A Novel Bioengineered Functional Motor Unit Platform to Study Neuromuscular Interaction

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Experimental Section
2.1. Immortalised Human Skeletal Muscle Cell Culture
2.2. Isolation of Rat Embryonic Spinal Cord Explants
2.3. Co-Culture
2.4. Immunocytochemistry
2.5. Assessment of Functional NMJ Formation
2.6. Statistical Analyses
3. Results
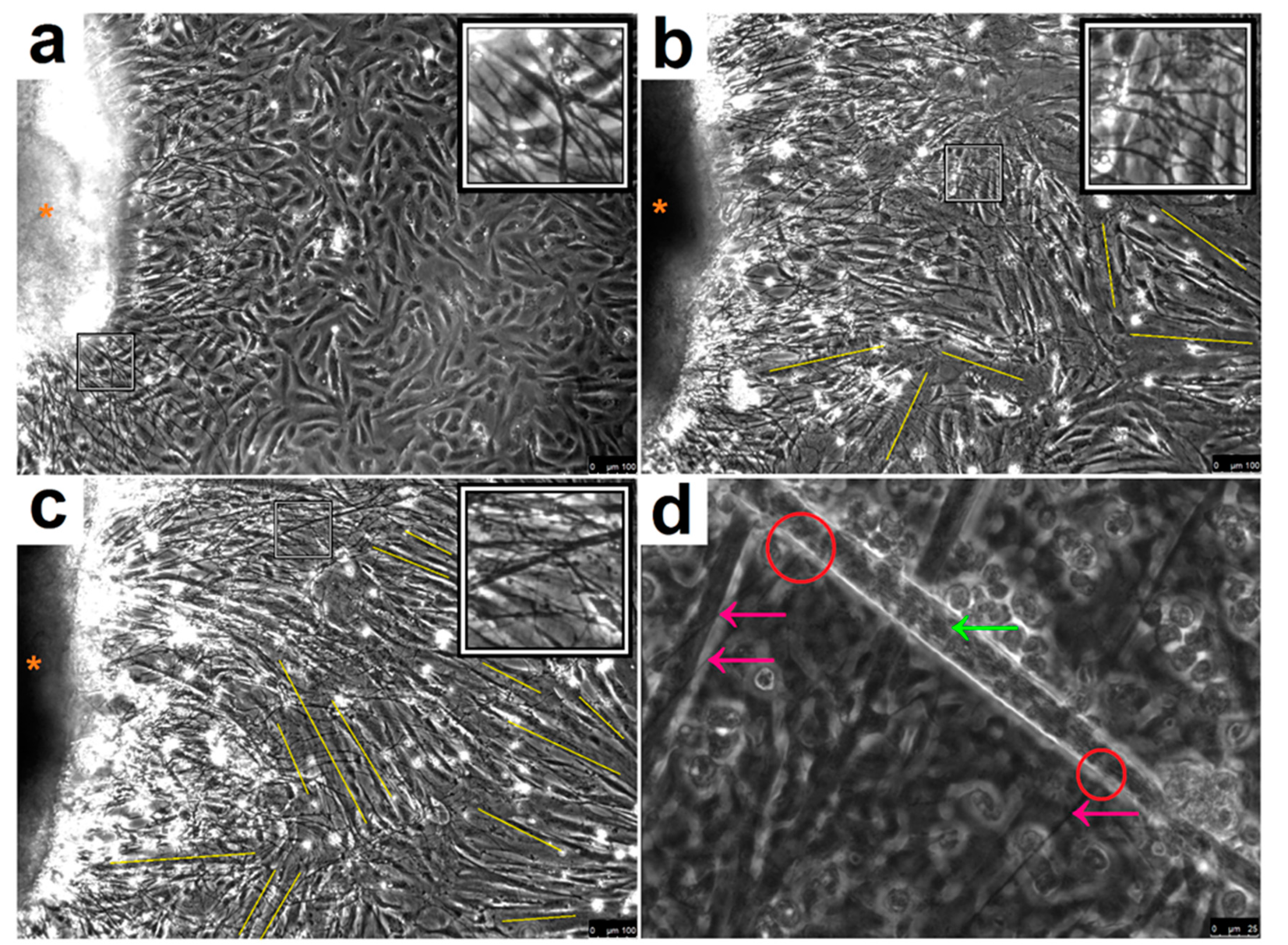
3.1. Co-Culture Morphological Characterisation
3.2. Spontaneous Myotube Contractions
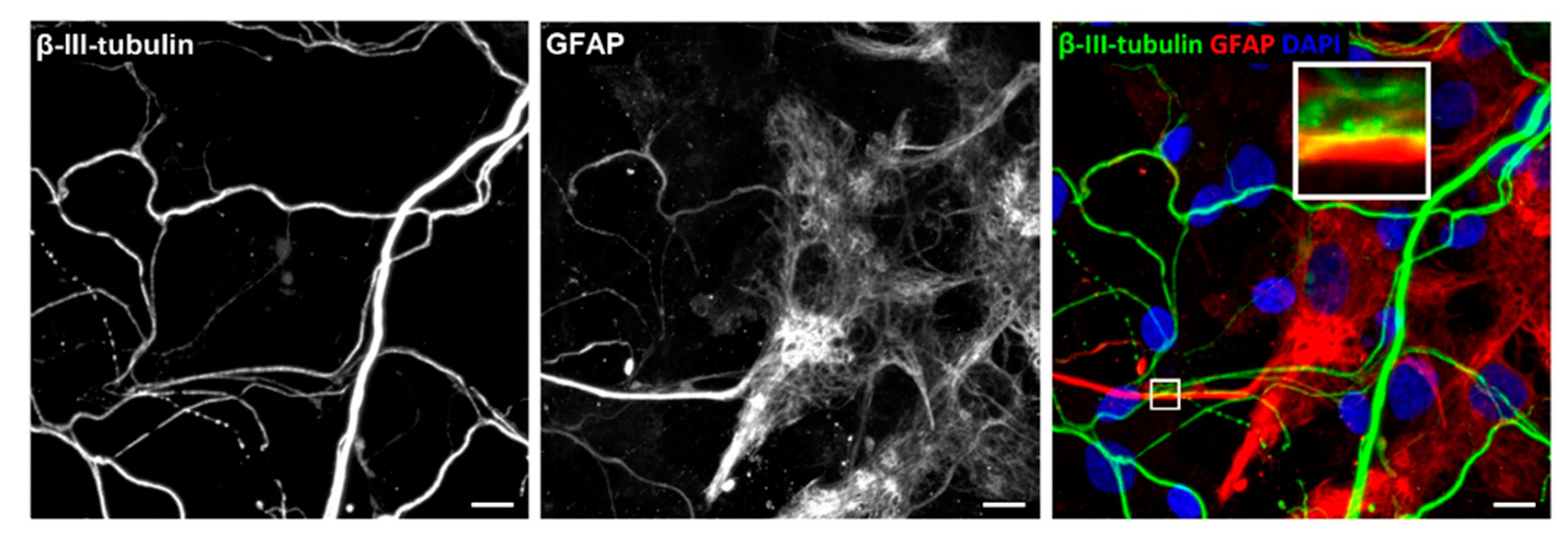
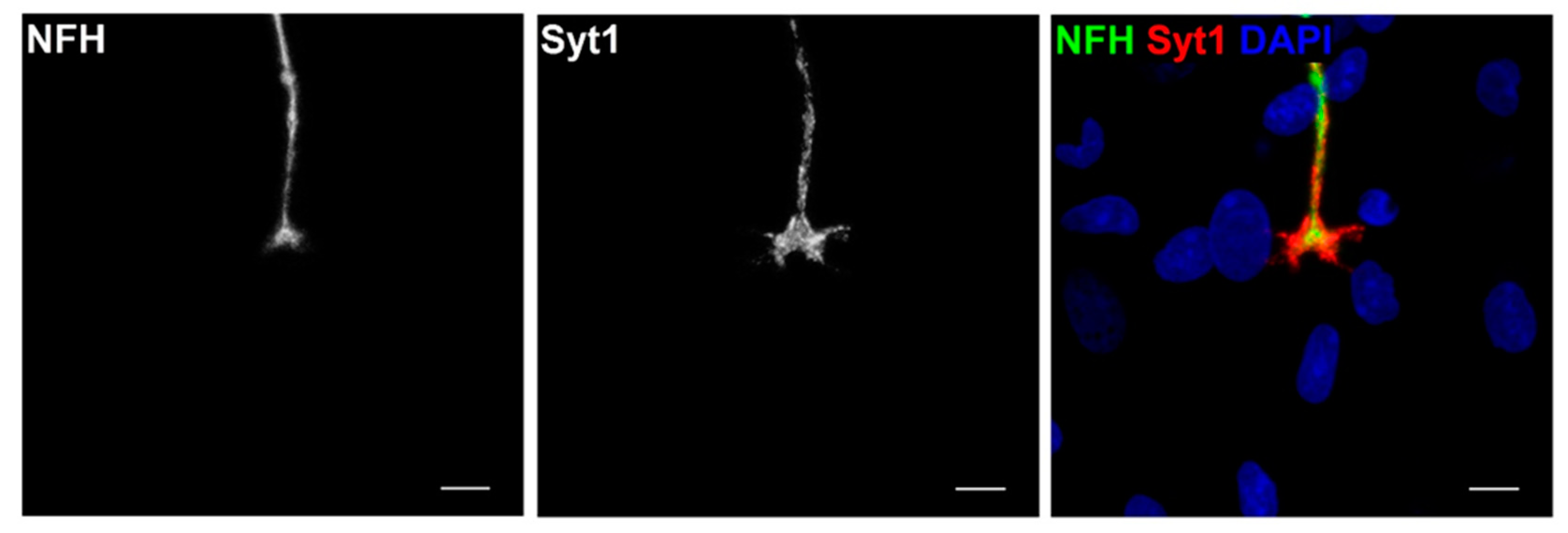
3.3. Characterisation of Neuronal Cells
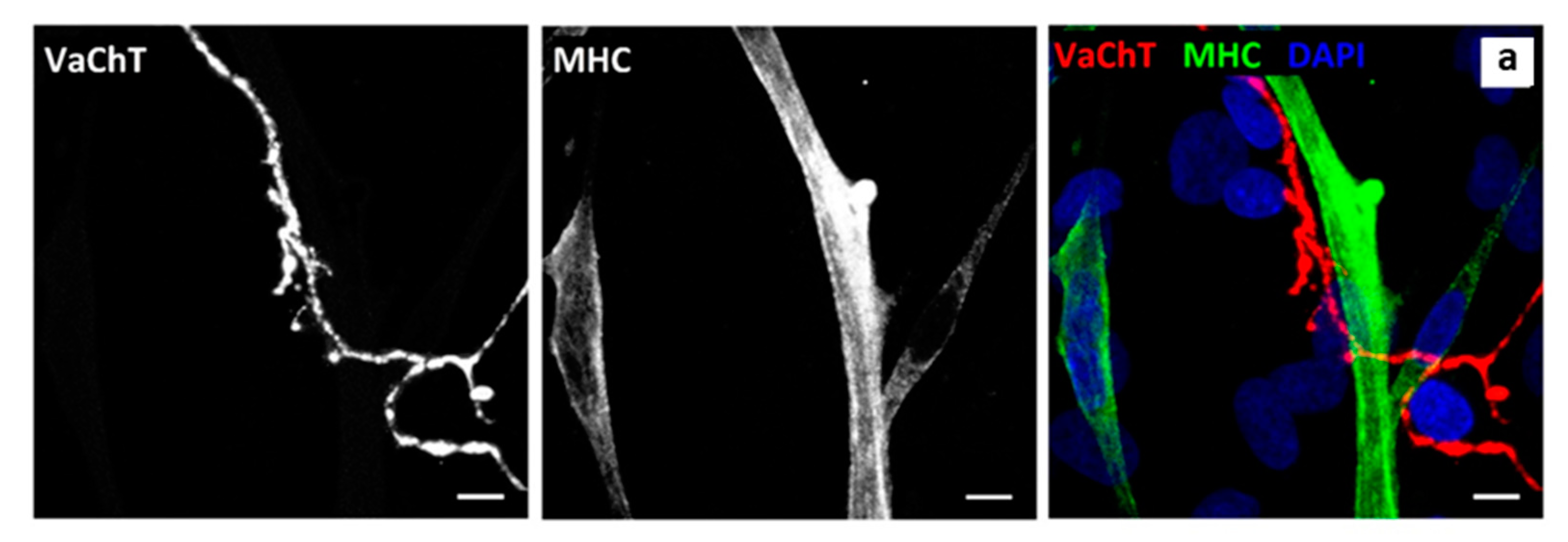
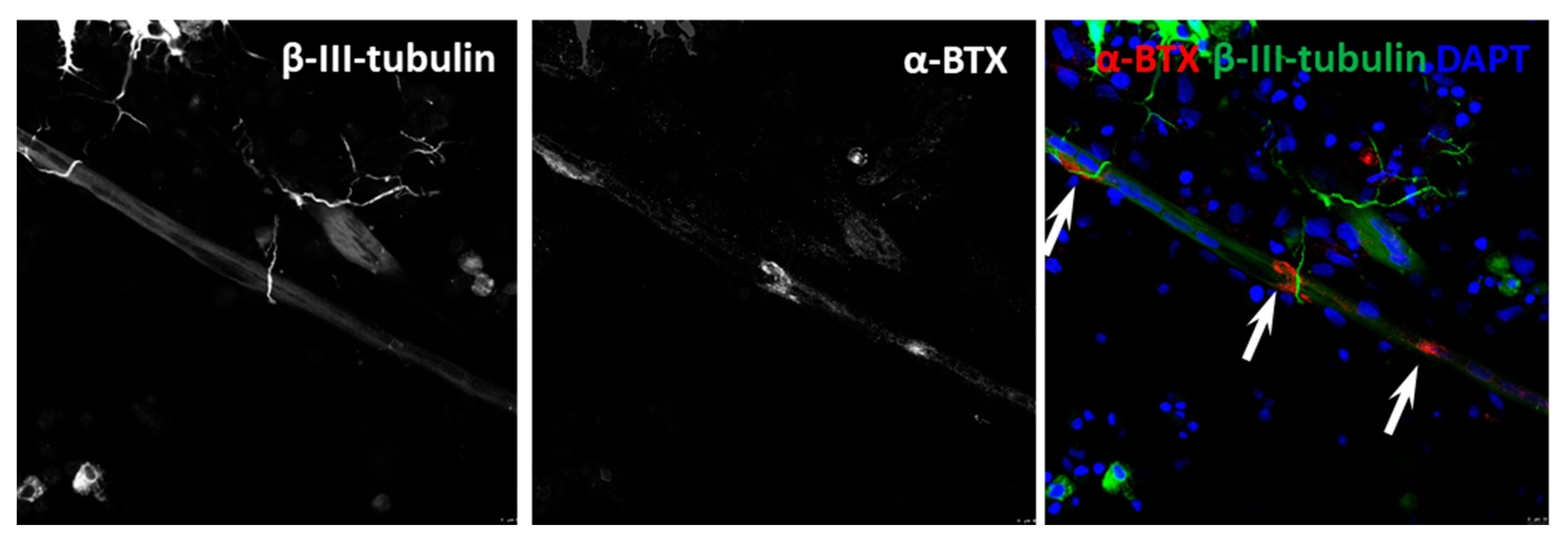
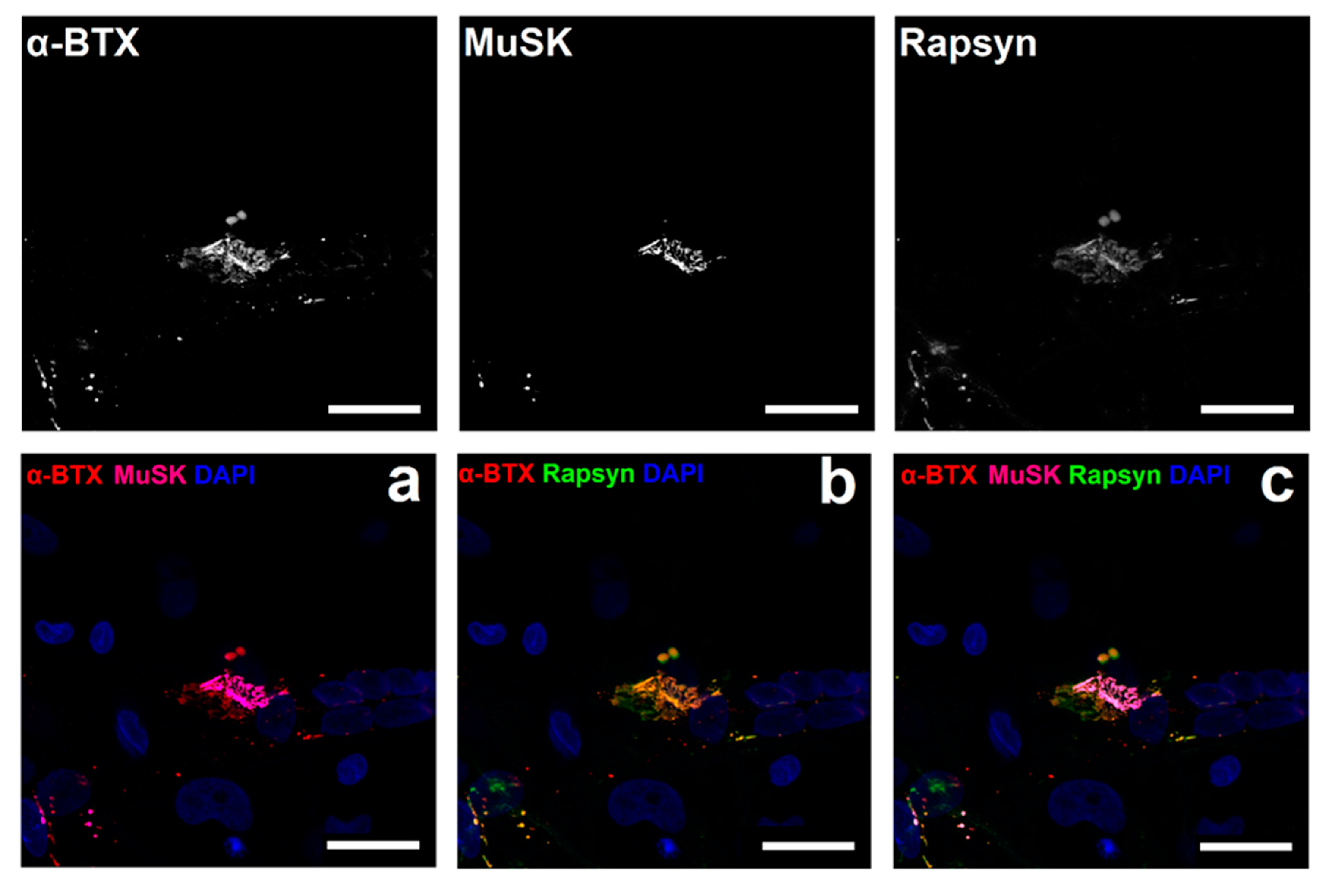
3.4. NMJ Formation
3.5. Functional Assessment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ferraro, E.; Molinari, F.; Berghella, L. Molecular control of neuromuscular junction development. J. Cachexia Sarcopenia Muscle 2012, 3, 13–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, I.Y.; Ehrlich, B.E. Signaling in muscle contraction. Cold Spring Harbor Perspect. Biol. 2015, 7, a006023. [Google Scholar] [CrossRef] [PubMed]
- Campanari, M.-L.; García-Ayllón, M.-S.; Ciura, S.; Sáez-Valero, J.; Kabashi, E. Neuromuscular junction impairment in amyotrophic lateral sclerosis: Reassessing the role of acetylcholinesterase. Front. Mol. Neurosci. 2016, 9, 160–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, A.; Palace, J.; Hilton-Jones, D. Myasthenia gravis. The numbers of limb motor neurons in the human lumbosacral cord throughout life. Lancet 2001, 357, 2122–2128. [Google Scholar] [CrossRef]
- Tomlinson, B.E.; Irving, D. The numbers of limb motor neurons in the human lumbosacral cord throughout life. J. Neurol. Sci. 1977, 34, 213–219. [Google Scholar] [CrossRef]
- van der Worp, H.B.; Howells, D.W.; Sena, E.S.; Porritt, M.J.; Rewell, S.; O’Collins, V.; Macleod, M.R. Can animal models of disease reliably inform human studies? PLoS Med. 2010, 7, e1000245. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Browne, K.D.; Laimo, F.A.; Maggiore, J.C.; Hilman, M.C.; Kaisaier, H.; Aguilar, C.A.; Ali, Z.S.; Mourkioti, F.; Cullen, D.K. Pre-innervated tissue-engineered muscle promotes a pro-regenerative microenvironment following volumetric muscle loss. Commun. Biol. 2020, 3, 330. [Google Scholar] [CrossRef]
- Cantor, S.; Zhang, W.; Delestrée, N.; Remédio, L.; Mentis, G.Z.; Burden, S.J. Preserving neuromuscular synapses in ALS by stimulating MuSK with a therapeutic agonist antibody. eLife 2018, 20, e34375. [Google Scholar] [CrossRef] [Green Version]
- Vila, O.F.; Qu, Y.; Vunjak-Novakovic, G. In vitro models of neuromuscular junctions and their potential for novel drug discovery and development. Exp. Opin. Drug Discov. 2020, 15, 307–317. [Google Scholar] [CrossRef]
- Haase, G. Motor neuron diseases: Cellular and animal models. In Reviews in Cell Biology and Molecular Medicine; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2006. [Google Scholar]
- Prather, R.S.; Lorson, M.; Ross, J.W.; Whyte, J.J.; Walters, E. Genetically engineered pig models for human diseases. Ann. Rev. Anim. Biosci. 2013, 1, 203–219. [Google Scholar] [CrossRef] [Green Version]
- Suuronen, E.J.; McLaughlin, C.R.; Stys, P.K.; Nakamura, M.; Munger, R.; Griffith, M. Functional innervation in tissue engineered models for in vitro study and testing purposes. Toxicol. Sci. 2004, 82, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Ashby, P.R.; Wilson, S.J.; Harris, A.J. Formation of primary and secondary myotubes in aneural muscles in the mouse mutant peroneal muscular atrophy. Dev. Biol. 1993, 156, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Umbach, J.A.; Adams, K.L.; Gundersen, C.B.; Novitch, B.G. Functional neuromuscular junctions formed by embryonic stem cell-derived motor neurons. PLoS ONE 2012, 7, e36049. [Google Scholar] [CrossRef] [PubMed]
- Demestre, M.; Orth, M.; Föhr, K.J.; Achberger, K.; Ludolph, A.C.; Liebau, S.; Boeckers, T.M. Formation and characterisation of neuromuscular junctions between hipsc derived motoneurons and myotubes. Stem Cell Res. 2015, 15, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Harper, J.M.; Krishnan, C.; Darman, J.S.; Deshpande, D.M.; Peck, S.; Shats, I.; Backovic, S.; Rothstein, J.D.; Kerr, D.A. Axonal growth of embryonic stem cell-derived motoneurons in vitro and in motoneuron-injured adult rats. Proc. Natl. Acad. Sci. USA 2004, 101, 7123–7128. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Greene, K.; Akanda, N.; Smith, A.; Stancescu, M.; Lambert, S.; Vandenburgh, H.; Hickman, J. In vitro differentiation of functional human skeletal myotubes in a defined system. Biomater. Sci. 2014, 2, 131–138. [Google Scholar] [CrossRef]
- Arnold, A.-S.; Christe, M.; Handschin, C. A functional motor unit in the culture dish: Co-culture of spinal cord explants and muscle cells. J. Vis. Exp. JoVE 2012, 12, 3616. [Google Scholar] [CrossRef] [Green Version]
- Behringer, R.; Gertsenstein, M.; Nagy, K.V.; Nagy, A. Testing serum batches for mouse embryonic stem cell culture. Cold Spring Harbor Protoc. 2017, 2017, 092411. [Google Scholar] [CrossRef]
- Rumsey, J.W.; Das, M.; Stancescu, M.; Bott, M.; Fernandez-Valle, C.; Hickman, J.J. Node of ranvier formation on motoneurons in vitro. Biomaterials 2009, 30, 3567–3572. [Google Scholar] [CrossRef] [Green Version]
- Mouly, V.; Aamiri, A.; Perie, S.; Mamchaoui, K.; Barani, A.; Bigot, A.; Bouazza, B.; Francois, V.; Furling, D.; Jacquemin, V.; et al. Myoblast transfer therapy: Is there any light at the end of the tunnel? Acta Myol. 2005, 24, 128–133. [Google Scholar]
- Webster, C.; Blau, H.M. Accelerated age-related decline in replicative life-span of duchenne muscular dystrophy myoblasts: Implications for cell and gene therapy. Som. Cell Mol. Genet. 1990, 16, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; Woltjen, K.; Miyake, K.; Hotta, A.; Ikeya, M.; Yamamoto, T.; Nishino, T.; Shoji, E.; Sehara-Fujisawa, A.; Manabe, Y.; et al. Efficient and reproducible myogenic differentiation from human ips cells: Prospects for modeling miyoshi myopathy in vitro. PLoS ONE 2013, 8, e61540. [Google Scholar] [CrossRef]
- Stockmann, M.; Linta, L.; Fohr, K.J.; Boeckers, A.; Ludolph, A.C.; Kuh, G.F.; Udvardi, P.T.; Proepper, C.; Storch, A.; Kleger, A.; et al. Developmental and functional nature of human ipsc derived motoneurons. Stem Cell Rev. 2013, 9, 475–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.J.; Du, Z.W.; Zarnowska, E.D.; Pankratz, M.; Hansen, L.O.; Pearce, R.A.; Zhang, S.C. Specification of motoneurons from human embryonic stem cells. Nat. Biotechnol. 2005, 23, 215–221. [Google Scholar] [CrossRef]
- Whiteside, T.L. The tumor microenvironment and its role in promoting tumor growth. Oncogene 2008, 27, 5904–5912. [Google Scholar] [CrossRef] [Green Version]
- Saini, J.; Faroni, A.; Abd Al Samid, M.; Reid, A.J.; Lightfoot, A.P.; Mamchaoui, K.; Mouly, V.; Butler-Browne, G.; McPhee, J.S.; Degens, H.; et al. Simplified in vitro engineering of neuromuscular junctions between rat embryonic motoneurons and immortalized human skeletal muscle cells. Stem Cells Cloning 2019, 12, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mamchaoui, K.; Trollet, C.; Bigot, A.; Negroni, E.; Chaouch, S.; Wolff, A.; Kandalla, P.K.; Marie, S.; Di Santo, J.; St Guily, J.L.; et al. Immortalized pathological human myoblasts: Towards a universal tool for the study of neuromuscular disorders. Skelet. Muscle 2011, 1, 34. [Google Scholar] [CrossRef] [Green Version]
- Das, M.; Rumsey, J.W.; Gregory, C.A.; Bhargava, N.; Kang, J.F.; Molnar, P.; Riedel, L.; Guo, X.; Hickman, J.J. Embryonic motoneuron-skeletal muscle co-culture in a defined system. Neuroscience 2007, 146, 481–488. [Google Scholar] [CrossRef]
- Guo, X.; Gonzalez, M.; Stancescu, M.; Vandenburgh, H.H.; Hickman, J.J. Neuromuscular junction formation between human stem cell-derived motoneurons and human skeletal muscle in a defined system. Biomaterials 2011, 32, 9602–9611. [Google Scholar] [CrossRef] [Green Version]
- Morimoto, Y.; Kato-Negishi, M.; Onoe, H.; Takeuchi, S. Three-dimensional neuron-muscle constructs with neuromuscular junctions. Biomaterials 2013, 34, 9413–9419. [Google Scholar] [CrossRef]
- Sanes, J.R.; Lichtman, J.W. Development of the vertebrate neuromuscular junction. Ann. Rev. Neurosci. 1999, 22, 389–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oda, Y. Choline acetyltransferase: The structure, distribution and pathologic changes in the central nervous system. Pathol. Int. 1999, 49, 921–937. [Google Scholar] [CrossRef] [PubMed]
- Arvidsson, U.; Riedl, M.; Elde, R.; Meister, B. Vesicular acetylcholine transporter (vacht) protein: A novel and unique marker for cholinergic neurons in the central and peripheral nervous systems. J. Comp. Neurol. 1997, 378, 454–467. [Google Scholar] [CrossRef]
- Low, L.K.; Cheng, H.-J. Axon pruning: An essential step underlying the developmental plasticity of neuronal connections. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1531–1544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balice-Gordon, R.J. Schwann cells: Dynamic roles at the neuromuscular junction. Curr. Biol. 1996, 6, 1054–1056. [Google Scholar] [CrossRef] [Green Version]
- Jessen, K.R.; Morgan, L.; Stewart, H.J.; Mirsky, R. Three markers of adult non-myelin-forming schwann cells, 217c(ran-1), a5e3 and gfap: Development and regulation by neuron-schwann cell interactions. Development 1990, 109, 91–103. [Google Scholar]
- Katsetos, C.D.; Legido, A.; Perentes, E.; Mork, S.J. Class iii beta-tubulin isotype: A key cytoskeletal protein at the crossroads of developmental neurobiology and tumor neuropathology. J. Child Neurol. 2003, 18, 851–866. [Google Scholar] [CrossRef]
- Lees, J.F.; Shneidman, P.S.; Skuntz, S.F.; Carden, M.J.; Lazzarini, R.A. The structure and organization of the human heavy neurofilament subunit (nf-h) and the gene encoding it. EMBO J. 1988, 7, 1947–1955. [Google Scholar] [CrossRef]
- Young, H.S.; Herbette, L.G.; Skita, V. Alpha-bungarotoxin binding to acetylcholine receptor membranes studied by low angle X-ray diffraction. Biophys. J. 2003, 85, 943–953. [Google Scholar] [CrossRef] [Green Version]
- Brose, N.; Petrenko, A.G.; Sudhof, T.C.; Jahn, R. Synaptotagmin: A calcium sensor on the synaptic vesicle surface. Science 1992, 256, 1021–1025. [Google Scholar] [CrossRef]
- Apel, E.D.; Glass, D.J.; Moscoso, L.M.; Yancopoulos, G.D.; Sanes, J.R. Rapsyn is required for musk signaling and recruits synaptic components to a musk-containing scaffold. Neuron 1997, 18, 623–635. [Google Scholar] [CrossRef] [Green Version]
- Lavoie, P.A.; Collier, B.; Tenehouse, A. Comparison of alpha-bungarotoxin binding to skeletal muscles after inactivity or denervation. Nature 1976, 260, 349–350. [Google Scholar] [CrossRef]
- Cooke, I.M.; Grinnell, A.D. Effect of tubocurarine on action potentials in normal and denervated skeletal muscle. J. Physiol. 1964, 175, 203–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rekling, J.C.; Funk, G.D.; Bayliss, D.A.; Dong, X.W.; Feldman, J.L. Synaptic control of motoneuronal excitability. Physiol. Rev. 2000, 80, 767–852. [Google Scholar] [CrossRef] [PubMed]
- Burgess, C.; Lai, D.; Siegel, J.; Peever, J. An endogenous glutamatergic drive onto somatic motoneurons contributes to the stereotypical pattern of muscle tone across the sleep-wake cycle. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 4649–4660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feher, J. The neuromuscular junction and excitation–contraction coupling. In Quantitative Human Physiology, 2nd ed.; Academic Press: Boston, MA, USA, 2017; pp. 318–333. [Google Scholar]
- Bandi, E.; Jevsek, M.; Mars, T.; Jurdana, M.; Formaggio, E.; Sciancalepore, M.; Fumagalli, G.; Grubic, Z.; Ruzzier, F.; Lorenzon, P. Neural agrin controls maturation of the excitation-contraction coupling mechanism in human myotubes developing in vitro. Am. J. Physiol. Cell Physiol. 2008, 294, C66–C73. [Google Scholar] [CrossRef] [Green Version]
- Arnold, A.S.; Gueye, M.; Guettier-Sigrist, S.; Courdier-Fruh, I.; Coupin, G.; Poindron, P.; Gies, J.P. Reduced expression of nicotinic achrs in myotubes from spinal muscular atrophy i patients. Laboratory investigation. J. Tech. Methods Pathol. 2004, 84, 1271–1278. [Google Scholar]
- Askanas, V.; Kwan, H.; Alvarez, R.B.; Engel, W.K.; Kobayashi, T.; Martinuzzi, A.; Hawkins, E.F. De novo neuromuscular junction formation on human muscle fibres cultured in monolayer and innervated by foetal rat spinal cord: Ultrastructural and ultrastructural—Cytochemical studies. J. Neurocytol. 1987, 16, 523–537. [Google Scholar] [CrossRef]
- Hong, I.H.; Etherington, S.J. Neuromuscular Junction (els); John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2011. [Google Scholar]
- Das, M.; Rumsey, J.W.; Bhargava, N.; Stancescu, M.; Hickman, J.J. A defined long-term in vitro tissue engineered model of neuromuscular junctions. Biomaterials 2010, 31, 4880–4888. [Google Scholar] [CrossRef] [Green Version]
- Rumsey, J.W.; Das, M.; Bhalkikar, A.; Stancescu, M.; Hickman, J.J. Tissue engineering the mechanosensory circuit of the stretch reflex arc: Sensory neuron innervation of intrafusal muscle fibers. Biomaterials 2010, 31, 8218–8227. [Google Scholar] [CrossRef] [Green Version]
- Mears, S.C.; Frank, E. Formation of specific monosynaptic connections between muscle spindle afferents and motoneurons in the mouse. J. Neurosci. Off. J. Soc. Neurosci. 1997, 17, 3128–3135. [Google Scholar] [CrossRef] [Green Version]
- Riethmacher, D.; Sonnenberg-Riethmacher, E.; Brinkmann, V.; Yamaai, T.; Lewin, G.R.; Birchmeier, C. Severe neuropathies in mice with targeted mutations in the erbb3 receptor. Nature 1997, 389, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Jevsek, M.; Mars, T.; Mis, K.; Grubic, Z. Origin of acetylcholinesterase in the neuromuscular junction formed in the in vitro innervated human muscle. Eur. J. Neurosci. 2004, 20, 2865–2871. [Google Scholar] [CrossRef] [PubMed]
- Domet, M.A.; Webb, C.E.; Wilson, D.F. Impact of α-bungarotoxin on transmitter release at the neuromuscular junction of the rat. Neurosci. Lett. 1995, 199, 49–52. [Google Scholar] [CrossRef]
- Bowman, W.C. Neuromuscular block. Br. J. Pharmacol. 2006, 147, S277–S286. [Google Scholar] [CrossRef]
- Nurullin, L.F.; Nikolsky, E.E.; Malomouzh, A.I. Elements of molecular machinery of GABAergic signaling in the vertebrate cholinergic neuromuscular junction. Acta Histochem. 2018, 120, 298–301. [Google Scholar] [CrossRef]
- Guo, X.; Das, M.; Rumsey, J.; Gonzalez, M.; Stancescu, M.; Hickman, J. Neuromuscular junction formation between human stem-cell-derived motoneurons and rat skeletal muscle in a defined system. Tissue Eng. Part C Methods 2010, 16, 1347–1355. [Google Scholar] [CrossRef] [Green Version]
- Bakooshli, A.M.; Lippmann, E.S.; Mulcahy, B.; Iyer, N.; Nguyen, C.T.; Tung, K.; Stewart, B.A.; van den Dorpel, H.; Fuehrmann, T.; Shoichet, M.; et al. A 3D culture model of innervated human skeletal muscle enables studies of the adult neuromuscular junction. eLife 2019, 8, e44530. [Google Scholar] [CrossRef]
- Abd Al Samid, M.; McPhee, J.S.; Saini, J.; McKay, T.R.; Fitzpatrick, L.M.; Mamchaoui, K.; Bigot, A.; Mouly, V.; Butler-Browne, G.; Al-Shanti, N. A functional human motor unit platform engineered from human embryonic stem cells and immortalized skeletal myoblasts. Stem Cells Cloning 2018, 9, 85–93. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Media Components/Company | Concentration | Catalogue # |
|---|---|---|
| Dulbecco’s Modified Eagle Media (DMEM) from Lonza | 59% (v/v) | 12–914F |
| Medium 199 with Earle’s Balanced Salt Solution from Lonza | 20% (v/v) | 12-119F |
| Heat-inactivated fetal bovine serum (FBS) from Thermo Fisher Scientific | 20% (v/v) | 10500-064 |
| L-glutamine from Lonza | 1% (v/v) | 17-605E |
| Fetuin from fetal bovine serum from Sigma-Aldrich (St Louis, MO, USA) | 25 μg/mL | F3004 |
| Recombinant human basic fibroblast growth factor (FGFb) from Thermo Fisher Scientific | 0.5 ng/mL | PHG0311 |
| Recombinant human epidermal growth factor (EGF) from Thermo Fisher Scientific | 5 ng/mL | PHG0311 |
| Recombinant human hepatocyte growth factor (HGF) from Sino Biological Inc. (Beijing, China) | 2.5 ng/mL | 10463-HNAS |
| Recombinant human insulin from Sigma-Aldrich | 5 μg/mL | 91077C |
| Dexamethasone from Sigma-Aldrich | 0.2 μg/mL | 10103483 |
| Gentamicin from Thermo Fisher Scientific | 10 μg/mL | 15710-049 |
| Antibody/Company | Concentration | Catalogue # |
|---|---|---|
| Anti-Vesicular Acetylcholine Transporter (VAChT) from Merck Millipore (Burlington, MA, USA) | 1:100 | ABN100 |
| Anti-Choline Acetyltransferase (ChAT) from Merck Millipore | 1:100 | AB144 |
| Anti-Glial Fibrillary Acidic Protein (GFAP) from Sigma-Aldrich | 1:100 | G3893 |
| Anti-Neurofilament Heavy (NFH) from Merck Millipore | 1:100 | AB5539 |
| Anti-Synaptotagmin (Syt1) from Abcam (Cambridge, UK) | 1:100 | ab13259 |
| Anti-Ryanodine Receptor 1 (RyR) from Merck Millipore | 1:100 | AB9078 |
| Anti-Calcium channel L type DHPR alpha 2 subunit (DHPR) from Abcam | 1:100 | ab2864 |
| Anti-Receptor-Associated Protein of the Synapse (Rapsyn) from Abcam | 1:100 | ab11423 |
| Anti-Muscle-Specific Kinase (MuSK) from Abcam | 1:100 | ab92950 |
| Anti-Beta Tubulin Class III Alexa Fluor® 488 conjugate from Thermo Fisher Scientific | 1:400 | 53-4510-82 |
| α-Bungarotoxin, Alexa Fluor® 647 conjugate from Thermo Fisher Scientific | 1:400 | B35450 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saini, J.; Faroni, A.; Reid, A.J.; Mamchaoui, K.; Mouly, V.; Butler-Browne, G.; Lightfoot, A.P.; McPhee, J.S.; Degens, H.; Al-Shanti, N. A Novel Bioengineered Functional Motor Unit Platform to Study Neuromuscular Interaction. J. Clin. Med. 2020, 9, 3238. https://doi.org/10.3390/jcm9103238
Saini J, Faroni A, Reid AJ, Mamchaoui K, Mouly V, Butler-Browne G, Lightfoot AP, McPhee JS, Degens H, Al-Shanti N. A Novel Bioengineered Functional Motor Unit Platform to Study Neuromuscular Interaction. Journal of Clinical Medicine. 2020; 9(10):3238. https://doi.org/10.3390/jcm9103238
Chicago/Turabian StyleSaini, Jasdeep, Alessandro Faroni, Adam J. Reid, Kamel Mamchaoui, Vincent Mouly, Gillian Butler-Browne, Adam P. Lightfoot, Jamie S. McPhee, Hans Degens, and Nasser Al-Shanti. 2020. "A Novel Bioengineered Functional Motor Unit Platform to Study Neuromuscular Interaction" Journal of Clinical Medicine 9, no. 10: 3238. https://doi.org/10.3390/jcm9103238