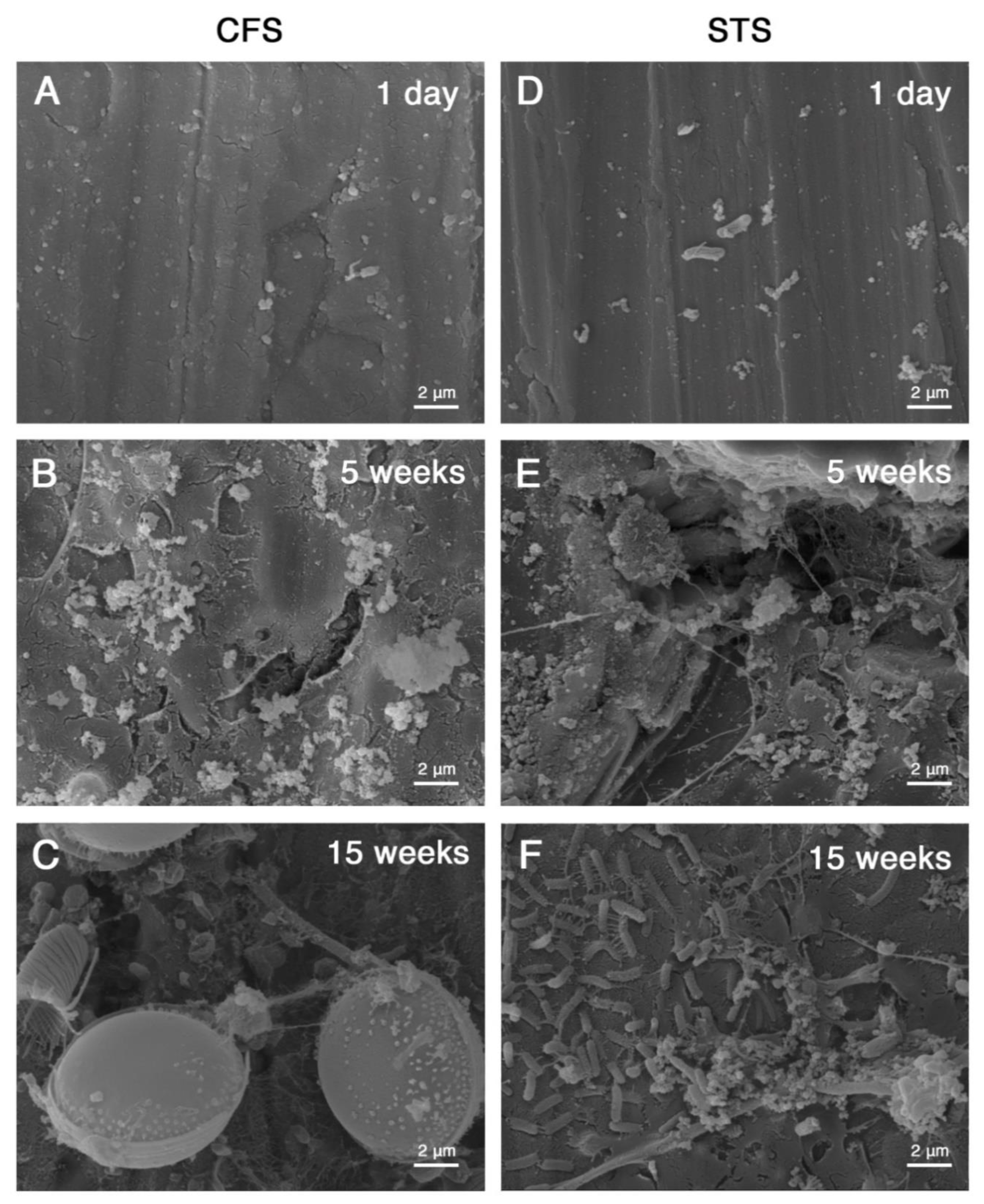
3.1. Microbial Cell Morphologies within the Biofilms
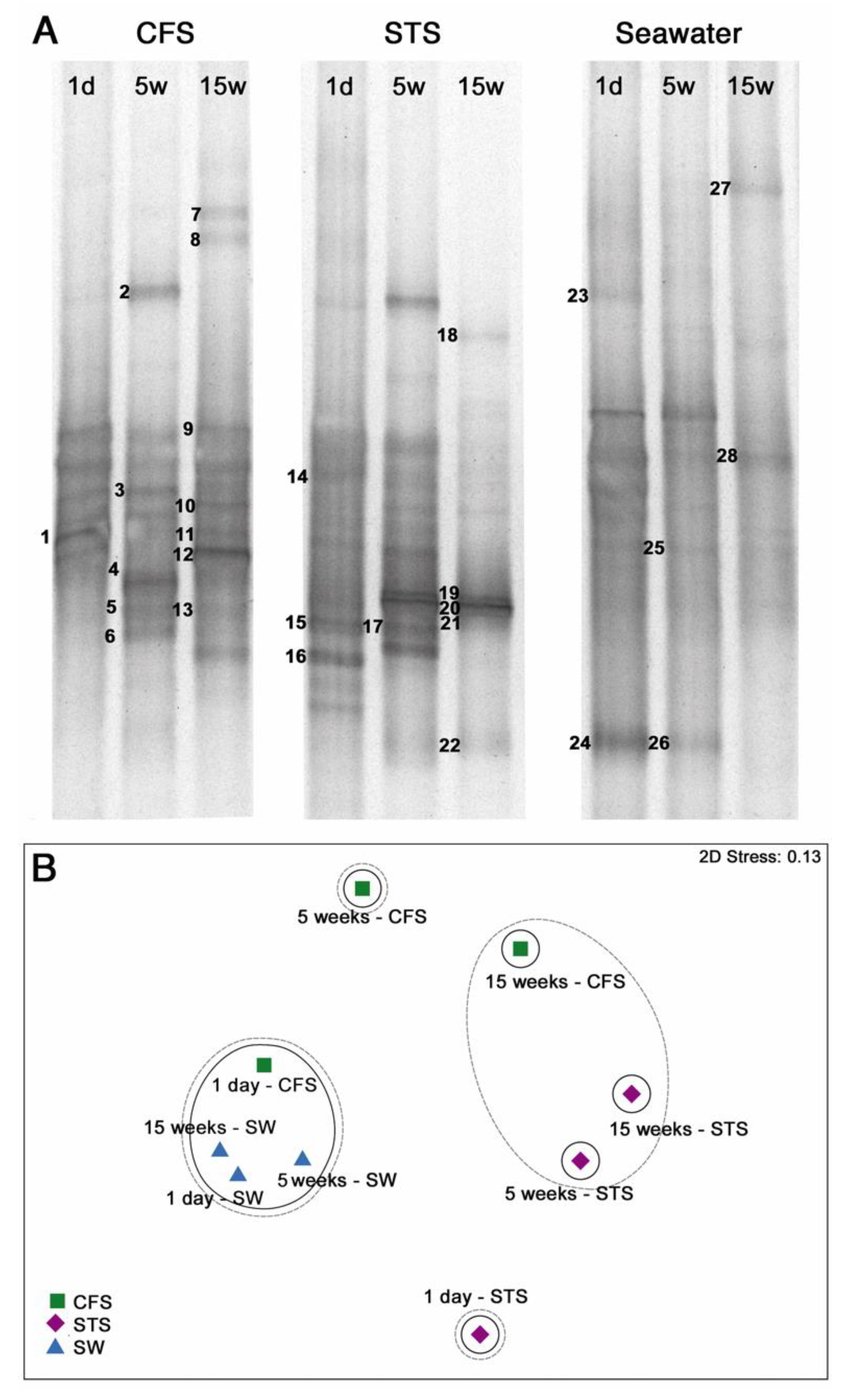
3.2. Bacterial Community Composition
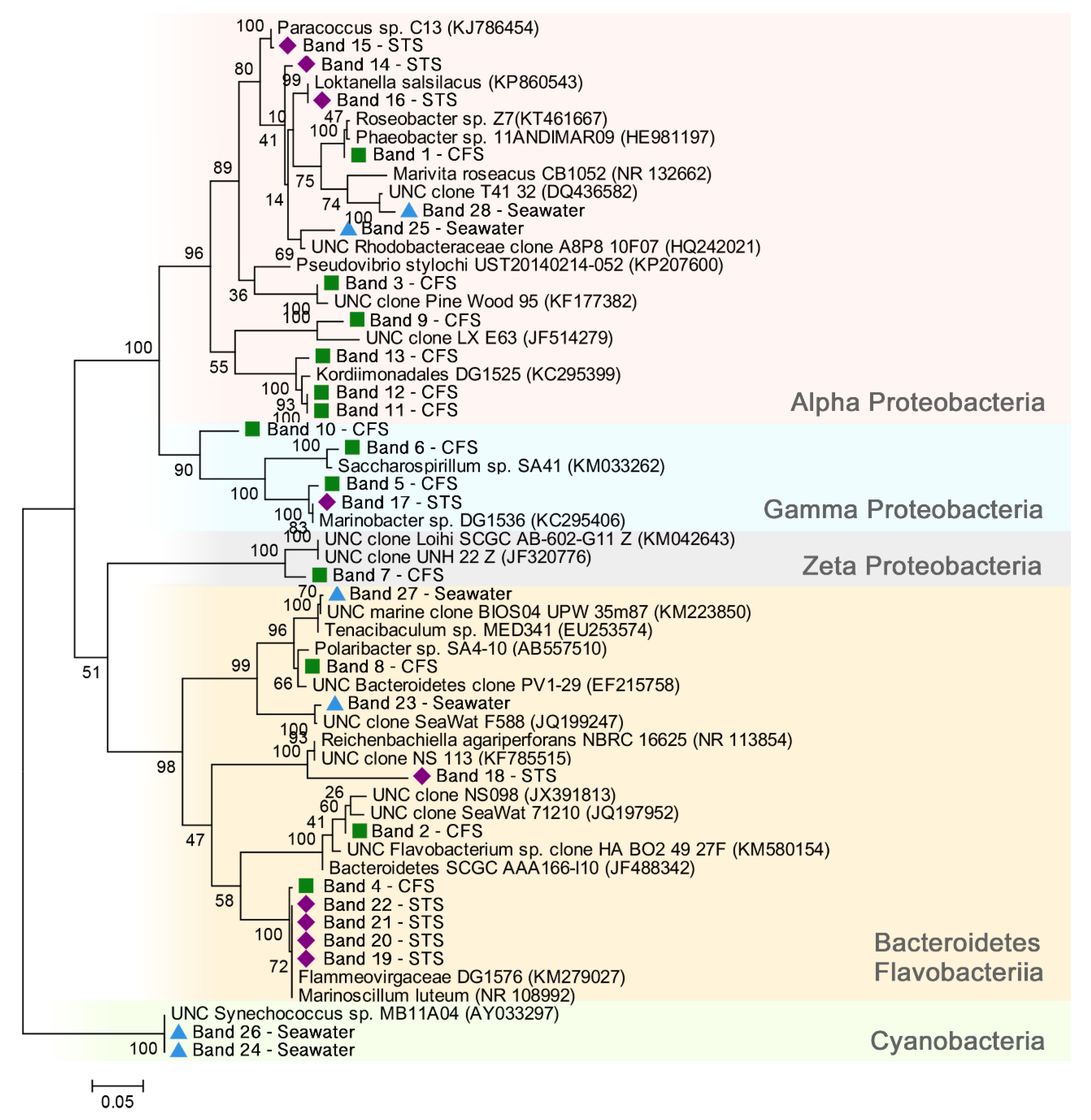
3.3. Bacterial Community—Phylogenetic Analysis of the Main Members
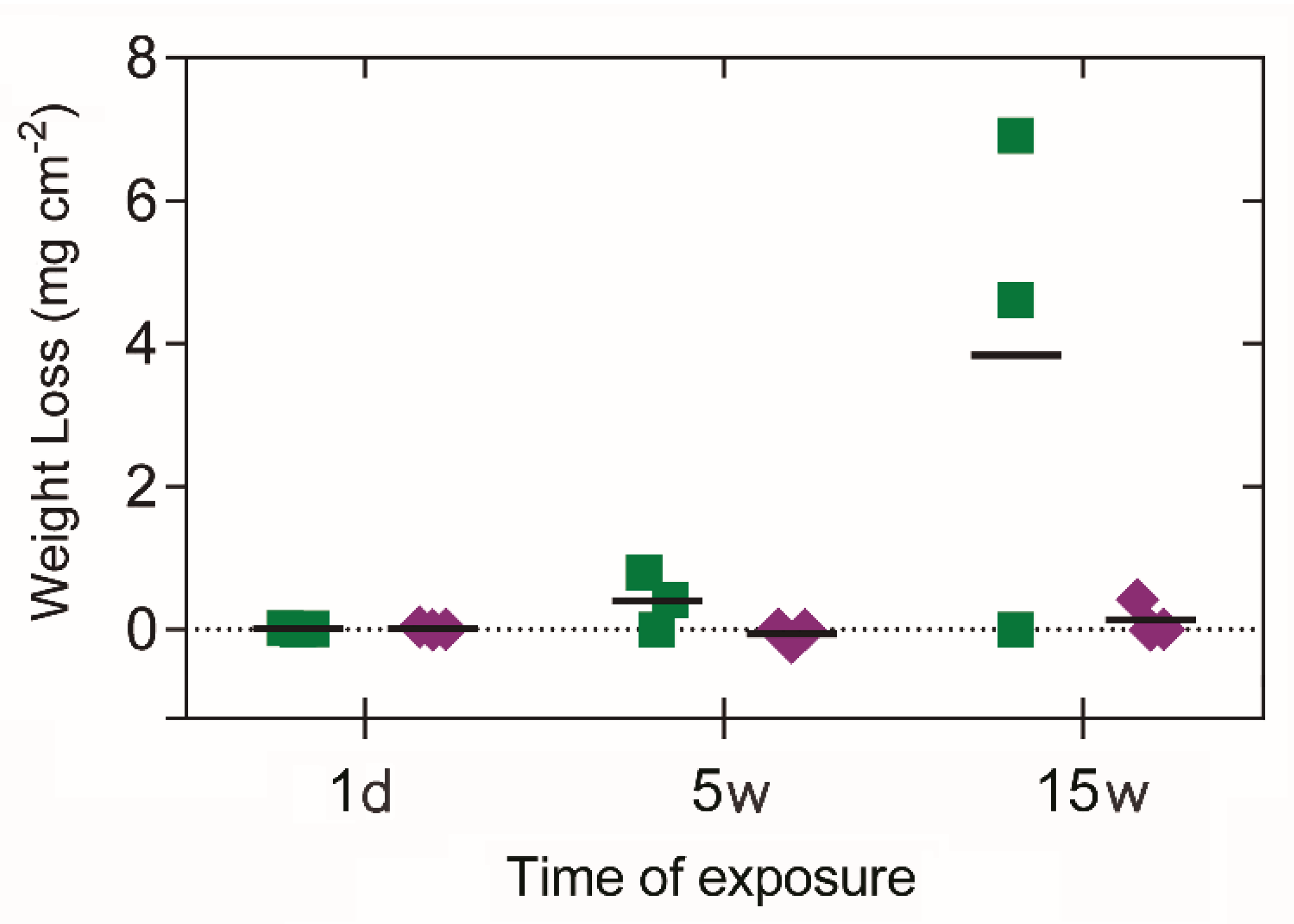
3.4. Corrosion Performance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pugh, D.T. Tides, Surges and Mean Sea-Level, 1st ed.; John Wiley & Sons: Hoboken, NJ, USA, 1987; ISBN 0-471-91505-X. [Google Scholar]
- European Federation of Corrosion. A Working Party Report on Marine Corrosion of Stainless Steels: Chlorination and Microbial Effects; Woodhead Pub Ltd.: Cambridge, UK, 1993; ISBN 0901716332. [Google Scholar]
- European Federation of Corrosion. Marine Corrosion of Stainless Steels; Feron, D., Ed.; IOM Communications, Maney Publishing: Leeds, UK, 1993; ISBN 1861251513. [Google Scholar]
- Melchers, R. Effect on marine immersion corrosion of carbon content of low alloy steels. Corros. Sci. 2003, 45, 2609–2625. [Google Scholar] [CrossRef]
- Melchers, R.E. Long-term immersion corrosion of steels in seawaters with elevated nutrient concentration. Corros. Sci. 2014, 81, 110–116. [Google Scholar] [CrossRef]
- Païssé, S.; Ghiglione, J.F.; Marty, F.; Abbas, B.; Gueuné, H.; Amaya, J.M.S.; Muyzer, G.; Quillet, L. Sulfate-reducing bacteria inhabiting natural corrosion deposits from marine steel structures. Appl. Microbiol. Biotechnol. 2013, 97, 7493–7504. [Google Scholar] [CrossRef]
- Koch, G.H.; Brongers, M.P.H.; Thompson, N.G.; Virmani, P.Y.; Payer, J.H. Corrosion Costs and Preventive Strategies in the United States; NACE International Report FHWA-RD-01; NACE International: Houston, TX, USA, 2002. [Google Scholar]
- Stipanicev, M.; Turcu, F.; Esnault, L.; Rosas, O.; Basseguy, R.; Sztyler, M.; Beech, I.B. Corrosion of carbon steel by bacteria from North Sea offshore seawater injection systems: Laboratory investigation. Bioelectrochemistry 2014, 97, 76–88. [Google Scholar] [CrossRef] [Green Version]
- Landoulsi, J.; El Kirat, K.; Richard, C.; Féron, D.; Pulvin, S. Enzymatic Approach of Microbial Influenced Corrosion, a Review Based on Stainless Steel in Natural Seawater. Environ. Sci. Technol. 2008, 42, 2233–2242. [Google Scholar] [CrossRef]
- Beech, I.B.; Sunner, J. Biocorrosion: Towards understanding interactions between biofilms and metals. Curr. Opin. Biotechnol. 2004, 15, 181–186. [Google Scholar] [CrossRef]
- Dexter, S.C.; Dexter, S.C. Corrosion in Seawater. Environments 2006, 13, 27–41. [Google Scholar]
- Lotze, H.K.; Worm, B.; Molis, M.; Wahl, M. Effects of UV radiation and consumers on recruitment and succession of a marine macrobenthic community. Mar. Ecol. Prog. Ser. 2002, 243, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Yeo, S.K.; Huggett, M.J.; Eiler, A.; Rappé, M.S. Coastal Bacterioplankton Community Dynamics in Response to a Natural Disturbance. PLoS ONE 2013, 8, e56207. [Google Scholar] [CrossRef]
- Eashwar, M.; Subramanian, G.; Palanichamy, S.; Rajagopal, G. The influence of sunlight on the localized corrosion of UNS S31600 in natural seawater. Biofouling 2011, 27, 837–849. [Google Scholar] [CrossRef]
- Eashwar, M.; Lakshman Kumar, A.; Sreedhar, G.; Kennedy, J.; Suresh Bapu, R.H. Stainless steel in coastal seawater: Sunlight counteracts biologically enhanced cathodic kinetics. Biofouling 2014, 30, 929–939. [Google Scholar] [CrossRef]
- Melchers, R.E.; Jeffrey, R. Corrosion of long vertical steel strips in the marine tidal zone and implications for ALWC. Corros. Sci. 2012, 65, 26–36. [Google Scholar] [CrossRef]
- Jeffrey, R.; Melchers, R.E. Effect of vertical length on corrosion of steel in the tidal zone. Corrosion 2009, 65, 695–702. [Google Scholar] [CrossRef]
- Hamilton, W.A. Sulphate-Reducing Bacteria and Anaerobic Corrosion. Annu. Rev. Microbiol. 1985, 39, 195–217. [Google Scholar] [CrossRef]
- Lichter, J.A.; Thompson, M.T.; Delgadillo, M.; Nishikawa, T.; Rubner, M.F.; Van Vliet, K.J. Substrata mechanical stiffness can regulate adhesion of viable bacteria. Biomacromolecules 2008, 9, 1571–1578. [Google Scholar] [CrossRef] [Green Version]
- King, R.A.; Miller, J.D.; Wakerleyt, D.S. Corrosion of Mild Steel in Cultures of Sulphate-Reducing Bacteria. Br. Corros. J. 1972, 8, 89–93. [Google Scholar] [CrossRef]
- Beech, I.B.; Sunner, J.A.; Hiraoka, K. Microbe-surface interactions in biofouling and biocorrosion processes. Int. Microbiol. 2005, 8, 157–168. [Google Scholar]
- Mansfeld, F. The interaction of bacteria and metal surfaces. Electrochim. Acta 2007, 52, 7670–7680. [Google Scholar] [CrossRef]
- Féron, D.; Dupont, I. Marine Biofilms on Stainless Steels: Effects on the Corrosion Behaviour. In Developments in Marine Corrosion; Woodhead Pub Ltd.: Cambridge, UK, 1998; pp. 89–102. [Google Scholar]
- Landolt, D. Corrosion and Surface Chemistry of Metals; Engineering Sciences: Materials; EPFL Press: Lausanne, Switzerland, 2007; Volume 10, ISBN 978-0-8493-8233-8. [Google Scholar]
- Videla, H. Biofilms and corrosion interactions on stainless steel in seawater. Int. Biodeterior. Biodegrad. 1994, 34, 245–257. [Google Scholar] [CrossRef]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as Complex Differentiated Communities. Annu. Rev. Microbiol. 2002, 56, 187–209. [Google Scholar] [CrossRef] [Green Version]
- Jia, R.; Unsal, T.; Xu, D.; Lekbach, Y.; Gu, T. Microbiologically influenced corrosion and current mitigation strategies: A state of the art review. Int. Biodeterior. Biodegrad. 2019, 137, 42–58. [Google Scholar] [CrossRef]
- Powell, C.; Francis, R. Corrosion Performance of Metals for the Marine Environment: A Basic Guide (EFC 63); CRC Press: Boca Raton, FL, USA, 2012; ISBN 978-1-907975-58-5. [Google Scholar]
- Aguirre, J.; Daille, L.; Fischer, D.A.; Galarce, C.; Pizarro, G.; Vargas, I.; Walczak, M.; de la Iglesia, R.; Armijo, F. Study of poly(3,4-ethylendioxythiphene) as a coating for mitigation of biocorrosion of AISI 304 stainless steel in natural seawater. Prog. Org. Coat. 2017, 113, 175–184. [Google Scholar] [CrossRef]
- Saha, J. Corrosion of Constructional Steels in Marine and Industrial Environment; Springer: Berlin/Heidelberg, Germany, 2013; ISBN 9783642076442. [Google Scholar]
- Kip, N.; van Veen, J.A. The dual role of microbes in corrosion. ISME J. 2015, 9, 542–551. [Google Scholar] [CrossRef]
- Videla, H.A. An overview of mechanisms by which sulphate-reducing bacteria influence corrosion of steel in marine environments. Biofouling 2000, 15, 37–47. [Google Scholar] [CrossRef]
- Sheng, X.; Ting, Y.-P.; Pehkonen, S.O. The influence of sulphate-reducing bacteria biofilm on the corrosion of stainless steel AISI 316. Corros. Sci. 2007, 49, 2159–2176. [Google Scholar] [CrossRef]
- Zhang, P.; Xu, D.; Li, Y.; Yang, K.; Gu, T. Electron mediators accelerate the microbiologically influenced corrosion of 304 stainless steel by the Desulfovibrio vulgaris biofilm. Bioelectrochemistry 2015, 101, 14–21. [Google Scholar] [CrossRef]
- Herrera, L.K.; Videla, H.A. Role of iron-reducing bacteria in corrosion and protection of carbon steel. Int. Biodeterior. Biodegrad. 2009, 63, 891–895. [Google Scholar] [CrossRef]
- Marty, F.; Gueuné, H.; Malard, E.; Sánchez-amaya, J.M.; Sjögren, L.; Abbas, B. Identification of key factors in Accelerated Low Water Corrosion through experimental simulation of tidal conditions: Influence of stimulated indigenous microbiota. Biofouling 2014, 30, 281–297. [Google Scholar] [CrossRef]
- Zhang, T.; Fang, H.H.P.; Ko, B.C.B. Methanogen population in a marine biofilm corrosive to mild steel. Appl. Microbiol. Biotechnol. 2003, 63, 101–106. [Google Scholar] [CrossRef]
- Rajasekar, A.; Anandkumar, B. Characterization of corrosive bacterial consortia isolated from petroleum-product-transporting pipelines. Appl. Microbiol. Biotechnol. 2010, 85, 1175–1188. [Google Scholar] [CrossRef]
- Lee, Y.K.; Kwon, K.; Cho, K.H.; Kim, H.W.; Park, J.H.; Lee, H.K. Culture and Identification of Bacteria from Marine Biofilms. J. Microbiol. 2003, 41, 183–188. [Google Scholar]
- Celikkol-Aydin, S.; Gaylarde, C.C.; Lee, T.; Melchers, R.E.; Witt, D.L.; Beech, I.B. 16S rRNA gene profiling of planktonic and biofilm microbial populations in the Gulf of Guinea using Illumina NGS. Mar. Environ. Res. 2016, 122, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Eashwar, M.; Subramanian, G.; Palanichamy, S.; Rajagopal, G.; Madhu, S.; Kamaraj, P. Cathodic behaviour of stainless steel in coastal Indian seawater: Calcareous deposits overwhelm biofilms. Biofouling 2009, 25, 191–201. [Google Scholar] [CrossRef] [PubMed]
- ASTM. ASTM International ASTM G1-03 (Reapproved 2017) Standard Practice for Preparing, Cleaning, and Evaluating Corrosion Test 1–9; ASTM: West Conshohocken, PA, USA, 2017. [Google Scholar]
- Fischer, D.A.; Daille, L.; Aguirre, J.; Galarce, C.; Armijo, F. Corrosion of Stainless Steel in Simulated Tide of Fresh Natural Seawater of South East Pacific. Int. J. Electrochem. Sci. 2016, 11, 6873–6885. [Google Scholar] [CrossRef]
- Schädler, S.; Burkhardt, C.; Kappler, A. Evaluation of Electron Microscopic Sample Preparation Methods and Imaging Techniques for Characterization of Cell-Mineral Aggregates. Geomicrobiol. J. 2008, 25, 228–239. [Google Scholar] [CrossRef]
- Fuhrman, J.A.; Comeau, D.E.; Hagström, A.; Chan, A.M. Extraction from natural planktonic microorganisms of DNA suitable for molecular biological studies. Appl. Environ. Microbiol. 1988, 54, 1426–1429. [Google Scholar] [CrossRef] [Green Version]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Huber, T.; Faulkner, G.; Hugenholtz, P. Bellerophon: A program to detect chimeric sequences in multiple sequence alignments. Bioinformatics 2004, 20, 2317–2319. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [PubMed]
- Li, S.; Li, L.; Qu, Q.; Kang, Y.; Zhu, B.; Yu, D.; Huang, R. Extracellular electron transfer of Bacillus cereus biofilm and its effect on the corrosion behaviour of 316L stainless steel. Colloids Surf. B Biointerfaces 2019, 173, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Nam, J.H.; Kim, Y.H.; Lee, K.H.; Lee, D.H. Bacterial communities in the initial stage of marine biofilm formation on artificial surfaces. J. Microbiol. 2008, 46, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Rampadarath, S.; Bandhoa, K.; Puchooa, D.; Jeewon, R.; Bal, S. Early bacterial biofilm colonizers in the coastal waters of Mauritius. Electron. J. Biotechnol. 2017, 29, 13–21. [Google Scholar] [CrossRef]
- Pollet, T.; Berdjeb, L.; Garnier, C.; Durrieu, G.; Le Poupon, C.; Misson, B.; Briand, J.-F. Prokaryotic community successions and interactions in marine biofilms: The key role of Flavobacteriia. FEMS Microbiol. Ecol. 2018, 94, fiy083. [Google Scholar] [CrossRef] [Green Version]
- Phan, H.C.; Wade, S.A.; Blackall, L.L. Is marine sediment the source of microbes associated with accelerated low water corrosion? Appl. Microbiol. Biotechnol. 2019, 103, 449–459. [Google Scholar] [CrossRef]
- Makita, H. Iron-oxidizing bacteria in marine environments: Recent progresses and future directions. World J. Microbiol. Biotechnol. 2018, 34, 1–13. [Google Scholar] [CrossRef]
- McBeth, J.M.; Emerson, D. In Situ Microbial Community Succession on Mild Steel in Estuarine and Marine Environments: Exploring the Role of Iron-Oxidizing Bacteria. Front. Microbiol. 2016, 7, 767. [Google Scholar] [CrossRef] [Green Version]
- McBeth, J.M.; Little, B.J.; Ray, R.I.; Farrar, K.M.; Emerson, D. Neutrophilic iron-oxidizing “Zetaproteobacteria” and mild steel corrosion in nearshore marine environments. Appl. Environ. Microbiol. 2011, 77, 1405–1412. [Google Scholar] [CrossRef] [Green Version]
- Dang, H.; Chen, R.; Wang, L.; Shao, S.; Dai, L.; Ye, Y.; Guo, L.; Huang, G.; Klotz, M.G. Molecular characterization of putative biocorroding microbiota with a novel niche detection of Epsilon- and Zetaproteobacteria in Pacific Ocean coastal seawaters. Environ. Microbiol. 2011, 13, 3059–3074. [Google Scholar] [CrossRef]
- Rosenberg, E.; DeLong, E.F.; Lory, S.; Stackebrandt, E.; Thompson, F. (Eds.) The Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2014; ISBN 978-3-642-38953-5. [Google Scholar]
- Dang, H.; Klotz, M.G.; Lovell, C.R.; Sievert, S.M. Editorial: The Responses of Marine Microorganisms, Communities and Ecofunctions to Environmental Gradients. Front. Microbiol. 2019, 10, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Moradi, M.; Song, Z.; Xiao, T. Exopolysaccharide produced by Vibrio neocaledonicus sp. as a green corrosion inhibitor: Production and structural characterization. J. Mater. Sci. Technol. 2018, 34, 2447–2457. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Time | OCP (V vs. Ag/AgCl) | Corrosion Rate a ± SD (mm·yr−1) |
|---|---|---|---|
| Reference | – | −0.02 | – |
| CFS | 1d | ND | 0.0046 ± 0.0045 |
| 5w | +0.25 | 0.0047 ± 0.0046 | |
| 15w | +0.28 | 0.0148 ± 0.0135 | |
| STS | 1d | ND | 0.0063 ± 0.0066 |
| 5w | +0.01 | 0.0007 ± 0.0012 | |
| 15w | +0.17 | 0.0006 ± 0.0009 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daille, L.K.; Aguirre, J.; Fischer, D.; Galarce, C.; Armijo, F.; Pizarro, G.E.; Walczak, M.; De la Iglesia, R.; Vargas, I.T. Effect of Tidal Cycles on Bacterial Biofilm Formation and Biocorrosion of Stainless Steel AISI 316L. J. Mar. Sci. Eng. 2020, 8, 124. https://doi.org/10.3390/jmse8020124
Daille LK, Aguirre J, Fischer D, Galarce C, Armijo F, Pizarro GE, Walczak M, De la Iglesia R, Vargas IT. Effect of Tidal Cycles on Bacterial Biofilm Formation and Biocorrosion of Stainless Steel AISI 316L. Journal of Marine Science and Engineering. 2020; 8(2):124. https://doi.org/10.3390/jmse8020124
Chicago/Turabian StyleDaille, Leslie K., Javiera Aguirre, Diego Fischer, Carlos Galarce, Francisco Armijo, Gonzalo E. Pizarro, Magdalena Walczak, Rodrigo De la Iglesia, and Ignacio T. Vargas. 2020. "Effect of Tidal Cycles on Bacterial Biofilm Formation and Biocorrosion of Stainless Steel AISI 316L" Journal of Marine Science and Engineering 8, no. 2: 124. https://doi.org/10.3390/jmse8020124