Antibacterial Activity of Some Flavonoids and Organic Acids Widely Distributed in Plants



1
Department of Botany, Breeding and Agricultural Technology of Medicinal Plants, Institute of Natural Fibres and Medicinal Plants, Kolejowa 2, 62-064 Plewiska, Poland
2
Department of Biotechnology, Institute of Natural Fibres and Medicinal Plants, Wojska Polskiego 71b, 60-630 Poznań, Poland
3
Department of Medical Microbiology, Poznań University of Medical Sciences, Wieniawskiego 3, 61-712 Poznań, Poland
*
Author to whom correspondence should be addressed.
J. Clin. Med. 2020, 9(1), 109; https://doi.org/10.3390/jcm9010109
Submission received: 21 November 2019
/
Revised: 24 December 2019
/
Accepted: 27 December 2019
/
Published: 31 December 2019
(This article belongs to the Special Issue Bioactive Phytochemicals in Health and Disease)
Abstract
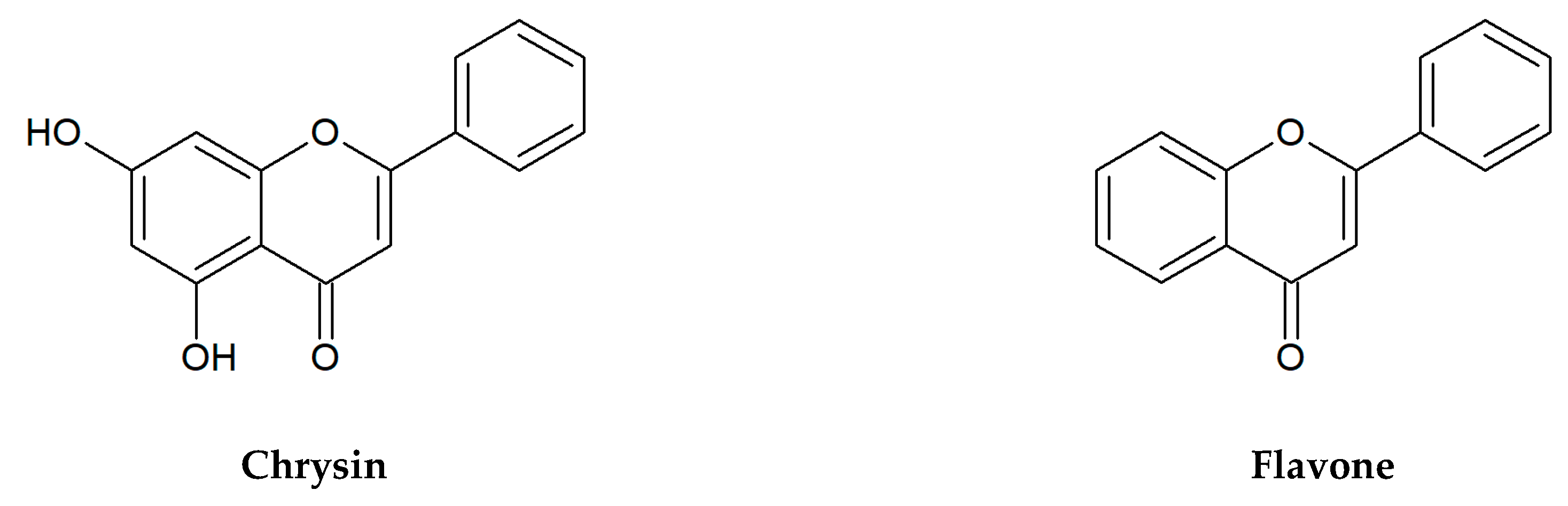
:Among natural substances widespread in fruits, vegetables, spices, and medicinal plants, flavonoids and organic acids belong to the promising groups of bioactive compounds with strong antioxidant and anti-inflammatory properties. The aim of the present work was to evaluate the antibacterial activity of 13 common flavonoids (flavones, flavonols, flavanones) and 6 organic acids (aliphatic and aromatic acids). The minimal inhibitory concentrations (MICs) of selected plant substances were determined by the micro-dilution method using clinical strains of four species of pathogenic bacteria. All tested compounds showed antimicrobial properties, but their biological activity was moderate or relatively low. Bacterial growth was most strongly inhibited by salicylic acid (MIC = 250–500 μg/mL). These compounds were generally more active against Gram-negative bacteria: Escherichia coli and Pseudomonas aeruginosa than Gram-positive ones: Enterococcus faecalis and Staphylococcus aureus. An analysis of the antibacterial effect of flavone, chrysin, apigenin, and luteolin showed that the presence of hydroxyl groups in the phenyl rings A and B usually did not influence on the level of their activity. A significant increase in the activity of the hydroxy derivatives of flavone was observed only for S. aureus. Similarly, the presence and position of the sugar group in the flavone glycosides generally had no effect on the MIC values.
Keywords:
kaempferol; naringin; orientin; rutin; vitexin; chlorogenic acid; citric acid; malic acid; quinic acid; rosmarinic acid1. Introduction
Screening biological studies of chemical compounds of natural origin allow for assessment of their activity and determine further research stages in order to search for new therapeutic solutions based on active compounds known in plants. This is especially important during the observed increasing resistance of bacteria and fungi to antibiotics. Multidrug resistance (MDR) is a serious threat to human health, but also to crops and animals. MDR is a growing challenge in medicine. Recently, several multinational studies have been carried out to determine the prevalence of herbal medicine use in infections due to pathogenic microorganisms [1,2]. It is considered that extracts of medicinal plants can be an alternative source of resistance modifying substances [2]. It is well known that plant extracts and other herbal products are complex mixtures containing the wide variety of primary and secondary metabolites, and their action may be the result of the synergy of different chemical components. Moreover, these extracts may show various mechanisms of biological and pharmacological activity, i.e., ability to bind to protein domains, modulation of the immune response, mitosis, apoptosis, and signal transduction [2]. However, it should be noted that plants interact with the environment and other organisms, therefore their chemical composition and the level of active substances can be very diverse [3,4]. In addition, the manufacturing process of herbal medicinal products is very complex because it encompasses non-standardized processes like the cultivation of plants, obtaining the vegetable raw material from various parts of the world, preparing of extract, and producing a product in accordance with local guidelines of the good manufacturing practice. Therefore, it can be concluded that using pure chemical compounds of natural origin would be an interesting complementary option due to their easier therapeutic dosage, the study of mechanisms of the pharmacological action and monitoring of their side effects.
A lot of widespread plant substances, including alkaloids, organosulfur compounds, phenolic acids, flavonoids, carotenoids, coumarins, terpenes, tannins, and some primary metabolites (amino acids, peptides, organic acids) exhibit antimicrobial properties [1,5,6,7,8]. Among them, flavonoids are a promising group of bioactive substances with low systemic toxicity. Natural flavonols, flavones, flavanones, and other compounds of this class belong to the common secondary metabolites found in various fruits, vegetables, and medicinal plants [9] showing strong antioxidant and anti-inflammatory properties [10,11]. Dietary polyphenols such as flavonoids and phenolic acids, consumed in large quantities in foods of plant origin, exhibit a number of beneficial effects and play an important role in the prevention of chronic and degenerative diseases. Not only their antioxidant and anti-inflammatory activities, but also neuroprotective, anticancer, immunomodulatory, antidiabetic, and anti-adipogenic properties have been shown [12,13]. Biological availability of dietary polyphenols is low as compared with micro- and macronutrients. Their absorption in the small intestine amounts only about 5–10%. However, recent studies showed that these phytochemicals exhibit prebiotic properties and antimicrobial activity against pathogenic intestinal microflora [13].
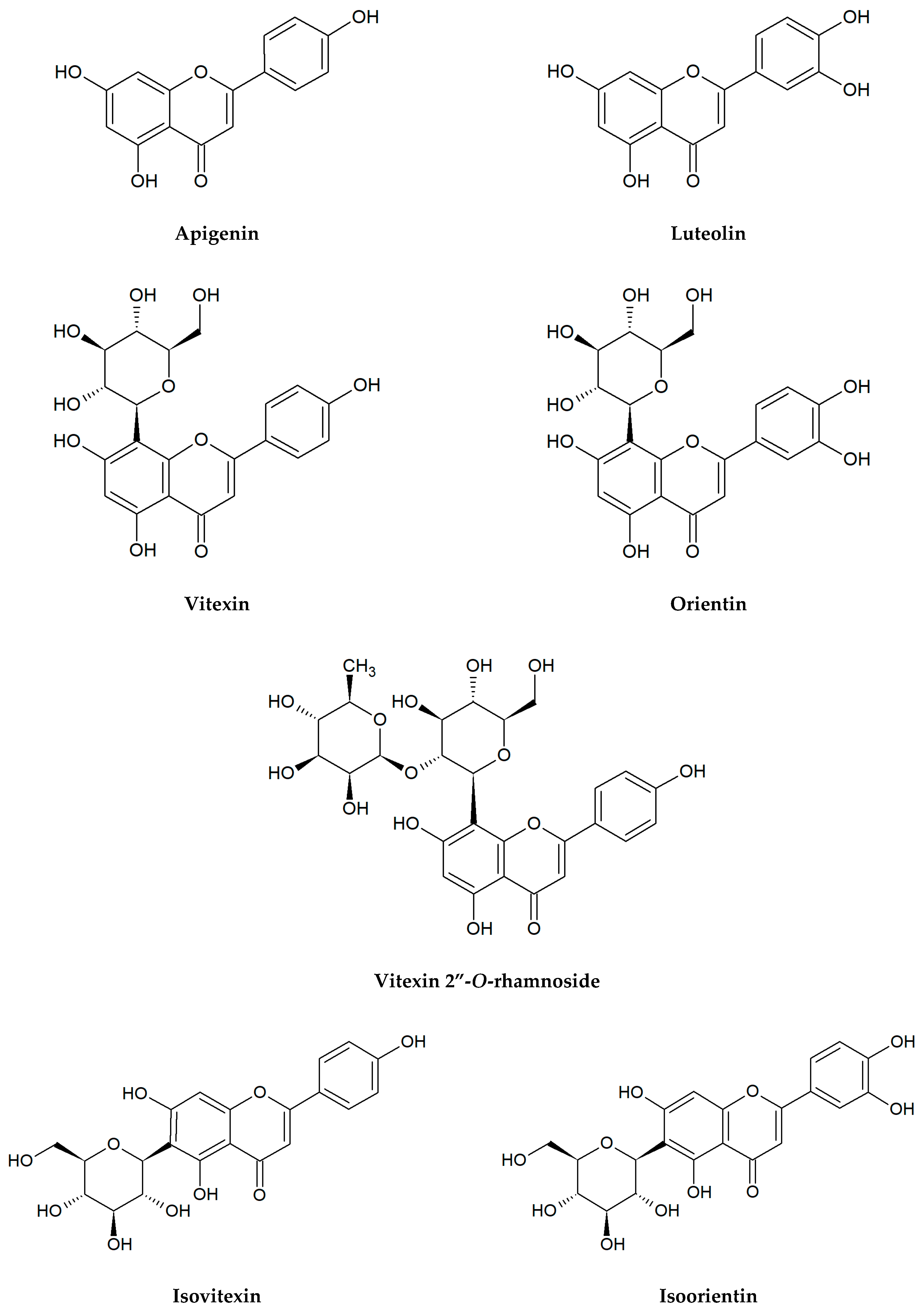
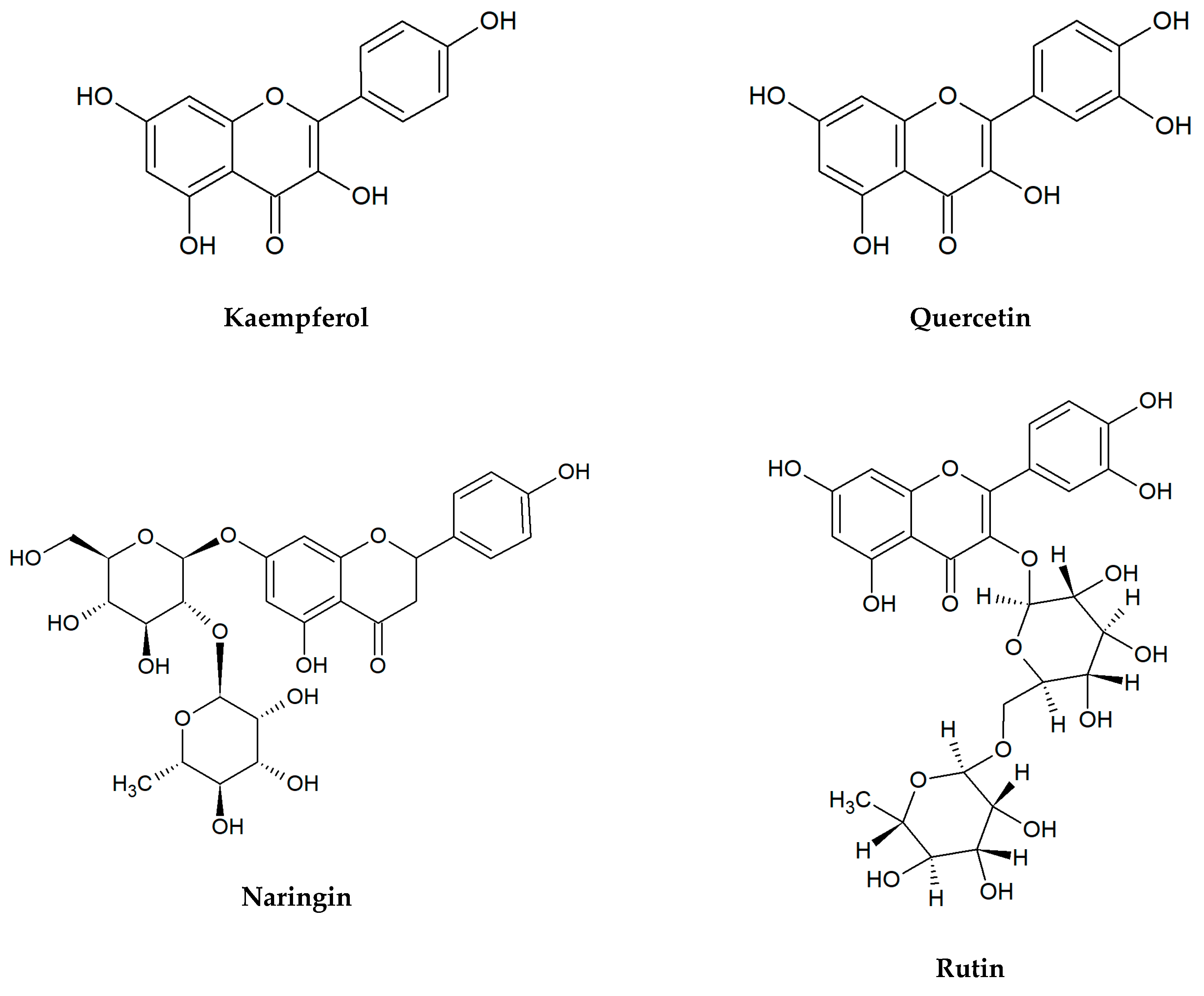
Flavonoids selected for our microbiological tests are presented in Figure 1. In the large quantities, they occur in stems and leaves, flowers as well as fruits of the species from the families of Apiaceae, Asteraceae, Betulaceae, Brassicaceae, Ericaceae, Fabaceae, Hypericaceae, Lamiaceae, Liliaceae, Passifloraceae, Polygonaceae, Primulaceae, Ranunculaceae, Rosaceae, Rubiaceae, Rutaceae, Scrophulariaceae, Tiliaceae, and Violaceae. Two flavonols: quercetin, kaempferol, and flavones: apigenin, luteolin belong to the most ubiquitous plant flavonoids [14]. A glycoside form of quercetin—rutin (sophorin, rutoside) is present in the highest concentrations in buckwheat (Fagopyrum esculentum Moench), rue (Ruta graveolens L.), flower buds of Styphnolobium japonicum (L.) Schott (Sophora japonica L.), apricots, peaches, and citrus fruits [15,16]. Apigenin derivatives, such as vitexin, isovitexin, and vitexin 2″-O-rhamnoside constitute the main bioactive compounds of leaves and flowers of hawthorn (Crataegus spp.) [17]. The 8- and 6-C-glucosides of luteolin: orientin and isoorientin are reported from different crop plants, including buckwheat, corn silk (Zea mays L.), acai fruits (Euterpe oleracea Mart., E. precatoria Mart.), and Moso bamboo leaves (Phyllostachys edulis/Carrière/J.Houz.) [18,19]. In turn, passion fruits (Passiflora spp.), skullcap roots (Scutellaria spp.) as well as honey and propolis are the main natural sources of chrysin [20,21,22,23]. Naringin is a flavanone glycoside isolated from grapes and citrus fruits, and it imparts a bitter taste to grapefruit juice [24].
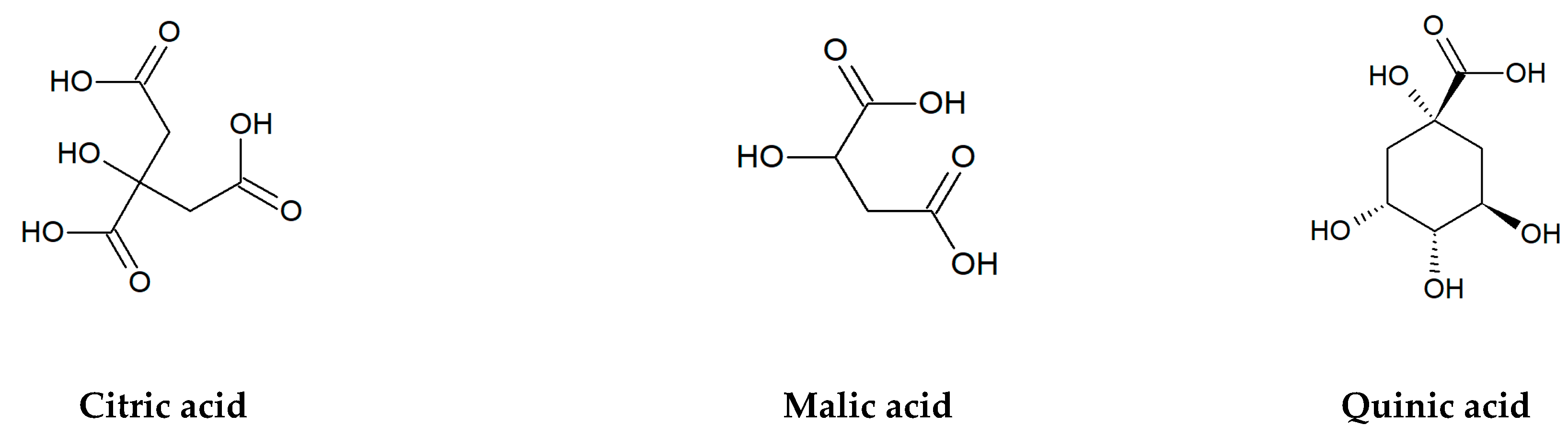
In addition to flavonoids, a lot of organic acids: both aliphatic and aromatic ones, especially phenolics are the important bioactive compounds of edible and medicinal plants (Figure 2). Among non-aromatic, short-chain hydroxy acids, malic, citric, and quinic acids belong to the most abundant substances with a key role in plant metabolism and physiology. Malic and citric acids are mainly produced in the tricarboxylic acid cycle (Krebs cycle) and, to a lesser degree, in the glyoxylate cycle, while quinic acid is a byproduct of the shikimic acid pathway [4]. High accumulation of these compounds is observed in various berry fruits of wild and cultivated plants from the Ericaceae, Rosaceae, and Grossulariaceae families, including cranberry (Vaccinium macrocarpon Aiton and V. oxycoccos L.), bilberry (V. myrtillus L.), blueberry (V. corymbosum L.), blackberry (Rubus spp.), raspberry (Rubus idaeus L.), black chokeberry (Aronia melanocarpa/Michx./Elliott), red currant (Ribes rubrum L.), black currant (Ribes nigrum L.), and many others [25,26,27,28,29,30]. For example, the total content of citric, malic, and quinic acids in fruits of European cranberry can reach almost 37% of dry matter [31].
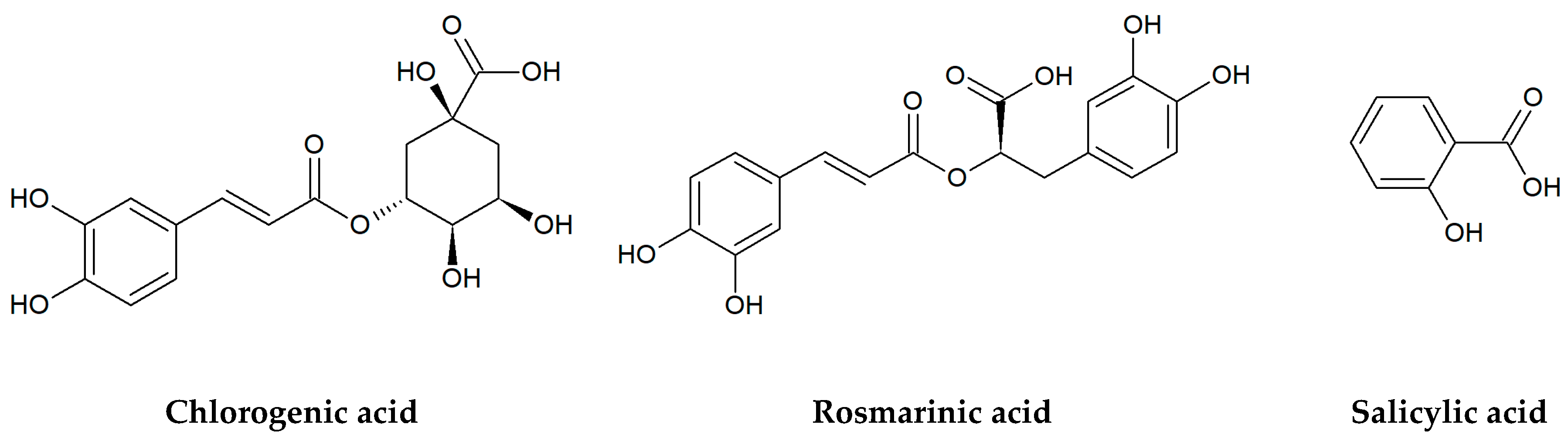
Berries are also a rich source of hydroxycinnamic acids and their derivatives, including chlorogenic (5-O-caffeoylquinic) and neochlorogenic (3-O-caffeoylquinic) acids, which are the esters formed between caffeic (3,4-dihydroxycinnamic) and quinic acids [32,33]. A great amount of chlorogenic acid isomers has been found, among others, in yerba mate (Ilex paraguariensis A.St-Hil.), coffee (Coffea spp.), and tea plant (Camellia sinensis/L./Kuntze) [34]. In turn, rosmarinic acid, an ester of caffeic and 3,4-dihydroxyphenyllactic acids, was isolated for the first time from the rosemary leaves (Rosmarinus officinalis L.). It commonly occurs in many aromatic and medicinal plants of the Lamiaceae family, especially mint (Mentha spp.) and thyme (Thymus spp.) species, lemon balm (Melissa officinalis L.), common sage (Salvia officinalis L.), oregano (Origanum vulgare L.), and sweet basil (Ocimum basilicum L.) [35,36,37]. Another well-known secondary metabolite, the phytohormone salicylic acid (SA), is a key signaling compound that participates in the plant response to pathogens, herbivores, and abiotic stress [38]. Natural salicylates such as salicylic acid and salicin (salicyl alcohol glucoside) were found in large amounts in the willow bark (Salix spp.), the buds of black poplar (Populus nigra L.), elm leaves (Ulmus spp.), and meadowsweet herb (Filipendula ulmaria/L./Maxim.) [39,40].
Our studies were focused on the estimation of antibacterial activity of selected flavonoids and organic acids widespread in fruits, vegetables, spices, and popular medicinal plants which are very often used for the prevention and treatment of various diseases. For example, many herbal preparations utilized as natural diuretics, and plant extracts with other main pharmacological activities (i.e., drugs against cardiovascular diseases, sedatives, anti-inflammatory agents) exhibit additional beneficial effects by the antimicrobial action [41,42]. Recent data show that flavonoids have protective potential against cutaneous inflammatory reactions and affect wound healing [43,44]. In addition, organic acids (especially citric acid) seem to be of significant importance in the antimicrobial activity and health of the skin [45,46]. In the present studies, we tested the biological activity of chosen flavonoids and organic acids against four widespread pathogens: Staphylococcus aureus, Enterococcus faecalis, Pseudomonas aeruginosa, and Escherichia coli. These Gram-positive and Gram-negative bacteria can cause many diseases in humans, including opportunistic infections and belong to the most common etiological factors of the skin and wound infections [47,48].
Microbiological screening tests included 19 plant metabolites from the various flavonoid classes: flavones, flavonols, flavanones, and simple organic acids: aliphatic and aromatic ones. The chosen flavonoids differed in the number of hydroxyl groups on the aromatic rings as well as the presence and position of the sugar group, which gave the opportunity to test the effect of these parameters on the biological activity of the natural compounds.
2. Materials and Methods
2.1. Chemicals
Chemicals used in this study were purchased from Merck (Sigma-Aldrich, Supelco, Poland). Plant compounds selected for the microbiological tests are presented in Table 1. All substances were dissolved in 20% water solution of dimethyl sulfoxide DMSO (Sigma-Aldrich, Poland) in a final concentration of 1 mg/mL. Additionally, DMSO was used as a negative control, while two antibiotics, ciprofloxacin (Sigma, cat. no. 17850) and gentamicin sulfate (Sigma-Aldrich, cat. no. G1914) as positives.
2.2. Bacterial Strains and Antimicrobial Activity
In the in vitro tests, there were investigated clinical isolates of two Gram-positive (Staphylococcus aureus, Enterococcus faecalis) and Gram-negative bacteria (Escherichia coli, Pseudomonas aeruginosa). For each species, four strains obtained from the collection of the Department of Medical Microbiology at Poznań University of Medical Sciences (Poland) were tested. None of them were multidrug-resistant. The species of bacteria were grown at 35 °C for 24 h, in tryptone soy agar (TSA; Graso, Poland).
The minimal inhibitory concentrations (MICs) of selected plant substances were determined by the micro-dilution method using the 96-well plates (Nest Scientific Biotechnology). Studies were conducted according to the Clinical and Laboratory Standards Institute (CLSI) [49], European Committee on Antimicrobial Susceptibility Testing (EUCAST) recommendations [50], and as described in our previous publications [48,51]. Primarily, 90 µL of Mueller–Hinton broth (Graso, Poland) was placed in each well. Serial dilutions of each of the substances were performed so that concentrations in the range of 15.6–1000 µg/mL were obtained. In the initial tests of antibacterial activity of phytochemicals, the lowest concentration amounted to 1.95 µg/mL (Figure 3), while for positive controls (antibiotics) it was 0.98 µg/mL. The inoculums were adjusted to contain approximately 108 CFU/mL bacteria. 10 µL of the proper inoculums were added to the wells, obtaining concentration 105 CFU/mL. The plates were incubated at 35 °C for 24 h, then 20 μL of 1% MTT water solution (3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide, Sigma-Aldrich) was added to the wells. Next, the plates were incubated 2–4 h at 37 °C. This assay is based on the reduction of yellow tetrazolium salt (MTT) to a soluble purple formazan product [48]. The MIC value was taken as the lowest concentration of the substance that inhibited any visible bacterial growth. The analyses were repeated three times.
In our investigations, we adopted the range of tested concentrations of phytochemicals for the MICs between 15.6 and 1000 µg/mL, although some authors determine the antimicrobial activity of natural compounds at the level of 2000–4000 µg/mL or more [52,53,54]. However, in our opinion, such high values indicate a very weak effect of these substances. During the description of the results, it was taken that the MIC = 250 μg/mL shows a relatively high antibacterial activity of plant chemicals, while the MICs = 500 and 1000 μg/mL mean moderate and low effects, respectively.
3. Results
Our research exhibited antibacterial properties of all tested flavonoids and organic acids, but their activity was quite diverse. These compounds were generally more active against Gram-negative than Gram-positive bacteria. The following tendency of microbial sensitivity to plant substances was observed: E. coli > P. aeruginosa > E. faecalis > S. aureus (Table 2). Salicylic acid showed the highest biological effect on all bacterial species (MIC = 250–500 μg/mL). However, other chemicals demonstrated a similar activity, especially against E. coli and P. aeruginosa (MIC = 500 μg/mL). Among 19 investigated phytochemicals, only three: kaempferol, quercetin, and chlorogenic acid had no significant influence on P. aeruginosa, while up to 10 compounds were relatively inactive against S. aureus (MIC > 1000 µg/mL). It was interesting that the individual strains of a given bacterial species most often did not show differences in the sensitivity to one plant substance. Only salicylic acid, rosmarinic acid, and apigenin exhibited differentiating effects on individual strains.
Although flavonol aglycones kaempferol and quercetin displayed a moderate activity only against E. coli, quercetin glycoside rutin demonstrated influence on all strains tested (MIC = 500–1000 μg/mL). A similar activity level was found for the glycosides from the other classes of flavonoids: flavanones (naringin) and flavones (vitexin, isovitexin, vitexin 2″-O-rhamnoside, orientin, isoorientin). Differences were determined only in the case of S. aureus. Naringin, vitexin and its derivatives showed no significant activity, while orientin and isoorientin were clearly stronger antibacterial agents than rutin.
Among organic acids, the highest variability in the microbiological effect was found against S. aureus and P. aeruginosa. Some metabolites such as citric, quinic, and rosmarinic acids for S. aureus, and also chlorogenic acid for P. aeruginosa were relatively inactive. The aliphatic acids: citric, malic and quinic ones showed the same level of activity within individual species of E. faecalis, E. coli, and P. aeruginosa (MIC = 500–1000 μg/mL). In turn, phenolic compounds: chlorogenic, rosmarinic, and salicylic acids exhibited variation within all bacterial species with the MIC values from 250 to above 1000 μg/mL.
4. Discussion
In recent years, a rapid increase in the number of studies concerning the antibacterial properties of plant extracts rich in phenolic compounds, including flavonoids and phenolic acids has been observed. However, due to the enormous wealth of species and natural substances, the degree of their examination is very diverse and still insufficient. Particularly, works on the antibacterial activity of individual pure compounds are relatively few. There is a small number of microbiological investigations describing the effects of some common flavonoid glycosides such as vitexin [55,56,57,58], isovitexin [59,60], vitexin 2″-O-rhamnoside [61], orientin [62,63], and isoorientin [56,60,62].
In addition, literature data are difficult to compare due to the use of various methods for assessing antibacterial activity, different solvents, and the origin and purity of test compounds, often isolated from various plant extracts [55,57,59,60,62,63,64,65]. Antimicrobial properties of natural chemicals were described not only by the minimum inhibitory concentration (MIC) [33,54,57,62,64,66,67,68,69,70,71,72] and by the minimum bactericidal concentration (MBC) [72], but also by the agar well or disc-diffusion methods [59,60,63,65]. Some authors expressed results as the IC50 or MIC80 values [33,67,73]. Moreover, the plant substances were tested in various concentrations. The kind of solvent used for the dissolution of pure compounds is the next important point in the assessment of in vitro activity. Although most authors utilized dimethyl sulfoxide, sometimes they did not give its concentration [59,62,63,65] or it is 100% DMSO [54], which may affect the level of antimicrobial activity of the tested solutions. The other solvents used were, for example, acetone [67], chloroform [59], Mueller Hinton II broth [68], and water [71]. In several cases, there was no information about dissolving procedures [57,58,60]. In this context, there is still a need for extensive screening studies that would compare the activity of a large number of plant metabolites against the same bacterial strains by a standardized method.
Our investigations exhibited moderate antibacterial properties of tested flavonoids and organic acids against clinical strains of Gram-negative pathogens: E. coli and P. aeruginosa (MIC = 500 µg/mL). Among 19 selected plant substances, only three: kaempferol, quercetin, and chlorogenic acid were inactive against P. aeruginosa at all concentrations tested (15.6–1000 µg/mL). However, for up to 10 compounds, no significant activity was found against the Gram-positive bacteria S. aureus. Additionally, another microorganism from this group E. faecalis showed low sensitivity (MIC = 1000 µg/mL) to most analyzed metabolites (Table 2). The above-described observations confirm the results of works which indicate a higher activity of natural plant substances, including flavonoids, against some Gram-negative bacteria than Gram-positive ones, although it is usually considered that this regularity is the opposite [48,55]. The general tendency of bacterial sensitivity to selected plant substances was observed as follows: E. coli > P. aeruginosa > E. faecalis > S. aureus (Table 2). Some screening studies showed the greater activity of alkaloids, flavonoids, and phenolic acids especially against P. aeruginosa, and also E. coli than S. aureus [5,55]. However, this relationship seems to have significant limitations and requires further detailed research. For example, all strains of S. aureus, E. coli, and P. aeruginosa tested by us had the same level of sensitivity to flavones chrysin, luteolin, orientin, isoorientin, and some clinical isolates of them to apigenin and salicylic acid. No differences in the inhibitory potency of bacterial growth of above-mentioned species were previously reported, among others, for luteolin, orientin, isoorientin [62], and in the case of S. aureus and E. coli for chrysin [67], luteolin [74], and glycosides of quercetin hyperoside and rutin [72].
Numerous studies allow to state that in antibacterial mechanisms of flavonoids are included mainly: inhibition of synthesis of nucleic acid, inhibition of cytoplasmic membrane function by influence the biofilm formation, porins, permeability, and by interaction with some crucial enzymes [6,8,75,76]. It was shown that apigenin inhibits the DNA gyrase of E. coli [77], and has inhibitory effects on the formation of E. coli biofilm [78]. Recently, a liposomal formulation of apigenin was examined, and it was observed increasing of its antibacterial property by the interaction of apigenin liposomes with the membrane of tested bacteria resulted in the lysis of the bacterial cells. Comparison of results exhibited much greater efficiency of liposomal apigenin against both Gram-positive and Gram-negative bacteria: B. subtilis (MIC = 4 µg/mL), S. aureus (MIC = 8 µg/mL), and E. coli (MIC = 16 µg/mL), P. aeruginosa (MIC = 64 µg/mL) [68]. Other flavones, including apigenin C-glucosides such as vitexin and isovitexin, have also been tested in order to study their effect on bacterial surface hydrophobicity and biofilm formation [57,58,59]. Das et al. [58] reported that vitexin reduces the hydrophobicity of cell surface and membrane permeability of S. aureus at the sub-MIC dose of 126 µg/mL. This flavone down-regulated the icaAB and agrAC gene expression showing antibiofilm activity and bactericidal effect. In similar work, Das et al. [57] demonstrated that vitexin exerts the MIC of 260 µg/mL against P. aeruginosa, and exhibits moderate antibiofilm activity. In turn, isovitexin (200–500 µg/mL) decreased the adhesion of methicillin-sensitive S. aureus ATCC 29213, and simultaneously increased the adhesion of two strains of E. coli [59]. Currently, it was shown that isovitexin has the potent antibacterial properties described as the diameter of the zone of growth inhibition (ZOI) for B. subtilis (19.5 mm), P. aeruginosa (17.5 mm), E. coli (14.1 mm), and Staphylococcus aureus (12.8 mm). The even stronger activity was found for isoorientin (luteolin C-glucoside), and it was as follows: B. subtilis (20.1 mm), P. aeruginosa (19.1 mm), S. aureus (18.7 mm), and E. coli (14.8 mm) [60].
Microbiological literature provides interesting data on the mechanism of action of two main flavonols: kaempferol and quercetin. It was shown that quercetin increases the cytoplasmic membrane permeability of S. pyogenes which resulted in the inhibitory influence on this Gram-positive bacterium at the MIC value of 128 µg/mL [79]. Moreover, in this study, the synergistic effect of quercetin with antibiotic ceftazidime was observed. Barbieri et al. [6] concluded that this flavonol is active not only against Gram-positive pathogens: S. aureus, S. haemolyticus, and S. pyogenes, but also against Gram-negative ones: E. coli and K. pneumoniae. Additionally, Betts et al. [80] showed a strongly inhibiting effect against methicillin-resistant S. aureus, which was significantly increased in the presence of epigallocatechin gallate. Studies of the mechanism of antimicrobial action allowed to state that quercetin diacyl glycosides show dual inhibition of DNA gyrase and topoisomerase IV [81]. In turn, our investigations exhibited the moderate effect of these plant metabolites against E. coli (MIC = 500 µg/mL), and lack of significant activity in the case of S. aureus (MIC > 1000 µg/mL). Research of Chen and Huang [82] concerning quercetin and kaempferol reported inhibition of the interaction of DNA B helicase of K. pneumoniae with deoxynucleotide triphosphates (dNTPs). Further study showed that the ATPase activity of this helicase KpDnaB was decreased to 75% and 65% in the presence of quercetin and kaempferol, respectively [83]. In the next work, Huang et al. [84] observed that kaempferol inhibits the DNA PriA helicase of S. aureus, and these results showed that the concentration of phosphate from ATP hydrolysis by this DNA helicase was decreased to 37% in the presence of 35 µM kaempferol. Thus, it was summarized that kaempferol can bind to DNA helicase and then inhibit its ATPase activity and this is a new mechanism of action for this chemical compound. According to the results, this flavonol may be taken into consideration as an active natural molecule in the development of new antibiotics against S. aureus [84]. Currently, Huang [73] demonstrated the inhibitory effect of kaempferol on the activity of a dihydropyrimidinase from P. aeruginosa with the IC50 value of 50 ± 2 μM.
Nowadays, it is believed that the structure-activity relationship in the antimicrobial effect of flavonoids should be further examined because it is a very large group of compounds, and many issues have not yet been clarified. Xie et al. [75] concluded that hydroxyl groups at special positions on the aromatic rings of flavonoids improve the antibacterial effect. Flavonoids have the C6-C3-C6 carbon structure consisting of two phenyl rings (A and B) and a heterocyclic ring (C). Generally, it was observed that at least one hydroxyl group in the ring A (especially at C-7) is vital for the antibacterial activity of flavones, and in another position such as C-5 and C-6 can increase this biological effect [85]. In this context, it is interesting to compare our results regarding the antibacterial activity of flavones with the hydroxyl groups at C-5 and C-7 (chrysin, apigenin, luteolin, and their glycosides) and flavone devoid of them. Just like other chemicals from this flavonoid class, flavone showed moderate inhibitory influence on the growth of E. coli and P. aeruginosa (MIC = 500 µg/mL). The same level of flavone activity was found against E. faecalis, and it was the highest value among the flavonoids tested. Only against S. aureus, the above-mentioned substance was inactive at concentrations tested (15.6–1000 µg/mL). Furthermore, we observed that a number of hydroxyl groups at two aromatic rings do not correspond with higher antimicrobial activity of flavonoids, i.e., quercetin has five hydroxyl groups, but it was not active against E. faecalis, S. aureus, and P. aeruginosa. In addition, some studies displayed a low effect of quercetin on B. subtilis, E. cloacae, E. coli, and K. pneumoniae [86]. The structure-activity relationships of flavonoids were discussed by Xie et al. [87], and it was summarized that two hydroxyl substituents on C-5 and C-7 of ring A of quercetin, rutin, and naringenin lead to their antibacterial activities. Moreover, it was found that the saturation of the C2=C3 double bond (in naringin) increased the antibacterial activity. However, in our study naringin was the most active against P. aeruginosa only in comparison with kaempferol and quercetin. On the other side, a recent study showed that the presence of glycosyl conjugated groups to polyphenols may reduce antibacterial activity [88]. We showed that glycosides of flavonoids (vitexin, vitexin 2″-O-rhamnoside, isovitexin, orientin, isoorientin, naringin, rutin) have some antibacterial effects (Table 2). The aglycone apigenin exhibited higher activity against S. aureus in comparison with its glycosides vitexin, isovitexin, and vitexin 2″-O-rhamnoside, however the aglycon luteolin had the same antibacterial effects on all bacterial strains as its C-glucosides orientin and isoorientin.
According to the literature, the level of sensitivity of the bacterial species studied by us to plant substances is very diverse and strongly depends not only on the type of active compound but also on the selected strains, as shown by comparative analyses in this regard [5,53]. It may also affect large discrepancies in the results between individual investigations. Some literature data suggest that standard strains are generally much more sensitive to antibiotics and natural plant compounds than current clinical isolates. For example, the MIC values of quercetin, apigenin, naringin, chlorogenic, and quinic acids for E. coli ATCC 35218, P. aeruginosa ATCC 10145, S. aureus ATCC 25923, and E. faecalis ATCC 29212 reached 2–16 µg/mL, while for the clinical strains it ranged between 32 and 128 µg/mL or above this [5]. In turn, research conducted by Su et al. [53] showed a slightly higher sensitivity of some clinical isolates of methicillin-resistant S. aureus to luteolin and quercetin (MIC = 31.2–62.5 µg/mL) than methicillin-sensitive strains (MIC = 125 µg/mL). In the study of Morimoto et al. [89], quinolone-resistant S. aureus Mu50 was much more sensitive to apigenin (MIC = 4 µg/mL) than quinolone-susceptible S. aureus strain FDA 209P (MIC > 128 µg/mL). Compared to the above-cited works [5,89], it was interesting that apigenin and chlorogenic acid were practically inactive (MIC > 4000 µg/mL) against all 34 strains of S. aureus tested by Su et al. [53]. Our investigations exhibited the moderate or weak activity of these two compounds against S. aureus (MIC = 500–1000 µg/mL). A recent review of the literature [33] showed that chlorogenic acid has a broad spectrum of antimicrobial activity, but its effect is very diverse. This phenolic acid strongly inhibited the growth of E. faecalis (MIC = 64 µg/mL), while it was inactive against P. aeruginosa (MIC80 = 10,000 µg/mL). For S. aureus and E. coli, its MIC values ranged from 40–80 to 10,000 µg/mL. The above data are largely consistent with the results of the current work (Table 2). We exhibited the moderate activity of chlorogenic acid against E. coli (MIC = 500 µg/mL) and confirmed the lack of significant influence of this substance on P. aeruginosa at the concentrations tested (MIC > 1000 µg/mL).
In addition to chlorogenic acid, we also studied the biological influence of other phenolic acids: rosmarinic and salicylic ones. In addition, their antibacterial activity was compared with some aliphatic acids: citric, malic, and quinic. Generally, there were no clear differences in the activity of these two groups of substances. However, the simple phenolic compound salicylic acid showed the highest activity with the MIC values of 250–500 µg/mL. Many studies proved that rosmarinic acid has an antimicrobial effect on Gram-positive and Gram-negative bacteria [65,71,90]. Sometimes, the level of this activity was not high. Recently, Akhtar et al. [65] indicated the moderate growth inhibition zones of clinical isolates of P. aeruginosa (13 mm in diameter), S. aureus (12 mm), Proteus vulgaris (11 mm), and E. coli (10 mm) at 1 µg/mL concentration of rosmarinic acid. Similarly, Matejczyk et al. [71] observed a not very strong antibacterial effect of this phenolic acid on E. coli (MIC > 250 µg/mL), Bacillus sp. (MIC > 500 µg/mL), S. epidermidis (MIC > 500 µg/mL), and S. pyogenes (MIC > 500 µg/mL) in comparison with an antibiotic kanamycin (MIC > 100 µg/mL). In turn, Ekambaram et al. [69] demonstrated the MIC values of rosmarinic acid against S. aureus and MRSA on the level of 800 and 10,000 µg/mL, respectively. Blaskovich et al. [54] carried out experimental research and made a critical review of the antimicrobial activity of salicylic acid. Results of these studies demonstrated that salicylic acid was practically inactive against various bacterial strains, including B. subtilis ATCC 6633, E. faecalis ATCC 29212, S. aureus, MSSA ATCC 25923, and S. pneumoniae ATCC 33400 (MIC = 32,000 µg/mL). The antibacterial properties are relatively well known for small aliphatic molecules tested by us: citric, malic, and quinic acids [91,92,93]. Investigations concerning the effect of quinic acid on cellular functions of S. aureus demonstrated that this organic acid could significantly decrease the intracellular pH and ATP concentration, and also reduce the DNA content [93]. Citric acid was previously shown to have the MICs of 900 µg/mL for S. aureus and 1500 µg/mL for E. coli, and to be very effective in the treatment of chronic wound infections in a dose of 3 g of citric acid dissolved in 100 mL of distilled water [91]. In turn, Gao et al. [85] demonstrated no clear activity of citric and malic acids against E. coli (MIC = 1667 and 2000 µg/mL, respectively) B. subtilis (MIC = 2000 µg/mL), and S. suis (MIC = 8000 and 6667 µg/mL). However, Jensen et al. [92] exhibited that cranberry juice and its main compounds (citric, malic, quinic, and shikimic acids) reduce E. coli colonization of the bladder. These organic acids decreased bacterial levels when they were administered together or in a combination of malic acid and citric or quinic ones. Our research confirmed the antibacterial activity of citric, malic, and quinic acids not only against E. coli (MIC = 500 µg/mL), but also against P. aeruginosa and E. faecalis (Table 2).
5. Conclusions
Our research confirmed the antibacterial activity of all tested plant compounds. With the exception of kaempferol and quercetin, they showed a biological effect against clinical strains of 3–4 bacterial species. Microbiological screening of flavonoids and organic acids allowed to exhibit some interesting details and relationships. First of all, these metabolites were generally more potent against Gram-negative bacteria: E. coli and P. aeruginosa than Gram-positive ones: E. faecalis and S. aureus. On the other hand, the comparative study of antibacterial activity of flavone, chrysin, apigenin, and luteolin demonstrated that the presence of hydroxyl groups in the phenyl rings A (C-5, C-7) and B (C-3′, C-4′) usually did not affect the activity level of flavones. Only in the case of S. aureus, a clear increase in the activity of the hydroxy derivatives of flavone was observed. Similarly, the presence and position of the sugar group in the flavone glycosides generally had no effect on the MIC values.
A comparison of our results with the literature data exhibited that the level of sensitivity of the bacterial species to plant substances is very diverse, and strongly depends not only on the type of active compounds but also on the strains tested. Moreover, it seems that current clinical isolates are generally much less sensitive to the natural plant metabolites than standard strains. Numerous standard strains have been isolated many years ago, therefore, with the currently growing resistance of bacteria, their use for the screening microbiological tests is limited. In our investigations, we found the moderate or even low activity of flavonoids and organic acids compared to the traditional antibiotics and some plant substances. However, examples of the use of natural compounds with a relatively low in vitro activity in the treatment of urinary tract infections, chronic wound infections, etc. or as food additives show that widely distributed flavonoids and organic acids could find broad practical applications.
Author Contributions
Conceptualization, A.A. and T.M.K.; methodology, A.A. and T.M.K.; reagents and investigation, A.A., M.O., and T.M.K.; visualization of chemical structures, T.M.K.; literature search, A.A., and M.O.; writing—original draft preparation, A.A., and M.O.; writing—review and editing, A.A., and M.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded from the budget of the Department of Medical Microbiology, Poznań University of Medical Sciences and the Polish Multiannual Programme entitled ‘Creating the scientific basis of the biological progress and conservation of plant genetic resources as a source of innovation to support sustainable agriculture and food security of the country’.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chandra, H.; Bishnoi, P.; Yadav, A.; Patni, B.; Mishra, A.P.; Nautiyal, A.R. Antimicrobial resistance and the alternative resources with special emphasis on plant-based antimicrobials–A review. Plants 2017, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.D.; Birdi, T.J. Development of botanicals to combat antibiotic resistance. J. Ayur. Integr. Med. 2017, 8, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Mate, A. (Ed.) Medicinal and Aromatic Plants of the World. Scientific, Production, Commercial and Utilization Aspects; Springer Science + Business Media: Dordrecht, The Netherlands, 2015; Volume 1. [Google Scholar]
- Zheng, J.; Huang, C.; Yang, B.; Kallio, H. Regulation of phytochemicals in fruits and berries by environmental variation—Sugars and organic acids. J. Food Biochem. 2019, 43, e12642. [Google Scholar] [CrossRef] [PubMed]
- Özçelik, B.; Kartal, M.; Orhan, I. Cytotoxicity, antiviral and antimicrobial activities of alkaloids, flavonoids, and phenolic acids. Pharm. Biol. 2011, 49, 396–402. [Google Scholar] [CrossRef]
- Barbieri, R.; Coppo, E.; Marchese, A.; Daglia, M.; Sobarzo-Sánchez, E.; Nabavi, S.F.; Nabavi, S.M. Phytochemicals for human disease: An update on plant-derived compounds antibacterial activity. Microbiol. Res. 2017, 196, 44–68. [Google Scholar] [CrossRef]
- Fialova, S.; Rendekova, K.; Mucaji, P.; Slobodnikova, L. Plant natural agents: Polyphenols, alkaloids and essential oils as perspective solution of microbial resistance. Curr. Org. Chem. 2017, 21, 1875–1884. [Google Scholar] [CrossRef]
- Khameneh, B.; Iranshahy, M.; Soheili, V.; Bazzaz, B.S.F. Review on plant antimicrobials: A mechanistic viewpoint. Antimicrob. Resist. Infect. Control 2019, 8, 118. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Grijalva, E.P.; Picos-Salas, M.A.; Leyva-López, N.; Criollo-Mendoza, M.S.; Vazquez-Olivo, G.; Heredia, J.B. Flavonoids and phenolic acids from oregano: Occurrence, biological activity and health benefits. Plants 2018, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Jungbauer, A.; Medjakovic, S. Anti-inflammatory properties of culinary herbs and spices that ameliorate the effects of metabolic syndrome. Maturitas 2012, 71, 227–239. [Google Scholar] [CrossRef]
- Goncalves, S.; Moreira, E.; Grosso, C.; Andrade, P.B.; Valentao, P.; Romano, A. Phenolic profile, antioxidant activity and enzyme inhibitory activities of extracts from aromatic plants used in mediterranean diet. J. Food Sci. Technol. 2017, 54, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Mileo, A.M.; Nisticò, P.; Miccadei, S. Polyphenols: Immunomodulatory and therapeutic implication in colorectal cancer. Front. Immunol. 2019, 10, 729. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Cabral, C.; Kumar, R.; Ganguly, R.; Rana, H.K.; Gupta, A.; Lauro, M.R.; Carbone, C.; Reis, F.; Pandey, A.K. Beneficial effects of dietary polyphenols on gut microbiota and strategies to improve delivery efficiency. Nutrients 2019, 11, 2216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Firrman, J.; Liu, L.S.; Yam, K. A review on flavonoid apigenin: Dietary intake, ADME, antimicrobial effects, and interactions with human gut microbiota. BioMed Res. Int. 2019, 2019, 7010467. [Google Scholar] [CrossRef]
- Chua, L.S. A review on plant-based rutin extraction methods and its pharmacological activities. J. Ethnopharmacol. 2013, 150, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Enogieru, A.B.; Haylett, W.; Hiss, D.C.; Bardien, S.; Ekpo, O.E. Rutin as a potent antioxidant: Implications for neurodegenerative disorders. Oxid. Med. Cell Longev. 2018, 2018, 6241017. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.E.; Brown, P.N.; Talent, N.; Dickinson, T.A.; Shipley, P.R. A review of the chemistry of the genus Crataegus. Phytochemistry 2012, 79, 5–26. [Google Scholar] [CrossRef]
- Yamaguchi, K.K.L.; Pereira, L.F.R.; Lamarão, C.V.; Lima, E.S.; da Veiga-Junior, V.F. Amazon acai: Chemistry and biological activities: A review. Food Chem. 2015, 179, 137–151. [Google Scholar] [CrossRef]
- Yuan, L.; Wang, J.; Wu, W.; Liu, Q.; Liu, X. Effect of isoorientin on intracellular antioxidant defence mechanisms in hepatoma and liver cell lines. Biomed. Pharmacother. 2016, 81, 356–362. [Google Scholar] [CrossRef]
- Mani, R.; Natesan, V. Chrysin: Sources, beneficial pharmacological activities, and molecular mechanism of action. Phytochemistry 2018, 145, 187–196. [Google Scholar] [CrossRef]
- Ożarowski, M.; Piasecka, A.; Paszel-Jaworska, A.; Chaves, D.S.; Romaniuk, A.; Rybczyńska, M.; Gryszczynska, A.; Sawikowska, A.; Kachlicki, P.; Mikolajczak, P.L.; et al. Comparison of bioactive compounds content in leaf extracts of Passiflora incarnata, P. caerulea and P. alata and in vitro cytotoxic potential on leukemia cell lines. Rev. Bras. Farmacogn. 2018, 28, 79–191. [Google Scholar] [CrossRef]
- Naz, S.; Imran, M.; Rauf, A.; Orhan, I.E.; Shariati, M.A.; Haq, I.U.; Yasmin, I.; Shahbaz, M.; Qaisrani, T.B.; Shah, Z.A.; et al. Chrysin: Pharmacological and therapeutic properties. Life Sci. 2019, 235, 116797. [Google Scholar] [CrossRef]
- Przybyłek, I.; Karpiński, T.M. Antibacterial properties of propolis. Molecules 2019, 24, 2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, M.A.; Subhan, N.; Rahman, M.M.; Uddin, S.J.; Reza, H.M.; Sarker, S.D. Effect of Citrus flavonoids, naringin and naringenin, on metabolic syndrome and their mechanisms of action. Adv. Nutr. 2014, 5, 404–417. [Google Scholar] [CrossRef] [PubMed]
- Viljakainen, S.; Visti, A.; Laakso, S. Concentrations of organic acids and soluble sugars in juices from Nordic berries. Acta Agric. Scand. Sect. B Soil Plant Sci. 2002, 52, 101–109. [Google Scholar] [CrossRef]
- Pande, G.; Akoh, C.C. Organic acids, antioxidant capacity, phenolic content and lipid characterisation of Georgia-grown underutilized fruit crops. Food Chem. 2010, 120, 1067–1075. [Google Scholar] [CrossRef]
- Nour, V.; Trandafir, I.; Ionica, M.E. Ascorbic acid, anthocyanins, organic acids and mineral content of some black and red currant cultivars. Fruits 2011, 66, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Kaume, L.; Howard, L.R.; Devareddy, L. The blackberry fruit: A review on its composition and chemistry, metabolism and bioavailability, and health benefits. J. Agric. Food Chem. 2012, 60, 5716–5727. [Google Scholar] [CrossRef]
- Wang, Y.; Johnson-Cicalese, J.; Singh, A.P.; Vorsa, N. Characterization and quantification of flavonoids and organic acids over fruit development in American cranberry (Vaccinium macrocarpon) cultivars using HPLC and APCI-MS/MS. Plant Sci. 2017, 262, 91–102. [Google Scholar] [CrossRef]
- Denev, P.; Kratchanova, M.; Petrova, I.; Klisurova, D.; Georgiev, Y.; Ognyanov, M.; Yanakieva, I. Black chokeberry (Aronia melanocarpa (Michx.) Elliot) fruits and functional drinks differ significantly in their chemical composition and antioxidant activity. J. Chem. 2018, 2018, 9574587. [Google Scholar] [CrossRef] [Green Version]
- Adamczak, A.; Buchwald, W.; Kozłowski, J. Variation in the content of flavonols and main organic acids in the fruit of European cranberry (Oxycoccus palustris Pers.) growing in peatlands of North-Western Poland. Herba Pol. 2011, 57, 5–15. [Google Scholar]
- Jurikova, T.; Mlcek, J.; Skrovankova, S.; Sumczynski, D.; Sochor, J.; Hlavacova, I.; Snopek, L.; Orsavova, J. Fruits of black chokeberry Aronia melanocarpa in the prevention of chronic diseases. Molecules 2017, 22, 944. [Google Scholar] [CrossRef] [PubMed]
- Santana-Gálvez, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Chlorogenic Acid: Recent advances on its dual role as a food additive and a nutraceutical against metabolic syndrome. Molecules 2017, 22, 358. [Google Scholar] [CrossRef] [Green Version]
- Meinhart, A.D.; Damin, F.M.; Caldeirao, L.; Silveira, T.F.F.; Filho, J.T.; Godoy, H.T. Chlorogenic acid isomer contents in 100 plants commercialized in Brazil. Food Res. Int. 2017, 99, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Shekarchi, M.; Hajimehdipoor, H.; Saeidnia, S.; Gohari, A.R.; Hamedani, M.P. Comparative study of rosmarinic acid content in some plants of Labiatae family. Pharmacogn. Mag. 2012, 8, 37–41. [Google Scholar] [PubMed] [Green Version]
- Ożarowski, M.; Mikołajczak, P.; Bogacz, A.; Gryszczyńska, A.; Kujawska, M.; Jodynis-Libert, J.; Piasecka, A.; Napieczynska, H.; Szulc, M.; Kujawski, R.; et al. Rosmarinus officinalis L. leaf extract improves memory impairment and affects acetylcholinesterase and butyrylcholinesterase activities in rat brain. Fitoterapia 2013, 91, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Ożarowski, M.; Mikolajczak, P.L.; Piasecka, A.; Kachlicki, P.; Kujawski, R.; Bogacz, A.; Bartkowiak-Wieczorek, J.; Szulc, M.; Kaminska, E.; Kujawska, M.; et al. Influence of the Melissa officinalis leaf extract on long-term memory in scopolamine animal model with assessment of mechanism of action. Evid. Based Complem. Alternat. Med. 2016, 2016, 9729818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balcke, G.U.; Handrick, V.; Bergau, N.; Fichtner, M.; Henning, A.; Stellmach, H.; Tissier, A.; Hause, B.; Frolov, A. An UPLC-MS/MS method for highly sensitive high-throughput analysis of phytohormones in plant tissues. Plant Methods 2012, 8, 47. [Google Scholar] [CrossRef] [Green Version]
- Toiu, A.; Vlase, L.; Oniga, I.; Benedec, D.; Tămaş, M. HPLC analysis of salicylic derivatives from natural products. Farmacia 2011, 59, 106–112. [Google Scholar]
- Bijttebier, S.; van der Auwera, A.; Voorspoels, S.; Noten, B.; Hermans, N.; Pieters, L.; Apers, S. A first step in the quest for the active constituents in Filipendula ulmaria (meadowsweet): Comprehensive phytochemical identification by liquid chromatography coupled to quadrupole-orbitrap mass spectrometry. Planta Med. 2016, 82, 559–572. [Google Scholar] [CrossRef] [Green Version]
- Nabavi, S.F.; Habtemariam, S.; Ahmed, T.; Sureda, A.; Daglia, M.; Sobarzo-Sánchez, E.; Nabavi, S.M. Polyphenolic composition of Crataegus monogyna Jacq.: From chemistry to medical applications. Nutrients 2015, 7, 7708–7728. [Google Scholar] [CrossRef]
- Kerasioti, E.; Apostolou, A.; Kafantaris, I.; Chronis, K.; Kokka, E.; Dimitriadou, C.; Tzanetou, E.N.; Priftis, A.; Koulocheri, S.D.; Haroutounian, S.; et al. Polyphenolic composition of Rosa canina, Rosa sempervivens and Pyrocantha coccinea extracts and assessment of their antioxidant activity in human endothelial cells. Antioxidants 2019, 8, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.W.; Cho, S.Y.; Lee, S.R.; Lee, K.S. Onion extract and quercetin induce matrix metalloproteinase-1 in vitro and in vivo. Int. J. Mol. Med. 2010, 25, 347–352. [Google Scholar] [PubMed]
- Chuang, S.Y.; Lin, Y.K.; Lin, C.F.; Wang, P.W.; Chen, E.L.; Fang, J.Y. Elucidating the skin delivery of aglycone and glycoside flavonoids: How the structures affect cutaneous absorption. Nutrients 2017, 9, 1304. [Google Scholar] [CrossRef] [Green Version]
- Nagoba, B.S.; Suryawanshi, N.M.; Wadher, B.; Selkar, S. Acidic environment and wound healing: A review. Wounds 2015, 27, 5–11. [Google Scholar]
- Nagoba, B.; Davane, M.; Gandhi, R.; Wadher, B.; Suryawanshi, N.; Selkar, S. Treatment of skin and soft tissue infections caused by Pseudomonas aeruginosa—A review of our experiences with citric acid over the past 20 years. Wound Med. 2017, 19, 5–9. [Google Scholar] [CrossRef]
- Bessa, L.J.; Fazii, P.; Di Giulio, M.; Cellini, L. Bacterial isolates from infected wounds and their antibiotic susceptibility pattern: Some remarks about wound infection. Int. Wound J. 2013, 12, 47–52. [Google Scholar] [CrossRef]
- Karpiński, T.M. Efficacy of octenidine against Pseudomonas aeruginosa strains. Eur. J. Biol. Res. 2019, 9, 135–140. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Disk Susceptibility Tests. Approved Standard, 12th ed.; CLSI document M02-A12; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015; Volume 35, no 1. [Google Scholar]
- EUCAST. MIC Determination of Non-Fastidious and Fastidious Organisms. Available online: http://www.eucast.org/ast_of_bacteria/mic_determination (accessed on 26 July 2019).
- Karpiński, T.M.; Adamczak, A. Fucoxanthin—An antibacterial carotenoid. Antioxidants 2019, 8, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.; Shao, J.; Sun, H.; Zhong, W.; Zhuang, W.; Zhang, Z. Evaluation of different kinds of organic acids and their antibacterial activity in Japanese Apricot fruits. Afr. J. Agric. Res. 2012, 7, 4911–4918. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Ma, L.; Wen, Y.; Wang, H.; Zhang, S. Studies of the in vitro antibacterial activities of several polyphenols against clinical isolates of methicillin-resistant Staphylococcus aureus. Molecules 2014, 19, 12630–12639. [Google Scholar] [CrossRef] [Green Version]
- Blaskovich, M.A.; Elliott, A.G.; Kavanagh, A.M.; Ramu, S.; Cooper, M.A. In vitro antimicrobial activity of acne drugs against skin-associated bacteria. Sci. Rep. 2019, 9, 14658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basile, A.; Giordano, S.; López-Sáez, J.A.; Cobianchi, R.C. Antibacterial activity of pure flavonoids isolated from mosses. Phytochemistry 1999, 52, 1479–1482. [Google Scholar] [CrossRef]
- Afifi, F.U.; Abu-Dahab, R. Phytochemical screening and biological activities of Eminium spiculatum (Blume) Kuntze (family Araceae). Nat. Prod. Res. 2012, 26, 878–882. [Google Scholar] [CrossRef] [PubMed]
- Das, M.C.; Sandhu, P.; Gupta, P.; Rudrapaul, P.; De, U.C.; Tribedi, P.; Akhter, Y.; Bhattacharjee, S. Attenuation of Pseudomonas aeruginosa biofilm formation by vitexin: A combinatorial study with azithromycin and gentamicin. Sci. Rep. 2016, 6, 23347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, M.C.; Das, A.; Samaddar, S.; Dawarea, A.V.; Ghosh, C.; Acharjee, S.; Sandhu, P.; Jawed, J.J.; De Utpal, C.; Majumdar, S.; et al. Vitexin alters Staphylococcus aureus surface hydrophobicity to interfere with biofilm 2 formation. bioRxiv 2018. [Google Scholar] [CrossRef]
- Awolola, G.V.; Koorbanally, N.A.; Chenia, H.; Shode, F.O.; Baijnath, H. Antibacterial and anti-biofilm activity of flavonoids and triterpenes isolated from the extracts of Ficus sansibarica Warb. subsp. Sansibarica (Moraceae) extracts. Afr. J. Tradit. Complem. Altern. Med. 2014, 11, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Rammohan, A.; Bhaskar, B.V.; Venkateswarlu, N.; Rao, V.L.; Gunasekar, D.; Zyryanov, G.V. Isolation of flavonoids from the flowers of Rhynchosia beddomei Baker as prominent antimicrobial agents and molecular docking. Microb. Pathog. 2019, 136, 103667. [Google Scholar] [CrossRef]
- Aderogba, M.A.; Akinkunmi, E.O.; Mabusela, W.T. Antioxidant and antimicrobial activities of flavonoid glycosides from Dennettia tripetala G. Baker leaf extract. Nig. J. Nat. Prod. Med. 2011, 15, 49–52. [Google Scholar] [CrossRef]
- Cottiglia, F.; Loy, G.; Garau, D.; Floris, C.; Casu, M.; Pompei, R.; Bonsignore, L. Antimicrobial evaluation of coumarins and flavonoids from the stems of Daphne gnidium L. Phytomedicine 2001, 8, 302–305. [Google Scholar] [CrossRef]
- Ali, H.; Dixit, S. In vitro antimicrobial activity of flavanoids of Ocimum sanctum with synergistic effect of their combined form. Asian Pac. J. Trop. Dis. 2012, 2, S396–S398. [Google Scholar] [CrossRef]
- Celiz, G.; Daz, M.; Audisio, M.C. Antibacterial activity of naringin derivatives against pathogenic strains. J. Appl. Microbiol. 2011, 111, 731–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhtar, M.S.; Hossain, M.A.; Said, S.A. Isolation and characterization of antimicrobial compound from the stem-bark of the traditionally used medicinal plant Adenium obesum. J. Tradit. Complem. Med. 2017, 7, 296–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Govindarajan, R.; Rawat, A.K.S.; Khare, P.B. Antimicrobial flavonoid rutin from Pteris vittata L. against pathogenic gastrointestinal microflora. Am. Fern J. 2008, 98, 98–103. [Google Scholar] [CrossRef]
- Liu, H.; Mou, Y.; Zhao, J.; Wang, J.; Zhou, L.; Wang, M.; Wang, D.; Han, J.; Yu, Z.; Yang, F. Flavonoids from Halostachys caspica and their antimicrobial and antioxidant activities. Molecules 2010, 15, 7933–7945. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, K.; Banerjee, S.; Das, S.; Mandal, M. Probing the potential of apigenin liposomes in enhancing bacterial membrane perturbation and integrity loss. J. Colloid Interface Sci. 2015, 453, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Ekambaram, S.P.; Perumal, S.S.; Balakrishnan, A.; Marappan, N.; Gajendran, S.S.; Viswanathan, V. Antibacterial synergy between rosmarinic acid and antibiotics against methicillin-resistant Staphylococcus aureus. J. Intercult. Ethnopharmacol. 2016, 5, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Smiljkovic, M.; Stanisavljevic, D.; Stojkovic, D.; Petrovic, I.; Vicentic, M.J.; Popovic, J.; Golic Grdadolnik, S.; Markovic, D.; Sankovic-Babice, S.; Glamoclija, J.; et al. Apigenin-7-O-glucoside versus apigenin: Insight into the modes of anticandidal and cytotoxic actions. EXCLI J. 2017, 16, 795–807. [Google Scholar]
- Matejczyk, M.; Swisłocka, R.; Golonko, A.; Lewandowski, W.; Hawrylik, E. Cytotoxic, genotoxic and antimicrobial activity of caffeic and rosmarinic acids and their lithium, sodium and potassium salts as potential anticancer compounds. Adv. Med. Sci. 2018, 63, 14–21. [Google Scholar] [CrossRef]
- Ren, G.; Xue, P.; Sun, X.; Zhao, G. Determination of the volatile and polyphenol constituents and the antimicrobial, antioxidant, and tyrosinase inhibitory activities of the bioactive compounds from the by-product of Rosa rugosa Thunb. var. plena Regal tea. BMC Complem. Altern. Med. 2018, 18, 307. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.Y. Inhibition of a putative dihydropyrimidinase from Pseudomonas aeruginosa PAO1 by flavonoids and substrates of cyclic amidohydrolases. PLoS ONE 2015, 10, e0127634. [Google Scholar] [CrossRef] [Green Version]
- Bustos, P.S.; Deza-Ponzio, R.; Páez, P.L.; Cabrera, J.L.; Virgolini, M.B.; Ortega, M.G. Flavonoids as protective agents against oxidative stress induced by gentamicin in systemic circulation. Potent protective activity and microbial synergism of luteolin. Food Chem. Toxicol. 2018, 118, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Yang, W.; Tang, F.; Chen, X.; Ren, L. Antibacterial activities of flavonoids: Structure-activity relationship and mechanism. Curr. Med. Chem. 2015, 22, 132–149. [Google Scholar] [CrossRef] [PubMed]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef] [Green Version]
- Ohemeng, K.A.; Schwender, C.F.; Fu, K.P.; Barrett, J.F. DNA gyrase inhibitory and antibacterial activity of some flavones. Bioorg. Med. Chem. Lett. 1993, 3, 225–230. [Google Scholar] [CrossRef]
- Lee, J.H.; Regmi, S.C.; Kim, J.A.; Cho, M.H.; Yun, H.; Lee, C.S.; Lee, J. Apple flavonoid phloretin inhibits Escherichia coli O157:H7 biofilm formation and ameliorates colon inflammation in rats. Infect. Immun. 2011, 79, 4819–4827. [Google Scholar] [CrossRef] [Green Version]
- Siriwong, S.; Thumanu, K.; Hengpratom, T.; Eumkeb, G. Synergy and mode ofaction of ceftazidime plus quercetin or luteolin on Streptococcus pyogenes. Evid. Based Complem. Altern. Med. 2015, 2015, 759459. [Google Scholar] [CrossRef] [Green Version]
- Betts, J.W.; Sharili, A.S.; Phee, L.M.; Wareham, D.W. In vitro activity of epigallocatechin gallate and quercetin alone and in combination versus clinical isolates of methicillin-resistant Staphylococcus aureus. J. Nat. Prod. 2015, 78, 2145–2148. [Google Scholar] [CrossRef]
- Hossion, A.M.; Zamami, Y.; Kandahary, R.K.; Tsuchiya, T.; Ogawa, W.; Iwado, A. Quercetin diacylglycoside analogues showing dual inhibition of DNA gyrase and topoisomerase IV as novel antibacterial agents. J. Med. Chem. 2011, 54, 3686–3703. [Google Scholar] [CrossRef]
- Chen, C.C.; Huang, C.Y. Inhibition of Klebsiella pneumoniae DnaB helicase by the flavonol galangin. Protein J. 2011, 30, 59–65. [Google Scholar] [CrossRef]
- Lin, H.H.; Huang, C.Y. Characterization of flavonol inhibition of DnaB helicase: Real-time monitoring, structural modeling, and proposed mechanism. J. Biomed. Biotechnol. 2012, 2012, 735368. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.H.; Huang, C.C.; Chen, C.C.; Yang, K.; Huang, C.Y. Inhibition of Staphylococcus aureus PriA helicase by flavonol kaempferol. Protein J. 2015, 34, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, F.; Khameneh, B.; Iranshahi, M.; Iranshahy, M. Antibacterial activity of flavonoids and their structure-activity relationship: An update review. Phytother. Res. 2019, 33, 13–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echeverría, J.; Opazo, J.; Mendoza, L.; Urzúa, A.; Wilkens, M. Structure-activity and lipophilicity relationships of selected antibacterial natural flavones and flavanones of chilean flora. Molecules 2017, 22, 608. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Chen, J.; Xiao, A.; Liu, L. Antibacterial activity of polyphenols: Structure-activity relationship and influence of hyperglycemic condition. Molecules 2017, 22, 1913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouarab-Chibane, L.; Forquet, V.; Lantéri, P.; Clément, Y.; Léonard-Akkari, L.; Oulahal, N.; Degraeve, P.; Bordes, C. Antibacterial properties of polyphenols: Characterization and QSAR (Quantitative Structure–Activity Relationship) models. Front. Microbiol. 2019, 10, 829. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, Y.; Baba, T.; Sasaki, T.; Hiramatsu, K. Apigenin as an anti-quinolone-resistance antibiotic. Int. J. Antimicrob. Agents. 2015, 46, 666–673. [Google Scholar] [CrossRef] [Green Version]
- Amin, A.; Vincent, R.; Séverine, M. Rosmarinic acid and its methyl ester as antimicrobial components of the hydromethanolic extract of Hyptis atrorubens Poit. (Lamiaceae). Evid. Based Complem. Alternat. Med. 2013, 2013, 604536. [Google Scholar]
- Nagoba, B.S.; Gandhi, R.C.; Wadher, B.J.; Potekar, R.M.; Kolhe, S.M. Microbiological, histopathological and clinical changes in chronic infected wounds after citric acid treatment. J. Med. Microbiol. 2008, 57, 681–682. [Google Scholar] [CrossRef] [Green Version]
- Jensen, H.D.; Struve, C.; Christensen, S.B.; Krogfelt, K.A. Cranberry juice and combinations of its organic acids are effective against experimental urinary tract infection. Front. Microbiol. 2017, 8, 542. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Wu, Y.; Zhong, K.; Xiao, K.; Liu, L.; Huang, Y.; Wang, Z.; Gao, H. A comparative study on the effects of quinic acid and shikimic acid on cellular functions of Staphylococcus aureus. J. Food Prot. 2018, 81, 1187–1192. [Google Scholar] [CrossRef]
Figure 1.
Chemical structures of flavonoids tested in the present research.



Figure 2.
Chemical structures of organic acids tested in the present research.


Figure 3.
The minimal inhibitory concentrations (MICs) of selected plant substances against Pseudomonas aeruginosa strain according to the micro-dilution method.
Figure 3.
The minimal inhibitory concentrations (MICs) of selected plant substances against Pseudomonas aeruginosa strain according to the micro-dilution method.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Plant pure substances used in the microbiological assays.
| No | Merck (Sigma-Aldrich, Supelco) | CAS No | PubChem CID | Purity |
|---|---|---|---|---|
| 1 | Apigenin | 520-36-5 | 5280443 | ≥95.0% (HPLC) |
| 2 | Chrysin | 480-40-0 | 5281607 | ≥98.0% (HPLC) |
| 3 | Flavone | 525-82-6 | 10680 | ≥99.0% |
| 4 | Isoorientin | 4261-42-1 | 114776 | ≥98.0% (HPLC) |
| 5 | Isovitexin | 38953-85-4 | 162350 | ≥98.0% (HPLC) |
| 6 | Kaempferol | 520-18-3 | 5280863 | ≥97.0% (HPLC) |
| 7 | Luteolin | 491-70-3 | 5280445 | ≥97.0% (HPLC) |
| 8 | Naringin | 10236-47-2 | 442428 | ≥95.0% (HPLC) |
| 9 | Orientin | 28608-75-5 | 5281675 | ≥98.0% (HPLC) |
| 10 | Quercetin | 117-39-5 | 5280343 | ≥95.0% (HPLC) |
| 11 | Rutin | 153-18-4 | 5280805 | ≥95.0% (HPLC) |
| 12 | Vitexin | 3681-93-4 | 5280441 | ≥95.0% (HPLC) |
| 13 | Vitexin 2″-O-rhamnoside | 64820-99-1 | 5282151 | ≥98.0% (HPLC) |
| 14 | Chlorogenic acid | 327-97-9 | 1794427 | ≥95.0% (HPLC) |
| 15 | Citric acid | 77-92-9 | 311 | ≤100% |
| 16 | Malic acid | 6915-15-7 | 525 | ≤100% |
| 17 | Quinic acid | 77-95-2 | 6508 | analytical standard |
| 18 | Rosmarinic acid | 20283-92-5 | 5281792 | ≥98.0% (HPLC) |
| 19 | Salicylic acid | 69-72-7 | 338 | ≥99.0% |
Table 2.
Antibacterial activity of selected plant substances against Gram (+) and Gram (−) bacteria.
Table 2.
Antibacterial activity of selected plant substances against Gram (+) and Gram (−) bacteria.
| Plant Substance | Tested Bacteria | |||
|---|---|---|---|---|
| Staphylococcus aureus | Enterococcus faecalis | Escherichia coli | Pseudomonas aeruginosa | |
| MIC (µg/mL) | ||||
| Kaempferol | >1000 | >1000 | 500 | >1000 |
| Quercetin | >1000 | >1000 | 500 | >1000 |
| Rutin | 1000 | 1000 | 500 | 500 |
| Naringin | >1000 | 1000 | 500 | 500 |
| Flavone | >1000 | 500 | 500 | 500 |
| Chrysin | 500 | 1000 | 500 | 500 |
| Apigenin | 500, 1000 (3x) | 1000 | 500 | 500 |
| Vitexin | >1000 | 1000 | 500 | 500 |
| Isovitexin | >1000 | 1000 | 500 | 500 |
| Vitexin 2″-O-rhamnoside | >1000 | 1000 | 500 | 500 |
| Luteolin | 500 | 1000 | 500 | 500 |
| Orientin | 500 | 1000 | 500 | 500 |
| Isoorientin | 500 | 1000 | 500 | 500 |
| Citric acid | >1000 | 1000 | 500 | 500 |
| Malic acid | 1000 | 1000 | 500 | 500 |
| Quinic acid | >1000 | 1000 | 500 | 500 |
| Chlorogenic acid | 1000 | 1000 | 500 | >1000 |
| Rosmarinic acid | >1000 | 1000 | 500 | 500 (2x), 1000 (2x) |
| Salicylic acid | 250 (2x), 500 (2x) | 500 | 250 (3x), 500 | 500 |
| Median | >1000 | 1000 | 500 | 500 |
| 20% DMSO (negative control) | >1000 | >1000 | >1000 | >1000 |
| Ciprofloxacin (positive) | <1 | <1 | <1 | <1 |
| Gentamicin sulfate (positive) | <1 | <1–62.5 | <1–3.9 | <1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Adamczak, A.; Ożarowski, M.; Karpiński, T.M. Antibacterial Activity of Some Flavonoids and Organic Acids Widely Distributed in Plants. J. Clin. Med. 2020, 9, 109. https://doi.org/10.3390/jcm9010109
AMA Style
Adamczak A, Ożarowski M, Karpiński TM. Antibacterial Activity of Some Flavonoids and Organic Acids Widely Distributed in Plants. Journal of Clinical Medicine. 2020; 9(1):109. https://doi.org/10.3390/jcm9010109
Chicago/Turabian StyleAdamczak, Artur, Marcin Ożarowski, and Tomasz M. Karpiński. 2020. "Antibacterial Activity of Some Flavonoids and Organic Acids Widely Distributed in Plants" Journal of Clinical Medicine 9, no. 1: 109. https://doi.org/10.3390/jcm9010109
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.