
Recent Advances in the Study of Marine Microbial Biofilm: From the Involvement of Quorum Sensing in Its Production up to Biotechnological Application of the Polysaccharide Fractions

 ,
, 
 ,
, 
Abstract
:1. Introduction
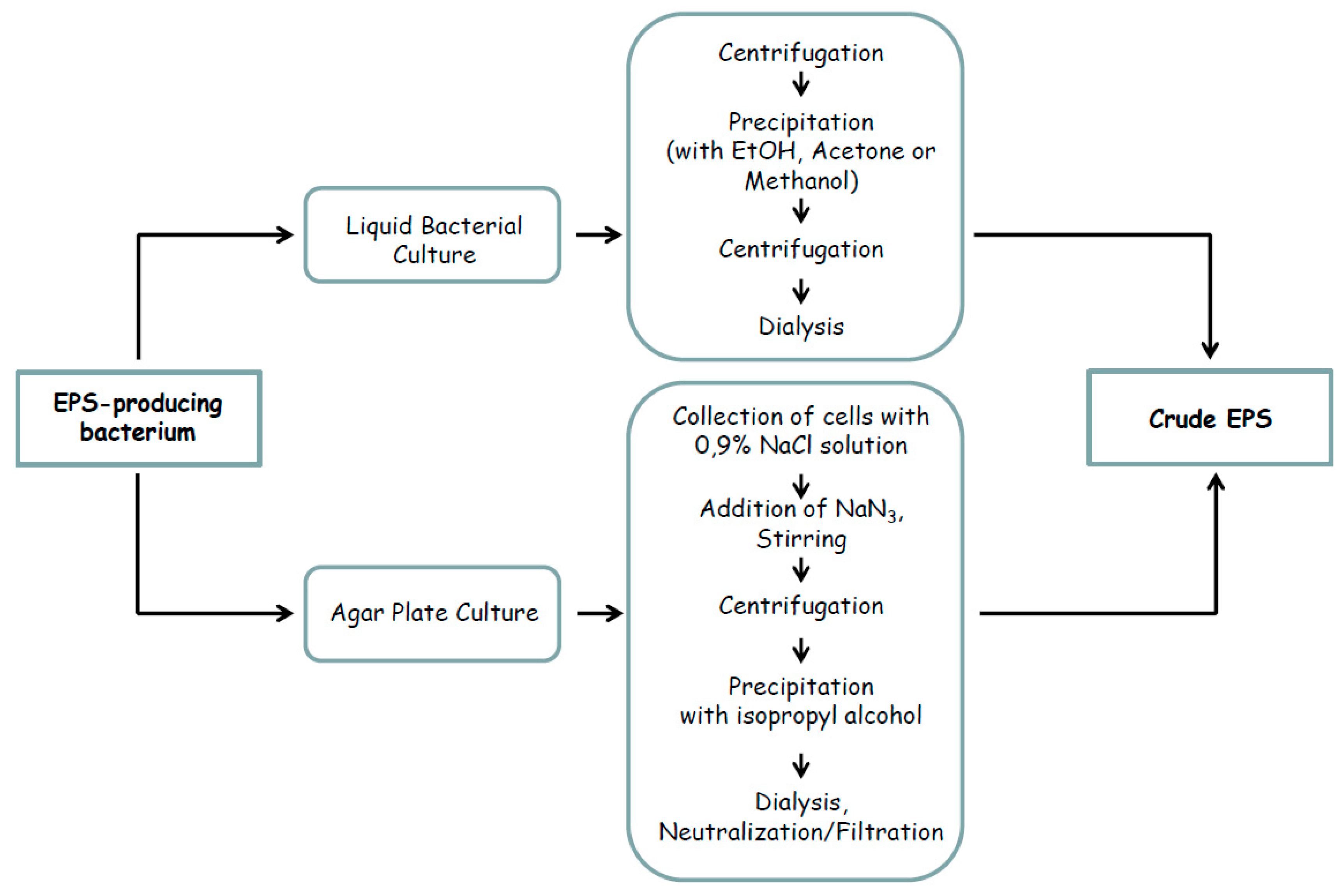
2. Marine Microbial Exopolysaccharides: Isolation and Purification Strategies
3. Marine Microbial Exopolysaccharides: Biotechnological Applications
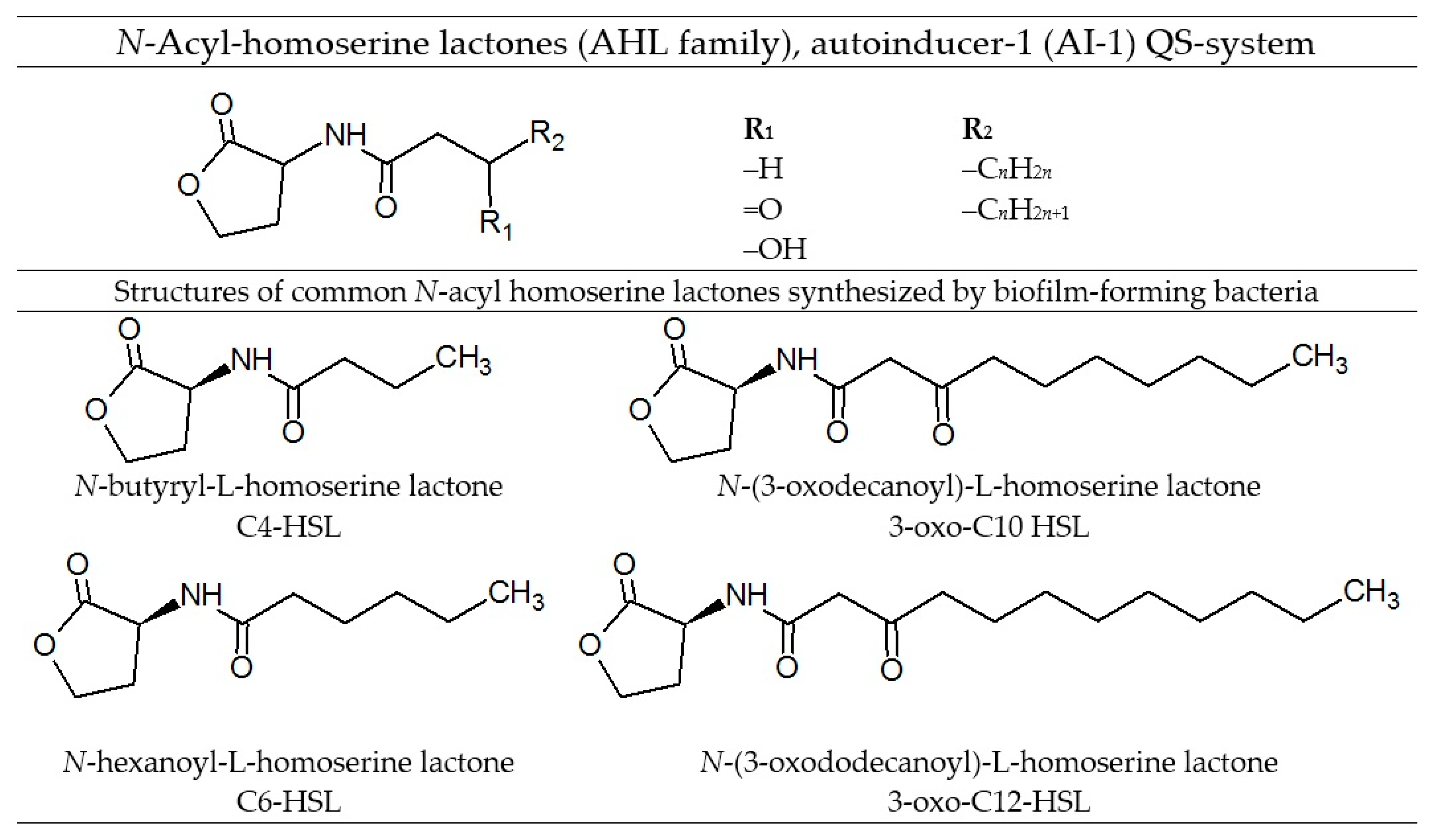
4. Quorum Sensing and Biofilm Development
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Albericio, F.; Álvarez, M.; Cuevas, C.; Francesch, A.; Pla, D.; Tulla-Puche, J. The Sea as a Source of New Drugs. In Molecular Imaging for Integrated Medical Therapy and Drug Development, 1st ed.; Tamaki, N.M.D., Yuji, K., Eds.; Springer Japan, Springer-Verlag: Tokyo, Japan, 2010; pp. 237–249. [Google Scholar]
- Jha, R.K.; Xu, Z. Biomedical Compounds from Marine organisms. Mar. Drugs 2004, 2, 123–146. [Google Scholar] [CrossRef]
- Manivasagana, P.; Venkatesana, J.; Sivakumarc, K.; Kima, S.K. Pharmaceutically active secondary metabolites of marine actinobacteria Panchanathan. Microbiol. Res. 2014, 169, 262–278. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, I.; Kim, S.K. Immense Essence of Excellence: Marine Microbial Bioactive Compounds. Mar. Drugs 2010, 8, 2673–2701. [Google Scholar] [CrossRef] [PubMed]
- Carte, B.K. Biomedical potential of marine natural products. Bioscience 1996, 46, 271–286. [Google Scholar]
- Erba, E.; Bergamaschi, D.; Ronzoni, S.; Faretta, M.; Taverna, S.; Bonfanti, M.; Catapano, C.V.; Faircloth, G.; Jimeno, J.; D’Incalci, M. Mode of action of thiocoraline, a natural marine compound with anti-tumour activity. Br. J. Canc. 1999, 80, 971–980. [Google Scholar] [CrossRef] [PubMed]
- Riedlinger, J.; Reicke, A.; Zähner, H.; Krismer, B.; Bull, A.T.; Maldonado, L.A. Abyssomicins, inhibitors of the para-aminobenzoic acid pathway produced by the marine Verrucosispora strain AB-18-032. J. Antibiot. 2004, 57, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.J.; Kim, T.S.; Lee, H.S.; Park, J.Y.; Choi, I.K.; Ho, Y.; Kwon, H.J. Streptopyrrolidine, an angiogenesis inhibitor from a marine-derived Streptomyces sp. KORDI-3973. Phytochemistry 2008, 69, 2363–2366. [Google Scholar] [CrossRef] [PubMed]
- Berenbaum, M.R. The chemistry of defense: Theory and practice. P.N.A.S. USA 1995, 92, 2–8. [Google Scholar] [CrossRef]
- Poli, A.; Anzelmo, G.; Nicolaus, B. Bacterial Exopolysaccharides from Extreme Marine Habitats: Production, Characterization and Biological Activities. Mar. Drugs 2010, 8, 1779–1802. [Google Scholar] [CrossRef] [PubMed]
- Decho, A.W. Microbial exopolymer secretions in ocean environments: Their role(s) in food webs and marine processes. Oceanogr. Mar. Biol. 1990, 28, 73–153. [Google Scholar]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial biofilms. Annu. Rev. Microbiol. 1995, 49, 711–745. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley, L.; Costerton, J.W.; Stoddley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microb. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Branda, S.; Vik, A.; Friedman, L.; Kolter, R. Biofilms: The matrix revisited. Trends Microbiol. 2005, 13, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Burmølle, M.; Ren, D.; Bjarnsholt, T.; Sørensen, S.J. Interactions in multispecies biofilms: Do they actually matter? Trends Microbiol. 2014, 22, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Matsumoto, S. Bacterial adhesion: From mechanism to control. Biochem. Eng. J. 2010, 48, 424–434. [Google Scholar] [CrossRef]
- De Rossi, B.P.; Calenda, M.; Vay, C.; Franco, M. Biofilm formation by Stenotrophomonasmaltophilia isolates from device-associated nosocomial infections. Rev. Argent. Microbiol. 2007, 39, 204–212. [Google Scholar]
- Palmer, J.; Flint, S.; Brooks, J. Bacterial cell attachment, the beginning of a biofilm. J. Ind. Microbiol. Biotechnol. 2007, 34, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J. Relevance of microbial extracellular polymeric substances (EPSs)—Part II: Technical aspects. Water Sci. Technol. 2001, 43, 9–16. [Google Scholar] [PubMed]
- Kenne, L.; Lindberg, B. Bacterial polysaccharides. In The Polysaccharides; Aspinall, G.O., Ed.; Academic Press: New York, NY, USA, 1983; Volume 2, pp. 287–363. [Google Scholar]
- Sutherland, I.W. The best and most comprehensive overview of the polysaccharide moiety of EPS. In Comprehensive Glycoscience; Kamerling, J.P., Ed.; Elsevier: Doordrecht, The Netherland, 2007; Volume 2, pp. 521–558. [Google Scholar]
- Nichols, C.A.; Guezennec, J.; Bowman, J.P. Bacterial Exopolysaccharides from Extreme Marine Environments with Special Consideration of the Southern Ocean, Sea Ice, and Deep-Sea Hydrothermal Vents: A Review. Mar. Biotechnol. 2005, 7, 253–271. [Google Scholar] [CrossRef] [PubMed]
- McDougald, D.; Rice, S.A.; Barraud, N.; Steinberg, P.D.; Kjelleberg, S. Should we stay or should we go: Mechanisms and ecological consequences for biofilm dispersal. Nat. Rev. Microb. 2011, 10, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, C.; Spanò, A.; Maugeri, T.L.; Poli, A.; Arena, A.; Nicolaus, B. Role of Bacterial Exopolysaccharides as Agents in Counteracting Immune Disorders Induced by Herpes Virus. Microorganisms 2015, 3, 464–483. [Google Scholar] [CrossRef]
- Vaningelgem, F.; Zamfir, M.; Mozzi, F.; Adriany, T.; Vancanneyt, M.; Swings, J.; de Vuyst, L. Biodiversity of exopolysaccharides produced by Streptococcus thermophilus strains is reflected in their production and their molecular and functional characteristics. Appl. Environ. Microb. 2004, 70, 900–912. [Google Scholar] [CrossRef]
- Ryder, C.; Byrd, M.; Wozniak, D.J. Role of exopolysaccharides in Pseudomonas aeruginosa biofilm development. Curr. Opin. Microbiol. 2007, 10, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Kumar, C.G.; Joo, H.S.; Choi, J.W.; Koo, Y.M.; Chang, C.S. Purification and characterization of an extracellular polysaccharide from haloalkalophilic Bacillus sp. I-450. Enzyme Microb. Tech. 2004, 34, 673–681. [Google Scholar] [CrossRef]
- Nadell, C.D.; Xavier, J.B.; Levin, S.A.; Foster, K.R. The Evolution of Quorum Sensing in Bacterial Biofilms. PLoS Biol 2008, 6, e14. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.H.; Koh, K.S.; Xie, C.; Tay, M.; Zhou, Y.; Williams, R.; Ng, W.J.; Rice, S.; Kjelleberg, S. The role of quorum sensing signalling in EPS production and the assembly of a sludge community into aerobic granules. ISME J. 2014, 8, 1186–1197. [Google Scholar] [CrossRef] [PubMed]
- Dobretsov, S.; Teplitski, M.; Paul, V. Mini review: Quorum sensing in the marine environment and its relationship to biofouling. Biofouling 2009, 25, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microb. 2001, 55, 165–199. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, A.; Burlingame, A.L.; Eberhard, C.; Kenyon, G.L.; Nealson, K.H.; Oppenheimer, N.J. Structural identification of autoinducer of Photobacterium fischeri luciferase. Biochemistry 1981, 20, 2444–2449. [Google Scholar] [CrossRef] [PubMed]
- Defroidt, T.; Boon, N.; Bossier, P.; Verstraete, W. Disruption of bacterial quorum sensing: An unexplored strategy to fight infections in aquaculture. Aquaculture 2004, 240, 69–88. [Google Scholar] [CrossRef]
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cel. Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef] [PubMed]
- Gram, L.; Grossart, H.P.; Schilingloff, A.; Kiroboe, T. Possible Quorum Sensing in Marine Snow Bacteria: Production of Acylated Homoserine Lactones by Roseobacter Strains Isolated from Marine Snow. Appl. Environ. Microb. 2002, 8, 4111–4116. [Google Scholar] [CrossRef]
- Frederick, M.R.; Kuttler, C.; Hense, B.; Eberl, H.J. A mathematical model of quorum sensing regulated EPS production in biofilm communities. Theor. Biol. Med. Mod. 2011, 8, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Dolfi, S.; Sveronis, A.; Silipo, A.; Rizzo, R.; Cescutti, P. A novel rhamno-mannan exopolysaccharide isolated from biofilms of Burkholderia multivorans C1576. Carbohydr. Res. 2015, 411, 42–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Santschi, P.H. Application of cross-flow ultrafiltration for isolating exopolymeric substances from a marine diatom (Amphora sp.). Limnol. Oceanogr. Methods 2009, 7, 419–429. [Google Scholar] [CrossRef]
- Bergmaier, D.; Lacroix, C.; Macedo, M.G.; Champagne, C.P. New method for exopolysaccharide determination in culture broth using stirred ultrafiltration cells. Appl. Microbiol. Biotechnol. 2001, 57, 401–406. [Google Scholar] [PubMed]
- Finore, I.; di Donato, P.; Mastascusa, V.; Nicolaus, B.; Poli, A. Fermentation Technologies for the Optimization of Marine Microbial Exopolysaccharide Production. Mar. Drugs 2014, 12, 3005–3024. [Google Scholar] [CrossRef] [PubMed]
- Senni, K.; Pereira, J.; Gueniche, F.; Delbarre-Ladrat, C.; Sinquin, C.; Ratiskol, J.; Godeau, G.; Fischer, A.; Helley, D.; Colliec-Jouault, S. Marine Polysaccharides: A Source of Bioactive Molecules for Cell Therapy and Tissue Engineering. Mar. Drugs 2011, 9, 1664–1681. [Google Scholar] [CrossRef] [PubMed]
- Laurienzo, P. Marine Polysaccharides in Pharmaceutical Applications: An Overview. Mar. Drugs 2010, 8, 2435–2465. [Google Scholar] [CrossRef] [PubMed]
- Courtois, A.; Berthou, C.; Guezennec, J.; Boisset, C.; Bordron, A. Exopolysaccharides Isolated from Hydrothermal Vent Bacteria Can Modulate the Complement System. PLoS ONE 2014, 9, e94965. [Google Scholar] [CrossRef] [PubMed]
- Lelchat, F.; Cozien, J.; Le Costaouec, T.; Brandilly, C.; Schmitt, S.; Baudoux, A.; Colliec-Jouault, S.; Boisset, C. Exopolysaccharide biosynthesis and biodegradation by a marine hydrothermal Alteromonas sp. Strain. Appl. Microb. Biotechnol. 2015, 99, 2637–2647. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Zhao, F.; Shi, M.; Zhang, X.; Zhou, B.; Zhang, Y.; Chen, X. Characterization and Biotechnological Analysis of a New Exopolysaccharide from the Arctic Marine Bacterium Polaribacter sp. SM1127. Nat. Sci. Rep. 2015, 5, 18435. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Liu, S.; Qiao, L.; Chen, X.; Pang, X.; Shi, M.; Zhang, X.; Qin, Q.; Zhou, B.; Zhang, Y.; et al. A novel exopolysaccharide from deep-sea bacterium Zunongwangia profunda SM-A87: Low-cost fermentation, moisture retention, and antioxidant activities. Appl. Microbiol. Biot. 2014, 98, 7437–7445. [Google Scholar] [CrossRef] [PubMed]
- Priyanka, P.; Arun, A.; Rekha, P. Sulfated exopolysaccharide produced by Labrenzia sp. PRIM-30, characterization and prospective applications. Int. J. Biol. Macromol. 2014, 69, 290–295. [Google Scholar]
- Lelchat, F.; Cerantola, S.; Brandily, C.; Colliec-Jouault, S.; Baudoux, A.; Ojima, T.; Boisset, C. The marine bacteria Cobetia marina DSMZ 4741 synthesizes an unexpected K-antigen-like exopolysaccharide. Carbohydr. Polym. 2015, 124, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Priyanka, P.; Arun, A.; Young, C.; Rekha, P. Prospecting exopolysaccharides produced by selected bacteria associated with marine organisms for biotechnological applications. Chin. J. Polym. Sci. 2015, 33, 236–244. [Google Scholar] [CrossRef]
- Hongpattarakere, T.; Cherntong, N.; Wichienchot, S.; Kolida, S.; Rastall, R. In vitro prebiotic evaluation of exopolysaccharides produced by marine isolated lactic acid bacteria. Carbohydr. Polym. 2012, 87, 846–852. [Google Scholar] [CrossRef]
- Deschatre, M.; Ghillebaert, F.; Guezennec, J.; Simon, C. Sorption of Copper(II) and Silver(I) by Four Bacterial Exopolysaccharides. Appl. Biochem. Biotechnol. 2013, 171, 1313–1327. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, P.; Melão, M.; Lombardi, A.; Nogueira, M.; Vieira, A. The effects of Anabaena spiroides exopolysaccharides on copper accumulation in an aquatic food chain. Aquat. Toxicol. 2009, 93, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Raj, R.; Dalei, K.; Chakraborty, J.; Das, S. Extracellular polymeric substances of a marine bacterium mediated synthesis of CdS nanoparticles for removal of cadmium from aqueous solution. J. Coll. Interf. Sci. 2016, 462, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Carrión, O.; Delgado, L.; Mercade, E. New emulsifying and cryoprotective exopolysaccharide from Antarctic Pseudomonas sp. ID1. Carbohydr. Polym. 2015, 117, 1028–1034. [Google Scholar] [CrossRef] [PubMed]
- Bramachari, P.; Kishor, P.K.; Ramadevi, R.; Kumar, R.; Rama, R.B.; Dubei, K. Isolation and Characterization of Mucous Exopolysaccharide (EPS) Produced by Vibrio furnissii Strain VB0S3. J. Microbiol. Biotechnol. 2007, 17, 44–51. [Google Scholar]
- Iyer, A.; Mody, K.; Jha, B. Enzyme and Emulsifying properties of a marine bacterial exopolysaccharide. Microb. Technol. 2006, 38, 220–222. [Google Scholar] [CrossRef]
- Liu, S.; Chen, X.; He, H.; Zhang, X.; Xie, B.; Yu, Y.; Chen, B.; Zhou, B.; Zhang, Y. Structure and Ecological Roles of a Novel Exopolysaccharide from the Arctic Sea Ice Bacterium Pseudoalteromonas sp. Strain SM20310. Appl. Env. Microbiol. 2013, 79, 224–230. [Google Scholar] [CrossRef] [PubMed]
- David, G.; Davies, M.; Parsek, R.; Pearson, J.P.; Iglewski, B.H.; Costerton, J.W.; Greenberg, E.P. The Involvement of Cell-to-Cell Signals in the Development of a Bacterial Biofilm. Science 1998, 280, 295–298. [Google Scholar] [CrossRef]
- Swift, S.; William, P.; Gsab, S. N-Acylhomoserine lactones and quorum sensing in proteobacteria. In Cell–Cell Signaling in Bacteria; Dunny, G.M., Winans, S.C., Eds.; ASM Press: Washington, D.C., USA, 1999; pp. 291–314. [Google Scholar]
- Bassler, B.L. A multichannel two-component signaling relay controls quorum sensing in Vibrio harveyi. In Cell–Cell Signaling in Bacteria; Dunny, G.M., Winans, S.C., Eds.; ASM Press: Washington, DC, USA, 1999; pp. 259–273. [Google Scholar]
- Tommonaro, G.; Abbamondi, G.R.; Iodice, C.; Tait, K.; de Rosa, S. Diketopiperazines Produced by the Halophilic Archaeon, Haloterrigena hispanica, Activate AHL Bioreporters. Microb. Ecol. 2012, 63, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Abbamondi, G.R.; de Rosa, S.; Iodice, C.; Tommonaro, G. Cyclic Dipeptides Produced by Marine Sponge-Associated Bacteria as Quorum Sensing Signals. Nat. Prod. Commun. 2014, 9, 229–232. [Google Scholar] [PubMed]
- Kendall, M.M.; Sperandio, V. What a Dinner Party! Mechanisms and Functions of Interkingdom Signaling in Host-Pathogen Associations. MBio 2016, 7, e01748-15. [Google Scholar] [CrossRef] [PubMed]
- Schikora, A.; Schenk, S.T.; Hartmann, A. Beneficial effects of bacteria-plant communication based on quorum sensing molecules of the N-acyl homoserine lactone group. Plant Mol. Biol. 2016, 90, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Nealson, K.H.; Platt, T.; Hastings, J.W. Cellular control of the synthesis and activity of the bacterial luminescence system. J. Bacteriol. 1970, 104, 313–322. [Google Scholar] [PubMed]
- Whiteley, M.; Lee, K.M.; Greenberg, E.P. Identification of genes controlled by quorum sensing in Pseudomonas aeruginosa. PNAS 1999, 96, 13904–13909. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.K.; Mangwani, N.; Rao, T.S.; Das, S. Biofilm-mediated bioremediation of polycyclic aromatic hydrocarbons. In Microbial Biodegradation and Bioremediation, 1st ed.; Das, S., Ed.; Elsevier: London, UK, 2014; pp. 203–232. [Google Scholar]
- De Kievit, T.R.; Gillis, R.; Marx, S.; Brown, C.; Iglewski, B.H. Quorum sensing genes in Pseudomonas aeruginosa biofilms: Their role and expression patterns. Appl. Environ. Microb. 2001, 67, 1865–1873. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zeng, Y.; Yu, Z.; Zhang, J.; Feng, H.; Lin, X. In silico and experimental methods revealed highly diverse bacteria with quorum sensing and aromatics biodegradation systems a potential broad application on bioremediation. Bioresour. Technol. 2013, 148, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Mangwani, N.; Kumari, S.; Das, S. Involvement of quorum sensing genes in biofilm development and degradation of polycyclic aromatic hydrocarbons by a marine bacterium Pseudomonas aeruginosa N6P6. Appl. Microb. Biotechnol. 2015, 99, 10283–10297. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.J.; Swift, S.; Kirke, D.F.; Keevil, C.W.; Dodd, C.E.R.; Williams, P. The regulation of biofilm development by quorum sensing in Aeromonas hydrophila. Environ. Microbiol. 2002, 4, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Husain, F.M.; Ahmad, I.; Khan, M.S.; Al-Shabib, N.A. Trigonella foenum-graceum (Seed) Extract Interferes with Quorum Sensing Regulated Traits and Biofilm Formation in the Strains of Pseudomonas aeruginosa and Aeromonas hydrophila. Evid. Based Complement. Alternat. Med. 2015. [Google Scholar] [CrossRef] [PubMed]
- Norizan, S.N.M.; Yin, W.F.; Chan, K.G. Caffeine as a potential quorum sensing inhibitor. Sensors 2013, 13, 5117–5129. [Google Scholar] [CrossRef] [PubMed]
- Fuqua, C.; Greenberg, E.P. Listening in on bacteria: Acyl-homoserine lactone signaling. Nat. Rev. 2002, 3, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, T.; Yin, W.F.; Chan, K.G. Inhibition of quorum sensing-controlled virulence factor production in Pseudomonas aeruginosa PAO1 by Ayurveda spice clove (Syzgium Aromaticum) bud extract. Sensors 2012, 12, 4016–4030. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.S.; Yunos, N.Y.; Tan, P.W.; Mohamad, N.I.; Adrian, T.G.; Yin, W.F.; Chan, K.G. Characterisation of a marine bacterium Vibrio brasiliensis T33 producing N-acyl homoserine lactone quorum sensing molecules. Sensors 2014, 14, 12104–12113. [Google Scholar] [CrossRef] [PubMed]
- Tan, P.W.; Tan, W.S.; Yunos, N.Y.; Mohamad, N.I.; Adrian, T.G.; Yin, W.F.; Chan, K.G. Short chain N-acyl homoserine lactone production in tropical marine Vibrio sinaloensis strain T47. Sensors 2014, 18, 12958–12967. [Google Scholar] [CrossRef] [PubMed]
- Brian-Jaisson, F.; Ortalo-Magné, A.; Guentas-Dombrowsky, L.; Armougom, F.; Blache, Y.; Molmeret, M. Identification of bacterial strains isolated from the Mediterranean Sea exhibiting different abilities of biofilm formation. Microb. Ecol. 2014, 68, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ayé, A.M.; Bonnin-Jusserand, M.; Brian-Jaisson, F.; Ortalo-Magné, A.; Culioli, G.; Nevry, R.K.; Rabah, N.; Blache, Y.; Molmeret, M. Modulation of violacein production and phenotypes associated with biofilm by exogenous quorum sensing N-acylhomoserine lactones in the marine bacterium Pseudoalteromonas ulvae TC14. Microbiol. 2015, 161, 2039–2051. [Google Scholar] [CrossRef]
- Long, R.A.; Azam, F. Antagonistic interactions among marine pelagic bacteria. Appl. Environ. Microbiol. 2001, 67, 4975–4983. [Google Scholar] [CrossRef] [PubMed]
- Kiørboe, T. Colonization of marine snow aggregates by invertebrate zooplankton: Abundance, scaling, and possible role. Limnol. Oceanogr. 2000, 45, 479–484. [Google Scholar] [CrossRef]
- Ploug, H.; Grossart, H.P. Bacterial growth and grazing on diatom aggregates: Respiratory carbon turnover as a function of aggregate size and sinking velocity. Limnol. Oceanogr. 2000, 45, 1467–1475. [Google Scholar] [CrossRef]
- Schweitzer, B.; Huber, I.; Amann, R.; Ludwig, W.; Simon, M. α- and β-Proteobacteria control the consumption and release of amino acids in lake snow aggregates. Appl. Environ. Microbiol. 2001, 67, 632–645. [Google Scholar] [CrossRef] [PubMed]
- Kjelleberg, S.; Molin, S. Is there a role for quorum sensing signals in bacterial biofilms? Cur. Opin. Microbiol. 2002, 5, 254–258. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Bacterial Source(EPS name) | Structure/Chemical Composition * | Activity/Applications | Ref. |
|---|---|---|---|
| Vibrio diabolicus, strain HE800T | Equal amounts of GlcA and hexosamine (GlcNAc and GalNAc) | Regenerating activity on bone and skin | [42] |
| Alteromonas infernus (GY785) | Repeating unit of uronic acids (GlcA and GalA) and neutral sugars (Gal and Glc) and substituted with one sulfate group | Increase the viability and the proliferation of chondrocytes. Cartilage tissue engineering applications. “Heparin-like” or “heparin-mimetic” activity (depolymerized and sulfated EPS). | [42,43] |
| Alteromonas macleodii subsp. Fijiensis, strain HYD657 | Monomer composition: Glc/Gal/Man/Rha/Fuc/GlcA/GalA (molar ratios: 1/1.9/0.4/0.6/0.2/1.2/2.8) | Protection of sensitive skin against chemical, mechanical and UVB aggressions | [45] |
| Polaribacter sp. SM1127 | Monomer composition: Rha/Fuc/GlcA/Man/Gal/Glc/GlcNAc (mol%: 0.8/7.4/21.4/23.4/17.3/1.6/28.0) | Food, cosmetic, pharmaceutical and biomedical fields | [46] |
| Weissella cibaria A2, Weissella confusa A9, Lactobacillus plantarum A3 and Pediococcus pentosaceus 5S4 | unknown | Prebiotic activity (Bifidobacteria group), high resistance to gastric and intestinal digestion | [51] |
| Paracoccus zeaxanthinifaciens subsp. payriae (EPS M1) | Proteins, neutral sugars, uronic acids, sulfates (w/w) 3%, 48%, 8%, 29%, respectively. Substituents: acetate and sulfate | Bioremediation of toxic metals | [52] |
| Anabaena spiroides | w/w composition: Glc 29.3%, Man 24.2%, Rha 21.9%, Xyl 7.8%, GlcA 6.6%, Fuc 5.6%, Gal 2.0%, GalA 1.8% and Ara 0.8%; proteins 12.2% | Bioremediation of metal-polluted environments (copper), reduction of copper accumulation in an aquatic microbial food chain | [53] |
| Pseudomonas aeruginosa JP-11 | unknown | Bioremediation of polluted waters by binding of toxic cadmium ions | [54] |
| Pseudomonas sp. ID1 | w/w composition: carbohydrates 33.8% (Glc 17.0%, Gal 8.6%, Fuc 8.2%)/uronic acids 2.4%/proteins 2.8% | Emulsifying and cryoprotectant properties. Potential applications in bioremediation and in food, pharmaceutical and cosmetic sectors. | [55] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Donato, P.; Poli, A.; Taurisano, V.; Abbamondi, G.R.; Nicolaus, B.; Tommonaro, G. Recent Advances in the Study of Marine Microbial Biofilm: From the Involvement of Quorum Sensing in Its Production up to Biotechnological Application of the Polysaccharide Fractions. J. Mar. Sci. Eng. 2016, 4, 34. https://doi.org/10.3390/jmse4020034
Di Donato P, Poli A, Taurisano V, Abbamondi GR, Nicolaus B, Tommonaro G. Recent Advances in the Study of Marine Microbial Biofilm: From the Involvement of Quorum Sensing in Its Production up to Biotechnological Application of the Polysaccharide Fractions. Journal of Marine Science and Engineering. 2016; 4(2):34. https://doi.org/10.3390/jmse4020034
Chicago/Turabian StyleDi Donato, Paola, Annarita Poli, Valentina Taurisano, Gennaro Roberto Abbamondi, Barbara Nicolaus, and Giuseppina Tommonaro. 2016. "Recent Advances in the Study of Marine Microbial Biofilm: From the Involvement of Quorum Sensing in Its Production up to Biotechnological Application of the Polysaccharide Fractions" Journal of Marine Science and Engineering 4, no. 2: 34. https://doi.org/10.3390/jmse4020034