
The Distribution of Pseudodiaptomus marinus in European and Neighbouring Waters—A Rolling Review

 , , , , ,
, , , , ,  , , , , ,
, , , , ,  , , , and add
Show full author list
, , , and add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
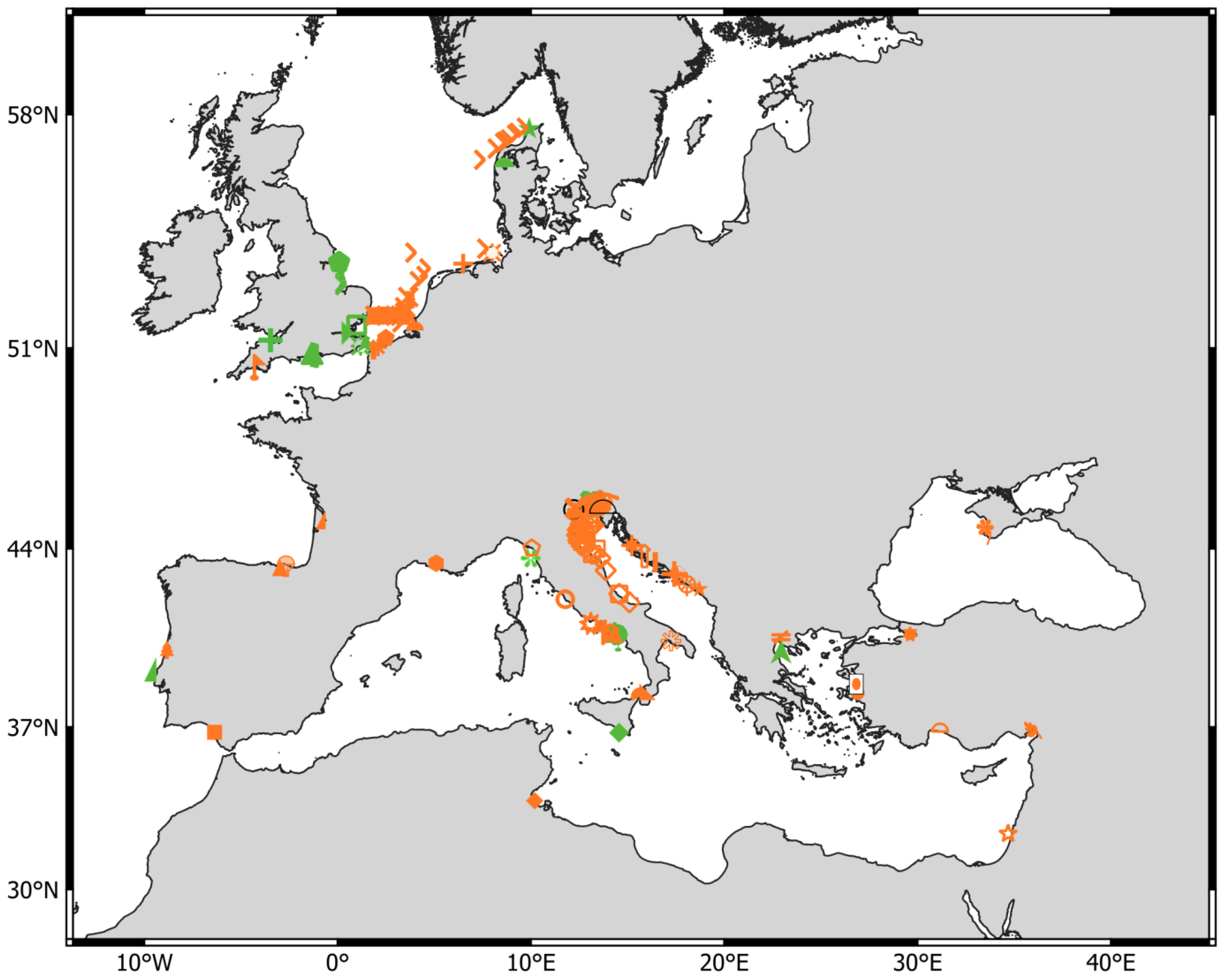
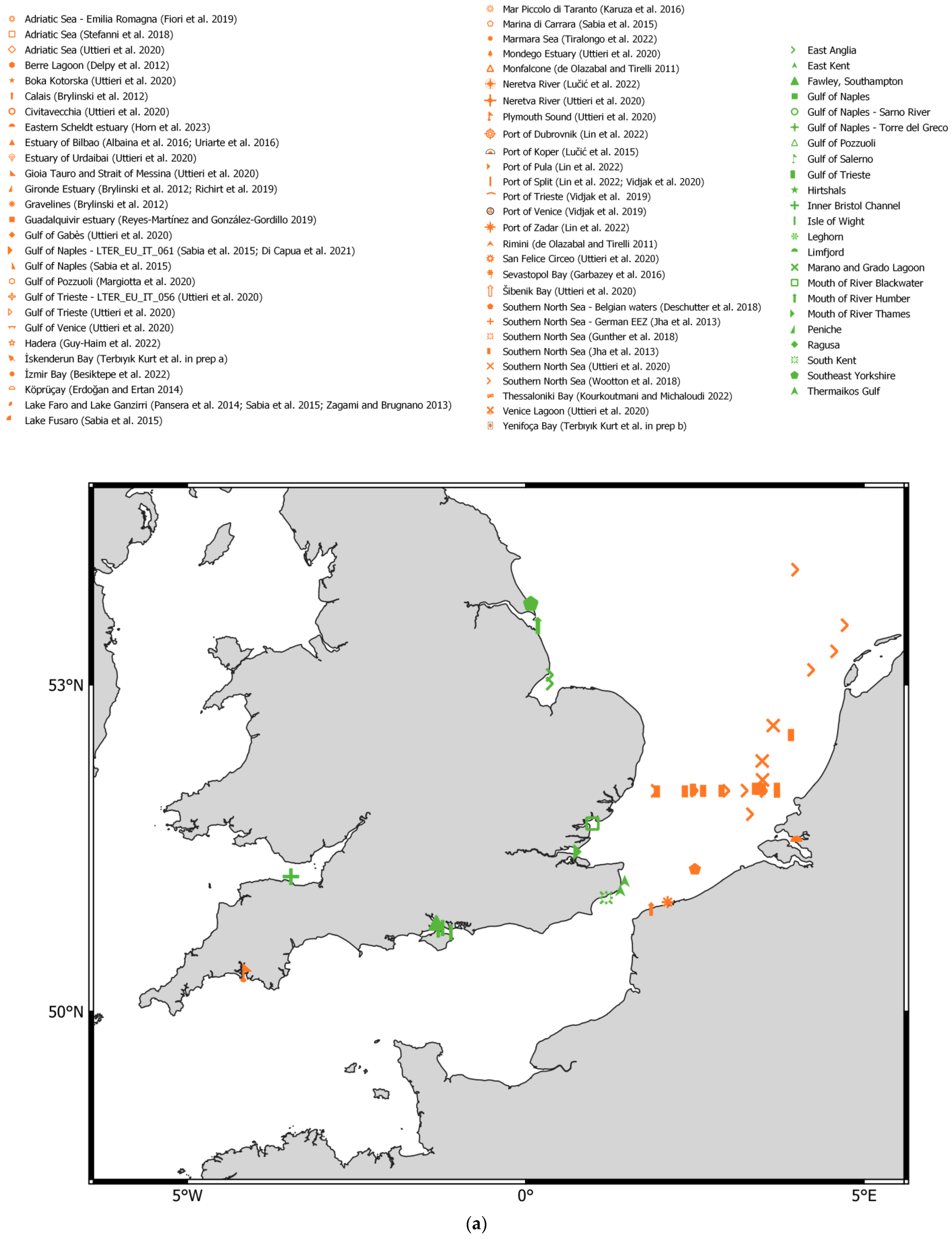
3. Results
3.1. Pseudodiaptomus marinus Records from Published Literature
- -
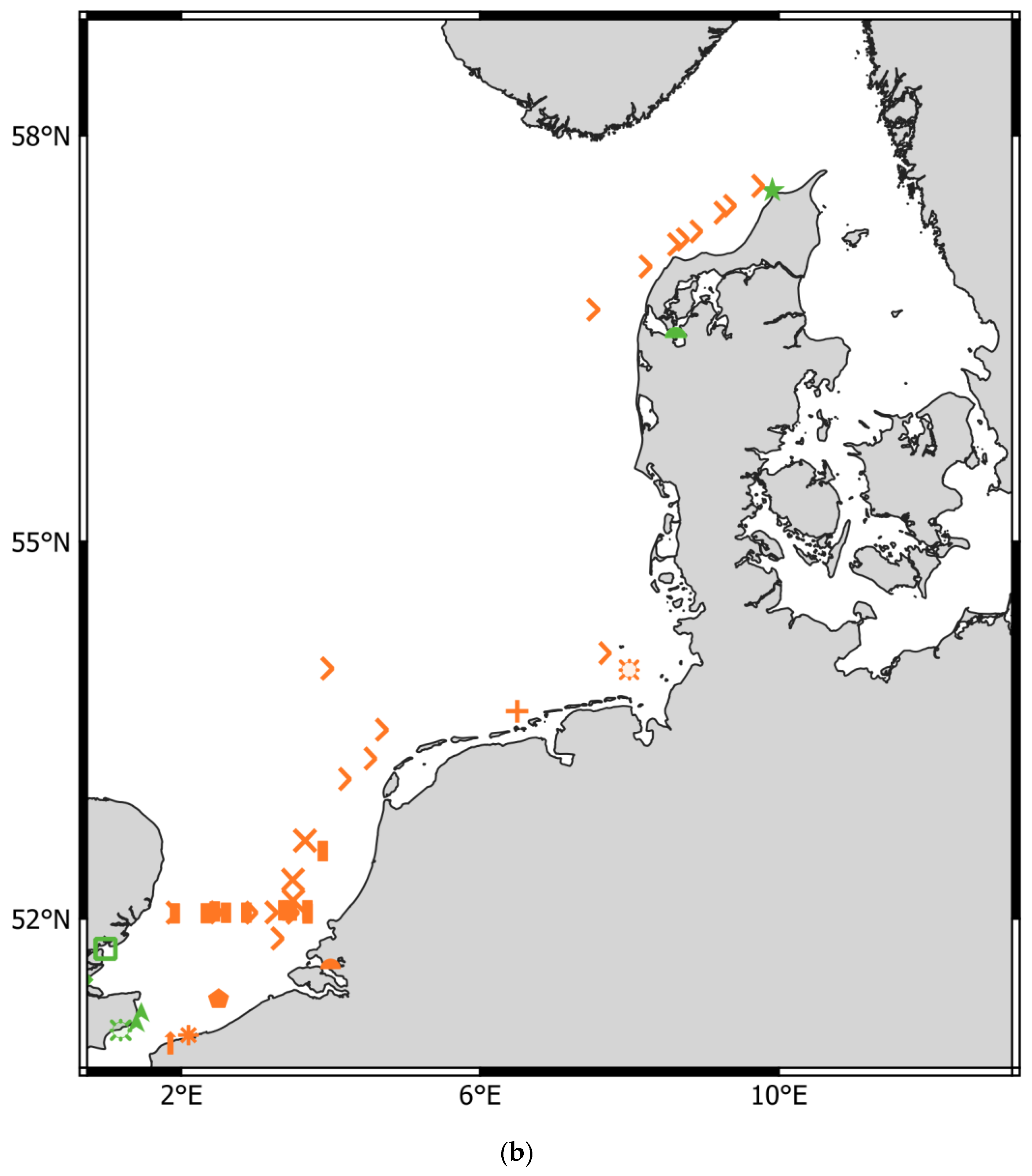
- Greater North Sea
- -
- Western Mediterranean Sea
- -
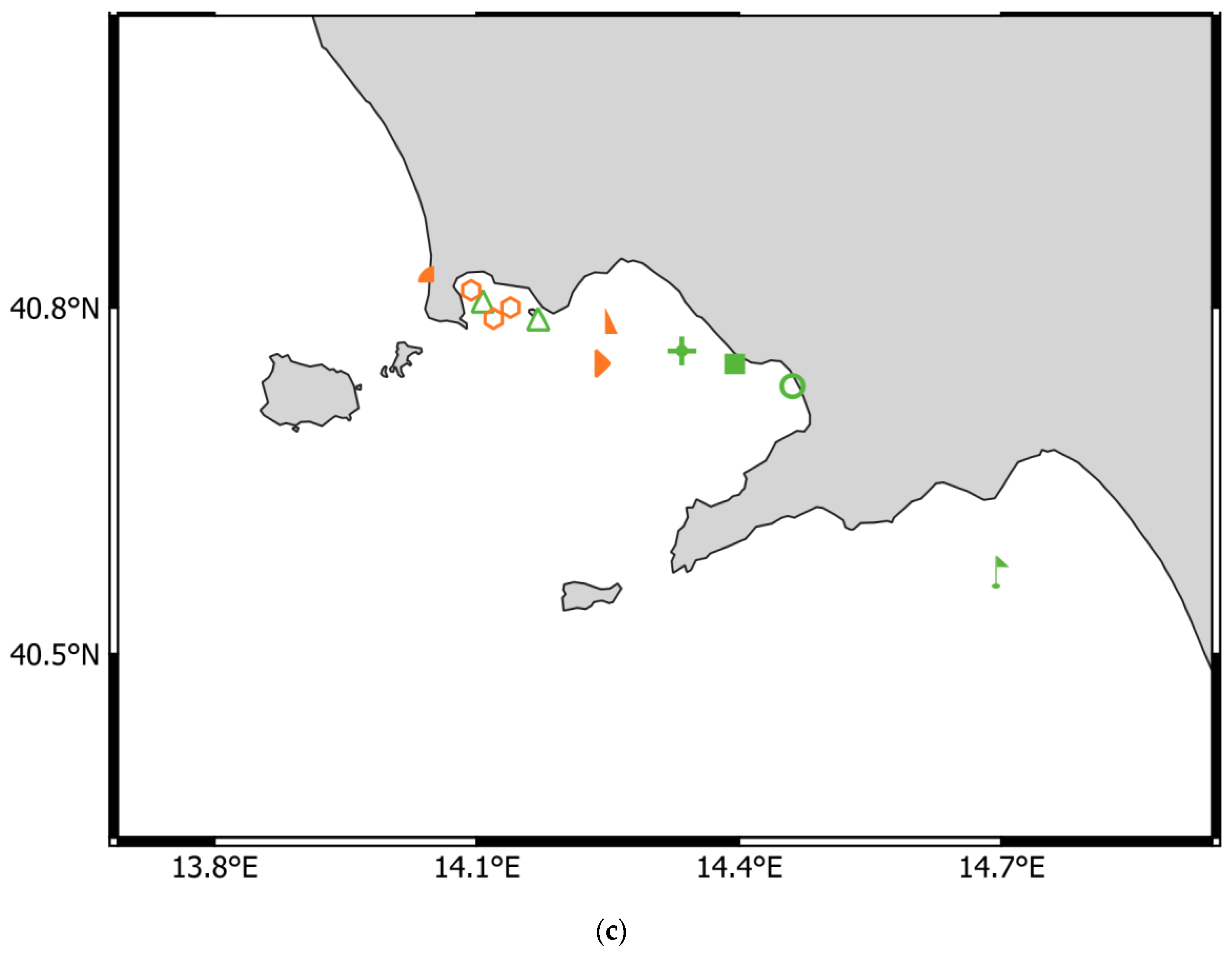
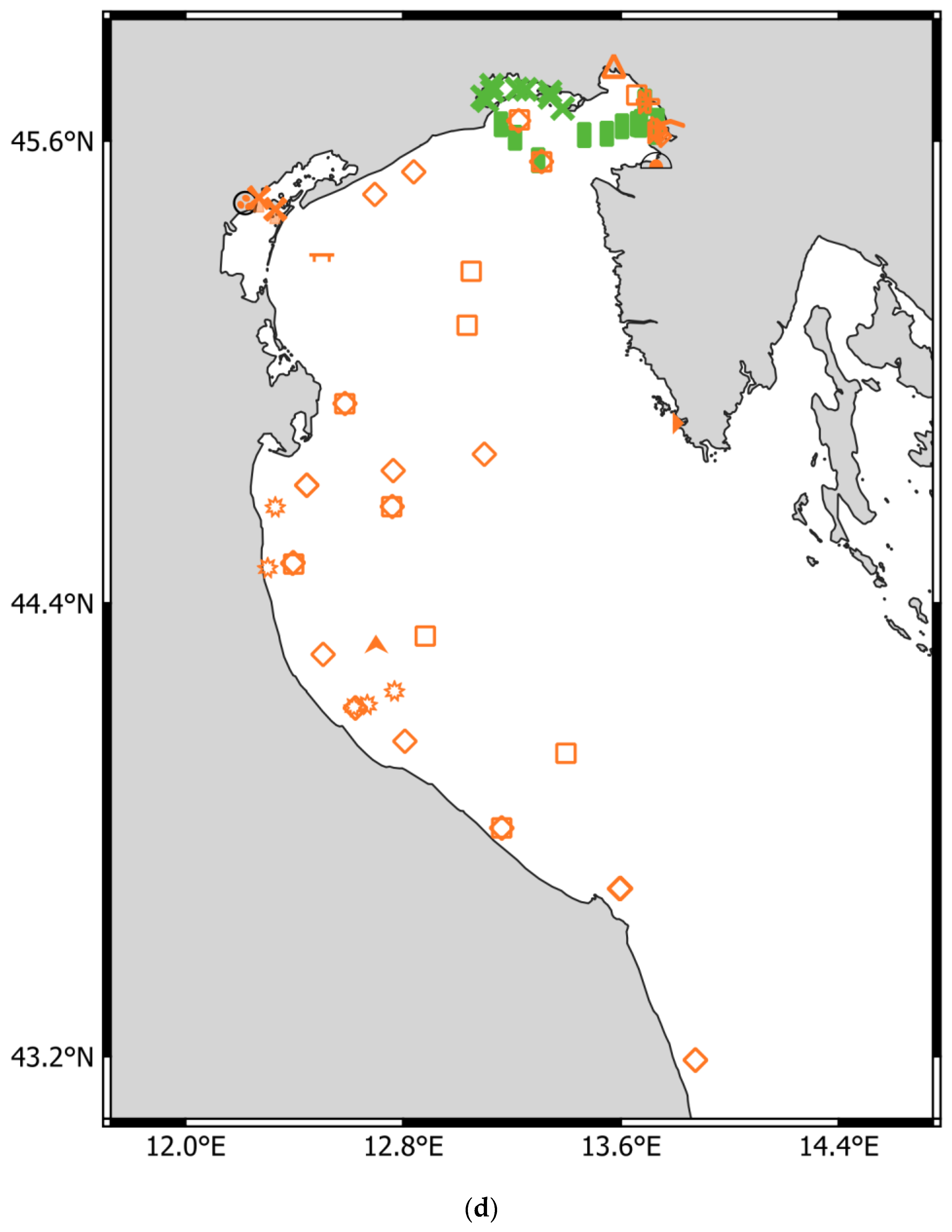
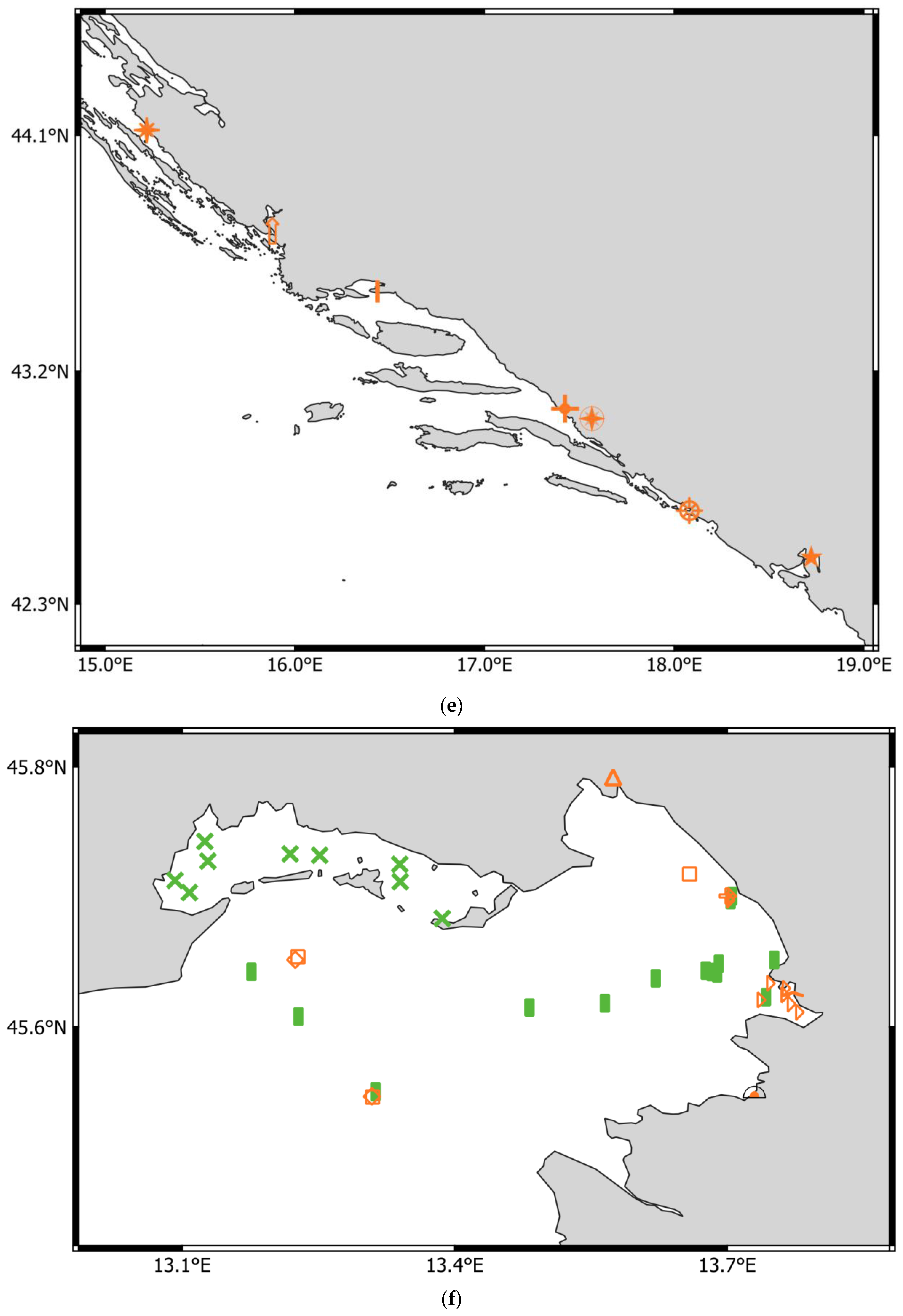
- Adriatic Sea
- -
- Aegean-Levantine Sea
- -
- Black Sea
3.2. New Records of Pseudodiaptomus marinus in ENW
- -
- Bay of Biscay and the Iberian Coast
- -
- Celtic Seas
- -
- Greater North Sea
- -
- Western Mediterranean Sea
- -
- Ionian Sea and the Central Mediterranean Sea
- -
- Adriatic Sea
- -
- Aegean-Levantine Sea
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geburzi, J.C.; McCarthy, M.L. How do they do it?—Understanding the success of marine invasive species. In YOUMARES 8—Oceans Across Boundaries: Learning from Each Other; Jungblut, S., Liebich, V., Bode, M., Eds.; Springer: Cham, Switzerland, 2018; pp. 109–124. [Google Scholar]
- Simberloff, D. Non-native invasive species and novel ecosystems. F1000Prime Rep. 2015, 7, 47. [Google Scholar] [CrossRef]
- Dexter, E.; Bollens, S.M. Zooplankton invasions in the early 21st century: A global survey of recent studies and recommendations for future research. Hydrobiologia 2020, 847, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.E. Evolutionary mechanisms of habitat invasions, using the copepod Eurytemora affinis as a model system. Evol. Appl. 2016, 9, 248–270. [Google Scholar] [CrossRef] [PubMed]
- Gollasch, S.; Lenz, J.; Dammer, M.; Andres, H.-G. Survival of tropical ballast water organisms during a cruise from the Indian Ocean to the North Sea. J. Plankton Res. 2000, 22, 923–937. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.H.; Kimmerer, W.; Smith, G.; Ruiz, G.M.; Lion, K. Post-exchange zooplankton in ballast water of ships entering the San Francisco Estuary. J. Plankton Res. 2005, 27, 707–714. [Google Scholar] [CrossRef] [Green Version]
- Cabrini, M.; Cerino, F.; de Olazabal, A.; Di Poi, E.; Fabbro, C.; Fornasaro, D.; Goruppi, A.; Flander-Putrle, V.; France, J.; Gollasch, S.; et al. Potential transfer of aquatic organisms via ballast water with a particular focus on harmful and non-indigenous species: A survey from Adriatic ports. Mar. Poll. Bull. 2019, 147, 16–35. [Google Scholar] [CrossRef] [PubMed]
- Velasquez, X.; Morov, A.R.; Terbıyık Kurt, T.; Meron, D.; Guy-Haim, T. Two-way bioinvasion: Tracking the neritic non-native cyclopoid copepods Dioithona oculata and Oithona davisae (Oithonidae) in the Eastern Mediterranean Sea. Mediterr. Mar. Sci. 2021, 22, 586–602. [Google Scholar] [CrossRef]
- Zagami, G.; Brugnano, C.; Granata, A.; Guglielmo, L.; Minutoli, R.; Aloise, A. Biogeographical distribution and ecology of the planktonic copepod Oithona davisae: Rapid invasion in Lakes Faro and Ganzirri (Central Mediterranean Sea). In Trends in Copepod Studies—Distribution, Biology and Ecology; Uttieri, M., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2018; pp. 59–82. [Google Scholar]
- Feis, M.E.; Goedknegt, M.A.; Arzul, I.; Chenuil, A.; den Boon, O.; Gottschalck, L.; Kondo, Y.; Ohtsuka, S.; Shama, L.N.S.; Thieltges, D.W.; et al. Global invasion genetics of two parasitic copepods infecting marine bivalves. Sci. Rep. 2019, 9, 12730. [Google Scholar] [CrossRef] [Green Version]
- Terbıyık Kurt, T.; Beşiktepe, Ş. First distribution record of the invasive copepod Oithona davisae Ferrari and Orsi, 1984, in the coastal waters of the Aegean Sea. Mar. Ecol. 2019, 40, e12548. [Google Scholar] [CrossRef]
- Pansera, M.; Camatti, E.; Schroeder, A.; Zagami, G.; Bergamasco, A. The non-indigenous Oithona davisae in a Mediterranean transitional environment: Coexistence patterns with competing species. Sci. Rep. 2021, 11, 8341. [Google Scholar] [CrossRef]
- Dragičević, B.; Anadoli, O.; Angel, D.; Benabdi, M.; Bitar, G.; Castriota, L.; Crocetta, F.; Deidun, A.; Dulčić, J.; Edelist, D.; et al. New Mediterranean biodiversity records (December 2019). Mediterr. Mar. Sci. 2019, 20, 645–656. [Google Scholar] [CrossRef] [Green Version]
- Bollens, S.M.; Breckenridge, J.K.; Cordell, J.R.; Rollwagen-Bollens, G.; Kalata, O. Invasive copepods in the Lower Columbia River Estuary: Seasonal abundance, co-occurrence and potential competition with native copepods. Aquat. Invasions 2012, 7, 101–109. [Google Scholar] [CrossRef]
- Bouley, P.; Kimmerer, W.J. Ecology of a highly abundant, introduced cyclopoid copepod in a temperate estuary. Mar. Ecol. Prog. Ser. 2006, 324, 219–228. [Google Scholar] [CrossRef]
- Barroeta, Z.; Villate, F.; Uriarte, I.; Iriarte, A. Impact of colonizer copepods on zooplankton structure and diversity in contrasting estuaries. Estuaries Coasts 2022, 45, 2592–2609. [Google Scholar] [CrossRef]
- Camatti, E.; Pansera, M.; Bergamasco, A. The copepod Acartia tonsa Dana in a microtidal Mediterranean lagoon: History of a successful invasion. Water 2019, 11, 1200. [Google Scholar] [CrossRef] [Green Version]
- Cordell, J.R.; Rasmussen, M.; Bollens, S.M. Biology of the introduced copepod Pseudodiaptomus inopinus in a northeastern Pacific estuary. Mar. Ecol. Prog. Ser. 2007, 333, 213–227. [Google Scholar] [CrossRef]
- Adams, J.B.; Bollens, S.M.; Bishop, J.G. Predation on the invasive copepod, Pseudodiaptomus forbesi, and native zooplankton in the lower Columbia River: An experimental approach to quantify differences in prey-specific feeding rates. PLoS ONE 2015, 10, e0144095. [Google Scholar] [CrossRef] [PubMed]
- de Olazabal, A.; Tirelli, V. First record of the egg-carrying calanoid copepod Pseudodiaptomus marinus in the Adriatic Sea. Mar. Biodivers. Rec. 2011, 4, e85. [Google Scholar] [CrossRef]
- Sabia, L.; Zagami, G.; Mazzocchi, M.G.; Zambianchi, E.; Uttieri, M. Spreading factors of a globally invading coastal copepod. Mediterr. Mar. Sci. 2015, 16, 460–471. [Google Scholar] [CrossRef] [Green Version]
- Uttieri, M.; Aguzzi, L.; Aiese Cigliano, R.; Amato, A.; Bojanić, N.; Brunetta, M.; Camatti, E.; Carotenuto, Y.; Damjanović, T.; Delpy, F.; et al. WGEUROBUS—Working Group “Towards a EURopean OBservatory of the non-indigenous calanoid copepod Pseudodiaptomus marinus”. Biol. Invasions 2020, 22, 885–906. [Google Scholar] [CrossRef]
- Svetlichny, L.; Hubareva, E.; Khanaychenko, A.; Uttieri, M. Response to salinity and temperature changes in the alien Asian copepod Pseudodiaptomus marinus introduced in the Black Sea. J. Exp. Zool. A 2019, 331, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Sabia, L.; Uttieri, M.; Schmitt, F.G.; Zagami, G.; Zambianchi, E.; Souissi, S. Pseudodiaptomus marinus Sato, 1913, a new invasive copepod in Lake Faro (Sicily): Observations on the swimming behaviour and the sex-dependent responses to food. Zool. Stud. 2014, 53, 49. [Google Scholar] [CrossRef] [Green Version]
- Sabia, L.; Di Capua, I.; Percopo, I.; Uttieri, M.; Amato, A. ITS2 in calanoid copepods: Reconstructing phylogenetic relationships and identifying a newly introduced species in the Mediterranean. Eur. Zool. J. 2017, 84, 104–115. [Google Scholar] [CrossRef] [Green Version]
- Brylinski, J.M.; Antajan, E.; Raud, T.; Vincent, D. First record of the Asian copepod Pseudodiaptomus marinus Sato, 1913 (Copepoda: Calanoida: Pseudodiaptomidae) in the southern bight of the North Sea along the coast of France. Aquat. Invasions 2012, 7, 577–584. [Google Scholar] [CrossRef]
- European Environment Agency. Marine Regions and Subregions. Available online: https://www.eea.europa.eu/data-and-maps/figures/marine-regions-and-subregions (accessed on 19 April 2023).
- Galanidi, M.; Zenetos, A. Data-driven recommendations for establishing threshold values for the NIS trend indicator in the Mediterranean Sea. Diversity 2022, 14, 57. [Google Scholar] [CrossRef]
- Tsiamis, K.; Palialexis, A.; Stefanova, K.; Gladan, Ž.N.; Skejić, S.; Despalatović, M.; Cvitković, I.; Dragičević, B.; Dulčić, J.; Vidjak, O.; et al. Non-indigenous species refined national baseline inventories: A synthesis in the context of the European Union’s Marine Strategy Framework Directive. Mar. Pollut. Bull. 2019, 145, 429–435. [Google Scholar] [CrossRef]
- Horn, H.G.; van Rijswijk, P.; Soetaert, K.; van Oevelen, D. Drivers of spatial and temporal micro- and mesozooplankton dynamics in an estuary under strong anthropogenic influences (the Eastern Scheldt, Netherlands). J. Sea Res. 2023, 192, 102357. [Google Scholar] [CrossRef]
- Di Capua, I.; Piredda, R.; Mazzocchi, M.G.; Zingone, A. Metazoan diversity and seasonality through eDNA metabarcoding at a Mediterranean long-term ecological research site. ICES J. Mar. Sci. 2021, 78, 3303–3316. [Google Scholar] [CrossRef]
- Margiotta, F.; Balestra, C.; Buondonno, A.; Casotti, R.; D’Ambra, I.; Di Capua, I.; Gallia, R.; Mazzocchi, M.G.; Merquiol, L.; Pepi, M.; et al. Do plankton reflect the environmental quality status? The case of a post-industrial Mediterranean Bay. Mar. Environ. Res. 2020, 160, 104980. [Google Scholar] [CrossRef]
- Fiori, E.; Benzi, M.; Ferrari, C.R.; Mazziotti, C. Zooplankton community structure before and after Mnemiopsis leidyi arrival. J. Plankton Res. 2019, 41, 803–820. [Google Scholar] [CrossRef]
- Itoh, H.; Tcachibana, A.; Nomura, H.; Tanaka, Y.; Furota, T.; Ishimaru, T. Vertical distribution of planktonic copepods in Tokyo Bay in summer. Plankton Benthos Res. 2011, 6, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Vidjak, O.; Damjanović, T.; Rožić, S.; Šegvić Bubić, T.; Bojanić, N.; Hrabar, J.; Arapov, J.; Skračić, M. Spreading of the non-indigenous Indo-Pacific copepod Pseudodiaptomus marinus Sato, 1913 in eastern Adriatic coastal and transitional waters. In 11th International Conference on Biological Invasions. The Human Role in Biological Invasions—A Case of Dr Jekyll and Mr Hyde? Jelaska, S.D., Ed.; Croatian Ecological Society: Zagreb, Croatia, 2020; p. 95. [Google Scholar]
- Lin, Y.; Vidjak, O.; Ezgeta-Balić, D.; Bojanić Varezić, D.; Šegvić-Bubić, T.; Stagličić, N.; Zhan, A.; Briski, E. Plankton diversity in Anthropocene: Shipping vs. aquaculture along the eastern Adriatic coast assessed through DNA metabarcoding. Sci. Total Environ. 2022, 807, 151043. [Google Scholar] [CrossRef]
- Lučić, D.; Onofri, I.; Garić, R.; Violić, I.; Vranješ, M.; Gangai Zovko, B.; Jurinović, J.; Njire, J.; Hure, M. Ingression of the hydromedusa Neotima lucullana (Delle Chiaje, 1822) into the ecosystem of the Neretva river estuary (south-eastern Adriatic, Croatia). Acta Adriat. 2022, 63, 165–174. [Google Scholar] [CrossRef]
- Glamuzina, B.; Tutman, P.; Glamuzina, L.; Vidović, Z.; Simonović, P.; Vilizzi, L. Quantifying current and future risks of invasiveness of non-native aquatic species in highly urbanised estuarine ecosystems—A case study of the River Neretva Estuary (Eastern Adriatic Sea: Croatia and Bosnia–Herzegovina). Fish. Manag. Ecol. 2021, 28, 138–146. [Google Scholar] [CrossRef]
- Njire, J.; Bojanić, N.; Lučić, D.; Violić, I. First record of the alien tintinnid ciliate Rhizodomus tagatzi Strelkow and Wirketis 1950 in the Adriatic Sea. Water 2023, 15, 1821. [Google Scholar] [CrossRef]
- Kourkoutmani, P.; Michaloudi, E. First record of the calanoid copepod Pseudodiaptomus marinus Sato, 1913 in the North Aegean Sea, in Thessaloniki Bay, Greece. BioInvasions Rec. 2022, 11, 738–746. [Google Scholar] [CrossRef]
- Krestenitis, Y.N.; Kombiadou, K.D.; Androulidakis, Y.S. Interannual variability of the physical characteristics of North Thermaikos Gulf (NW Aegean Sea). J. Mar. Syst. 2012, 96–97, 132–151. [Google Scholar] [CrossRef]
- Hyder, P.; Simpson, J.H.; Christopoulos, S.; Krestenitis, Y. The seasonal cycles of stratification and circulation in the Thermaikos Gulf Region of Freshwater Influence (ROFI), north-west Aegean. Cont. Shelf Res. 2002, 22, 2573–2597. [Google Scholar] [CrossRef]
- Angelidis, A. Fulvia fragilis (Forsskal in Niebuhr, 1775) (Bivalvia: Cardiidae), first record of an alien mollusk in the Gulf of Thessaloniki (Inner Thermaikos Gulf, North Aegean Sea, Greece). J. Biol. Res. 2013, 20, 228–232. [Google Scholar]
- Besiktepe, S.; Terbıyık Kurt, T.; Gubanova, A. Mesozooplankton composition and distribution in İzmir Bay, Aegean Sea: With special emphasis on copepods. Reg. Stud. Mar. Sci. 2022, 55, 102567. [Google Scholar] [CrossRef]
- Alyuruk, H.; Kontas, A. Seasonal variations and distributions of dissolved free and total carbohydrates at the İzmir Bay, Aegean Sea. Acta Oceanol. Sin. 2018, 37, 6–14. [Google Scholar] [CrossRef]
- Terbıyık Kurt, T.; Beşiktepe, S.; Velasquez, X.; Guy-Haim, T. New Record of Pseudodiaptomus marinus in the Yenifoça Bay (Aegean Sea); Department of Marine Biology, Faculty of Fisheries, Çukurova University: Adana, Turkey, in prep.
- TÜBİTAK-MRC and İzmir Metropolitan Municipality. Monitoring of İzmir Bay Water Quality and Terrestrial Inputs and Developing Recommendations for the Pollution Prevention (İZİZ); TÜBİTAK-MAM ve İzmir Metropolitan Municipality: Kocaeli, Turkey, 2023; p. 522T203. [Google Scholar]
- Guy-Haim, T.; Velasquez, X.; Terbiyik-Kurt, T.; Di Capua, I.; Mazzocchi, M.G.; Morov, A.R. A new record of the rapidly spreading calanoid copepod Pseudodiaptomus marinus (Sato, 1913) in the Levantine Sea using multi-marker metabarcoding. BioInvasions Rec. 2022, 11, 964–976. [Google Scholar] [CrossRef]
- Terbıyık Kurt, T.; Beşiktepe, Ş.; Velasquez, X.; Guy-Haim, T. Recent Introduction of Non-Indigeneous Copepods Species Pseudodiaptomus marinus in the İskenderun Bay (Northeastern Mediterranean); Department of Marine Biology, Faculty of Fisheries, Çukurova University: Adana, Turkey, in prep.
- Ministry of Environment Urbanization and Climate Change TÜBITAK-MRC. Integrated Marine Pollution Monitoring 2020–2022 Program: 2022, Mediterranean Sea Report; Ministry of Environment Urbanization and Climate Change TÜBITAK-MRC: Kocaeli, Turkey, 2023.
- Polat, S.; Uysal, Z. Abundance and biomass of picoplanktonic Synechococcus (Cyanobacteria) in a coastal ecosystem of the northeastern Mediterranean, the Bay of İskenderun. Mar. Biol. Res. 2009, 5, 363–373. [Google Scholar] [CrossRef]
- Terbıyık Kurt, T. Contribution and acclimatization of the swarming tropical copepod Dioithona oculata (Farran, 1913) in a Mediterranean coastal ecosystem. Turk. J. Zool. 2018, 42, 567–577. [Google Scholar] [CrossRef]
- Terbiyik, T.; Cevik, C.; Toklu-Alicli, B.; Sarihan, E. First record of Ferosagitta galerita (Dallot, 1971) [Chaetognatha] in the Mediterranean Sea. J. Plankton Res. 2007, 29, 721–726. [Google Scholar] [CrossRef] [Green Version]
- Tiralongo, F.; Akyol, O.; Al Mabruk, S.A.A.; Battaglia, P.; Beton, D.; Bitls, B.; Borg, J.A.; Bouchoucha, M.; Çinar, M.E.; Crocetta, F.; et al. New alien Mediterranean biodiversity records (August 2022). Mediterr. Mar. Sci. 2022, 23, 725–747. [Google Scholar] [CrossRef]
- Ergül, H.A. Evaluation of seasonal physicochemical conditions and chlorophyll-a concentrations in Izmit Bay, Marmara Sea. J. Black Sea Mediterr. Environ. 2016, 22, 201–217. [Google Scholar]
- Tan, I.; Beken, Ç.P.; Öncel, S. Pressure-impact analysis of the coastal waters of Marmara Sea. Fresenius Environ. Bull. 2017, 26, 2689–2699. [Google Scholar]
- Ergul, H.A.; Balkis-Ozdelice, N.; Koral, M.; Aksan, S.; Durmus, T.; Kaya, M.; Kayal, M.; Ekmekci, F.; Canli, O. The early stage of mucilage formation in the Marmara Sea during spring 2021. J. Black Sea Mediterr. Environ. 2021, 27, 232–257. [Google Scholar]
- Karadurmuş, U.; Sarı, M. Marine mucilage in the Sea of Marmara and its effects on the marine ecosystem: Mass deaths. Turk. J. Zool. 2022, 46, 93–102. [Google Scholar] [CrossRef]
- Leitão, F.; Baptista, V.; Vieira, V.; Silva, P.L.; Relvas, P.; Teodósio, M.A. A 60-year time series analyses of the upwelling along the Portuguese coast. Water 2019, 11, 1285. [Google Scholar] [CrossRef] [Green Version]
- Battuello, M.; Brizio, P.; Mussat Sartor, R.; Nurra, N.; Pessani, D.; Abete, M.C.; Squadrone, S. Zooplankton from a North Western Mediterranean area as a model of metal transfer in a marine environment. Ecol. Indic. 2016, 66, 440–451. [Google Scholar] [CrossRef]
- Donnarumma, L.; Sandulli, R.; Appolloni, L.; Ferrigno, F.; Rendina, F.; Di Stefano, F.; Russo, G.F. Bathymetrical and temporal variations in soft-bottom molluscan assemblages in the coastal area facing the Sarno River mouth (Mediterranean Sea, Gulf of Naples). Ecol. Quest. 2020, 31, 53–65. [Google Scholar] [CrossRef]
- Marino, D.; Modigh, M.; Zingone, A. General features of phytoplankton communities and primary production in the Gulf of Naples and adjacent waters. In Marine Phytoplankton Productivity. Lecture Notes on Coastal and Estuarine Studies; Holm-Hansen, O., Bolis, L., Gilles, R., Eds.; Springer: Berlin/Heidelberg, Germany, 1984; pp. 89–100. [Google Scholar]
- Reyes Suarez, N.C.; Cook, M.S.; Gačić, M.; Paduan, J.D.; Drago, A.; Cardin, V. Sea surface circulation structures in the Malta-Sicily Channel from remote sensing data. Water 2019, 11, 1589. [Google Scholar] [CrossRef] [Green Version]
- Consorzio di Sviluppo Economico del Friuli. Porto Nogaro. Available online: https://www.cosef.fvg.it/zona-industriale-aussa-corno/porto-nogaro.html (accessed on 19 April 2023).
- Leray, M.; Yang, J.Y.; Meyer, C.P.; Mills, S.C.; Agudelo, N.; Ranwez, V.; Boehm, J.T.; Machida, R.J. A new versatile primer set targeting a short fragment of the mitochondrial COI region for metabarcoding metazoan diversity: Application for characterizing coral reef fish gut contents. Front. Zool. 2013, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Geller, J.; Meyer, C.; Parker, M.; Hawk, H. Redesign of PCR primers for mitochondrial cytochrome c oxidase subunit I for marine invertebrates and application in all-taxa biotic surveys. Mol. Ecol. Resour. 2013, 13, 851–861. [Google Scholar] [CrossRef]
- Androulidakis, Y.; Kolovoyiannis, V.; Makris, C.; Krestenitis, Y.; Baltikas, V.; Stefanidou, N.; Chatziantoniou, A.; Topouzelis, K.; Moustaka-Gouni, M. Effects of ocean circulation on the eutrophication of a Mediterranean gulf with river inlets: The Northern Thermaikos Gulf. Cont. Shelf Res. 2021, 221, 104416. [Google Scholar] [CrossRef]
- Garbazey, O.A.; Popova, E.V.; Gubanova, A.D.; Altukov, D.A. First record of the occurrence of Pseudodiaptomus marinus (Copepoda: Calanoida: Pseudodiaptomidae) in the Black Sea (Sevastopol Bay). Mar. Biol. J. 2016, 1, 78–80. [Google Scholar] [CrossRef]
- Zenetos, A.; Tsiamis, K.; Galanidi, M.; Carvalho, N.; Bartilotti, C.; Canning-Clode, J.; Castriota, L.; Chainho, P.; Comas-González, R.; Costa, A.C.; et al. Status and trends in the rate of introduction of marine non-indigenous species in European seas. Diversity 2022, 14, 1077. [Google Scholar] [CrossRef]
- Zenetos, A.; Gofas, S.; Verlaque, M.; Cinar, M.E.; García Raso, J.E.; Bianchi, C.N.; Morri, C.; Azzurro, E.; Bilecenoglu, M.; Froglia, C.; et al. Alien species in the Mediterranean Sea by 2010. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part I. Spatial distribution. Mediterr. Mar. Sci. 2010, 11, 381–493. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.A.; Chain, F.J.J.; Zhan, A.; MacIsaac, H.J.; Cristescu, M.E. Early detection of aquatic invaders using metabarcoding reveals a high number of non-indigenous species in Canadian ports. Divers. Distrib. 2016, 22, 1045–1059. [Google Scholar] [CrossRef] [Green Version]
- Di Capua, I.; D’Angiolo, R.; Piredda, R.; Minucci, C.; Boero, F.; Uttieri, M.; Carotenuto, Y. From phenotypes to genotypes and back: Toward an integrated evaluation of biodiversity in calanoid copepods. Front. Mar. Sci. 2022, 9, 833089. [Google Scholar] [CrossRef]
- Albaina, A.; Uriarte, I.; Aguirre, M.; Abad, D.; Iriarte, A.; Villate, F.; Estonba, A. Insights on the origin of invasive copepods colonizing Basque estuaries; a DNA barcoding approach. Mar. Biodivers. Rec. 2016, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- Abad, D.; Albaina, A.; Aguirre, M.; Laza-Martínez, A.; Uriarte, I.; Iriarte, A.; Villate, F.; Estonba, A. Is metabarcoding suitable for estuarine plankton monitoring? A comparative study with microscopy. Mar. Biol. 2016, 163, 149. [Google Scholar] [CrossRef]
- Günther, B.; Knebelsberger, T.; Neumann, H.; Laakmann, S.; Martinez Arbizu, P. Metabarcoding of marine environmental DNA based on mitochondrial and nuclear genes. Sci. Rep. 2018, 8, 14822. [Google Scholar] [CrossRef] [Green Version]
- Ohtsuka, S.; Shimono, T.; Hanyuda, T.; Shang, X.; Huang, C.; Soh, H.Y.; Kimmerer, W.; Kawai, H.; Itoh, H.; Ishimaru, T.; et al. Possible origins of planktonic copepods, Pseudodiaptomus marinus (Crustacea: Copepoda; Calanoida), introduced from East Asia to the San Francisco Estuary based on a molecular analysis. Aquat. Invasions 2018, 13, 221–230. [Google Scholar] [CrossRef]
- Stefanni, S.; Stanković, D.; Borme, D.; de Olazabal, A.; Juretić, T.; Pallavicini, A.; Tirelli, V. Multi-marker metabarcoding approach to study mesozooplankton at basin scale. Sci. Rep. 2018, 8, 12085. [Google Scholar] [CrossRef] [Green Version]
- Carlton, J.T.; Ruiz, G.M. Vector science and integrated vector management in bioinvasion ecology: Conceptual framework. In Invasive Alien Species. A New Synthesis; Mooney, H.A., Mack, R.N., McNeely, J.A., Neville, L.E., Schei, P.J., Waage, J.K., Eds.; Island Press: Washington, DC, USA, 2005; pp. 36–58. [Google Scholar]
- Sündermann, J.; Pohlmann, T. A brief analysis of North Sea physics. Oceanologia 2011, 53, 663–689. [Google Scholar] [CrossRef] [Green Version]
- Jha, U.; Jetter, A.; Lindley, J.A.; Postel, L.; Wootton, M. Extension and distribution of Pseudodiaptomus marinus, an introduced copepod, in the North Sea. Mar. Biodivers. Rec. 2013, 6, e53. [Google Scholar] [CrossRef]
- Wootton, M.; Fischer, A.C.; Ostle, C.; Skinner, J.; Stevens, D.P.; Johns, D.G. Using the Continuous Plankton Recorder to study the distribution and ecology of marine pelagic copepods. In Trends in Copepod Studies—Distribution, Biology and Ecology; Uttieri, M., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2018; pp. 13–42. [Google Scholar]
- Deschutter, Y.; Vergara, G.; Mortelmans, J.; Deneudt, K.; De Schamphelaere, K.; De Troch, M. Distribution of the invasive calanoid copepod Pseudodiaptomus marinus (Sato, 1913) in the Belgian part of the North Sea. BioInvasions Rec. 2018, 7, 33–41. [Google Scholar] [CrossRef]
- Mutlu, E.; Özvarol, Y. Recent record of Oceania armata and near-past records of other gelatinous organisms in the Turkish waters presumably derived by basin-scale current. COMU J. Mar. Sci. Fish. 2022, 5, 48–55. [Google Scholar] [CrossRef]
- Civitarese, G.; Gačić, M.; Lipizer, M.; Eusebi Borzelli, G.L. On the impact of the Bimodal Oscillating System (BiOS) on the biogeochemistry and biology of the Adriatic and Ionian Seas (Eastern Mediterranean). Biogeosciences 2010, 7, 3987–3997. [Google Scholar] [CrossRef] [Green Version]
- Ozer, T.; Gertman, I.; Kress, N.; Silverman, J.; Herut, B. Interannual thermohaline (1979–2014) and nutrient (2002–2014) dynamics in the Levantine surface and intermediate water masses, SE Mediterranean Sea. Glob. Planet. Chang. 2017, 151, 60–67. [Google Scholar] [CrossRef]
- Fleminger, A.; Hendrix Kramer, S. Recent introduction of an Asian estuarine copepod, Pseudodiaptomus marinus (Copepoda: Calanoida), into southern California embayments. Mar. Biol. 1988, 98, 535–541. [Google Scholar] [CrossRef]
- Rajakaruna, H.; Lewis, M. Temperature cycles affect colonization potential of calanoid copepods. J. Theor. Biol. 2017, 419, 77–89. [Google Scholar] [CrossRef]
- Rajakaruna, H.; Strasser, C.; Lewis, M. Identifying non-invasible habitats for marine copepods using temperature-dependent R0. Biol. Invasions 2012, 14, 633–647. [Google Scholar] [CrossRef]
- Ojaveer, H.; Jaanus, A.; Mackenzie, B.R.; Martin, G.; Olenin, S.; Radziejewska, T.; Telesh, I.; Zettler, M.L.; Zaiko, A. Status of biodiversity in the Baltic Sea. PLoS ONE 2010, 5, e12467. [Google Scholar] [CrossRef] [Green Version]
- Leppäkoski, E.; Gollasch, S.; Gruszka, P.; Ojaveer, H.; Olenin, S.; Panov, V. The Baltic—A sea of invaders. Can. J. Fish. Aquat. Sci. 2002, 59, 1175–1188. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Temperature | Salinity |
|---|---|---|
| Bay of Biscay and the Iberian Coast | ||
| Peniche (PT) | 15.5–17.1 °C | 35.0–35.1 |
| Celtic Seas | ||
| Bristol Channel (UK) | n.a. | n.a. |
| Greater North Sea | ||
| East Anglia (UK) | n.a. | n.a. |
| East Kent (UK) | n.a. | n.a. |
| Isle of Wight (UK) | n.a. | n.a. |
| River Blackwater (UK) | n.a. | n.a. |
| River Humber (UK) | n.a. | n.a. |
| River Thames (UK) | n.a. | n.a. |
| Southampton (UK) | n.a. | n.a. |
| South Kent (UK) | n.a. | n.a. |
| Southeast Yorkshire (UK) | n.a. | n.a. |
| The Eastern Scheldt estuary (NL) [30] | ~21 °C | ~31 |
| Limfjord (DK) | 18 °C | 29 |
| Hirtshals (DK) | 12.2–12.8 °C | 31.0–32.5 |
| Western Mediterranean Sea | ||
| Leghorn (IT)—Nov 2020 §1 | 17.7 °C | 38.3 |
| —Nov 2021 §2 | 17.3 °C | 38.1 |
| Gulf of Naples—Sarno River (IT) §3 | 15.3 °C | 37.6 |
| Gulf of Naples—Torre del Greco (IT) §4 | 14.9 °C | 37.9 |
| Gulf of Naples—Between Torre del Greco and Torre Annunziata (IT) | 17.4–28.1 °C | 36.9–37.9 |
| Gulf of Pozzuoli (IT) | 15.2–27.0 °C | 37.6–38.0 |
| Gulf of Pozzuoli (IT) [32] | 17.4–27.3 °C | 37.5–37.7 |
| Gulf of Salerno (IT) §5 | 16.6 °C | 38.5 |
| Ionian Sea and the Central Mediterranean Sea | ||
| Marina di Ragusa (IT) §6 | 17.3 °C | 38.7 |
| Adriatic Sea | ||
| Emilia Romagna coasts (IT) [33] | 6.8–15.4 °C | 33.3–39.9 |
| Gulf of Trieste (IT) | 9.6–25.9 °C | 34.3–38.5 |
| Marano and Grado Lagoon (IT) | 5.0–30.0 °C | 4.7–35.4 |
| Croatian ports (HR) [35,36] | n.a. | n.a. |
| Neretva River (HR) [37] | 10.6–25.5 °C | 0.0–38.4 |
| Aegean-Levantine Sea | ||
| Thessaloniki Bay (GR) [40] | 17.2–31.0 °C | 34.8–38.5 |
| Thermaikos Gulf (GR) | 14.7–27.7 °C | 34.9–38.5 |
| İskenderun Bay (TR) [49,50] | 15.2–16.1 °C | 38.5–38.8 |
| İzmir Bay (TR) # [44] | 19 °C | 38.9 |
| Yenifoça Bay (TR) [46,47] | 14.6–15.1 °C | 39.2 |
| Hadera monitoring station (IL) [48] | 16.2–32.4 °C | 38.3–40.4 |
| Black Sea | ||
| İzmit Bay (TR) [54] | 15.6–23.2 °C | 29.5–38.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uttieri, M.; Anadoli, O.; Banchi, E.; Battuello, M.; Beşiktepe, Ş.; Carotenuto, Y.; Marques, S.C.; de Olazabal, A.; Di Capua, I.; Engell-Sørensen, K.; et al. The Distribution of Pseudodiaptomus marinus in European and Neighbouring Waters—A Rolling Review. J. Mar. Sci. Eng. 2023, 11, 1238. https://doi.org/10.3390/jmse11061238
Uttieri M, Anadoli O, Banchi E, Battuello M, Beşiktepe Ş, Carotenuto Y, Marques SC, de Olazabal A, Di Capua I, Engell-Sørensen K, et al. The Distribution of Pseudodiaptomus marinus in European and Neighbouring Waters—A Rolling Review. Journal of Marine Science and Engineering. 2023; 11(6):1238. https://doi.org/10.3390/jmse11061238
Chicago/Turabian StyleUttieri, Marco, Olga Anadoli, Elisa Banchi, Marco Battuello, Şengül Beşiktepe, Ylenia Carotenuto, Sónia Cotrim Marques, Alessandra de Olazabal, Iole Di Capua, Kirsten Engell-Sørensen, and et al. 2023. "The Distribution of Pseudodiaptomus marinus in European and Neighbouring Waters—A Rolling Review" Journal of Marine Science and Engineering 11, no. 6: 1238. https://doi.org/10.3390/jmse11061238